Содержание
Какие факторы окружающей среды оказывают влияние на рост и развитие корня?
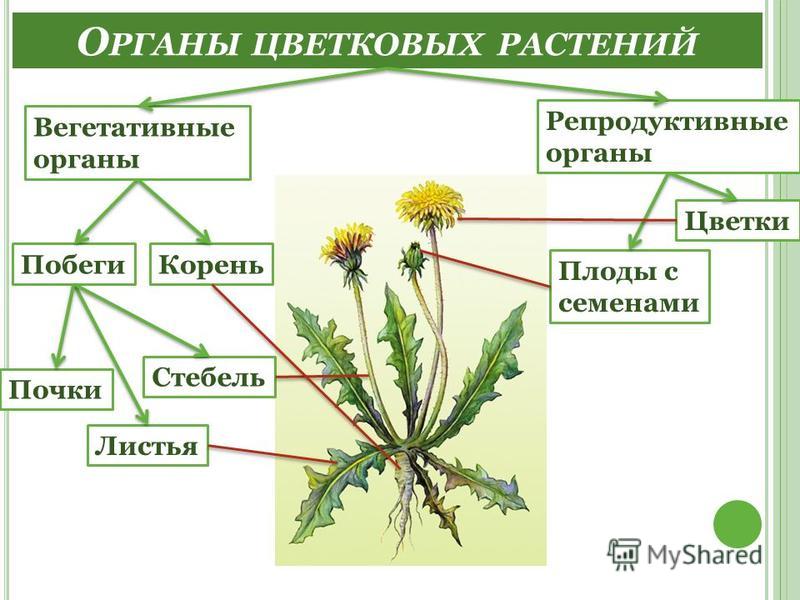
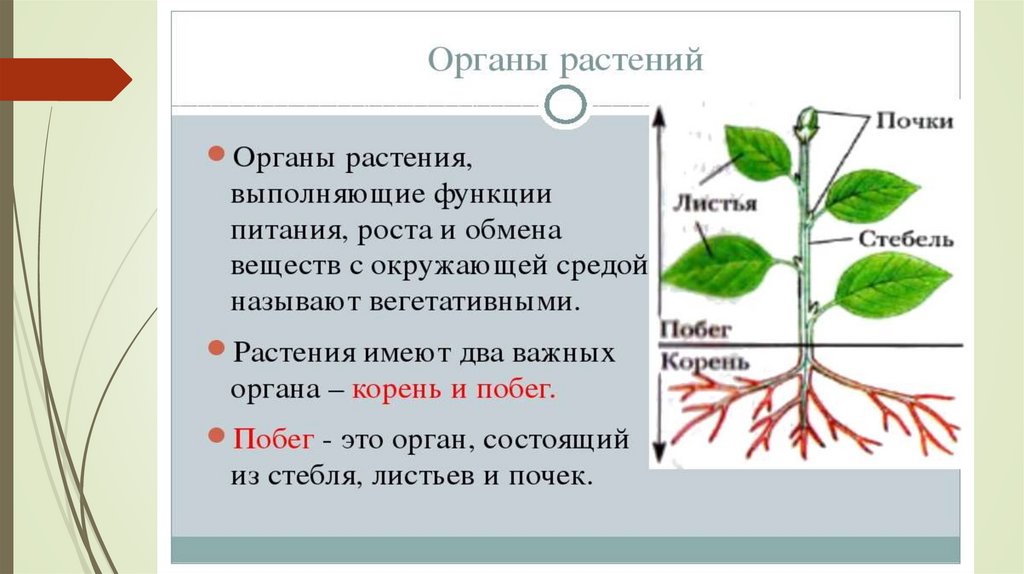
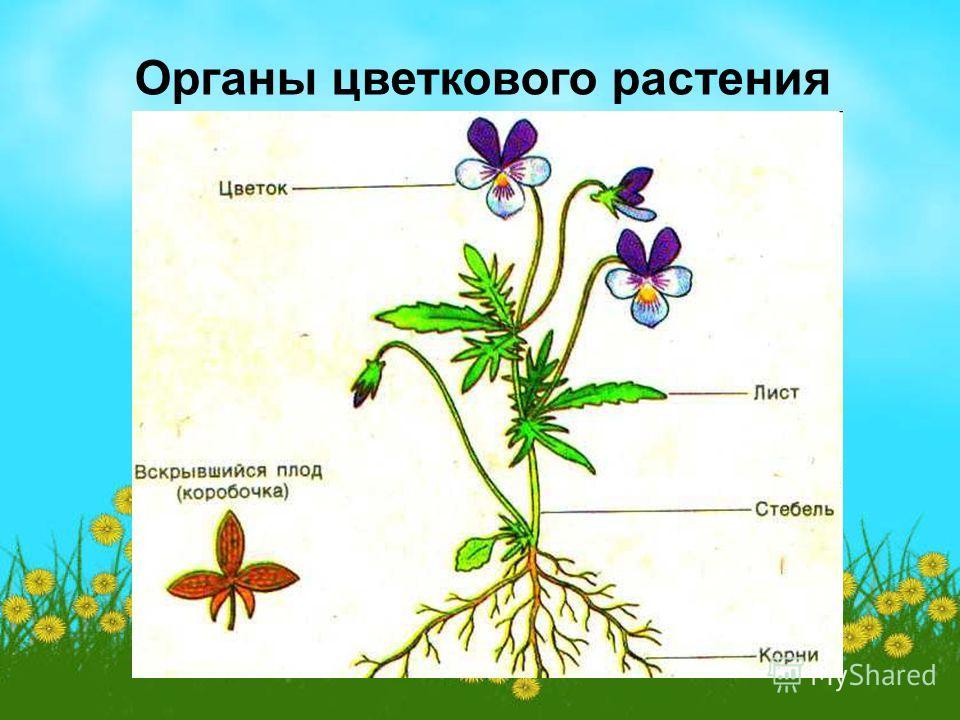
Корнем называют подземный орган растения, особенностями которого являются непрерывный рост и способность к ветвлению. Основной функцией корня является поглощение влаги и питательных веществ, находящихся в почве, которые необходимы для питания растения.
