
Содержание
Физиология растений. Часть 1 | Открытые видеолекции учебных курсов МГУ
Курс «Физиология растений. Часть 1» читается студентам третьего курса биологического факультета МГУ имени М. В. Ломоносова в 5 семестре.
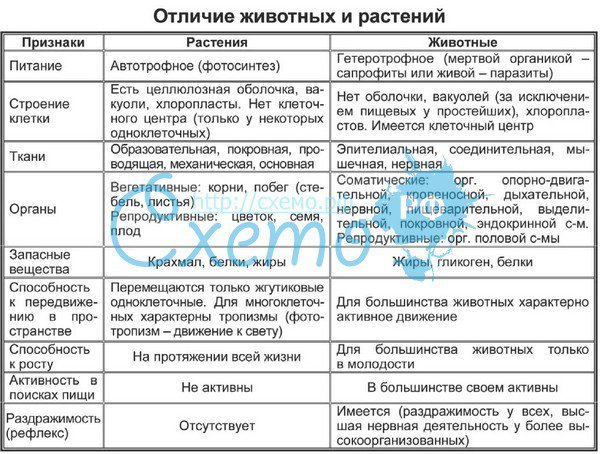
Физиология растений – наука об организации и координации функциональных систем зеленого растения. Физико-химический, экологический и эволюционный аспекты физиологии растений. Ее задача — познание закономерностей жизнедеятельности растений, раскрытие молекулярных основ сложных функций и механизмов их регуляции в системе целого организма.
Методологические основы фитофизиологии. Редукционизм, органицизм и интегратизм как подходы к изучению живых систем. Сочетание различных уровней исследования (субклеточный, клеточный, организменный, биоценотический) как необходимое условие прогресса физиологии растений. Специфические методы фитофизиологии как науки.
Задачи дисциплины углубленный анализ молекулярных механизмов различных функций интактного растительного организма (дыхание, фотосинтез, минеральное питание, водный обмен, рост и развитие, размножение, устойчивость к абиотическим и биотическим стрессам. ), их связь между собой, регуляция и приспособление к внешней среде, происхождение и становление в процессе эволюции и индивидуального развития.
), их связь между собой, регуляция и приспособление к внешней среде, происхождение и становление в процессе эволюции и индивидуального развития.
Цели дисциплины – изложение и закрепление знаний у студентов о физиологии растений как интегральной науки о жизнедеятельности интактного растительного организма – клеток, органов, функциональных систем.
Список всех тем лекций
Лекция 1. Физиология растений.
Методы и подходы
Происхождение растений, место в систематике.отличия высших растений от других организмов.
Дополнительная информация: экологические, политические, экономические и этические аспекты растений
Биоэнергетика
Энергетическая система клетки — дыхание
Процессы, происходящие в митохондриях
Лекция 2.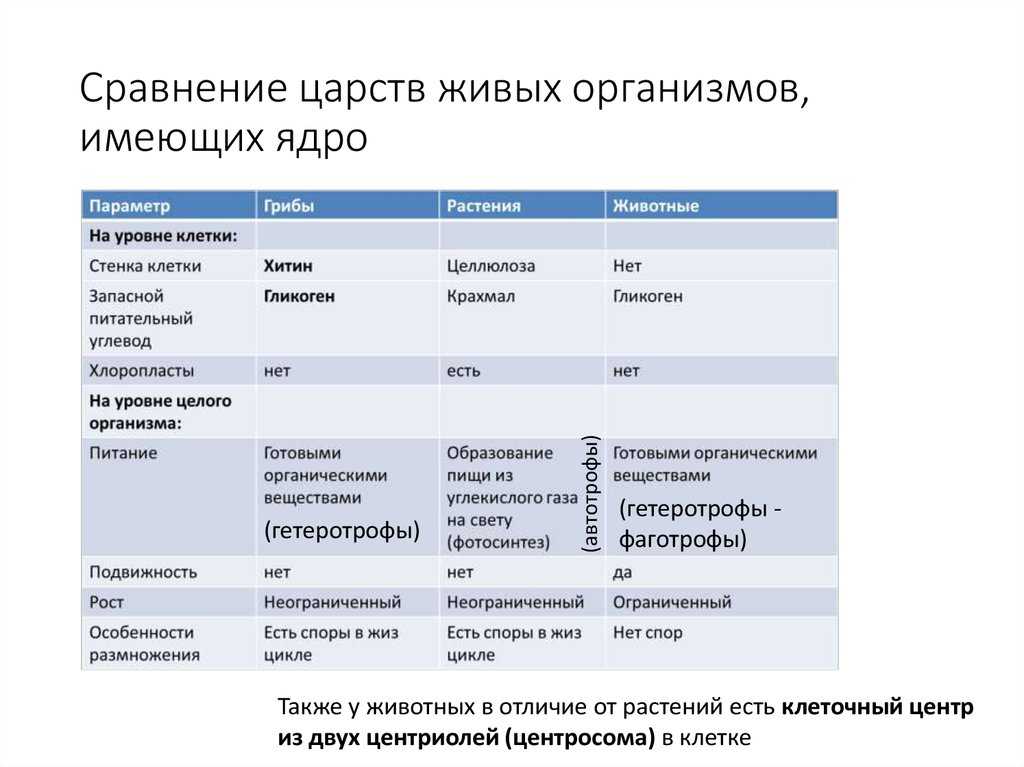 Физиология растений.
Физиология растений.
Биоэнергетика
Хиноны (убихиноны и пластохиноны)
Флавинадениндинуклеотид и флавинмононуклеотид
Железосерные белки
Гемы-коферменты цитохромов
Комплекс 1: никотинамидадениндинуклеотид-дегидрогеназный комплекс (НАДН-дегидрогеназный комплекс)
(НАДН-убихинон-оксидоредуктазы/НАДН-дегидрогеназы) митохондрий растений
комплекс) у растений
Схема работы
(Сукцинат дегидрогеназа)
(супрамолекулярные структуры, содержащие митохондриальные ферменты дыхательной цепи)
Особенности гликолиза у растений
Лекция 3. Физиология растений.
Специфика энергетических систем растительной клетки I
Два пути образования фруктоза-1,6-бисфосфатов у растений
Дифосфат-зависимая фосфофруктоза
Гликолиз в растительной клетке
Регулирование гликолиза у животных и растений
Специфика энергетических систем растительной клетки II
Общая схема цикла трикарбоновых кислот (цикла Кребса)
Транспорт веществ через мембрану митохондрий
Цикл трикарбоновых кислот в растительной митохондрии
Зачем нужна альтернативная оксидаза и как она включается
Транспорт интермедиатов дыхания через мембрану митохондрий
Электрон-транспортная цепь дыханий растений
Образование суперкомплекса
Система оксидаз растительной клетки
Лекция 4. Фотосинтез.
Фотосинтез.
Фотосинтез
Многообразие хлорофилов
Основные структурные особенности молекулы хлорофила
Биосинтез порфиринов у растений происходит в пластидах
Спектры поглощения хлорофиллов
Энергетические уровни хлорофилла
Первичные процессы фотосинтеза
Простейшие схемы фотосинтеза
Где взять источник электронов
Z-схема фотосинтеза
Кофакторы электрон-транспортной цепи фотосинтеза
Организация фотосинтетического аппарата
Фотосистема II (первый функциональный комплекс электрон-транспортной цепи хлоропластов)
Кинетика работы водоокисляющего комплекса
Фотосистема I (пластоцианин-ферредоксин-оксидоредуктаза): второй функциональный комплекс электрон-транспортной цепи хлоропластов
Лекция 5. Фотосинтез.
Фотосинтез.
Фотосинтез
Электрон-транспортная цепь хлоропластов
Варианты электронного транспорта в электрон-транспортной цепи хлоропластов
Антенны (светособирающие комплексы/антенные комплексы): пигмент-белковые комплексы фотосинтезирующих организмов
Антенны, фикобилисомы — светособирающий комплекс
Состав пигментов в антенных комплексах
Светособирающие комплексы различных организмов
Каротинойды: Каротины ксантофиллы
Развлечение генных инженеров
Схема мономер фотосистемы II с антенными комплексами
Светособирающие комплексы фотосистем I и II
Латеральная гетерогенность
(светособирающий комплекс 2)
Функции каротинойдов
Лекция 6. Фотосинтез.
Фотосинтез.
Механизм регулирования и защиты фотосистемы от фотодеструкции
Функции каротинойдов
Каротинойды в хлоропластах
Виолоксантиновый цикл
«Переключение» виолоксантиновой системы
Схема «Цикла Кальвина»
«Цикл Калвина»: Энергетика
фазы
Фаза кабоксилирования ВПЦ (восстановительного пентозофосфатного цикла)
Рибулёзобисфосфат-карбоксилаза оксигеназа (основной фермент листьев растений)
Решение проблем низкого углекислого газа: активация РуБисКо
Рубиско (РуБисКо — фермент, катализирующий присоединение углекислого газа к рибулёзо-1,5-бисфосфату)
РуБиско: катализирует две взаимоисключающие реакции
Решение проблем низкого углекислого газа
Восстановительная фаза Цикла Кальвина
Фаза регенерации: общая схема перегруппировок
бисфосфата
бисфосфата
Фаза регенерации: первая, вторая транскетолизная реакция
Фаза регенерации: образование рибулёзо-5-фосфата
Общий вид и регуляция
Крахмал в пластидах и сахароза в цитозоле
Гидролиз транзиторного крахмала в хлоропластах
Лекция 7. Фотосинтез, минеральное питание.
Фотосинтез, минеральное питание.
Фотосинтез
Схема к шизофреничному ферменту
Фотодыхание общая и подробная схемы
Соотношения карбоксилазной и оксигеназной функции Рубиско
Решение проблемы низкого СО2
Фотосинтез: Кранц-анатомия
С4-фотосинтез
Фосфоенолпируваткарбоксилаза
Регулирование фосфоенолпируваткарбоксилазы
Карбогидраза присутствует только в цитозоле
Участники игры
Три варианта С4-растений (растения, у которых третий этап фотосинтеза протекает с присоединением углекислого газа к фосфоенолпировиноградной кислоте)
Распределение ферментов С4-фотосинтеза
Три варианта С4-растений
и С4-растений в зависимости от ее концентрации
растений
САМ-метаболизм (Crassulacean Acid Metabolism): Временное карбоксилирование в фиксации углекислого газа
и С4-метаболизм: возможные варианты
Маленькая сенсация-одноклеточный С4-фотосинтез
Электронная микроскопия со схемой одноклеточного С4-фотосинтеза
Минеральный обмен растений
Минеральное питание
Восстановление нитратов
Нитратредуктаза: схема работы
Регуляция работы нитратредуктазы
Нитратредуктаза: строение
Лекция 8. Минеральный обмен растений.
Минеральный обмен растений.
Минеральный обмен растений
Азот
Использование азота
Флавоноид+белок NodD (активируют транскрипцию nod-генов, которые кодируют ферменты, обуславливающие формирование базовой структуры липо-хитонового олигосахарида)
Использование активности Nod (nucleotide-binding oligomerization domain)-генов-Nod-фактора (гены, кодирующие свойство клубеньковых бактерий образовывать клубеньки на корнях бобовых)
Структура нитрогеназы
В ядре клеток растений активируются гены, необходимые для симбиоза
Включение азота в органические соединения
Вторая система глутаматдегидрогенеза
«Добыча» азота растением
Транспорт азота
Включение азота в органические соединения при азотофиксации
Сера
Сера «работает» в разных степенях окисления
Метаболизм серы
Основные ферменты
Обмен серы в растении
Поглощение ионов корнем растений
Проницаемость биологических мембран
Варианты транспортных процессов через мембрану
Схема транспортных систем растительной клетки
Лекция 9. Минеральное питание, водный обмен растений.
