Содержание
Урок биологии в 10-м классе по теме «Сходство и различие в строении клеток растений, животных, бактерий и грибов»
Категория: Биология.
Оборудование: микропрепараты: растительная
клетка, вольвокс, инфузория туфелька, чешуя лука,
эвглена зеленая, животная клетка. Микроскопы,
цифровой микроскоп, компьютер, мультимедиа.
Таблицы: “Растительная клетка”, “Животная
клетка”, “Бактериальная клетка”, “Грибы”.
Ход урока
I. Организационный момент.
(приветствие учителя)
II. Актуализация знаний.
Слово учителя:
— На прошлом уроке вы познакомились с
особенностями строения прокариотических и
эукариотических клеток, нашли сходство и
различия в их строении.
Давайте обсудим следующие вопросы.
1. Работа по карточкам.
(Карточки раздаются 4 учащимся)
Карточка 1.
В чем особенность ДНК прокариот?
Карточка 2.
Чем в клетке бактерий заменены отсутствующие
органоиды?
Карточка 3.
В чем биологическое значение спорообразования?
Карточка 4.
Что общего в строении клеток эукариот и
прокариот?
2. Фронтальная работа.
Биологический диктант.
(Класс делится на 3 варианта. Учитель читает
текст, на экране этот текст дублируется)
(Слайд 1)
1 вариант. Выпишите организмы, ведущие
паразитический способ питания.
2 вариант. Выпишите организмы, ведущие
сапрофитный способ питания.
3 вариант. Выпишите организмы –
хемосинтетики.
Представители:
(Слайд 2)
1 – возбудители тифа,
2 – молочнокислые бактерии,
3 – гнилостные бактерии,
4 – серобактерии,
5 – железобактерии,
6 – возбудители ангины,
7 – холерный вибрион,
8 – азотобактерии,
9 – уксуснокислые бактерии,
10 – возбудители чумы.
Поверка выполнения задания.
(Устные ответы обучающихся)
(Слайд 3)
Задание 2:
На экране вы видите микропрепараты. Ваша задача
Ваша задача
— определить их название, к какому Царству они
относятся.
На экране проецируются через цифровой
микроскоп микропрепараты:
(1 – растительной клетки, 2 – вольвокса, 3 –
инфузории туфельки, 4 – чешуи лука, 5 – эвглены
зеленой, 6 – животной клетки, 7 – сенной палочки, 8
– дрожжей)
Поверка выполнения задания.
(Устные ответы обучающихся)
(Слайд 4, 5, 6)
Слово учителя
— Мы вспомнили особенности строения клеток
прокариот и эукариот, нашли сходство и различия в
их строении. Выяснили, что все живое состоит из
клеток.
III. Изучение нового материала.
Слово учителя
— Перечислите основные положения современной
клеточной теории.
(Устные ответы обучающихся)
Слово учителя
— На уроке мы остановимся на одном из них. Как
вы думаете, на каком из них?
(Устные ответы обучающихся)
Слово учителя
Верно. Тема урока “Сходство и различие в
строении клеток растений, животных, бактерий и
грибов”.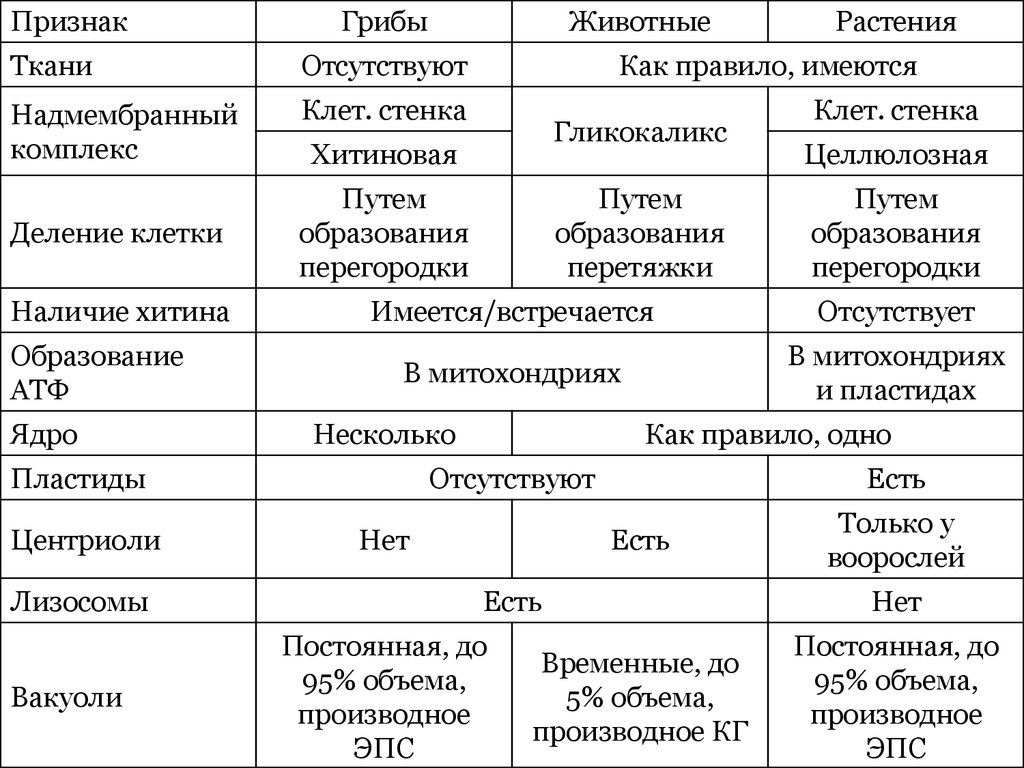
(Слайд 7)
Сегодня на уроке вам необходимо сравнить
строение растительной, грибной, бактериальной и
животной клетки.
У вас на столах лежат инструкции по выполнению
лабораторной работы.
Лабораторная работа (Слайд
8)
“Изучение строения растительной,
грибной, бактериальной и животной клеток под
микроскопом”
Цель: закрепить умение готовить
микропрепараты и рассматривать их под
микроскопом, находить особенности строения
клеток различных организмов, сравнивать их между
собой.
Оборудование: микроскопы, предметные
и покровные стекла, стаканы с водой, стеклянные
палочки, лук репчатый, разведенные дрожжи,
культура сенной палочки, микропрепараты клеток
многоклеточных животных.
Ход работы (Слайд 9)
- Приготовьте микропрепараты кожицы лука,
дрожжевых грибов, бактерии сенной палочки,
рассмотрите их под микроскопом. - Зарисуйте клетки в тетрадях и обозначьте
видимые в световой микроскоп органоиды.
- Рассмотрите готовый микропрепарат клеток
многоклеточного животного - (клетки эпителиальной ткани).
- Сопоставьте увиденное с изображением объектов
на таблицах. - Сравните между собой эти клетки. Ответьте в
выводе на следующие вопросы: - В чем заключается сходство и различие этих
клеток? - Каковы причины сходства и различия.
Поверка выполнения задания.
(Слайд 10)
IV. Первичное закрепление изученного
материала.
Слово учителя
— Перед вами лежат задания для каждой группы. На
эту работу вам отводится 5 минут.