Глубина, которой способен достичь корень, степень его ветвления, его форма зависят как от различных внешних факторов, так и от типа корневой системы, который присущ определенному виду растений.
Строение корня
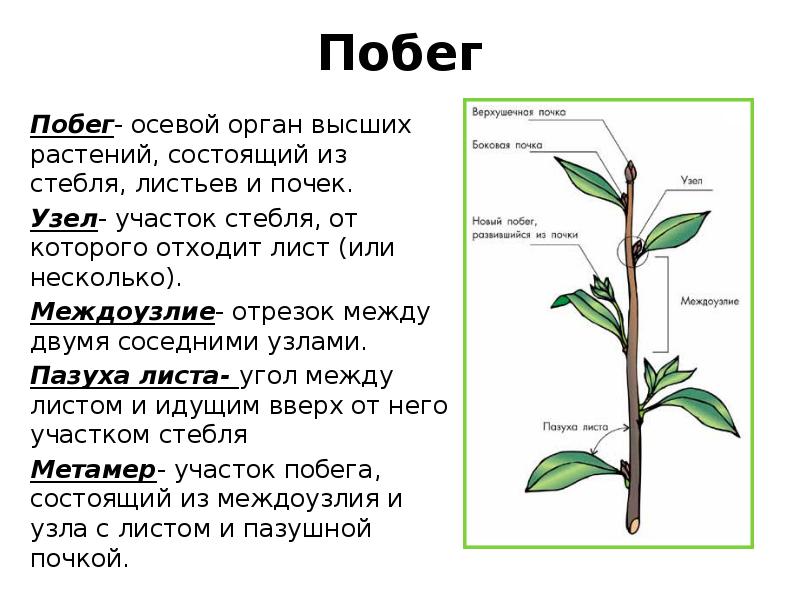
Каждый корень подразделяется на несколько частей:
- зона проведения;
- зона всасывания;
- зона роста;
- зона деления;
- корневой чехлик, которым заканчивается зона деления и который служит для оберегантя корня от повреждений в процессе роста.