Минеральное питание, водный обмен растений.
Варианты транспортных процессов через мембрану
Принципиальная схема транспортных процессов растительной клетки
Схема транспортных систем растительной клетки
Первичный транспорт
Три типа аденозинтрифосфата из растительной клетки
Структура белка Н-АТФазы (водородной аденозинтрифосфатазы) плазмалеммы
Н-АТФаза тонопласта
Образование электрогенного потенциала на плазмалемме и тонопласте
АВС-транспортёры (транспортные белки, использующие энергию гидролиза аденозинтрифосфатазы для транспорта через мембраны самых разных химических агентов)
АВС-транспортёры: строение работа
Вторичный транспорт
Две стратегии поглощения железа растениями
Железо восстанавливается и переносится через мембрану специфическим транспортёром
Стратегия 2: обеспечивает поглощение трехвалентного железа за счет образования специальных хелатирующих соединений фитосидерофор)
Пассивный транспорт
Системы транспорта калия в растительных клетках
Функции калия
Водный обмен растений
Водичка в растительной клетке
Осмотические явления
Лекция 10. Водный обмен растений.
Водный обмен растений.
Водный обмен растений
Осмотические явления
Растительная клетка как осмотическая система
Термодинамические показатели воды
Водный баланс клетки
Осмотический стресс
Направление воды в системе целого растения
Колебания водного потенциала в растительных тканях
Динамические характеристики потока воды в клетку
Аквапорины
Структура водных каналов аквапоринов
Капиллярные эффекты
Поглощение воды корнем
Устьица — особые клетки эпидермиса
Гуттация
Лекция 11. Водный обмен растений, растительная клетка.
Водный обмен растений, растительная клетка.
Водный обмен растений
Нижний концевой двигатель
Радиальное строение и механизм возникновения корневого давления
Радиальный транспорт воды в клетке
Механизм работы верхнего концевого двигателя
Схема флоэмного транспорта
Строение флоэмы
Состав флоэмного и ксилемного эксеудата
Вещества, не транспортируемые (А) и транспортируемые во флоэме (В)
Клетка: спутницы бывают трех типов
Симпластная и апопластная загрузка флоэмы
Симпластическая загрузка флоэмы
Взаимность ксилемного и флоэмного транспорта
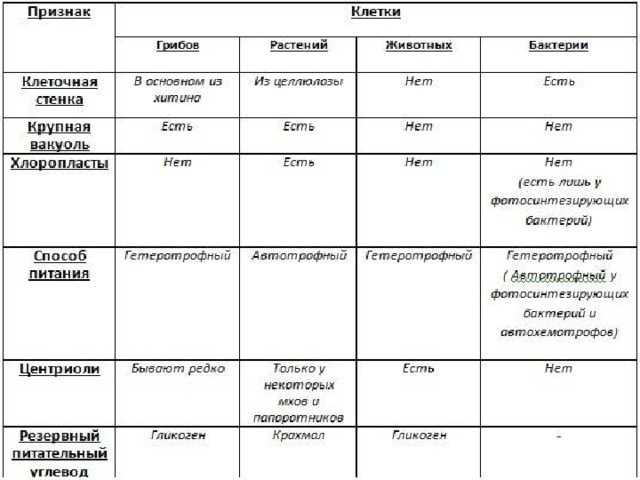
Растительная клетка
Органеллы эукариотической клетки
Классификация органелл
Два пути сортировки белков: цитоплазматический и секреторный
Сигнальные последовательности транспорта белков в растительной клетке
Транспорт ядерно-кодируемых белков в хлоропласт
Секреторный путь транспорта белков
Аппарат Гольджи
Цитология растительной клетки
Клеточная стенка
Лекция 12.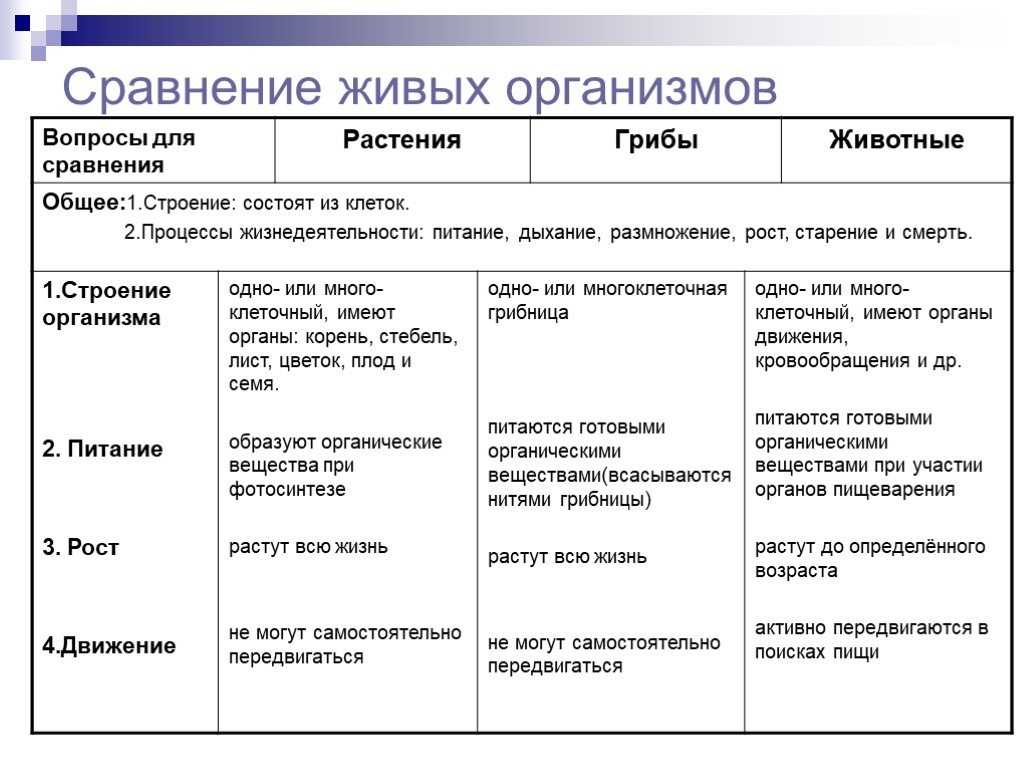 Клеточная стенка, плазмалеммы.
Клеточная стенка, плазмалеммы.
Клеточная стенка
сахаров
Строение микрофибрилл целлюлозы
Строение целлюлозо-синтазы
Сшивочные гликаны
Гемицеллюлозы: ксилоглюкан двудольных
Пектины
«Замковые зоны» пектиновой сети
Пектины: функциональная сеть клеточной стенки
Структурные белки клеточной стенки
Трехмерная модель двух типов клеточной стенки
Лигнины
Образование лигнина
Плазмалеммы
Лекция 13. Клеточная стенка, плазмалеммы.
Плазмалеммы
Некоторые особенности плазмалеммы
Функции плазмолеммы
Структура растительного аппарата Гольджи
Везикулярный транспорт, типы молекул
Вакуоли
Транспорт белков в вакуоли: варианты и сигналы
Пероксисомы
Разновидность пероксисом
Структура ядерных пор
Некоторые особенности ядерного генома растений
Лекция 14.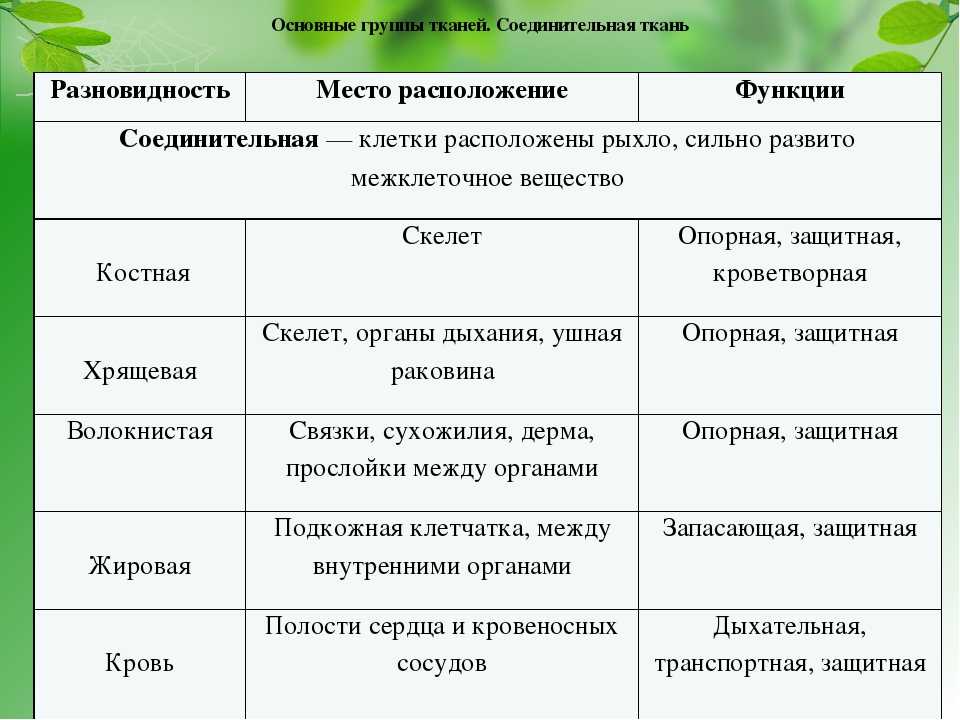 Физиология клетки.
Физиология клетки.
Некоторые особенности ядерного генома растений
Взаимопревращение пластид контролируется ядерным геномом
Фитоферритин в пластидах мезотила сои, аминопласт
Этиопласт, формирование хлоропласта
Структура хлоропластного генома риса
Структура хлоропластного генома разных видов растений
Сходства и отличия хлоропластного генома и белоксинтезирующей системы от бактериальных
Гены хлоропластов
Особенности работы пластидной системы РНК-полимеразы
Функции пластид
Растительные митохондрии: строение митохондрий
Структура митохондрий кукурузы
Сопоставление хлоропластного и митохондриального геномов риса
Гены митохондрий
Варианты редатбирования хлоропластных митохондриальных РНК растений
Метаболизм растительной клетки
Клеточный цикл и его регуляция
Культура клеток высших растений
Культура клеток как биологическая система
Каллусогенез и дифференцировка культуры клеток
Характеристика каллусных культур
Получение суспензионных культур клеток
Система выращивания культур клеток высших растений
Лекция 15.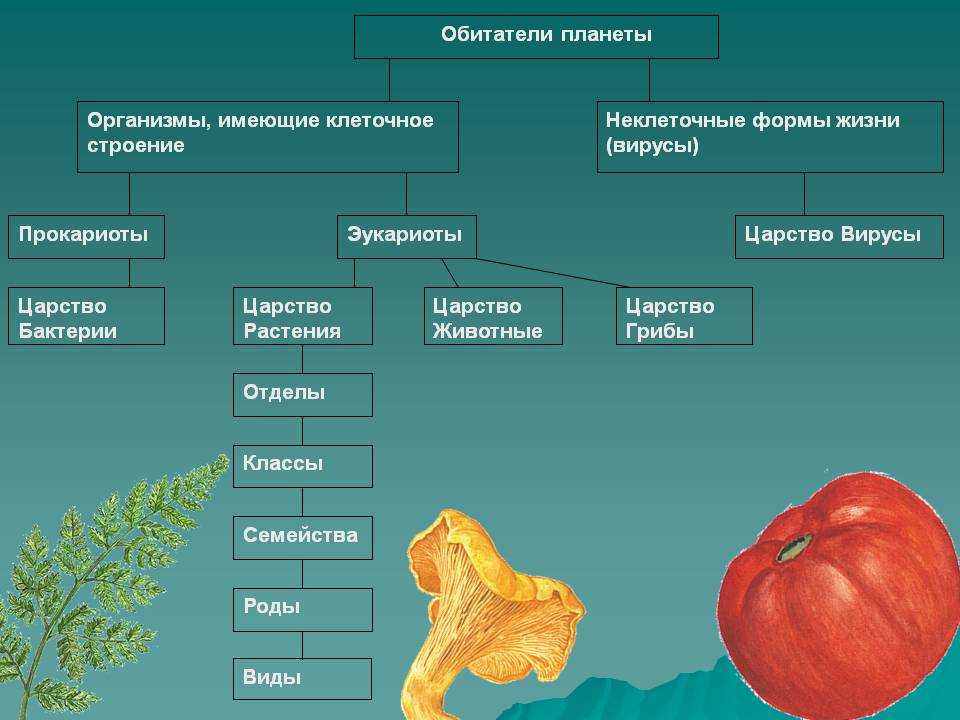 Культура клеток высших растений.