(Слайд 11)
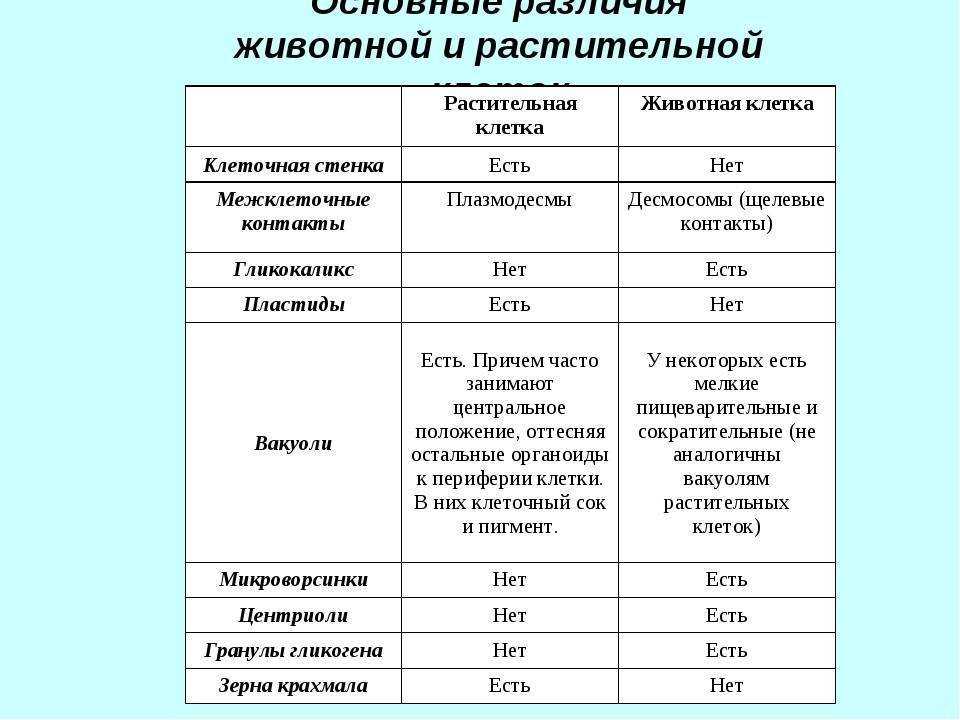
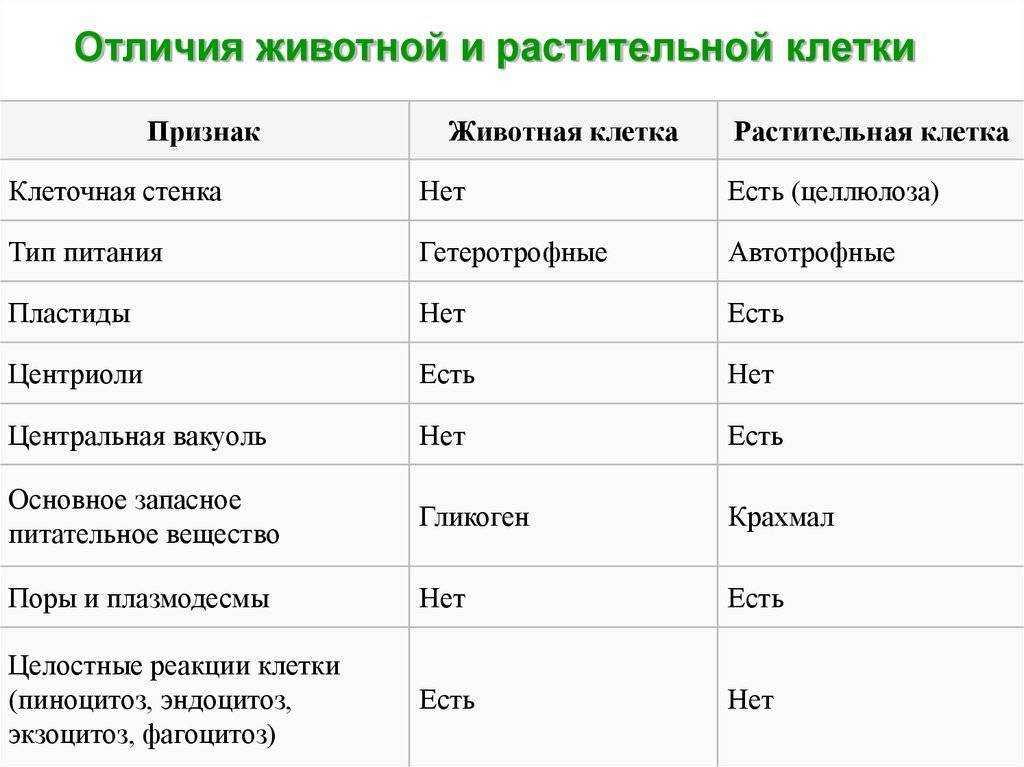
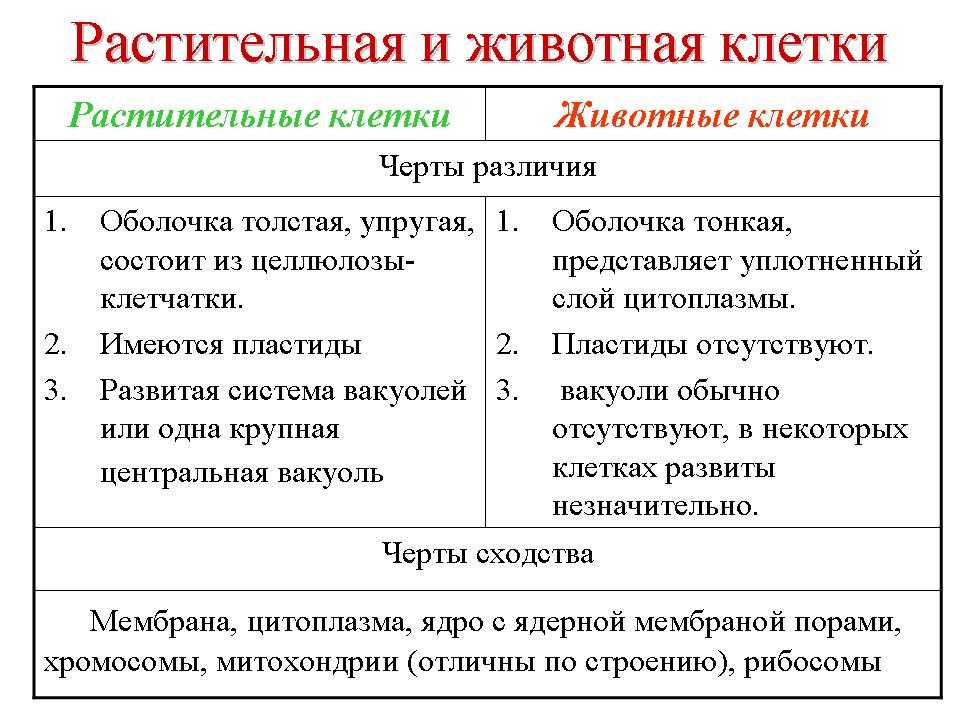
Задание 1. Найдите сходства и отличия в
строении у растительной и животной клетки.
Задание 2. О чем свидетельствует сходство в
строении клеток различных организмов?
Задание 3. О чем свидетельствует различие в
строении клеток различных организмов?
Задание 4. Докажите, что клетка является
основной структурной и функциональной единицей
живого.
Поверка выполнения задания
(Ответы обучающихся)
(Слайд 12, 13)
Слово учителя
— Какой вывод можно сделать?
(Ответы обучающихся)
(Слайд 14, 15)
V. Подведение итогов урока
Итак, мы расширили свои знания о строении
клетки и пришли к выводу, что единство клеточного
строения организмов находит подтверждение не
только в сходстве строения различных клеток, но и
в сходстве химического состава и метаболических
процессов. Нуклеиновые кислоты, белки, процессы
их синтеза и превращений универсальны и
принципиально близки в клетках всех живых
организмов.
VI. Выставление отметок.
Учащиеся сдают тетради с выполненной
лабораторной работой.
VII. Домашнее задание.
Подготовить сообщения и презентации на темы:
«Строение и жизнедеятельность растительной
клетки», «Строение и жизнедеятельность
животной клетки «, «История создания
клеточной теории».
23. 03.2009
03.2009
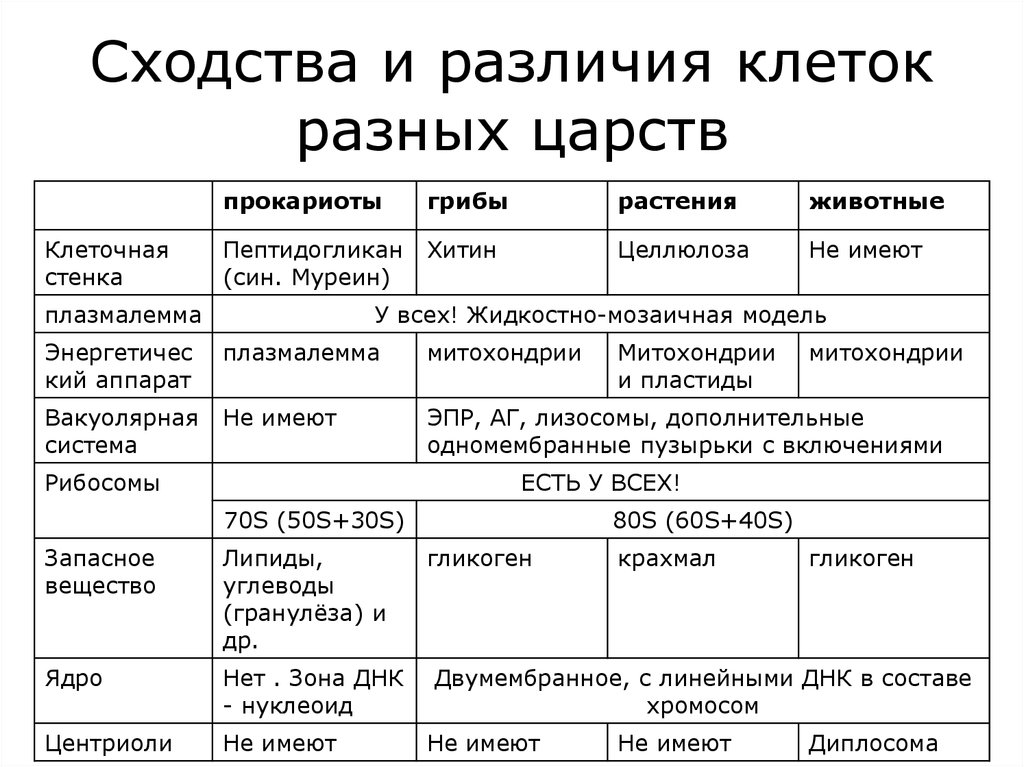
Различия клеток про- и эукариот, различия в строении клеток разных царств — что это, определение и ответ
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
Прокариоты (доядерные) – более простые по строению и возникли в процессе эволюции раньше;
Эукариоты (ядерные) – более сложные, возникли позже.
Клетки, составляющие тело человека, в основном, являются эукариотическими.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам:
● Общий план строения клетки: наличие клеточной мембраны, цитоплазмы, ядра
● Принципиальное сходство процессов обмена веществ и энергии в клетке
● Кодирование наследственной информации при помощи нуклеиновых кислот
● Единство химического состава клеток
● Сходные процессы деления клеток
Прокариоты (от лат. Pro – перед, до и греч. Κάρῠον – ядро, орех) – организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий).
Pro – перед, до и греч. Κάρῠον – ядро, орех) – организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий).
У прокариот генетический материал представлен кольцевой молекулой ДНК (нуклеоидом). Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи.
• Нуклеоид — не ограниченный мембранами участок цитоплазмы, в котором расположена кольцевая молекула ДНК — «бактериальная хромосома», где хранится весь генетический материал клетки.
· Плазмиды — небольшие дополнительные кольцевые молекулы ДНК, несущие обычно всего несколько генов. Плазмиды, в отличие от бактериальной хромосомы, не являются обязательным компонентом клетки. Обычно они придают бактерии определенные полезные для неё свойства, такие как устойчивость к антибиотикам.