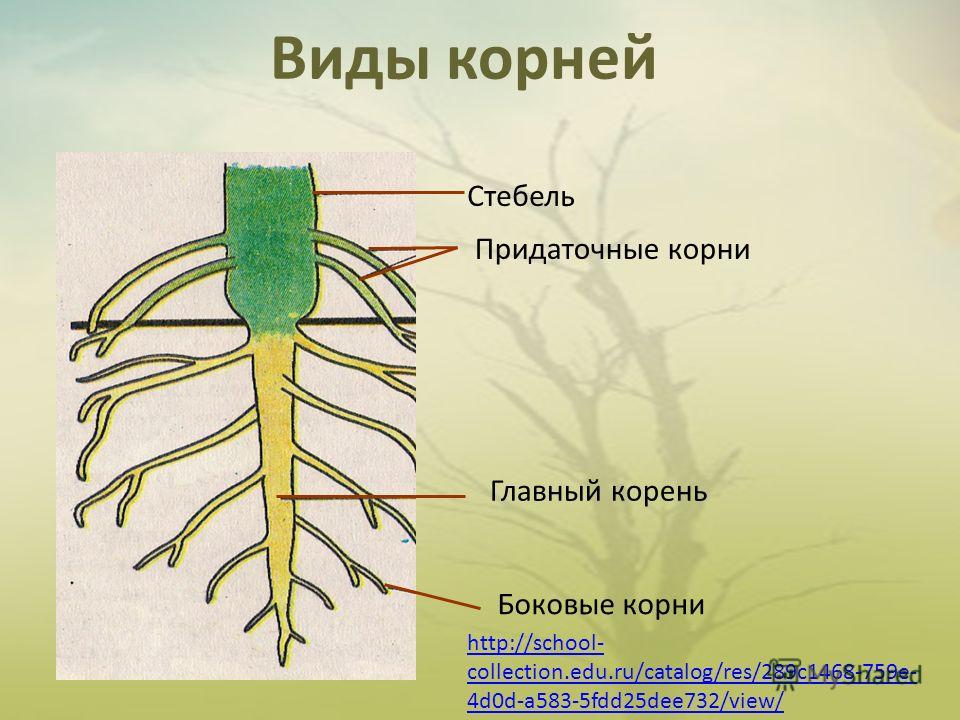
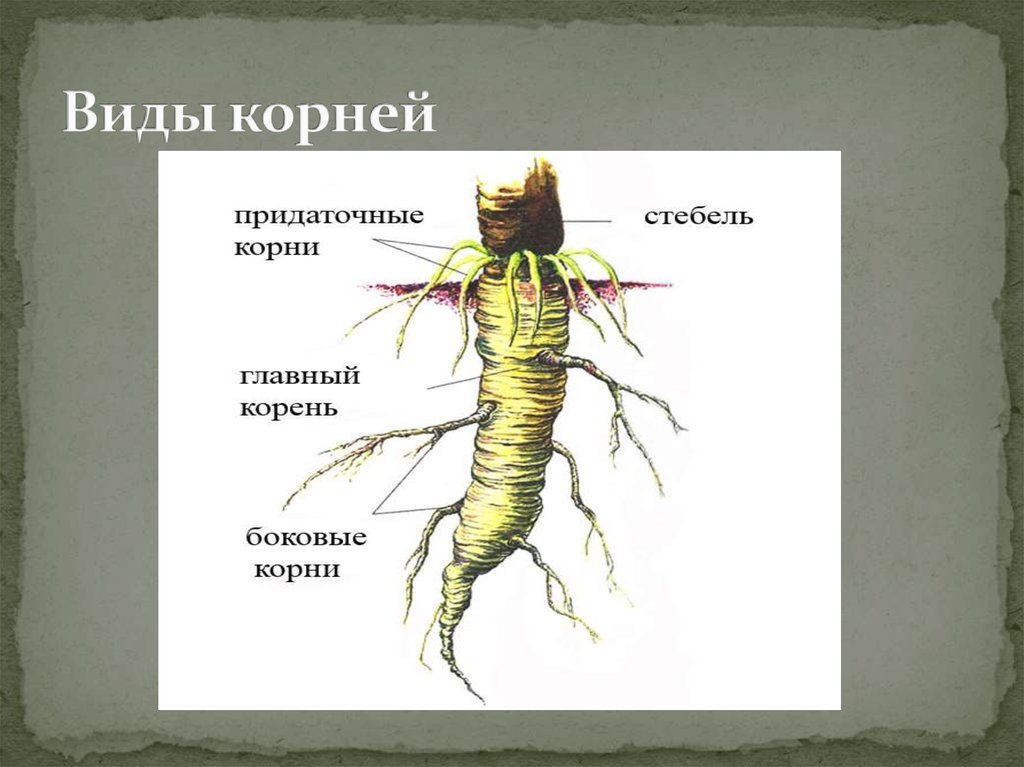
Основные виды корней
В зависимости от своего происхождения корни подразделяются на:
- главный корень, который образуется из зародышевого корешка;
- придаточные корни, которые вырастают из надземных частей растения;
- боковые корни, которые являются ответвлениями главного корня.