Культура клеток высших растений.
Три ипостаси культуры клеток высших растений
Основные свойства культуры клеток высших растений
Гетерогенность популяций культур клеток высших растений
Генетическая гетерогенность культур клеток
Перестройка хромосом в культуре клеток
Отбор клеток по максимальной или устойчивой пролиферации
Параметры клеточного цикла и митотический индекс культур клеток
Изменение плоидности клеток при выращивании культуры клеток диоскореи в проточном режиме
Протопласты
Фотосинтез
Минеральное питание
Использование элементов питания: азот
Культивирование клеток диоскореи — влияние фосфата
Лекция 16. Культура клеток высших растений.
Культура клеток высших растений.
Культура клеток высших растений
Протопласты
Основные свойства культуры клеток высших растений
Морфогенез диполярный — геммогенез, ризогенез
Сомаклональная вариабельность: механизмы и закономерности
Рост и развитие
Соматический эмбриогенез: этапы и факторы
Практическое использование культуры клеток высших растений
Коллекция растительных объектов
Коллекция «пробирочных» растений
Коллекция тканей и органов растений
Депозитарий живых систем МГУ
Сельскохозяйственная биотехнология
Клеточная селекция: схема клеточной селекции
Рынок лекарственных препаратов
Рост и сохранность ягнят в молочный период при использовании ферментно-пробиотического препарата «Бацелл»
А. Н.Ульянов, доктор с.-х. наук,
Н.Ульянов, доктор с.-х. наук,
А.Я.Куликова, доктор с.-х. наук,
Н.П.Журавлева, главный зоотехник
В настоящее время в животноводстве уделяется большое внимание повышению эффективности использования кормов, повышению энергетической ценности рационов за счет применения ферментных препаратов и других биологических веществ. Нами изучено влияние скармливания ягнятам мясошерстного направления продуктивности ферментно-пробиотического препарата «Бацелл». С этой целью были взяты две группы ягнят в возрасте 24 (контрольная) и 25 (опытная) дней. В дополнение к молоку матерей, ягнята обеих групп получали сено суданки и комбикорм с содержанием 14,4% сырого протеина. Ягнятам опытной группы вместе с комбикормом отдельно скармливали по 10 грамм пробиотика «Бацелл».
Ягнята контрольной (1) и опытной (2) групп по постановочной живой массе практически не различались (табл.1).
Таблица 1 — Живая масса ягнят, кг
| Группа ягнят | Пол ягнят | При постановке (25 дней) | В возрасте 59 дней | В возрасте 90 дней | |||
|---|---|---|---|---|---|---|---|
| n | M±m | n | M±m | n | M±m | ||
| 1 | Бараны | 14 | 6,71±0,28 | 14 | 9,98±0,49 | 13 | 13,93±0,62 |
| Ярки | 21 | 6,03±0,24 | 21 | 10,84±0,36 | 18 | 14,37±0,71 | |
| Среднее | 35 | 6,30±0,18 | 35 | 10,49±0,29 | 31 | 14,18±0,48 | |
| 2 | Бараны | 15 | 7,00±0,32 | 15 | 13,28±0,64 | 15 | 16,14±0,72 |
| Ярки | 20 | 6,15±0,34 | 20 | 12,45±0,499 | 18 | 16,61±0,76 | |
| Среднее | 35 | 6,55±0,24 | 35 | 12,81±0,39 | 33 | 16,38±0,52 | |
В возрасте 59 дней средняя живая масса ягнят опытной группы оказалась достоверно (td=4,77) выше, чем в контроле.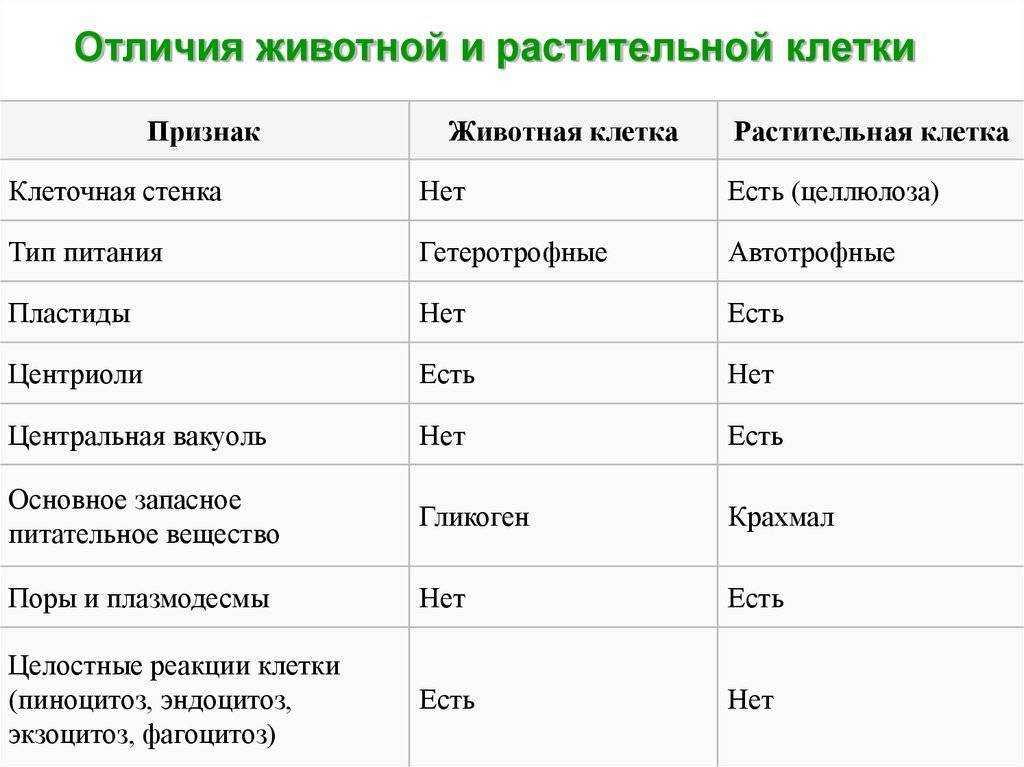
За 25-дневный период прирост живой массы ягнят контрольной группы составил 4,19 кг, в том числе у баранчиков — 4,13 кг, у ярок — 4,81 кг, а у ягнят опытной группы, получавших дополнительно «Бацелл», эти показатели составили 6,26, 7,13 и 6,30 кг соответственно, что на 49,4; 73,4 и 30,9% выше нежели у контрольных сверстников.
Среднесуточный прирост живой массы за этот же период у ягнят опытной группы составил 250,4, в том числе у баранов — 258,2 г, у ярок — 252 г, в то время как у контрольных сверстников 167,6, 165,2 и 192,4 г соответственно.
За возрастной период с 59 до 90 дней у ягнят контрольной группы живая масса возросла в среднем на 35,18%, в том числе у баранов — на 39,58%, у ярок — на 32,56%, а в опытной группе на 27,8%, 21,54 и 33,41% соответственно. Прирост живой массы за этот период у всех ягнят контрольной группы составил 3,69 кг, или 82,0 г/сут., а ягнят, получавших «Бацелл», 3,57 кг, или 79,3 г/сут.
Различия в интенсивности роста между ягнятами контрольной и опытной групп в этот период связанны с низким качеством суданкового сена, (содержавшем 38,0% клетчатки) и отъемом ягнят от их матерей в возрасте 65 дней.
В целом, за 65 дней опытного периода у ягнят контрольной группы прирост живой массы составил 7,88 кг (табл.2), в том числе у баранов — 7,22 кг, у ярок — 8,15 кг, или по 121,2; 111,1 и 125,4 г/сут.
Таблица 2 — Общий (кг) и среднесуточный (г) прирост живой массы ягнят за учетные периоды опыта (на голову)
| Группа ягнят | Пол ягнят | Прирост живой массы (кг) за периоды опыта, дней | Прирост, г/сут. | ||||
|---|---|---|---|---|---|---|---|
| С 25 до 59 | С 60 до 90 | С 25 до 90 | С 25 до 59 | С 60 до 90 | С 25 до 90 | ||
| 1 | Бараны | 4,13 | 3,95 | 7,22 | 165,2 | 87,8 | 110,1 |
| Ярки | 4,81 | 3,53 | 8,15 | 192,4 | 78,4 | 125,4 | |
| Среднее | 4,19 | 3,69 | 7,88 | 167,6 | 82,0 | 121,4 | |
| 2 | Бараны | 6,28 | 2,86 | 9,14 | 251,2 | 63,6 | 140,6 |
| Ярки | 6,40 | 4,16 | 10,46 | 252,2 | 92,4 | 160,9 | |
| Среднее | 6,26 | 3,57 | 9,83 | 250,4 | 79,3 | 151,2 | |
Прирост живой массы ягнят опытной группы за этот же период составил 9,83, 9,14 и 10,46кг, или 151,2, 140,6 и 160,9 г/сут. Превосходство ягнят опытной группы над контрольной по приросту живой массы составило 24,7, 26,6 и 28,3%.
Превосходство ягнят опытной группы над контрольной по приросту живой массы составило 24,7, 26,6 и 28,3%.
Для ягнят, получавших «Бацелл», характерно лучшее не только общее развитие, но и сохранность за опытный период. В числе выращенных ягнят с живой массой 12 кг и менее в этой группе имелся 1 ягненок, что составляет 2,85% к общему количеству, в то время как среди ягнят контрольной группы, отставших в развитии, было 5 голов, или 16,13%.
К отъему от матерей отход ягнят в контрольной группе составил 4 головы или 11,43% к их наличию на начало опыта, а в опытной группе таких ягнят не было.
В целях определения эффективности использования кормов проводился учет их поедаемости по результатам проведенных контрольных кормлений.
В результате отмечен низкий уровень поедаемости сена суданки, представленного крупностебельчатой, малооблиственной его формой (табл.3). В контрольной группе суточный рацион состоял из 451 г сена суданки и 207 г комбикорма. Поедаемость сена составила 60,5%, а комбикорма — 100%. В опытной группе эти показатели составили 59,7 и 100%.