Обычно они придают бактерии определенные полезные для неё свойства, такие как устойчивость к антибиотикам.
• Жгутики — органеллы движения некоторых бактерий. Бактериальный жгутик построен значительно проще эукариотического, и он в 10 раз тоньше, внешне не покрыт плазматической мембраной и состоит из одинаковых молекул белков, которые образуют цилиндр. В мембране жгутик закреплен при помощи базального тела.
• Пили — поверхностные структуры, присутствующие у многих бактериальных клеток и представляющие собой прямые белковые цилиндры. Пили участвуют в передаче генетического материала между бактериальными клетками (конъюгация).
• Споры (эндоспоры) — окруженные плотной оболочкой структуры, содержащие ДНК бактерии и обеспечивающее выживание в неблагоприятных условиях. К образованию спор способны лишь некоторые виды прокариот, в частности, возбудитель столбняка, возбудитель ботулизма и возбудитель сибирской язвы. Споры бактерий могут выдерживать довольно жесткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение.
Споры бактерий могут выдерживать довольно жесткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение.
• Слизистая капсула – слизистая структура, располагающаяся над клеточной стенкой, состоит из полисахаридов.
Эукариоты (от греч. Ευ – хорошо, полностью и κάρῠον – ядро, орех) – организмы, обладающие оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой.
Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства комплекс с белками-гистонами, называемый хроматином.
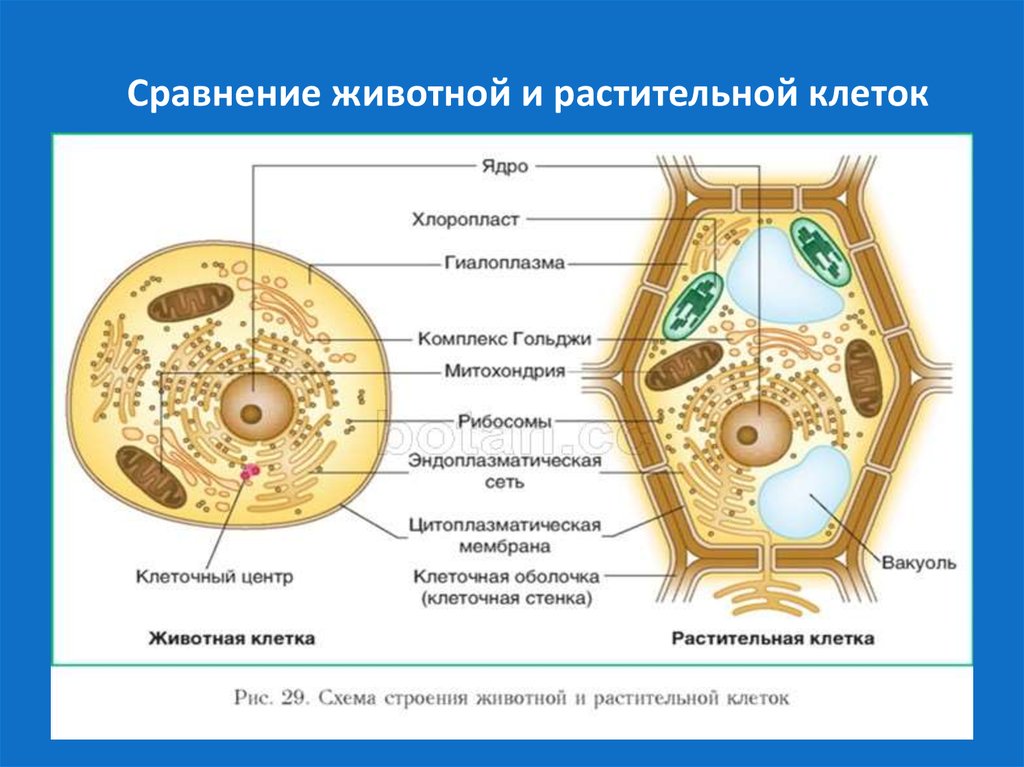
В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты – митохондрии, а у водорослей и растений – пластиды.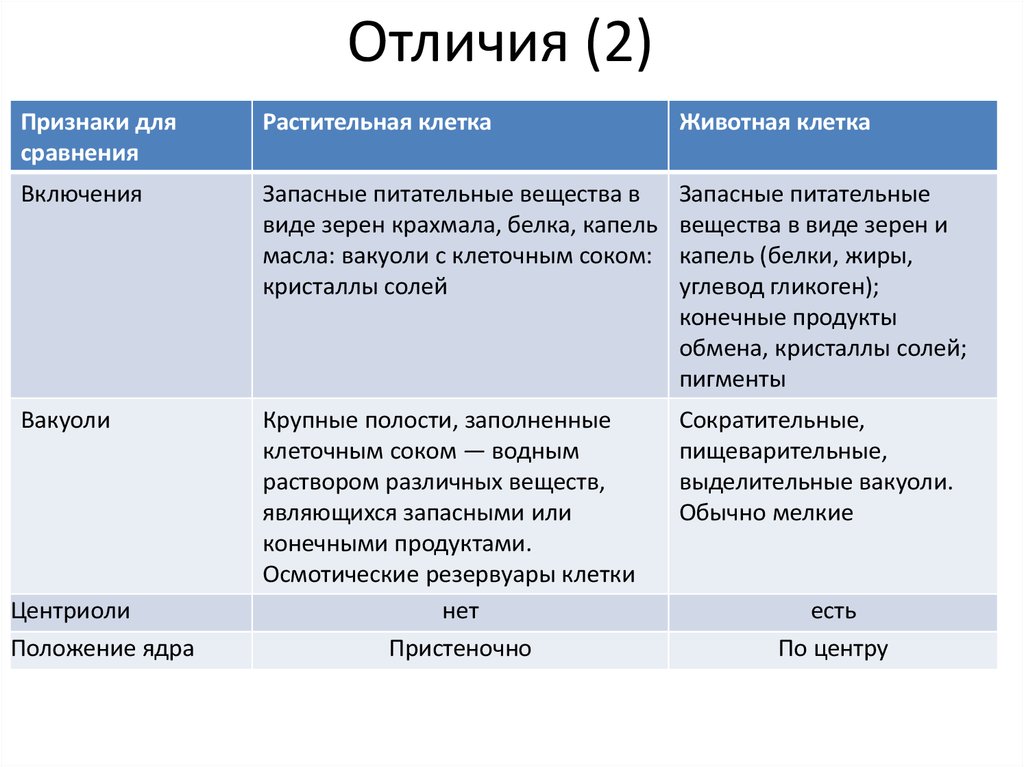
| Прокариоты | Эукариоты |
| Средний диаметр 0,5-10 мкм | Средний диаметр 10-100 мкм |
| Имеют одну кольцевую молекулу ДНК, размещенную в цитоплазме | Имеют несколько линейных молекул ДНК – хромосом, локализованных в ядре |
| Деление клетки происходит посредством бинарного деления, без образования веретена деления. | Деление клетки происходит путем мейоза или митоза. |
| Отсутствуют мембранные органоиды, иногда плазмалемма образует выпячивание внутрь клетки | Присутствует большое количество мембранных органелл |
В природе выделяют четыре царства живой природы.
Царство Бактерии
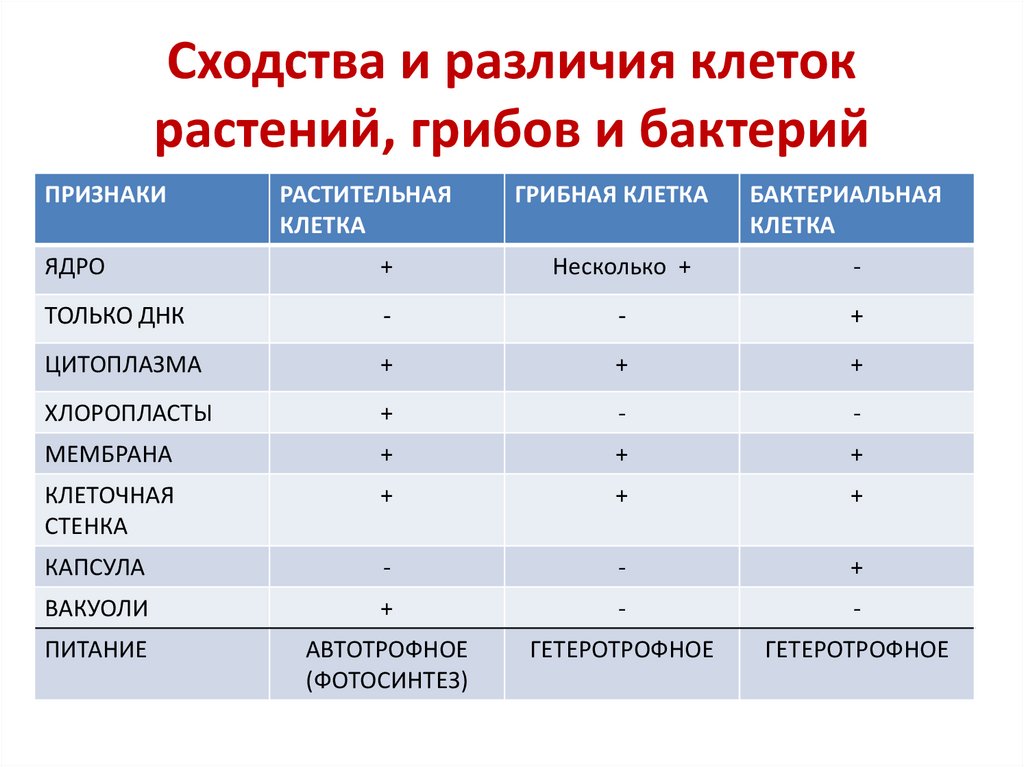
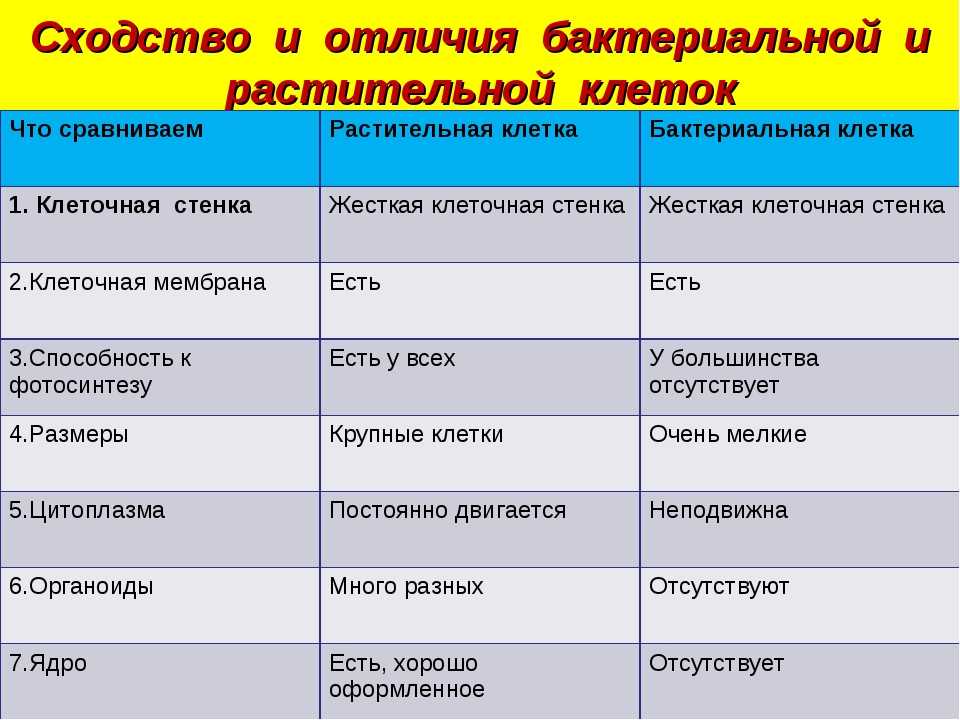
Бактериальная клетка – типичная клетка прокариот.