Все разнообразие корней растения в совокупности называется корневой системой.
Корневые системы
В соответствии с тем, как развиваются главный, боковые и придаточные корни, различают такие типы корневых систем:
- Стержневая система, при которой наиболее развитым является главный корень, а боковые и придаточные малоразвиты и поэтому форма корня кажется удлиненной.
- Мочковатая система, при которой придаточные и боковые корни хорошо развиты, в то время, как главный корень не выделяется среди них и создается впечатление, что главного корня вообще нет.
- Смешанная система, при которой главный корень является наиболее развитым, но придаточные и боковые корни тоже хорошо развиты.
Также в зависимости от распределения корней в почве, различают глубинный, поверхностный и универсальный типы корневых систем.
Рост и развитие корневых систем
Факторы, влияющие на рост и развитие корневой системы:
- плодородие почвы;
- влажность почвы;
- температура почвы;
- воздухопроницаемость почвы;
- состояние надземной части растения;
- экология;
- механические повреждения корней.
Важным фактором, влияющим на рост и развитие корня и всего растения в целом, является уровень плодородности почвы. Если почва не плодородна, то корни развиваются лучше, чем надземная часть растения, а если почва плодородна, то корневая система развивается хуже надземной части, так как растению нет необходимости в большом количестве корней.
Чрезмерная влажность почвы способствует торможению роста корней. Но в то же время, и недостаточная влажность задерживает их рост. Поэтому, каждому растению необходима собственная оптимальная влажность почвы для развития корневой системы.
Некоторые растения, приспособленные к существованию в засушливых регионах планеты выработали собственные механизмы добычи и сохранения влаги. Например, главный корень, уходящий глубоко под землю в поисках воды.
Температура, так же как и влажность, должна быть оптимальной для каждого вида растений. Слишком низкая температура почвы может стал причиной отмирания корней. При этом, если немного понижается температура, лучше развивается корневая система, а если увеличивается, то лучше развивается надземная часть растения.
Сильно уплотненные почвы, в которые проникает мало воздуха и которые являются слишком твердыми, замедляют рост корней.
Интенсивность фотосинтеза также оказывает влияние на рост подземной части растения. Если постоянно удалять с растения части, в которых происходит процесс фотосинтеза, то рост корней также замедляется.
Замедляет рост корневой системы также и обильный урожай.
Загрязнение почв мусором и отходами в большинстве случаев также сказывается негативно на развитии корней растений. Напротив, благоприятная экологическая обстановка способствует гармоничному развитию корневой системы.
Механическое повреждение корней может произойти в результате попадания растущего корня в скопления мусора (пластикового, металлического и др.), столкновения с твердыми породами, деятельности человека или животных, природных катаклизмов и т. д. Если у корня сильно повреждена верхушка, то он прекращает свой рост в длину, но при этом усиливают свой рост боковые ответвления и корневая система становится более разветвленной и обширной. Данное явление широко используется в сельском хозяйстве для увеличения урожайности различных культур.
Но в том случае, если главный корень слишком поврежден, растение может погибнуть. Также существуют виды растений, которые при остановке роста главного корня, также прекращают свой рост в целом.

 Данное явление широко используется в сельском хозяйстве для увеличения урожайности различных культур.
Данное явление широко используется в сельском хозяйстве для увеличения урожайности различных культур.Разработка урока . Тема: «Корень. Многообразие корней»
«Видоизменения корней».
Оборудование: рисунки , тексты.