В опытной группе эти показатели составили 59,7 и 100%.
Таблица 3 — Затраты корма и его оплата приростом живой массы (на 1 ягненка)
| Группа ягнят | Учетных дней опыта | Затрачено кормов | Получено прироста живой массы, кг | Затрачено на 1 кг прироста живой массы | ||
|---|---|---|---|---|---|---|
| Корм.ед., кг | Переваримого протеина, кг | Корм.ед., кг | Переваримого протеина, кг | |||
| 1 | 65 | 28,17 | 3,17 | 7,88 | 3,57 | 0,402 |
| 2 | 65 | 29,54 | 3,13 | 9,80 | 3,01 | 0,319 |
Использование в рационе «Бацелл» способствовало улучшению оплаты корма приростом живой массы. Так на 1 кг прироста живой массы ягнятами опытной группы было затрачено на 18,6% меньше кормовых единиц и на 26,0% меньше переваримого протеина (табл.3), нежели их сверстниками контрольной группы.
Результаты выполненных исследований дают основание считать, что использование ферментно — пробиотического препарата «Бацелл» при выращивании ягнят в молочный период (в первые 60-65 дней) обеспечивает повышение прироста живой массы и их сохранность.
Овцы, козы, шерстяное дело, № 4, 2008г.
Рост растений — закономерности, ограничения и модели — Неживая жизнь
Рис. 1 Проращивание сеянцев ели ситхинской. Почти весь рост до этого момента был обеспечен питательными веществами, хранящимися в семени. Когда появятся первые листья, они смогут фотосинтезировать и обеспечить материальные и энергетические потребности роста.
Одним из чудес жизни является рост, способность организмов становиться больше. Это особенно относится к растениям, поскольку, как описано ранее, растения растут «из воздуха». части окружающей их среды и собрать их в органическую сущность. Напротив, животным нужно только найти предварительно собранные фрагменты жизни, возможно, все еще живые, а возможно, и нет, и переконфигурировать их для собственного использования. И хотя контроль за ростом животных прост и связан с приемом пищи, то, что контролирует рост растений, менее очевидно.
И хотя контроль за ростом животных прост и связан с приемом пищи, то, что контролирует рост растений, менее очевидно.
Когда растения растут, они обычно увеличиваются как в размере, так и в весе, и процесс роста может быть связан с увеличением того и другого. Однако по ряду причин рост растений определяется как необратимое увеличение размеров, а не увеличение массы. Частично это связано с тем, что поглощение и потеря воды могут существенно изменить (влажную) массу растения из-за процессов, которые большинство не рассматривает в качестве роста. Например, за ночь деревья набирают значительное количество воды, чтобы восполнить потери, потерянные в течение дня; большинство не считает прибавку в весе за ночь ростом, а потерю веса в течение дня — «отрицательным ростом». , азот, фосфор и т. д., а также синтез органических молекул, таких как углеводы и белки. Но определение роста по увеличению сухого веса привело бы к некоторым противоречивым результатам. Большинство считает, что деревья растут весной, когда удлиняются побеги и появляются листья. В это время дерево фактически уменьшается в сухом весе. Летом в районах с умеренным климатом, когда деревья фотосинтезируют и поглощают питательные вещества, их сухая масса увеличивается, но многие из них не становятся больше с точки зрения более длинных побегов. Точно так же прорастающее семя, которое большинство сочло бы растущим, на самом деле уменьшается в сухом весе до тех пор, пока скорость его фотосинтеза не превысит скорость дыхания; обычно этого не происходит, пока сеянцу не исполнится пару недель, и он уже будет значительного размера. Следовательно, рост растений обычно определяется как необратимое увеличение размеров.
В это время дерево фактически уменьшается в сухом весе. Летом в районах с умеренным климатом, когда деревья фотосинтезируют и поглощают питательные вещества, их сухая масса увеличивается, но многие из них не становятся больше с точки зрения более длинных побегов. Точно так же прорастающее семя, которое большинство сочло бы растущим, на самом деле уменьшается в сухом весе до тех пор, пока скорость его фотосинтеза не превысит скорость дыхания; обычно этого не происходит, пока сеянцу не исполнится пару недель, и он уже будет значительного размера. Следовательно, рост растений обычно определяется как необратимое увеличение размеров.
- Процессы роста
- Ограничения роста растений
- Ростовые модели
Процессы роста
Увеличение размеров растений происходит по мере увеличения отдельных клеток, образующихся в результате клеточных делений в меристемах. Хотя на первый взгляд это может показаться простым явлением, рассмотрите следующие аспекты этого процесса:
- Все растительные клетки, даже маленькие, окружены ограничивающей клеточной стенкой, основной функцией которой является
- .
 0002 предотвращает расширение . Это позволяет клетке создавать давление, и это важно для водного баланса и функционирования клеток растений.
0002 предотвращает расширение . Это позволяет клетке создавать давление, и это важно для водного баланса и функционирования клеток растений. - По мере расширения клетки толщина клеточной мембраны и клеточной стенки снаружи , а не уменьшается. В отличие от этого, рассмотрим расширяющийся воздушный шар: по мере того, как вы вдуваете в него воздух, «кожа» воздушного шара растягивается все тоньше и тоньше, поскольку статический объем материала воздушного шара распределяется по все большему и большему объему. Этого не происходит по мере роста растительных клеток, следовательно, расширение должно быть согласовано с производством нового материала для клеточной мембраны и клеточной стенки; это сохраняет толщину границ ячейки постоянной. Значение этого, особенно в отношении мембраны, должно быть очевидным; растяжение плазматической мембраны невозможно, только ее разрыв, и это разрушило бы ее способность быть селективным барьером, удерживающим одни молекулы внутри, а другие вне клетки.
- Хотя расширение клетки является следствием поглощения воды, это не простое «разбавление» клетки. Подобно тому, что происходит в мембране и стенке, клетка добавляет внутриклеточные компоненты со скоростью, соответствующей ее расширению.
- Хотя цитозоль действительно увеличивает свой объем по мере роста клетки, обычно именно расширение вакуолей составляет большую часть увеличенного объема клетки. Предполагая, что более крупная клетка полезна для организма (поскольку она позволяет ему проникать в большую часть окружающей среды, что важно как для корней, так и для побегов), крупная центральная вакуоль является относительно «дешевым» способом увеличения клетки, поскольку ее содержимое Для получения вакуоли требуется меньше энергии, чем содержимое цитозоля. Увеличенная клетка — это не просто разбавленная и растянутая версия исходной, ее стенки и мембраны имеют ту же толщину, что и раньше, а цитозоль — того же состава, что и раньше. Цитозоль несколько увеличился в объеме, но большая часть увеличения общего объема клетки является результатом более крупной вакуоли, которая должна иметь ту же концентрацию растворенного вещества, что и цитозоль, но растворенные вещества другие, и «более дешевые» находятся в вакуоли. .
- Область произрастания растения отделена, часто на очень значительные расстояния, от источника материалов для этого произрастания. Какие материалы нужны для роста? Мы можем выделить три основные потребности: вода , которая представляет собой самый большой компонент «нового растительного» материала; углеводы , которые используются как в клеточном дыхании, для обеспечения энергией синтетических реакций, так и в качестве строительных материалов для изготовления клеточных стенок, клеточных мембран, внутренних мембран, белков, метаболитов, витаминов и т. д.; и минеральные питательные вещества , напр. фосфор для мембран, азот для аминокислот и т. д. Вода и питательные вещества поступают из почвы и, таким образом, находятся очень близко к растущим клеткам корневых меристем, но должны переноситься на значительные расстояния, чтобы добраться до кончиков побегов, до 350 футов. в случае красного дерева. Углеводы поставляются фотосинтезирующими листьями, которые могут располагаться относительно близко к растущим побегам, но могут находиться на расстоянии 100 м от растущей корневой меристемы. Углеводы часто не поступают напрямую из листьев в точки роста, а вместо этого могут поступать из листьев в места хранения, а затем из мест хранения в области роста.
Клеточному расширению противодействует прочность клеточной стенки, и расширение происходит, когда давление внутри клетки, создаваемое внутренней диффузией воды, превышает прочность стенки. Растущие клетки имеют «более мягкие» клеточные стенки, т. е. стенки, которые поддаются (расширяются) при более низком давлении, чем нерастущие клетки. И считается, что растения контролируют клеточный рост, контролируя «размягчение» клеточных стенок. Подводя итог, можно сказать, что рост растений включает скоординированный процесс синтеза мембран (как клеточной, так и вакуолярной), (2) клеточной стенки, (3) цитоплазматических материалов (белков, мембран, метаболитов) и (4) некоторых вакуолярных клеток. материалов, наряду с поглощением воды и минералов. Размер клетки определяется генетикой и условиями окружающей среды. Рост прекращается, когда стенка «затвердевает» и больше не поддается давлению, создаваемому диффузией воды.
В тех клетках, у которых есть вторичные клеточные стенки, дополнительный материал клеточной стенки откладывается после прекращения расширения клетки — обратите внимание, что, пока добавляются материалы вторичной клеточной стенки, клетка не увеличивается в размере, а увеличивается в массе.Большая часть роста растений является результатом расширения вакуолей. Это следует за клеточным делением, в результате которого образуются новые клетки, примерно такие же, как исходные, что приводит к так называемому цитоплазматическому росту и лишь незначительному увеличению размера органа. Деление клеток имеет важное значение для процесса роста, потому что оно обеспечивает клетки, которые могут демонстрировать вакуолярный рост. Деление клеток происходит в растениях в изолированных местах, называемых меристемами, и большая часть роста растений происходит в областях, прилегающих к этим меристемам, где новообразованные клетки демонстрируют вакуолярный рост. Расширение новообразованных клеток отодвигает меристематические области дальше от основного тела растения, увеличивая общий размер растения.
Таким образом, существует пространственное разделение между областью клеточного деления и областью клеточного роста (глава 8). Этот паттерн наиболее заметен при апикальном росте, но также имеет место и при вторичном (латеральном) росте (глава 9).), хотя и в сосудистом камбии, и в пробковом камбии рост клеток происходит с обеих сторон меристемы. Активность обычно возникает одновременно в области клеточного деления и в области клеточного роста. Наибольший рост органа происходит всякий раз, когда новым клеткам, продуцируемым меристемой, позволяют расширяться. Для некоторых растений рост более или менее непрерывен и стабилен, пока условия окружающей среды (в частности, температура, вода и свет) стабильны. Тем не менее, большинство растений демонстрируют эпизодический рост со вспышками образования и расширения клеток, за которыми следуют периоды бездействия, даже при постоянных и благоприятных условиях. Для многих многолетних растений, обитающих в районах с сезонным климатом, рост строго сезонный и происходит только часть года. Часто период роста составляет лишь очень небольшую часть «благоприятного» периода времени. Например, многие деревья в северных районах с умеренным климатом растут только в течение двух-четырех недель в мае.Паттерны, описанные выше, представляют собой экспансионный рост, проявляемый апикальными меристемами в корнях и побегах (первичный рост), и экспансионный рост, вызываемый боковыми меристемами (вторичный рост). Рост листьев и плодов разный; эти детерминированные органы имеют паттерн, сходный с таковым у многих животных, у которых весь орган имеет период клеточного деления, за которым следует период клеточной экспансии, т. е. клеточное деление и экспансия разделены во времени, а не в пространстве. Может быть период перекрытия, когда происходят как деление, так и экспансия, но обычно задолго до прекращения роста клетки перестают делиться, и эмбриональные клетки больше не образуются. Рост поддерживается по мере расширения новых клеток. В конце концов рост органа прекращается и остается структура неизменного размера.