1. Ядерный аппарат. Кольцевая ДНК – нуклеоид. Есть плазмиды.
2. Клеточная стенка из муреина.
Клеточная стенка из муреина.
3. Не имеет мембранных органоидов. Есть только рибосомы, редко бывают вакуоли.
4. Запасает различные полисахариды.
5. По способу питания может быть, как автотрофом, так и гетеротрофом.
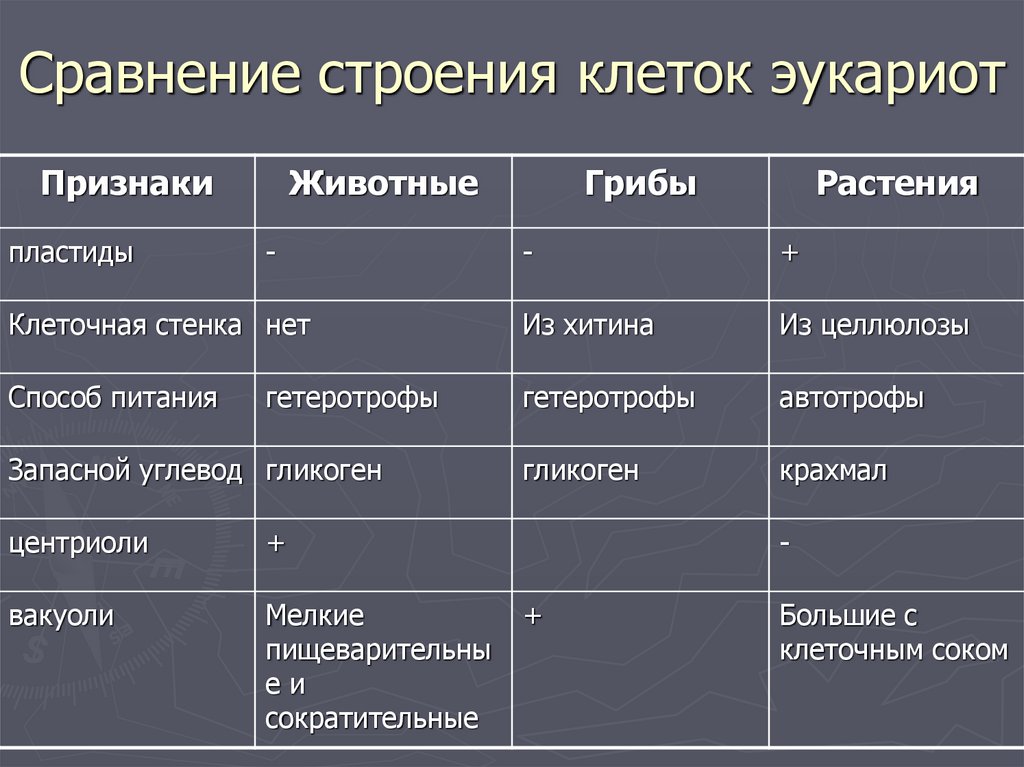
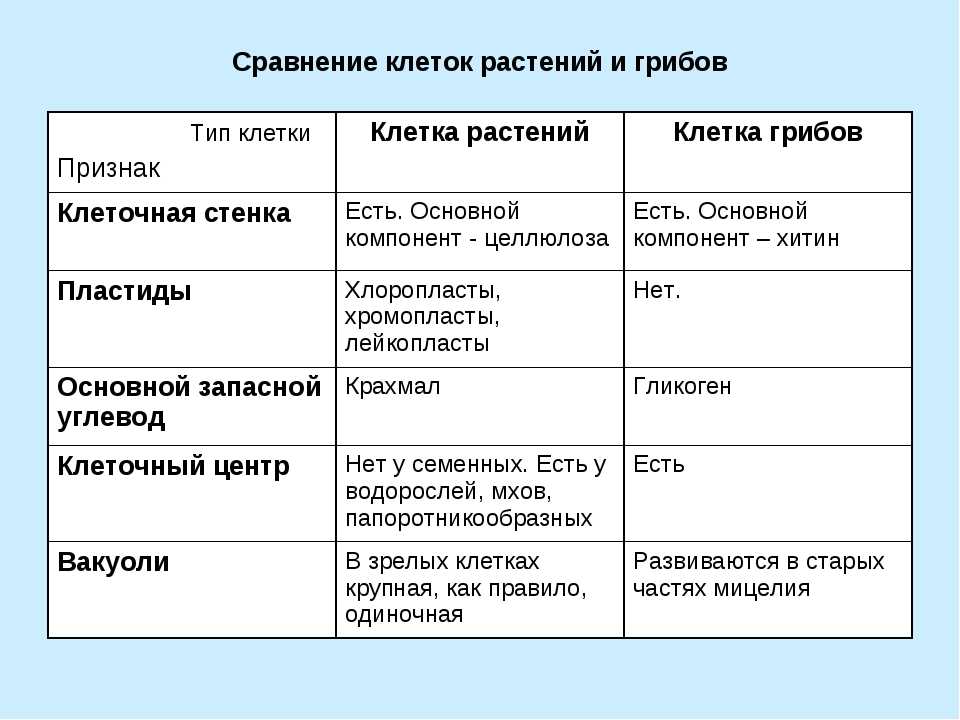
Царство Грибы
Клетка грибов:
1. Имеет ядро. Линейные хромосомы.
2. Клеточная стенка есть, содержит хитин.
3. Имеет мембранные органоиды. Вакуоли развиваются у старых клеток.
4. Запасающий углевод – гликоген.
5. По способу питания гетеротрофы.
Тело гриба образовано нитевидными структурами в один ряд клеток – гифами. У некоторых грибов перегородки между клетками утрачиваются, и возникает грибница, состоящая из одной гигантской многоядерной клетки. Грибы не способны к активному движению, зато они могут расти неограниченно – это признаки, которые объединяют грибы с растениями.
Царство Растения
Клетка растений:
1. Имеет ядро. Линейные хромосомы.
Имеет ядро. Линейные хромосомы.
2. Клеточная стенка есть, содержит целлюлозу.
3. Имеет мембранные органоиды.
Есть крупные вакуоли с клеточным соком и пластиды – главное отличие растительной клетки от других.
4. Запасающий углевод – крахмал.
5. По способу питания автотрофы.
Для обеспечения фотосинтеза в клетках растений содержатся пластиды, несущие хлорофилл и другие пигменты.
Царство Животные
Клетка животных:
1. Имеет ядро. Линейные хромосомы.
2. Клеточная стенка отсутствует. Поэтому клетка животных способна к фаго –и пиноцитозу. Имеет гликокаликс, который выполняет рецепторную и маркерную функции.
3. Имеет мембранные органоиды. Есть клеточный центр. В некоторых клетках есть мелкие сократительные и пищеварительные вакуоли.
4. Запасающий углевод – гликоген.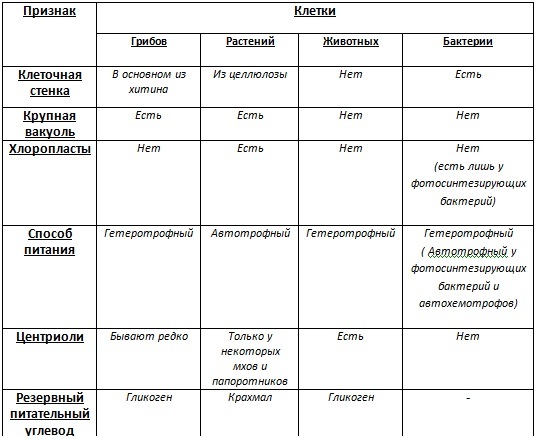
5. По способу питания гетеротрофы.