Ход урока:
Д.з. Пересказ параграфа и ответы на вопросы.(10 минут)
Вызов: 7 минут
Вы уже несколько уроков изучаете тему корень.
Самые длинные корни – 120м у дикого инжира, он растет в восточной Африке. В Шотландии вяз имеет корневую систему – 110м. Глубокие корни имеют растения, растущие в пустынях от 20 до 50м. Подумайте? От чего это зависит?
Кроме функции доставки раствора минеральных солей, т. е корневого питания какие еще выполняет функции корень?
е корневого питания какие еще выполняет функции корень?
Чтобы выполнять эти функции корню необходимо иметь соответствующее строение. Что же нам предстоит выяснить?
ОТВЕТЫ:
1. Изучить видоизмененные корни.
2. Узнать их значение.
3. Выяснить, что видоизмененные
корни результат приспособленности
растений к условиям существования.
Отгадайте загадки:
1) Круглая, а не месяц
Желтая, а не масло
С хвостиком, а не мышь (репа).
2) За хохол, да в котел, а потом – борщок на стол. (свёкла)
Рассмотрите рисунки корней отгаданных вами растений.
Какой вид корня у них разросся?( Утолщение главного корня и у некоторых утолщаются ещё и нижние участки стебля)
Какую функцию они выполняют? (Запасающую)
Посмотрите внимательно на изображение подземной части георгина и батата.
Что это?
Как они образуются?
Рассмотрите рисунок орхидеи
Наружная часть коры, свисающая вниз придаточных воздушных корней этих растений состоит из крупных клеток, которые могут впитывать воду подобно губке. Во время дождя эти клетки наполняются водой. Для чего растению такие корни?
Во время дождя эти клетки наполняются водой. Для чего растению такие корни?
Посмотрите внимательно на следующий рисунок болотного кипариса, скажите с какими трудностями, сталкивается это растение?
Чего не хватает этому растению?
Какую функцию выполняет корень?
Посмотрите на рисунок плюща.
Что имеется у плюща по всему стеблю?
Какова роль корней таких корней?
Содержание
Это все только наши с вами предположения на основе наблюдений и сравнения.
Чтобы выяснить где же мы были правы, а где заблуждались изучим информацию текста ВИДОИЗМЕНЕНИЯ КОРНЕЙ и заполним таблицу.
Заполняют таблицу с опорой на текст 10 минут
Название видоизменения корня | Особенности строения | Примеры растений |
ФИЗКУЛЬМИНУТКА
Для разминки из-за парт
Поднимаемся. На старт! .
На старт! .
Объявление результатов работы. 5 минут
Название видоизменения корня | Особенности строения | Примеры растений |
Корнеплоды | Разросшийся главный корень. | Морковь, редис, свёкла, репа. |
Корневые клубни | Утолщение придаточных и боковых корней. | Георгин, батат. |
Воздушные корни | Длинные придаточные корни, всасывающие воду. | Орхидея, монстера. |
Дыхательные корни | Боковые корни, которые растут вертикально вверх. | Кипарис болотный, ива ломкая. |
Корни — прицепки | Короткие придаточные корни с мощными прицепками. | Плющ. |
1. У разных растений корневая система имеет видоизменения. От чего это зависит? От условий- экологических факторов.
Каких? (влажности, плодородия почв, климата, освещения, плотности почв)
Таким образом мы выяснили, что кроме функции питания корень способен выполнять еще ряд функций. Обобщите данные таблицы и статьи «Корень- это важный орган растения» и оформите схему: 7 минут.
Рефлексия
Тест до конца урока
Индивидуально
1.(1,5 балла). Выберите три верных ответа из шести и запишите цифры, под которыми они указаны.
Корнеплод имеют:
Георгин
Репа
Турнепс
Плющ
Баньян
Брюква.
2. (1 балл)Какую из перечисленных функций корни не выполняют
(1 балл)Какую из перечисленных функций корни не выполняют
1) закрепление растений в почве
2) всасывание воды и минеральных солей из почвы
3) запасание питательных веществ
4) образование питательных веществ в растении
3. (1 балл) В результате утолщения боковых и придаточных корней образуются
А) воздушные корни
Б) корневые клубни
В) корнеплоды
Г) корни-присоски
Батат Георгин
Орхидея Плющ
Корень — это важный орган растения.