Однако эти структуры do продолжают развиваться: проявляют старение и опадение в случае листьев и в случае плодов, демонстрирующих созревание таким образом, что это способствует распространению семян (и это часто также включает опадение).Два примера роста:
Почки деревьев Почки деревьев умеренного пояса формируются во второй половине лета. Почка содержит апикальную меристему, очень короткий участок стебля с несколькими примордиальными листьями, упакованными особыми листьями (чешуйками почек), которые покрывают всю структуру. Почки увеличиваются в размерах с момента их образования до того момента, когда они становятся бездействующими ранней осенью, но увеличение размера незначительно, и обычно образуются бутоны размером от 0,5 до 3 см. Весной, при «распускании почек», содержимое почки резко увеличивается в размерах, образуя участок стебля длиной до 20 см с несколькими листьями сопоставимой длины. Как и в случае с плодами, большинство клеток, присутствующих в этом новом участке стебля, образовались ранее, и большая часть роста, наблюдаемого весной, является результатом расширения этих клеток за счет вакуолярного роста.
Рис. 2 Рост веток — рост веток неопределенный, но происходит ежегодными «приливами». Рост листьев детерминированный. Вся структура справа от стрелки присутствовала в шишке, полученной в конце прошлого лета. И большинство клеток как ветки, так и листьев присутствовали в зародыше в течение девяти месяцев, прежде чем быстро разрослись в течение 2-3 недель в мае.
Плоды Типичный плод яблони начинает свое развитие после оплодотворения, обычно в конце апреля. Месяц спустя плод вырос до лишь небольшой части своего окончательного размера, с диаметром 1-2 см и объемом около 5 кубических см, однако все клетки плода (примерно 75 миллионов (!) клетки) присутствуют. В течение лета яблоко вырастает до своего зрелого размера, возможно, 10 см с объемом 1300 куб. В течение этого времени новые ячейки не добавляются; происходит только рост уже существующих клеток. У яблок и многих других фруктов конечный размер плода определяется количеством образующихся клеток и, таким образом, зависит от условий поздней весной, а не от летних.
Для поддержания роста необходим запас материалов, не только воды, обеспечивающей клеточное расширение, но и материалов для создания новых клеточных стенок, клеточных мембран и всех компонентов цитоплазмы по мере расширения клетки. Материалы нужны не только для создания увеличивающихся клеток, но и для снабжения энергией, необходимой для этих процессов. Каждая пептидная связь требует гидролиза АТФ, как и каждая дополнительная единица глюкозы в растущем полимере целлюлозы. Помимо прямых «затрат на строительство», энергия необходима для других клеточных процессов, например, для транспорта молекул через мембраны.
Ограничения роста
Многие люди заинтересованы в том, чтобы растения росли быстрее, производя больше материала за более короткий период времени. Что ограничивает рост? Ниже перечислены некоторые важные факторы, некоторые из которых действуют по-разному. Хотя увеличение всех этих факторов может увеличить рост, эта реакция непостоянна и часто ослабевает при дальнейшем увеличении фактора, что приводит к феномену насыщения , когда дальнейшее увеличение фактора вызывает незначительные изменения в росте.
Более того, для всех этих факторов может быть «слишком много хорошего», и дальнейшее увеличение фактически снижает рост (токсичность).Рис. 3 Изменения роста в ответ на увеличение количества питательного вещества. Когда питательное вещество находится на низком уровне, оно влияет на рост последовательным, часто линейным образом. Но с дальнейшим увеличением количества питательных веществ увеличение роста уменьшается, т. е. происходит насыщение, и в конечном итоге рост больше не увеличивается по мере добавления большего количества питательных веществ. Кроме того, большинство питательных веществ токсичны при достаточно высоком уровне, т. е. рост замедляется по мере поступления большего количества питательных веществ. Рис. 4 Типичное удобрение содержит азот, калий и фосфор. Это по 13% от каждого в заказе; если бы было указано 20-10-8, это было бы 20% азота, 10% калия и 8% фосфора.
Питательные вещества
Все необходимые минеральные элементы могут ограничивать рост.
Ограничение может возникать как из-за отсутствия этого элемента в почве, так и из-за того, что элемент хотя и присутствует, но недоступен из-за почвенных условий. Например, железо часто недоступно в основных почвах, хотя оно может присутствовать в изобилии. Проблема в том, что в аэробных, основных условиях очень мало железа присутствует в форме, которая легко растворяется.Где-то на земле есть почвы с дефицитом всех 14 минеральных элементов, необходимых растениям, и дефицит может развиться даже в таких элементах, как молибден, которые необходимы в очень малых количествах. В начале 1920-го века Карл Шпренгель разработал идею, позже отстаиваемую Юстусом ван Либихом, под названием «Закон минимума»: рост растений будет ограничен не доступностью питательных веществ в целом, а любым питательным веществом, которое находится в наименьшем количестве относительно того, сколько необходимо. Например, хотя добавление азота часто увеличивает рост растений, если доступного молибдена недостаточно, такие добавки не приведут к какому-либо усилению роста.
Выращивание сельскохозяйственных культур можно сравнить с выпечкой торта: если рецепт торта требует пяти ингредиентов, приготовление торта может быть ограничено любым из пяти ингредиентов, а недостаток одного не компенсируется избытком других. Это очень простая идея, применимая во многих ситуациях. Но это противоречит общепринятой идее о том, что реакция на факторы всегда будет постоянной: «если немного хорошо, то многое должно быть лучше».0002 не случай!В то время как слишком мало основных питательных веществ может ограничить рост, слишком много тех же элементов (токсичности) также может замедлить рост. Наиболее распространенная токсичность является результатом засоленных почв с высоким уровнем K, Ca, Cl, SO 4 и Na, но уникальные почвенные условия (заболачивание) также могут вызывать токсичность железа и марганца в незасоленных почвах.
Вода
Рис. 5 Круговое орошение на хлопковом поле. Система вращается вокруг центрального источника воды.
Вода является наиболее важным фактором, ограничивающим наземный фотосинтез во всем мире. Вода играет несколько ролей в росте растений: как реагент в фотосинтезе, как основной компонент любой новой клетки, которая образуется, как транспортная среда, которая перемещает материалы по растению и, в частности, в растущие регионы. Хотя все вышеперечисленное потенциально может играть определенную роль, эффект воды возникает в первую очередь из-за взаимодействия между потерей воды и накоплением углекислого газа. В засушливых местообитаниях растения держат устьица закрытыми, чтобы избежать потери воды. Это снижает концентрацию углекислого газа внутри листа и уменьшает количество фотосинтеза. Кроме того, растения могут уменьшить потерю воды за счет более мелких листьев или меньшего их количества, что может ограничивать рост, поскольку общая площадь листа определяет количество фотосинтеза, которое может происходить. В то время как нехватка воды может замедлить рост, избыток воды также вреден для большинства растений, прежде всего потому, что переувлажненные почвы становятся анаэробными, а корни плохо растут и/или отмирают.
Рис. 6. Вид со спутника на пахотные земли Канзаса. Круги отражают использование центральных круговых оросительных систем, в которых большая штанга вращается вокруг источника воды. Стрела моторизована для кругового движения; вода, подаваемая из колодца, откачивается из бонового заграждения и используется для полива сельскохозяйственных культур.
Свет
Без света невозможен фотосинтез, а без фотосинтеза невозможен рост. Свет может оказывать очень значительное влияние на фотосинтез и рост, но только тогда, когда другие условия благоприятны для поддержания роста, и только при уровне освещенности, сравнимом с тем, который обычно испытывает растение. Слишком много света может быть чрезвычайно вредным по ряду причин, и растения, адаптированные к тени, обычно очень плохо себя чувствуют при воздействии яркого света. Для домашнего садовода правильное расположение декоративных растений во многом зависит от соображений освещения. Количество света, которое получает растение, контролируется как интенсивностью света, так и продолжительностью светового воздействия; эффекты не всегда компенсируют друг друга, т.
Е. Короткие периоды очень яркого света не эквивалентны более длительным периодам менее яркого света. Большинство видов сельскохозяйственных культур адаптированы к условиям высокой освещенности и очень плохо себя чувствуют при выращивании в затененных условиях, и, вероятно, рост урожая может быть снижен в результате продолжительных облачных условий. Однако такие условия часто связаны с частыми дождями, которые также могут быть причиной замедления роста в случае затопления почв.Распределение листьев и продолжительность жизни являются важными параметрами растений, на которые влияет свет из-за проблем, связанных с самозатенением. Как правило, листья образуются таким образом, чтобы уменьшить самозатенение и обеспечить больший фотосинтез. Старые листья, находящиеся в затененных условиях, часто выбрасываются (т. е. они стареют и опадают), потому что они больше не получают достаточно света, чтобы приносить пользу в энергетическом/материальном смысле. Это отражено в следующем уравнении:
Чистый фотосинтез = общий фотосинтез – дыхание
Предполагая, что стоимость обслуживания листа (т.
е. количество дыхания, необходимое листу для поддержания его жизненных условий) постоянна, затенение снизит общий фотосинтез до такой степени, что чистый фотосинтез отрицательный, т. е. содержание листа обходится дороже, чем он «зарабатывает» в процессе фотосинтеза. В этот момент растение может сократить свои потери, удалив лист.Температура
Растения пойкилотермные, их температура не регулируется внутренне, а определяется условиями окружающей среды. Более того, у растений есть диапазон температур, в которых они могут выжить (часто этот диапазон толерантности меняется в зависимости от сезона). На все биологические процессы, и в особенности на фотосинтез, дыхание и рост, влияет температура (глава 26), и, за несколькими важными исключениями, основная реакция заключается в том, что активность растений, включая рост, увеличивается при более высоких температурах в диапазоне от 0 до 20°С (от 32 до 68°F). Однако у всех растений есть оптимальная температура для роста, выше которой рост замедляется с повышением температуры.
Частично это объясняется тем, что при более высоких температурах дыхание более чувствительно к температуре (т. е. больше увеличивается с повышением температуры), чем фотосинтез. температура (см. уравнение выше).Взаимодействие между водой, светом и температурой
Эти факторы часто переплетаются: больше света повышает температуру, а более высокие температуры усиливают транспирацию и могут привести к осложнениям из-за нехватки воды. Насколько тесно связаны эти три фактора, зависит от множества факторов.
Площадь листьев
Как и следовало ожидать, растения с большей площадью листьев обычно растут быстрее, чем растения с меньшей площадью листьев. Это обсуждается в следующем разделе.
Модели роста растений
Что ограничивает рост растений? Это важный вопрос, который имеет множество практических последствий. Ясно, что рост растений может быть ограничен неблагоприятными условиями окружающей среды, например, отсутствием осадков.