Для лучшего запоминания закрепим информацию о клетках разных царств в таблице.
Недавнее удивительное сходство между растительными клетками и нейронами
. 2010 февраль;5(2):87-9.
doi: 10.4161/psb.5.2.11237.
Epub 2010 19 февраля.
Франтишек Балуска
1
принадлежность
- 1 IZMB, Боннский университет, Бонн, Германия. [email protected]
PMID:
20150757
PMCID:
PMC2884105
DOI:
10.
4161/псб.5.2.11237
 4161/псб.5.2.11237
4161/псб.5.2.11237Бесплатная статья ЧВК
Франтишек Балуска.
Поведение сигналов растений.
2010 9 февраля0003
Бесплатная статья ЧВК
. 2010 февраль;5(2):87-9.
doi: 10.4161/psb.5.2.11237.
Epub 2010 19 февраля.
Автор
Франтишек Балуска
1
принадлежность
- 1 IZMB, Боннский университет, Бонн, Германия. [email protected]
PMID:
20150757
PMCID:
PMC2884105
DOI:
10.
4161/псб.5.2.11237
 4161/псб.5.2.11237
4161/псб.5.2.11237Абстрактный
Растительные клетки и нейроны имеют несколько общих черт, включая нецентросомные микротрубочки, подвижные пост-Гольджи органеллы, отделенные как пространственно/структурно, так и функционально от аппарата Гольджи и участвующие в везикулярной эндоцитарной рециркуляции, а также домены межклеточной адгезии на основе актина. /миозиновый цитоскелет, служащий для межклеточной коммуникации. Клетки растений, растущие на кончиках, такие как корневые волоски и пыльцевые трубки, также напоминают нейроны, расширяющие свои аксоны. Недавно были сделаны удивительные открытия в отношении молекулярной основы нейродегенеративных заболеваний, известных как наследственная спастическая параплегия, и рост корневых волосков на кончиках. Все эти достижения кратко обсуждаются в контексте других сходств между растительными клетками и нейронами.
Похожие статьи
Новые аспекты организации ER при росте кончиков корневых волос: уроки RHD3 и Atlastin.
Чжэн Х., Чен Дж.
Чжэн Х и др.
Поведение сигналов растений. 2011 ноябрь;6(11):1710-3. doi: 10.4161/psb.6.11.17477. Epub 2011 1 ноября.
Поведение сигналов растений. 2011.PMID: 22057320
Бесплатная статья ЧВК.Рассмотрение.
Транспорт везикул, цитоскелет и передача сигналов в корневых волосках и пыльцевых трубках.
Самай Дж., Мюллер Дж., Бек М., Бём Н., Менцель Д.
Самай Дж. и др.
Тенденции Растениевод. 2006 Декабрь; 11 (12): 594-600. doi: 10.1016/j.tplants.2006.10.002. Epub 2006 7 ноября.
Тенденции Растениевод. 2006.PMID: 17092761
Рассмотрение.
Глутаматная сигнализация в корнях.
Форде БГ.
Форде БГ.
J Опытный бот. 2014 март; 65(3):779-87. дои: 10.1093/jxb/ert335. Epub 2013 22 октября.
J Опытный бот. 2014.PMID: 24151303
Рассмотрение.
Фармакологическое восстановление дефектов роста аксонов в модели иПСК человека наследственной спастической параплегии SPG3A.
Zhu PP, Denton KR, Pierson TM, Li XJ, Blackstone C.
Чжу П.П. и др.
Хум Мол Жене. 2014 1 ноября; 23 (21): 5638-48. дои: 10.1093/хмг/дду280. Epub 2014 6 июня.
Хум Мол Жене. 2014.PMID: 24908668
Бесплатная статья ЧВК.Белок, активирующий ГТФазу, фактор АДФ-рибозилирования класса I, имеет решающее значение для поддержания направленного роста корневых волосков у арабидопсиса.
Yoo CM, Wen J, Motes CM, Sparks JA, Blancaflor EB.
Ю КМ и др.
Завод Физиол. 2008 г., август; 147 (4): 1659-74. doi: 10. 1104/стр.108.119529. Epub 2008 6 июня.
Завод Физиол. 2008.PMID: 18539780
Бесплатная статья ЧВК.
 Epub 2013 22 октября.
Epub 2013 22 октября. 1104/стр.108.119529. Epub 2008 6 июня.
1104/стр.108.119529. Epub 2008 6 июня.Посмотреть все похожие статьи
Цитируется
Наведение мостов: электрофизиологическая связь между растениями, опосредованная мицелием.
Томас М.А., Купер Р.Л.
Томас М.А. и соавт.
Поведение сигналов растений. 2022 31 декабря; 17 (1): 2129291. дои: 10.1080/15592324.2022.2129291.
Поведение сигналов растений. 2022.PMID: 36384396
Бесплатная статья ЧВК.Открытие на основе растительной мутантной модели хантингтина нарушенной экспрессии GTPCH и DHFR.
Хун С.И., Чжу С., Киттур Ф.С., Хе М., Арнинг Э., Чжан Дж., Джонсон А.Дж., Джава Г.С., Томас М.Д., Дин Т.
Т., Се Дж.
Хун С.И. и соавт.
Cell Mol Life Sci. 2022 17 октября; 79 (11): 553. doi: 10.1007/s00018-022-04587-6.
Cell Mol Life Sci. 2022.PMID: 36251090
Бесплатная статья ЧВК.Внедрение робототехники, вдохновленной растениями.
Ли Дж., Кальво П.
Ли Дж. и др.
Передний нейроробот. 2022 31 января; 15:772012. doi: 10.3389/fnbot.2021.772012. Электронная коллекция 2021.
Передний нейроробот. 2022.PMID: 35173596
Бесплатная статья ЧВК.Наши сестры растения? заметки из филогенетики и ботаники о слепоте родства растений.
Буто Ф., Гресильон Э., Шартье Д., Арбеле-Боннин Д., Кавано Т., Балушка Ф., Манкузо С., Кальво П., Лауренти П.
Буто Ф. и др.
Поведение сигналов растений. 2 декабря 2021 г . ; 16 (12): 2004769. дои: 10.1080/15592324.2021.2004769. Epub 2021 16 декабря.
Поведение сигналов растений. 2021.PMID: 34913409
Бесплатная статья ЧВК.Два — это компания, но четыре — это вечеринка — проблемы тетраплоидизации для динамики клеточных стенок и эффективного роста кончиков пыльцы.
Вестерманн Дж.
Вестерманн Дж.
Растения (Базель). 2021 5 ноября; 10 (11): 2382. doi: 10.3390/plants10112382.
Растения (Базель). 2021.PMID: 34834745
Бесплатная статья ЧВК.Рассмотрение.
 Т., Се Дж.
Т., Се Дж.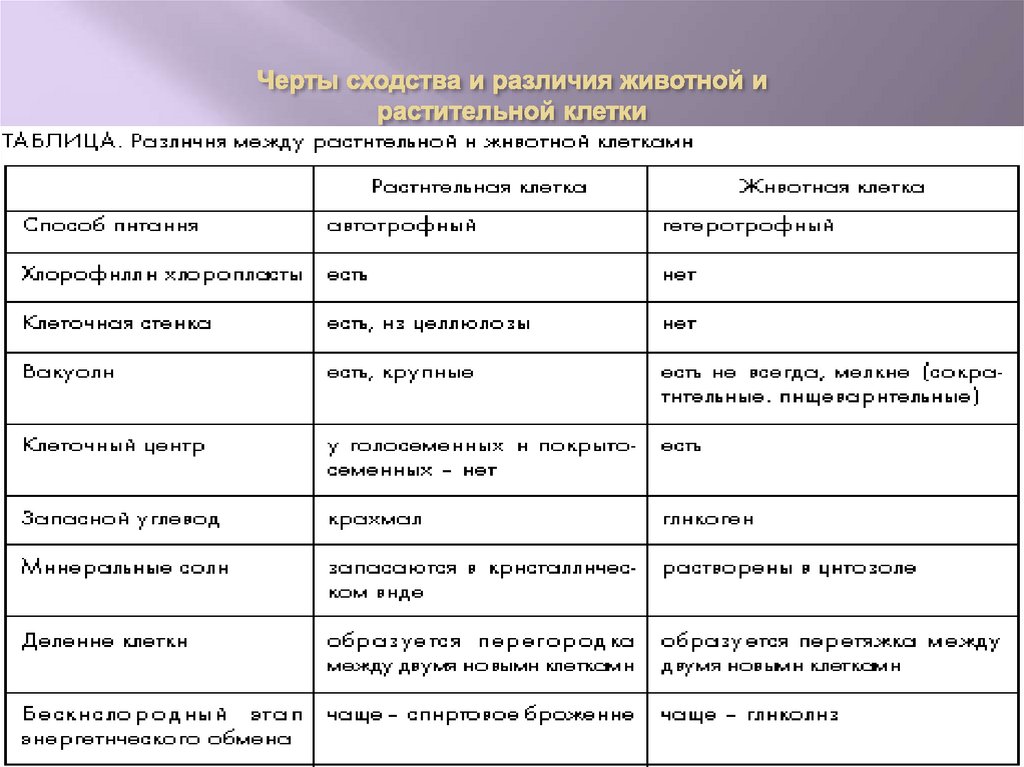 ; 16 (12): 2004769. дои: 10.1080/15592324.2021.2004769. Epub 2021 16 декабря.
; 16 (12): 2004769. дои: 10.1080/15592324.2021.2004769. Epub 2021 16 декабря.Просмотреть все статьи «Цитируется по»
термины MeSH
вещества
Недавние удивительные сходства между растительными клетками и нейронами
1. Palanivelu R, Preuss D. Ориентация на пыльцевые трубки и направление аксонов: параллели в механизмах роста кончиков. Тенденции клеточной биологии. 2000; 10: 517–524. [PubMed] [Google Scholar]
Тенденции клеточной биологии. 2000; 10: 517–524. [PubMed] [Google Scholar]
2. Лев-Ядун С. Интрузивный рост — растительный аналог роста дендритов и аксонов у животных. Новый Фитол. 2001; 150: 508–512. [Google Scholar]
3. Бреннер Э., Штальберг Р., Манкузо С., Виванко Дж., Балушка Ф., Ван Волкенбург Э. Нейробиология растений: интегрированный взгляд на сигнализацию растений. Тенденции Растениевод. 2006; 11: 413–419.. [PubMed] [Google Scholar]
4. Балушка Ф., Фолькманн Д., Менцель Д. Синапсы растений: домены адгезии на основе актина для межклеточной коммуникации. Тенденции Растениевод. 2005; 10:106–111. [PubMed] [Google Scholar]
5. Baluška F, Hlavacka A. Растительные формины стареют: что-то особенное в поперечных стенках. Новый Фитол. 2005; 168: 499–503. [PubMed] [Google Scholar]
6. Балушка Ф., Шамай Дж., Войташек П., Фолькманн Д., Менцель Д. Цитоскелет — плазматическая мембрана — континуум клеточной стенки у растений: новый взгляд на возникающие связи. Завод Физиол. 2003; 133: 482–49.1. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Завод Физиол. 2003; 133: 482–49.1. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Barlow PW, Volkmann D, Baluška F. Полярность корней. В: Линдси К., редактор. Полярность растений. Издательство Блэквелл; 2004. стр. 192–241. [Google Scholar]
8. Балушка Ф., Фолькманн Д., Барлоу П.В. Эукариотические клетки и их клеточные тела: новый взгляд на клеточную теорию. Энн Бот. 2004; 94:9–32. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Шамай Дж., Рид Н.Д., Фолькманн Д., Мензель Д., Балушка Ф. Эндоцитарная сеть у растений. Тенденции клеточной биологии. 2005; 15: 425–433. [PubMed] [Академия Google]
10. Horton AC, Rácz B, Monson EE, Lin AL, Weinberg RJ, Ehlers MD. Поляризованный секреторный транспорт направляет груз для асимметричного роста дендритов и морфогенеза. Нейрон. 2005; 48: 757–771. [PubMed] [Google Scholar]
11. Прюдз К., Дик Г., Твейт Х. Сколько путей через лабиринт Гольджи? Трафик. 2008: 299–304. [PubMed] [Google Scholar]
12. Тан Б.Л. Новые аспекты мембранного трафика при росте нейрональных дендритов. Биохим Биофиз Акта. 2008; 1783: 169–176. [PubMed] [Академия Google]
Тан Б.Л. Новые аспекты мембранного трафика при росте нейрональных дендритов. Биохим Биофиз Акта. 2008; 1783: 169–176. [PubMed] [Академия Google]
13. Lam SK, Cai Y, Tse YC, Wang J, Law AHY, Pimpl P, et al. Индуцированные BFA компартменты из аппарата Гольджи и транс-сети Гольджи/ранней эндосомы различны в растительных клетках. Плант Дж. 2009; 60: 865–881. [PubMed] [Google Scholar]
14. Dettmer J, Hong-Hermesdorf A, Stierhof YD, Schumacher K. Vacuolar H + -АТФазная активность необходима для эндоцитарного и секреторного транспорта арабидопсиса. Растительная клетка. 2006; 18: 715–730. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Šamaj J, Baluška F, Voigt B, Schlicht M, Volkmann D, Menzel D. Эндоцитоз, актиновый цитоскелет и передача сигналов. Завод Физиол. 2004; 135:1150–1161. [PMC free article] [PubMed] [Google Scholar]
16. Baluška F, Šamaj J, Menzel D. Полярный транспорт ауксина: транспорт-опосредованный поток через плазматическую мембрану или секреция, подобная нейротрансмиттеру? Тенденции клеточной биологии.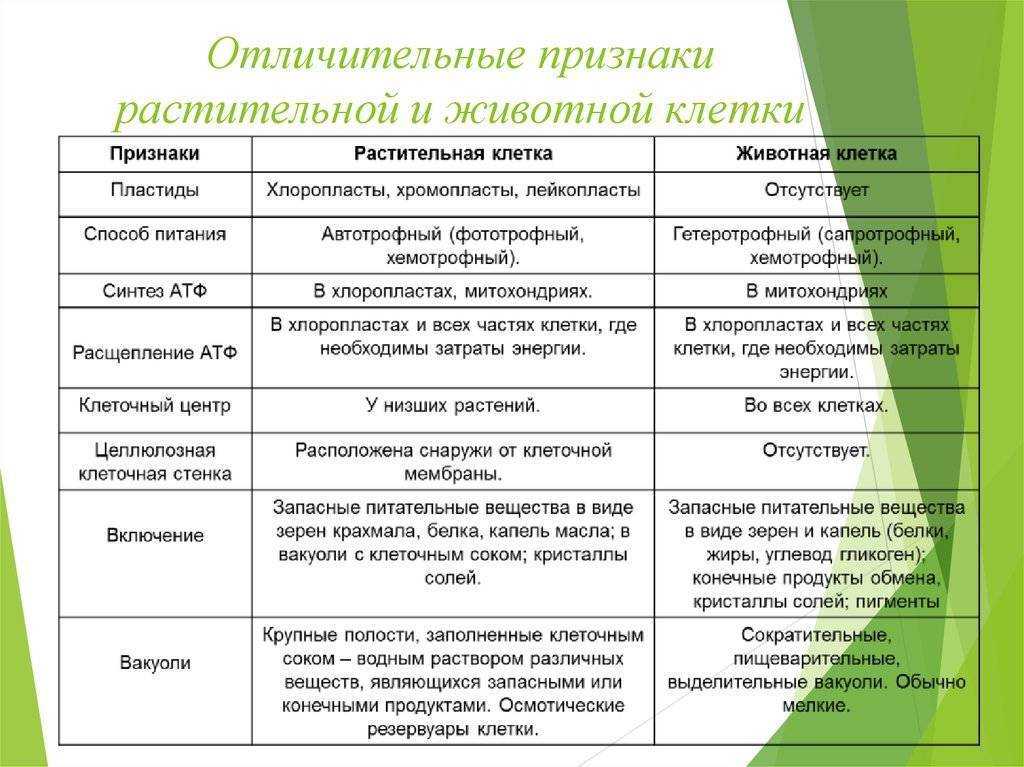 2003; 13: 282–285. [PubMed] [Google Scholar]
2003; 13: 282–285. [PubMed] [Google Scholar]
17. Favre P, Degli Agosti R. Потенциал-зависимые потенциалы действия в Arabidopsis thaliana . Завод Физиол. 2007; 131: 263–272. [PubMed] [Google Scholar]
18. Felle HH, Zimmermann MR. Системная передача сигналов в ячмене через потенциалы действия. Планта. 2007; 226: 203–214. [PubMed] [Google Scholar]
19. Stolarz M, Król E, Dziubinska H, Kurenda A. Глутамат индуцирует серию потенциалов действия и снижение скорости циркуляции у Helianthus annuus. Завод Физиол. 2010 В печати. [PubMed] [Google Scholar]
20. Masi E, Ciszak M, Stefano G, Renna L, Azzarello E, Pandolfi C, et al. Пространственно-временная динамика активности электрической сети в верхушке корня. Proc Natl Acad Sci USA. 2009 г.;106:4048–4053. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Салинас С., Проукакис С., Кросби А., Уорнер Т.Т. Наследственная спастическая параплегия: клиника и патогенетические механизмы. Ланцет Нейрол. 2008;7:1127–1138. [PubMed] [Google Scholar]
2008;7:1127–1138. [PubMed] [Google Scholar]
22. Lee Y, Paik D, Bang S, Kang J, Chun B, Lee S, et al. Потеря гена спастической параплегии атластина вызывает возрастную гибель дофаминергических нейронов у дрозофилы. Нейробиол Старение. 2008; 29:84–94. [PubMed] [Академия Google]