Но если условия идеальны для роста, что его ограничивает? Как и все организмы, растения растут, приобретая материал и встраивая его в свою собственную структуру. Можно предположить, что способность растения усваивать материал напрямую связана с его размером, причем более крупные организмы способны усваивать больше, чем маленькие организмы. Это привело бы к процессу роста с положительной обратной связью: приобретение ресурсов —> рост —> большее растение —> большее приобретение ресурсов —> больший рост —> еще большее растение и т. д. и т. д. Эту идею можно смоделировать в наборе уравнений, основанных на идее, что скорость роста является линейной функцией размера растения: чем больше растения, тем больше рост; больше роста больше растений.На словах эту идею можно выразить двумя способами:
(1) скорость роста определяется размером растения
(2) скорость роста, выраженная на единицу растения, является константой
В математическом выражении эти два утверждения:
(1) скорость роста = ∆S/∆t = k * S, где ∆S/∆t — скорость роста, изменение размера, деленное на изменение во времени, S — общий размер растения k — константа
(2) (∆S/∆t) * (1/S) = k,
Для большинства студентов-биологов это должно быть знакомо, потому что: (1) это звучит как «экспоненциальный» рост населения, или, возможно, «геометрический» рост населения, (2) это начинает походить на исчисление, курс, который часто требуется для студентов-биологов (и, возможно, теперь вы понимаете, почему!)
Вычисления могут привести к следующему, представив их в дифференциальной форме:
(1 a) dS/dt = k * S
(2 a) (dS/dt) * (1/S) = k
( 3) размер (S) в любой момент времени (t) определяется как S(t) = S o e (k*t)
Обратите внимание, что хотя переход к уравнению (3) требует исчисления, идеи уравнения 1 и 1а и их преобразование в уравнения 2 и 2а должны иметь смысл без него.
Уравнение (3) следует из любого из первых двух.Таким образом, можно было бы ожидать, что рост растений будет экспоненциальным, точно так же, как вы могли бы ожидать, что рост населения будет экспоненциальным. Обратите внимание, что значение слова «экспоненциальный» имеет математическое значение, не эквивалентное общеупотребительному. Экспоненциальный рост не обязательно является «быстрым», и действительно, быстрый — это субъективное прилагательное. В математическом смысле экспонента описывается уравнениями 1-3, хотя только 3 имеет в ней показатель степени. Можно разработать аналогичные уравнения, основанные на площади листа (то есть, что скорость роста на единицу площади листа является постоянной), с аргументом, что площадь листа, контролируя фотосинтез, диктует скорость роста. Однако аналогичный аргумент можно привести и в отношении корней, поскольку без воды и питательных веществ фотосинтез невозможен. Проще всего просто предположить, что необходимы корни, листья и все остальное, и пусть S будет просто «общим размером растения» и работать с предположением, что растения могут получать больше материалов со скоростью, которая является линейной функцией их размера (т.
, уравнение 1).Но рост растений, как и рост популяций, обычно не экспоненциальный, по крайней мере, ненадолго. Почему рост растений не экспоненциальный? Основной ответ заключается в том, что рост не контролируется скоростью приобретения материала. Это внутренне регулируемый процесс, и было бы слишком упрощенно предполагать, что скорость роста есть простая функция способности листьев и корней приобретать материалы, необходимые для роста. Внутренний контроль растения, включая как гормональный контроль, так и молекулярный контроль (например, какие гены активированы), регулирует процессы клеточного деления и расширения клеток и, таким образом, процесс роста. В фундаментальном смысле это ничем не отличается от того, что обсуждалось ранее в отношении развития одноклеточных организмов (см. главу 7 о развитии организмов). Второй фактор включает меристемы или, в более общем смысле, области роста. Если у организма есть только ограниченная область, где начинается рост, и эта область имеет конечную способность производить рост, тогда рост не будет экспоненциальным.
В случае нитчатых водорослей, которые растут из одной апикальной клетки, легко понять, почему рост не может быть экспоненциальным. Точно так же нельзя ожидать, что неразветвленный стебель с единственной апикальной меристемой будет демонстрировать экспоненциальный рост, хотя можно ожидать, что стебель с ветвями и с ветвями, которые могут давать больше ветвей, будет.Экономическая модель роста растений
Полезной аналогией для роста растений является экономическая «бизнес-модель». Валовой доход сначала делится на расходы (расходы на ведение бизнеса) и чистый доход («прибыль», то, что остается после оплаты расходов). Прибыль можно «инвестировать» различными способами: инвестиции, которые позволяют увеличить прибыль, инвестиции, которые защищают существующие структуры, или инвестиции, которые являются «легкомысленными», т. е. не имеющими очевидной выгоды. Например, пекарь получает доход от продажи хлеба. Часть своего дохода он использует на «обслуживание», покрывая расходы своей пекарни — на покупку муки, уплату налогов, оплату электроэнергии для работы печей, ремонт сломанного оборудования и т.
д. Доход, оставшийся после расходов на содержание, был уплаченные деньги можно инвестировать различными способами: (1) инвестиции в дополнительные печи и миксеры или, возможно, в совершенно новую пекарню. Эти инвестиции увеличат его доходы и приведут к экспоненциальному росту. (2) в средствах защиты, таких как спринклерная система для защиты его пекарни от огня или, возможно, система безопасности, которая снижает вероятность ограбления. Это может совсем не увеличить его заработок, но обеспечивает защиту от множества потенциальных проблем. (3) деньги могут быть потрачены на «легкомысленные» предметы, которые не защищают пекарню. Деньги могут просто храниться под его матрасом. Теоретически растения работают одинаково, приобретенный материал (прежде всего углеводы, но также и минеральные элементы) используется для обслуживания, а то, что осталось, может быть «инвестировано» в структуры (например, листья), которые позволят получить больше ресурсов, или могут быть вложены в структуры (например, химикаты против травоядных), которые защищают существующие структуры, или, возможно, приобретенный материал может вообще не быть вложен во что-либо «стоящее». Обратите внимание, что пока пекарь (или завод) инвестирует определенную часть своего заработка таким образом, чтобы увеличить его способность получать больший доход (даже если это очень небольшая часть), результатом будет экспоненциальный рост.Экспоненциальная модель роста чрезмерно упрощена и обычно плохо отражает реальность. Однако он дает отправную точку для анализа роста и основную идею о том, что по мере роста организмов (особенно растений) их способность к росту (скорость роста) увеличивается просто потому, что они больше.
- «Характеристики роста деревьев» Дженнифер Франклин и др. Рост дерева.
- https://extension.tennessee.edu/publications/Documents/W227.pdf
- «Выращивание яблок и управление урожаем», Стив МакАртни. Яблочный рост.
- http://umassfruitnotes.com/v76n1/a3.pdf
Media Attributions
 _Sitka_Spruce.jpg»> Рассада Picea sitchensis © Rosser1954 находится под лицензией CC BY-SA (Attribution ShareAlike) лицензия
_Sitka_Spruce.jpg»> Рассада Picea sitchensis © Rosser1954 находится под лицензией CC BY-SA (Attribution ShareAlike) лицензия - .
- Дом
- Диаграмма роста растений
- Войдите в свою учетную запись . Зарегистрируйтесь, указав свой адрес электронной почты и пароль, или создайте бесплатную учетную запись, чтобы попробовать сервис перед обновлением подписки.
- Загрузить документ . Перетащите файл со своего устройства или импортируйте его из других служб, таких как Google Диск, OneDrive, Dropbox или по внешней ссылке.
- Редактировать диаграмму роста растений . Быстро добавляйте и подчеркивайте текст, вставляйте изображения, галочки и значки, добавляйте новые заполняемые поля, а также меняйте порядок страниц или удаляйте их из документа.
- Заполнить таблицу роста растений . Загрузите обновленный документ, экспортируйте его в облако, распечатайте из редактора или поделитесь им с другими участниками с помощью общей ссылки или в виде вложения электронной почты.
 0002 предотвращает расширение . Это позволяет клетке создавать давление, и это важно для водного баланса и функционирования клеток растений.
0002 предотвращает расширение . Это позволяет клетке создавать давление, и это важно для водного баланса и функционирования клеток растений.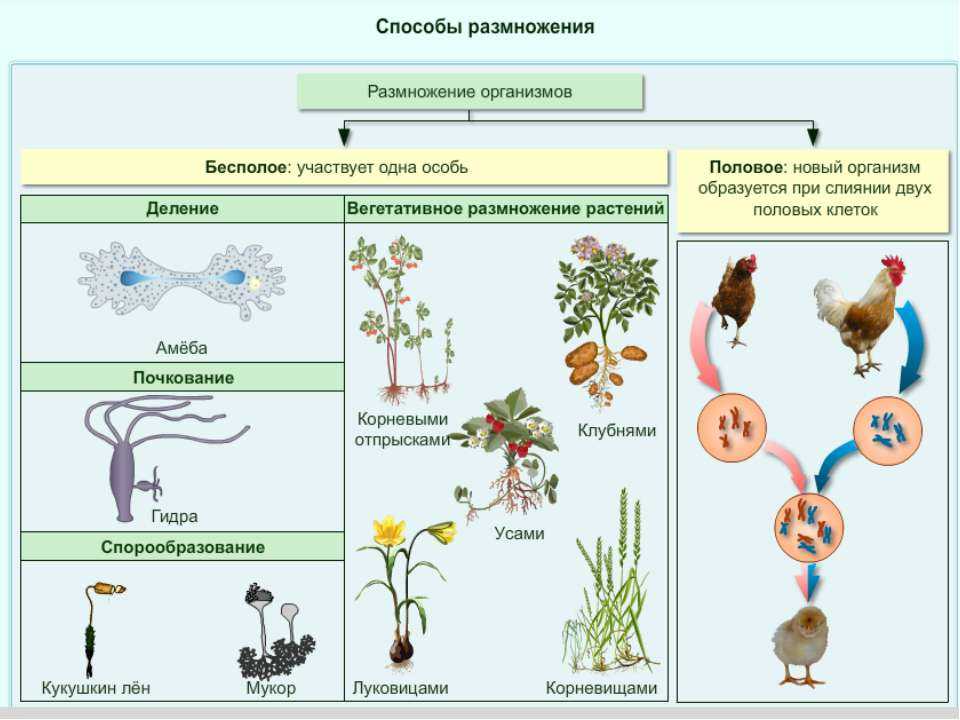
 .
. Углеводы часто не поступают напрямую из листьев в точки роста, а вместо этого могут поступать из листьев в места хранения, а затем из мест хранения в области роста.
Углеводы часто не поступают напрямую из листьев в точки роста, а вместо этого могут поступать из листьев в места хранения, а затем из мест хранения в области роста. В тех клетках, у которых есть вторичные клеточные стенки, дополнительный материал клеточной стенки откладывается после прекращения расширения клетки — обратите внимание, что, пока добавляются материалы вторичной клеточной стенки, клетка не увеличивается в размере, а увеличивается в массе.
В тех клетках, у которых есть вторичные клеточные стенки, дополнительный материал клеточной стенки откладывается после прекращения расширения клетки — обратите внимание, что, пока добавляются материалы вторичной клеточной стенки, клетка не увеличивается в размере, а увеличивается в массе. Таким образом, существует пространственное разделение между областью клеточного деления и областью клеточного роста (глава 8). Этот паттерн наиболее заметен при апикальном росте, но также имеет место и при вторичном (латеральном) росте (глава 9).), хотя и в сосудистом камбии, и в пробковом камбии рост клеток происходит с обеих сторон меристемы. Активность обычно возникает одновременно в области клеточного деления и в области клеточного роста. Наибольший рост органа происходит всякий раз, когда новым клеткам, продуцируемым меристемой, позволяют расширяться. Для некоторых растений рост более или менее непрерывен и стабилен, пока условия окружающей среды (в частности, температура, вода и свет) стабильны. Тем не менее, большинство растений демонстрируют эпизодический рост со вспышками образования и расширения клеток, за которыми следуют периоды бездействия, даже при постоянных и благоприятных условиях. Для многих многолетних растений, обитающих в районах с сезонным климатом, рост строго сезонный и происходит только часть года.