23. Hu J, Shibata Y, Zhu PP, Voss C, Rismanchi N, Prinz W, et al. Класс динаминоподобных ГТФаз, участвующих в формировании тубулярной сети ЭР. Клетка. 138: 549–561. [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Wang H, Lockwood SK, Hoeltzel MF, Schiefelbein JW. Ген ROOT HAIR DEFECTIVES кодирует эволюционно консервативный белок с GTP-связывающими мотивами и необходим для регулируемого увеличения клеток у арабидопсиса. Генерал Дев. 1997; 11: 799–811. [PubMed] [Академия Google]
25. Ван Х., Ли М.М., Шифельбейн Дж.В. Регуляция гена экспансии клеток RHD3 в процессе развития арабидопсиса. Завод Физиол. 2002; 129: 638–649. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Yuen CYL, Sedbrook JC, Perrin RM, Carroll KL, Masson PH. Мутации с потерей функции ROOT HAIR DEFECTIVE3 подавляют волнистость корней, перекос и ротацию эпидермальных клеток у Arabidopsis. Завод Физиол. 2005; 138: 701–714. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Yuen CYL, Sedbrook JC, Perrin RM, Carroll KL, Masson PH. Мутации с потерей функции ROOT HAIR DEFECTIVE3 подавляют волнистость корней, перекос и ротацию эпидермальных клеток у Arabidopsis. Завод Физиол. 2005; 138: 701–714. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Rismanchi N, Soderblom C, Stadler J, Zhu PP, Blackstone C. Atlastin GTPases необходимы для аппарата Гольджи и морфогенеза ER. Хум Мол Жене. 2008;17:1591–1604. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Hu Y, Zhong R, Morrison WH, 3rd, Ye ZH. Ген RHD3 арабидопсиса необходим для биосинтеза клеточной стенки и организации актина. Планта. 2003; 217:912–921. [PubMed] [Google Scholar]
29. Zheng H, Kunst L, Hawes C, Moore I. Анализ на основе GFP показывает роль RHD3 в транспорте между эндоплазматическим ретикулумом и аппаратом Гольджи. Плант Дж. 2004; 37: 398–414. [PubMed] [Google Scholar]
30. Эванс К., Келлер С., Павур К., Глазго К., Конн Б., Лоринг Б. Взаимодействие двух генных продуктов наследственной спастической параплегии, спастина и атластина, предполагает общий путь для поддержания ионона.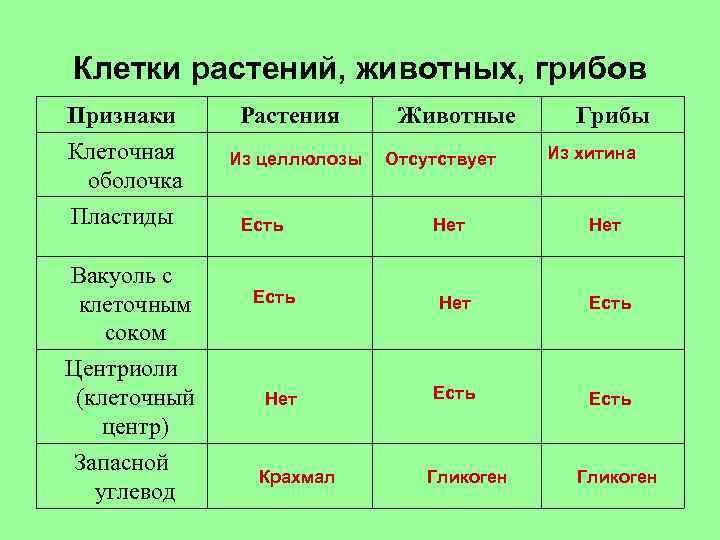 Proc Natl Acad Sci USA. 103:10666–10671. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Proc Natl Acad Sci USA. 103:10666–10671. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Zhu PP, Soderblom C, Tao-Cheng JH, Stadler J, Blackstone C. Белок SPG3A атластин-1 обогащен конусами роста и способствует удлинению аксонов во время развития нейронов. Хум Мол Жене. 2006; 15:1343–1353. [PubMed] [Google Scholar]
32. Lee M, Paik SK, Lee MJ, Kim YJ, Kim S, Nahm M, et al. Атластин дрозофилы регулирует стабильность мышечных микротрубочек и необходим для развития синапсов. Дев биол. 2009; 330: 250–262. [PubMed] [Google Scholar]
33. Тихонов Д.Б., Магазинник Л.Г. Происхождение и молекулярная эволюция ионотропных рецепторов глутамата. Neurosci Behav Physiol. 2009 г.;39:763–773. [PubMed] [Google Scholar]
34. Балушка Ф., Манкузо С. Глубокие эволюционные истоки нейробиологии: переворачивание сущности «нейронного» с ног на голову. Коммун Интегр Биол. 2009;2:60–65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Давенпорт Р. Рецепторы глутамата в растениях.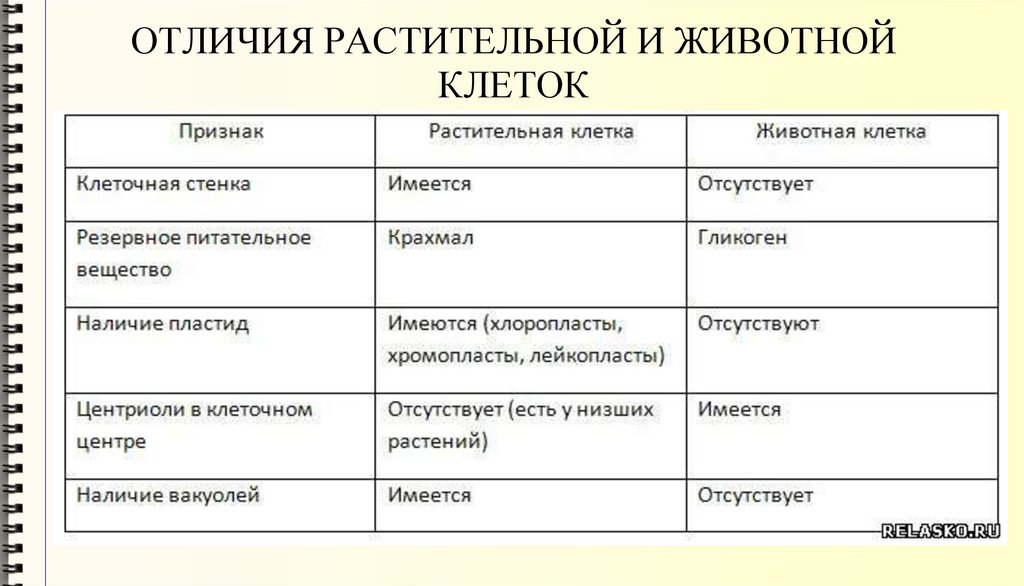 Энн Бот. 2002; 90: 549–557. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Энн Бот. 2002; 90: 549–557. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Gilliham M, Campbell M, Dubois C, Becker D, Davenport R. Семейство глутамат-подобных рецепторов Arabidopsis thaliana (AtGLR) In: Baluška, Mancuso S, Volkmann Д, редакторы. Коммуникация в растениях. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 187–204. [Академия Google]
37. Tapken D, Hollmann M. Функция ионных каналов рецептора глутамата Arabidopsis thaliana, продемонстрированная трансплантацией ионных пор. Дж Мол Биол. 2008; 383:36–48. [PubMed] [Google Scholar]
38. Walch-Liu P, Liu LH, Remans T, Tester M, Forde BG. Доказательства того, что L-глутамат может действовать как эндогенный сигнал для модуляции роста и ветвления корней у Arabidopsis thaliana. Физиология клеток растений. 2006;47:1045–1057. [PubMed] [Google Scholar]
39. Форде Б.Г., Леа П. Глутамат в растениях: метаболизм, регуляция и передача сигналов. J Опытный бот. 2007;58:2339–2358. [PubMed] [Google Scholar]
40. Li J, Zhu S, Song X, Shen Y, Chen H, Yu J и др. Ген, подобный рецептору глутамата риса, имеет решающее значение для деления и выживания отдельных клеток в апикальной меристеме корня. Растительная клетка. 2006; 18: 340–349. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Li J, Zhu S, Song X, Shen Y, Chen H, Yu J и др. Ген, подобный рецептору глутамата риса, имеет решающее значение для деления и выживания отдельных клеток в апикальной меристеме корня. Растительная клетка. 2006; 18: 340–349. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Балушка Ф., Фолькманн Д., Барлоу П.В. Перекресток полярности в переходной зоне роста верхушек корней кукурузы: последствия для цитоскелета и развития. J Регулятор роста растений. 2001; 20: 170–181. [Академия Google]