Таким образом, существует пространственное разделение между областью клеточного деления и областью клеточного роста (глава 8). Этот паттерн наиболее заметен при апикальном росте, но также имеет место и при вторичном (латеральном) росте (глава 9).), хотя и в сосудистом камбии, и в пробковом камбии рост клеток происходит с обеих сторон меристемы. Активность обычно возникает одновременно в области клеточного деления и в области клеточного роста. Наибольший рост органа происходит всякий раз, когда новым клеткам, продуцируемым меристемой, позволяют расширяться. Для некоторых растений рост более или менее непрерывен и стабилен, пока условия окружающей среды (в частности, температура, вода и свет) стабильны. Тем не менее, большинство растений демонстрируют эпизодический рост со вспышками образования и расширения клеток, за которыми следуют периоды бездействия, даже при постоянных и благоприятных условиях. Для многих многолетних растений, обитающих в районах с сезонным климатом, рост строго сезонный и происходит только часть года. Часто период роста составляет лишь очень небольшую часть «благоприятного» периода времени. Например, многие деревья в северных районах с умеренным климатом растут только в течение двух-четырех недель в мае.
Часто период роста составляет лишь очень небольшую часть «благоприятного» периода времени. Например, многие деревья в северных районах с умеренным климатом растут только в течение двух-четырех недель в мае.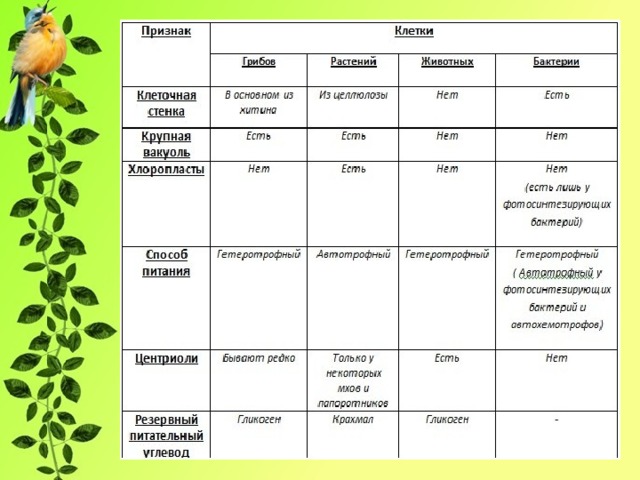 Однако эти структуры do продолжают развиваться: проявляют старение и опадение в случае листьев и в случае плодов, демонстрирующих созревание таким образом, что это способствует распространению семян (и это часто также включает опадение).
Однако эти структуры do продолжают развиваться: проявляют старение и опадение в случае листьев и в случае плодов, демонстрирующих созревание таким образом, что это способствует распространению семян (и это часто также включает опадение).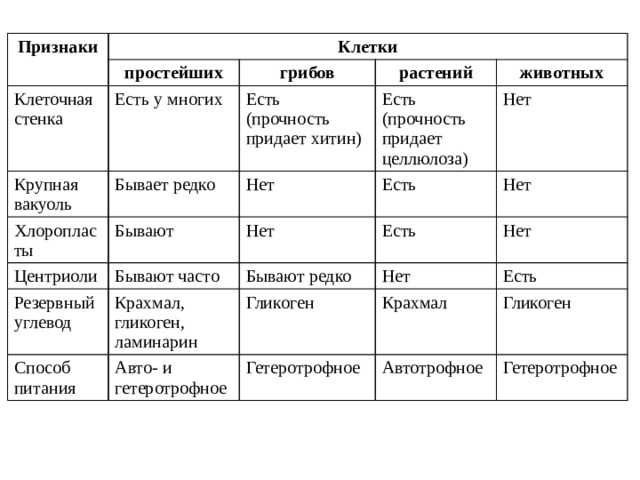

 Более того, для всех этих факторов может быть «слишком много хорошего», и дальнейшее увеличение фактически снижает рост (токсичность).
Более того, для всех этих факторов может быть «слишком много хорошего», и дальнейшее увеличение фактически снижает рост (токсичность).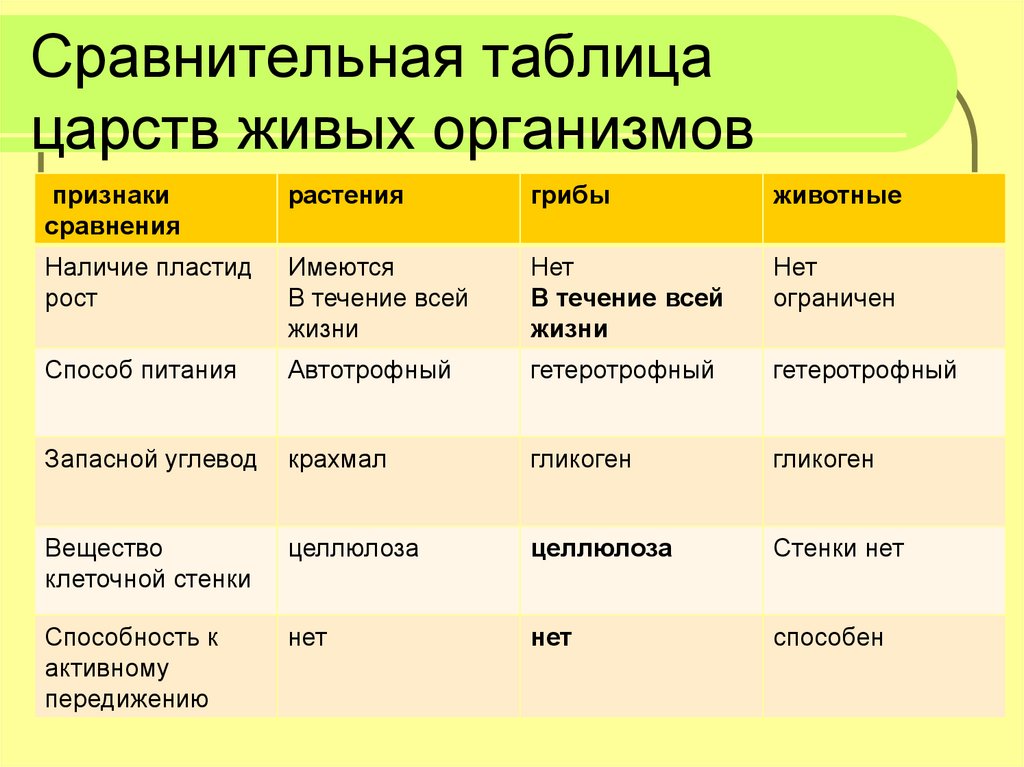 Ограничение может возникать как из-за отсутствия этого элемента в почве, так и из-за того, что элемент хотя и присутствует, но недоступен из-за почвенных условий. Например, железо часто недоступно в основных почвах, хотя оно может присутствовать в изобилии. Проблема в том, что в аэробных, основных условиях очень мало железа присутствует в форме, которая легко растворяется.
Ограничение может возникать как из-за отсутствия этого элемента в почве, так и из-за того, что элемент хотя и присутствует, но недоступен из-за почвенных условий. Например, железо часто недоступно в основных почвах, хотя оно может присутствовать в изобилии. Проблема в том, что в аэробных, основных условиях очень мало железа присутствует в форме, которая легко растворяется. Выращивание сельскохозяйственных культур можно сравнить с выпечкой торта: если рецепт торта требует пяти ингредиентов, приготовление торта может быть ограничено любым из пяти ингредиентов, а недостаток одного не компенсируется избытком других. Это очень простая идея, применимая во многих ситуациях. Но это противоречит общепринятой идее о том, что реакция на факторы всегда будет постоянной: «если немного хорошо, то многое должно быть лучше».0002 не случай!
Выращивание сельскохозяйственных культур можно сравнить с выпечкой торта: если рецепт торта требует пяти ингредиентов, приготовление торта может быть ограничено любым из пяти ингредиентов, а недостаток одного не компенсируется избытком других. Это очень простая идея, применимая во многих ситуациях. Но это противоречит общепринятой идее о том, что реакция на факторы всегда будет постоянной: «если немного хорошо, то многое должно быть лучше».0002 не случай!

 Е. Короткие периоды очень яркого света не эквивалентны более длительным периодам менее яркого света. Большинство видов сельскохозяйственных культур адаптированы к условиям высокой освещенности и очень плохо себя чувствуют при выращивании в затененных условиях, и, вероятно, рост урожая может быть снижен в результате продолжительных облачных условий. Однако такие условия часто связаны с частыми дождями, которые также могут быть причиной замедления роста в случае затопления почв.
Е. Короткие периоды очень яркого света не эквивалентны более длительным периодам менее яркого света. Большинство видов сельскохозяйственных культур адаптированы к условиям высокой освещенности и очень плохо себя чувствуют при выращивании в затененных условиях, и, вероятно, рост урожая может быть снижен в результате продолжительных облачных условий. Однако такие условия часто связаны с частыми дождями, которые также могут быть причиной замедления роста в случае затопления почв.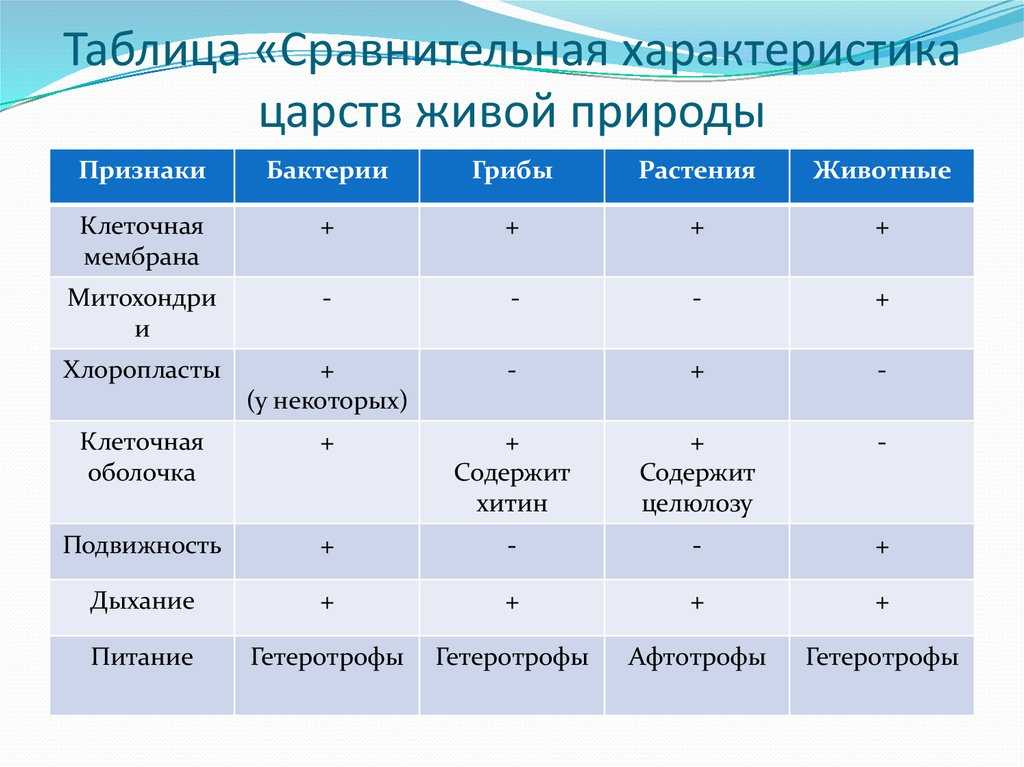 е. количество дыхания, необходимое листу для поддержания его жизненных условий) постоянна, затенение снизит общий фотосинтез до такой степени, что чистый фотосинтез отрицательный, т. е. содержание листа обходится дороже, чем он «зарабатывает» в процессе фотосинтеза. В этот момент растение может сократить свои потери, удалив лист.