42. Verbelen J-P, De Cnodder T, Le J, Vissenberg K, Baluška F. Верхушка корня Arabidopsis thaliana состоит из четырех отдельных зон клеточной активности: меристематическая зона, переходная зона, зона быстрого удлинения и зона прекращения роста. Поведение сигналов растений. 2006; 1: 296–304. [Статья бесплатно PMC] [PubMed] [Google Scholar]
43. Сивагуру М., Пайк С., Гассманн В., Баскин Т. Алюминий быстро деполимеризует микротрубочки и деполяризует плазматическую мембрану: свидетельство того, что ответы опосредованы глутаматным рецептором. Физиология клеток растений. 2003; 44: 667–675. [PubMed] [Академия Google]
Физиология клеток растений. 2003; 44: 667–675. [PubMed] [Академия Google]
44. Де Смет И., Тецумура Т., Де Рибе Б., Фрей Н.Ф., Лаплаз Л., Касимиро И. и др. Ауксин-зависимая регуляция положения боковых корней в базальной меристеме арабидопсиса. Разработка. 2007; 134: 681–690. [PubMed] [Google Scholar]
45. Brenner ED, Martinez-Barboza N, Clark AP, Liang QS, Stevenson DW, Coruzzi GM. Мутанты арабидопсиса, устойчивые к S(+)-бета-метил-альфа, бетадиаминопропионовой кислоте, агонисту рецептора глутамата, полученному из саговника. Завод Физиол. 2000; 124:1615–1624. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Бреннер ЭД, Файнберг П, Рунко С, Коруцци ГМ. Мутация в протеосомной регуляторной частице ААА-АТФазе-3 у арабидопсиса нарушает светоспецифическую реакцию удлинения гипокотиля, вызываемую агонистом рецептора глутамата, ВМАА. Дж Мол Биол. 2009; 70: 523–533. [PubMed] [Google Scholar]
47. Иллеш П., Шлихт М., Павловкин Дж., Лихтшайдль И., Балушка Ф., Овецка М.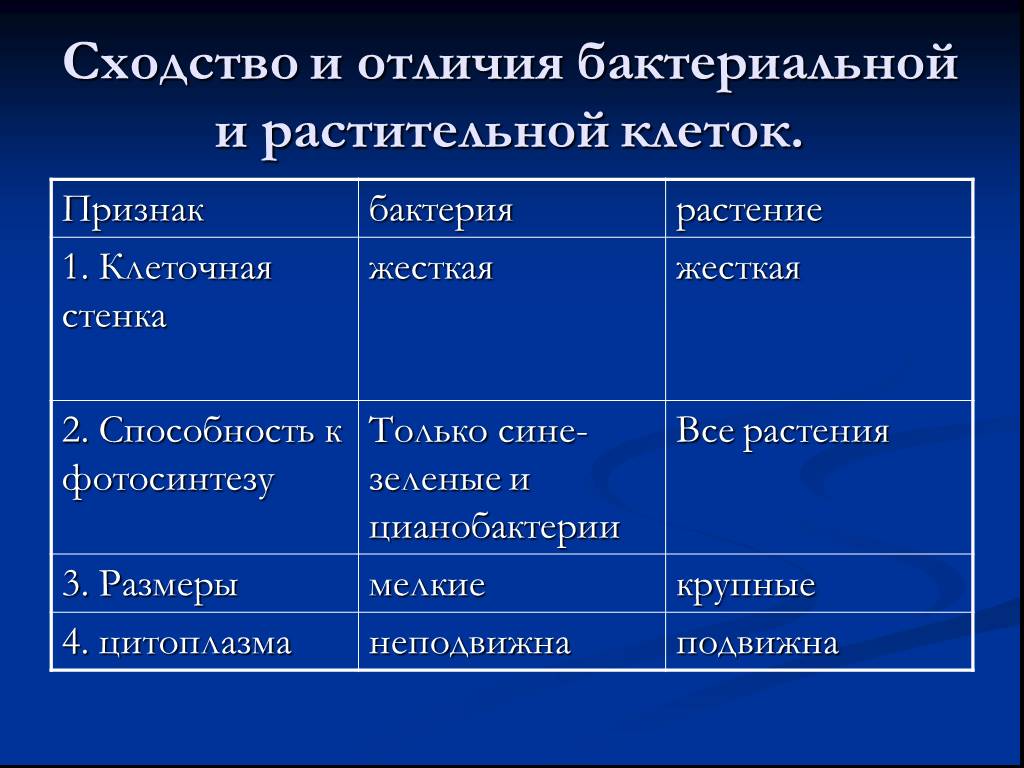 Токсичность алюминия в растениях: интернализация алюминия в клетки переходной зоны в верхушках корней арабидопсиса связана с изменениями в потенциале плазматической мембраны, эндосомальном поведении и продукции оксида азота. J Опытный бот. 2006;57:4201–4213. [PubMed] [Академия Google]
Токсичность алюминия в растениях: интернализация алюминия в клетки переходной зоны в верхушках корней арабидопсиса связана с изменениями в потенциале плазматической мембраны, эндосомальном поведении и продукции оксида азота. J Опытный бот. 2006;57:4201–4213. [PubMed] [Академия Google]
48. Ринкон-Захари М., Тистер Н.Д., Спаркс Дж.А., Валстер А.Х., Мотес К.М., Бланкафлор Э.Б. Сенсибилизированное излучение флуоресцентной резонансной энергии Yellow Cameleon 3.60 выявляет специфичные для корневой зоны кальциевые сигнатуры арабидопсиса в ответ на алюминий и другие трехвалентные катионы. Завод Физиол. 2010 В печати. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Sivaguru M, Horst WJ. Дистальная часть переходной зоны является наиболее чувствительной к алюминию апикальной корневой зоной кукурузы. Завод Физиол. 1998;116:155–163. [Google Scholar]
50. Сивагуру М., Балушка Ф., Фолькманн Д., Фелле Х., Хорст В.Дж. Воздействие алюминия на цитоскелет верхушки корня кукурузы: кратковременное воздействие на дистальную часть переходной зоны.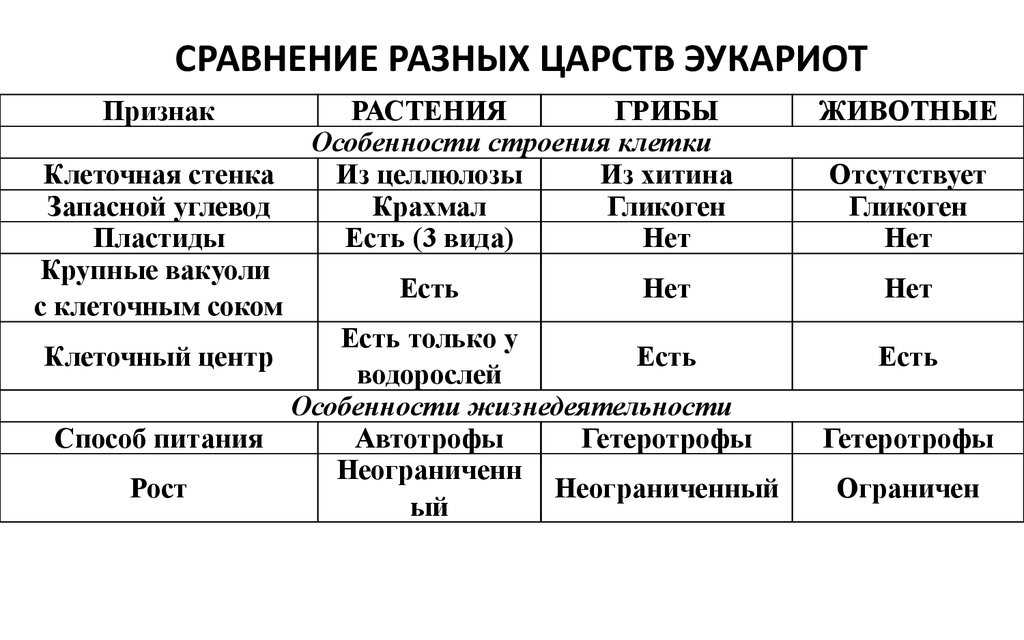 Завод Физиол. 1999;119:1073–1082. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Завод Физиол. 1999;119:1073–1082. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Kollmeier M, Felle HH, Horst WJ. Генотипические различия в устойчивости кукурузы к алюминию выражены в дистальной части переходной зоны. Участвует ли сниженный поток базипетального ауксина в ингибировании удлинения корня алюминием? Завод Физиол. 2000;122:945–956. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Дончева С., Аменос М., Пошенридер С., Барсело Дж. Структурирование корневых клеток: основная цель токсичности алюминия в кукурузе. J Опытный бот. 2005;56:1213–1220. [PubMed] [Google Scholar]
53. Джонс Д.Л., Гилрой С., Ларсен П.Б., Хауэлл С.Х., Кочиан Л.В. Влияние алюминия на цитоплазматический гомеостаз Ca 2+ в корневых волосках Arabidopsis thaliana (L.) Planta. 1998; 206: 378–387. [PubMed] [Академия Google]
54. Ма Л.Г., Фан QS, Ю. ZQ, Чжоу Х.Л., Чжан Ф.С., Сунь Д.Ю. Ингибирует ли алюминий прорастание пыльцы через внеклеточный кальмодулин? Физиология клеток растений.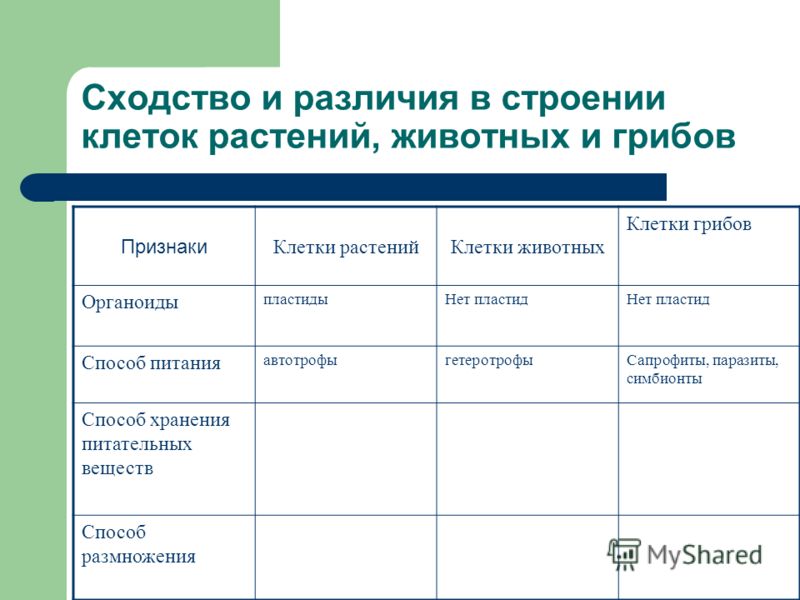 2000;41:372–376. [PubMed] [Google Scholar]
2000;41:372–376. [PubMed] [Google Scholar]
55. Бабурина О., Ренгель З. Поглощение алюминия клетками корней арабидопсиса, измеренное с помощью флуоресцентной визуализации. Энн Бот. 2009; 104: 189–195. [Статья бесплатно PMC] [PubMed] [Google Scholar]
56. Shen H, Hou NY, Schlicht M, Wan Y, Mancuso S, Baluška F. Токсичность алюминия нацелена на PIN2 в верхушках корней арабидопсиса: влияние на эндоцитоз PIN2, везикулярную рециркуляцию и полярный транспорт ауксина. Чин Научный Бык. 2008; 53: 2480–2487. [Академия Google]
57. Mancuso S, Marras AM, Mugnai S, Schlicht M, Zarsky V, Li G, et al. Фосфолипаза Dζ2 управляет везикулярной секрецией ауксина для его полярного межклеточного транспорта в переходной зоне верхушки корня. Поведение сигналов растений. 2007; 2: 240–244. [Бесплатная статья PMC] [PubMed] [Google Scholar]
58. Ezaki B, Kiyohara H, Matsumoto H, Nakashima S. Сверхэкспрессия ауксилиноподобного гена (F9E10.5) может подавлять поглощение Al корнями арабидопсиса.