е. количество дыхания, необходимое листу для поддержания его жизненных условий) постоянна, затенение снизит общий фотосинтез до такой степени, что чистый фотосинтез отрицательный, т. е. содержание листа обходится дороже, чем он «зарабатывает» в процессе фотосинтеза. В этот момент растение может сократить свои потери, удалив лист. Частично это объясняется тем, что при более высоких температурах дыхание более чувствительно к температуре (т. е. больше увеличивается с повышением температуры), чем фотосинтез. температура (см. уравнение выше).
Частично это объясняется тем, что при более высоких температурах дыхание более чувствительно к температуре (т. е. больше увеличивается с повышением температуры), чем фотосинтез. температура (см. уравнение выше). Но если условия идеальны для роста, что его ограничивает? Как и все организмы, растения растут, приобретая материал и встраивая его в свою собственную структуру. Можно предположить, что способность растения усваивать материал напрямую связана с его размером, причем более крупные организмы способны усваивать больше, чем маленькие организмы. Это привело бы к процессу роста с положительной обратной связью: приобретение ресурсов —> рост —> большее растение —> большее приобретение ресурсов —> больший рост —> еще большее растение и т. д. и т. д. Эту идею можно смоделировать в наборе уравнений, основанных на идее, что скорость роста является линейной функцией размера растения: чем больше растения, тем больше рост; больше роста больше растений.
Но если условия идеальны для роста, что его ограничивает? Как и все организмы, растения растут, приобретая материал и встраивая его в свою собственную структуру. Можно предположить, что способность растения усваивать материал напрямую связана с его размером, причем более крупные организмы способны усваивать больше, чем маленькие организмы. Это привело бы к процессу роста с положительной обратной связью: приобретение ресурсов —> рост —> большее растение —> большее приобретение ресурсов —> больший рост —> еще большее растение и т. д. и т. д. Эту идею можно смоделировать в наборе уравнений, основанных на идее, что скорость роста является линейной функцией размера растения: чем больше растения, тем больше рост; больше роста больше растений. Уравнение (3) следует из любого из первых двух.
Уравнение (3) следует из любого из первых двух. , уравнение 1).
, уравнение 1). В случае нитчатых водорослей, которые растут из одной апикальной клетки, легко понять, почему рост не может быть экспоненциальным. Точно так же нельзя ожидать, что неразветвленный стебель с единственной апикальной меристемой будет демонстрировать экспоненциальный рост, хотя можно ожидать, что стебель с ветвями и с ветвями, которые могут давать больше ветвей, будет.
В случае нитчатых водорослей, которые растут из одной апикальной клетки, легко понять, почему рост не может быть экспоненциальным. Точно так же нельзя ожидать, что неразветвленный стебель с единственной апикальной меристемой будет демонстрировать экспоненциальный рост, хотя можно ожидать, что стебель с ветвями и с ветвями, которые могут давать больше ветвей, будет. д. Доход, оставшийся после расходов на содержание, был уплаченные деньги можно инвестировать различными способами: (1) инвестиции в дополнительные печи и миксеры или, возможно, в совершенно новую пекарню. Эти инвестиции увеличат его доходы и приведут к экспоненциальному росту. (2) в средствах защиты, таких как спринклерная система для защиты его пекарни от огня или, возможно, система безопасности, которая снижает вероятность ограбления. Это может совсем не увеличить его заработок, но обеспечивает защиту от множества потенциальных проблем. (3) деньги могут быть потрачены на «легкомысленные» предметы, которые не защищают пекарню. Деньги могут просто храниться под его матрасом. Теоретически растения работают одинаково, приобретенный материал (прежде всего углеводы, но также и минеральные элементы) используется для обслуживания, а то, что осталось, может быть «инвестировано» в структуры (например, листья), которые позволят получить больше ресурсов, или могут быть вложены в структуры (например, химикаты против травоядных), которые защищают существующие структуры, или, возможно, приобретенный материал может вообще не быть вложен во что-либо «стоящее».
д. Доход, оставшийся после расходов на содержание, был уплаченные деньги можно инвестировать различными способами: (1) инвестиции в дополнительные печи и миксеры или, возможно, в совершенно новую пекарню. Эти инвестиции увеличат его доходы и приведут к экспоненциальному росту. (2) в средствах защиты, таких как спринклерная система для защиты его пекарни от огня или, возможно, система безопасности, которая снижает вероятность ограбления. Это может совсем не увеличить его заработок, но обеспечивает защиту от множества потенциальных проблем. (3) деньги могут быть потрачены на «легкомысленные» предметы, которые не защищают пекарню. Деньги могут просто храниться под его матрасом. Теоретически растения работают одинаково, приобретенный материал (прежде всего углеводы, но также и минеральные элементы) используется для обслуживания, а то, что осталось, может быть «инвестировано» в структуры (например, листья), которые позволят получить больше ресурсов, или могут быть вложены в структуры (например, химикаты против травоядных), которые защищают существующие структуры, или, возможно, приобретенный материал может вообще не быть вложен во что-либо «стоящее». Обратите внимание, что пока пекарь (или завод) инвестирует определенную часть своего заработка таким образом, чтобы увеличить его способность получать больший доход (даже если это очень небольшая часть), результатом будет экспоненциальный рост.
Обратите внимание, что пока пекарь (или завод) инвестирует определенную часть своего заработка таким образом, чтобы увеличить его способность получать больший доход (даже если это очень небольшая часть), результатом будет экспоненциальный рост.Шаблон диаграммы роста растений
: Заполните и подпишите онлайн Шаблон диаграммы роста растений
: Заполните и подпишите онлайн | докхаб
Получить форму
4.2 из 5
29 голосов
Отзывы DocHub
44 отзыва
Отзывы DocHub
23 оценки
15 005
10 000 000+
303
100 000+ пользователей
Вот как это работает
01. Редактируйте диаграмму роста растений онлайн
Редактируйте диаграмму роста растений онлайн
Введите текст, добавьте изображения, затемните конфиденциальные данные, добавьте комментарии, выделение и многое другое.
02. Подпишите в несколько кликов
Нарисуйте свою подпись, введите ее, загрузите ее изображение или используйте мобильное устройство в качестве панели для подписи.
03. Поделитесь своей формой с другими
Отправьте рабочую таблицу диаграммы роста по электронной почте, по ссылке или по факсу. Вы также можете скачать его, экспортировать или распечатать.
Самый простой способ изменить диаграмму роста растений в формате PDF онлайн
9.5
Простота настройки
Рейтинги пользователей DocHub на G2
9.0
Простота использования
Рейтинги пользователей DocHub на G2
Работать с документами с помощью нашего универсального и интуитивно понятного редактора PDF очень просто. Выполните следующие действия, чтобы легко и быстро заполнить диаграмму роста растений онлайн:
 Зарегистрируйтесь, указав свой адрес электронной почты и пароль, или создайте бесплатную учетную запись, чтобы попробовать сервис перед обновлением подписки.
Зарегистрируйтесь, указав свой адрес электронной почты и пароль, или создайте бесплатную учетную запись, чтобы попробовать сервис перед обновлением подписки.Максимально используйте DocHub — самый простой редактор для быстрого управления документацией в Интернете!
будьте готовы получить больше
Заполните эту форму за 5 минут или меньше
Получить форму
Есть вопросы?
У нас есть ответы на самые популярные вопросы наших клиентов. Если вы не можете найти ответ на свой вопрос, пожалуйста, свяжитесь с нами.
Если вы не можете найти ответ на свой вопрос, пожалуйста, свяжитесь с нами.
Свяжитесь с нами
Сколько растений посадили на 3-й неделе?
(c) Растение А выросло = 9 см \ u2013 7 см = 2 см в течение 3-й недели.
Сколько недель растет растение?
Большинство растений прорастают в течение 1-2 недель, а некоторые плодоносящие растения в некоторых случаях занимают до 3 недель. Если ваши растения не взошли в течение 3 недель, то обязательно обратитесь в нашу службу поддержки, которая поможет вам решить любые проблемы.
Каковы 5 стадий роста растений?
Они следуют циклическому процессу начала новой жизни, роста и последующего возвращения к исходной стадии (размножения). Различают 5 стадий жизненного цикла растений. Этапы посева, прорастания, роста, размножения, опыления и распространения семян.
Различают 5 стадий жизненного цикла растений. Этапы посева, прорастания, роста, размножения, опыления и распространения семян.
Каковы 3 стадии роста растений?
Станьте лучшим садоводом: изучите 3 стадии роста растений прямо сейчас! Стадия семян (прорастание семян) Стадия вегетации (рост) Стадия размножения, цветения и плодов.
Каковы 3 стадии роста растений?
Станьте лучшим садоводом: изучите 3 стадии роста растений прямо сейчас! Стадия семян (прорастание семян) Стадия вегетации (рост) Стадия размножения, цветения и плодов.
таблица роста растений
таблица роста растений для дошкольников
Связанные формы
будьте готовы получить больше
Заполните эту форму за 5 минут или меньше
Получить форму
Люди также спрашивают
Каковы 4 стадии прорастания?
Процесс прорастания семян: набухание: вода наполняет семена. Вода активирует ферменты, которые запускают рост растения. Из семени вырастает корень, чтобы получить доступ к подземной воде. Из семени вырастают побеги, которые растут к солнцу. На побегах вырастают листья и начинается фотоморфогенез.
Вода активирует ферменты, которые запускают рост растения. Из семени вырастает корень, чтобы получить доступ к подземной воде. Из семени вырастают побеги, которые растут к солнцу. На побегах вырастают листья и начинается фотоморфогенез.
Какое растение может вырасти за 3 недели?
14 быстрорастущих овощей для вашего весеннего сада Кресс-салат: 14 дней. Всего за две недели вы можете собрать кресс-салат, траву с острым, острым вкусом. … Руккола: от 2 до 3 недель. … Всходы гороха: 2-3 недели. … Редька: 3 недели. … Мизуна: 3 недели. … Зеленый лук: 3 недели. … Детская капуста: 3-4 недели. … Бэби Бок Чой: 3-4 недели.
Каковы закономерности роста растений и животных?
— Мы знаем, что растения продолжают расти, но животные перестают расти до определенного периода; он растет в течение определенного периода и останавливается. Рост растений локализован в определенных областях, таких как корень, листья. Рост у животных диффузный, происходит в теле и активных областях.
Рост растений локализован в определенных областях, таких как корень, листья. Рост у животных диффузный, происходит в теле и активных областях.
Что такое модель роста растений?
Рост растений происходит по мере удлинения стеблей и корней. Некоторые растения, особенно древесные, также увеличиваются в толщине в течение жизни. Увеличение длины побега и корня называется первичным ростом и является результатом деления клеток в апикальной меристеме побега.
Каковы 4 стадии роста растений?
История вопроса: Жизненный цикл растения состоит из четырех стадий; семя, росток, маленькое растение и взрослое растение. Когда семя посадят в почву с водой и солнцем, оно начнет расти в маленький росток.