Корень выполняет ряд жизненно необходимых функций, и ими определяется его значение для растений.
Корень осуществляет почвенное питание растения, он всасывает воду и минеральные вещества. Поэтому чем лучше у растения развита корневая система, тем оно лучше его почвенное питание.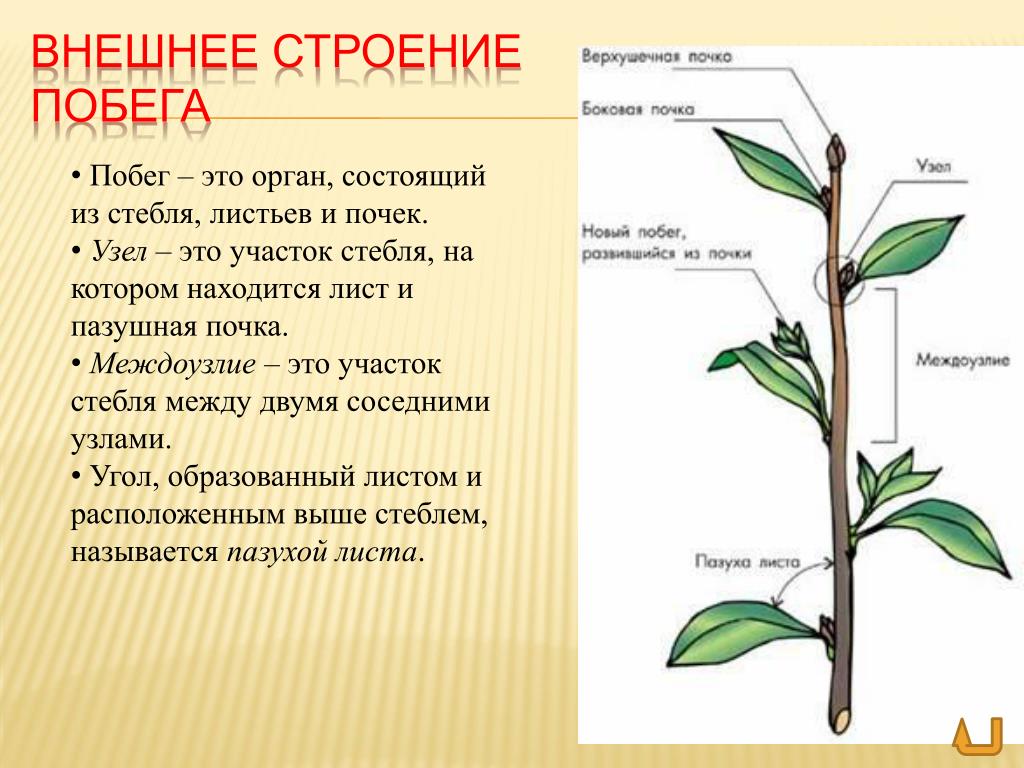 Среди минеральных веществ, которые в растение поступают из почвы, есть соли калия, кальция, магния, фосфора, ряд соединений азота и серы. Все эти вещества необходимы для обмена веществ, образования клеток и их органелл. Например, магний важен для образования хлорофилла.
Среди минеральных веществ, которые в растение поступают из почвы, есть соли калия, кальция, магния, фосфора, ряд соединений азота и серы. Все эти вещества необходимы для обмена веществ, образования клеток и их органелл. Например, магний важен для образования хлорофилла.
Помимо почвенного питания корень выполняет еще одну важную функцию. Он удерживает растение в почве. Причем удерживает его достаточно прочно, чтобы ни сильный ветер, ни проливные дожди не выдернули или не вымыли растение из почвы. Корень не только прочный, но и гибкий. Это достигается расположением проводящих сосудов и механической ткани в его центре.
С помощью корня может осуществляться вегетативное размножение. В этом случае на корнях образуются придаточные почки. Из них потом развиваются побеги. Они могут быть отделены от растения и жить самостоятельно. Такими корневыми отпрысками размножаются, например, одуванчик, слива, малина, сирень.
Еще одной функцией корня является запас питательных веществ.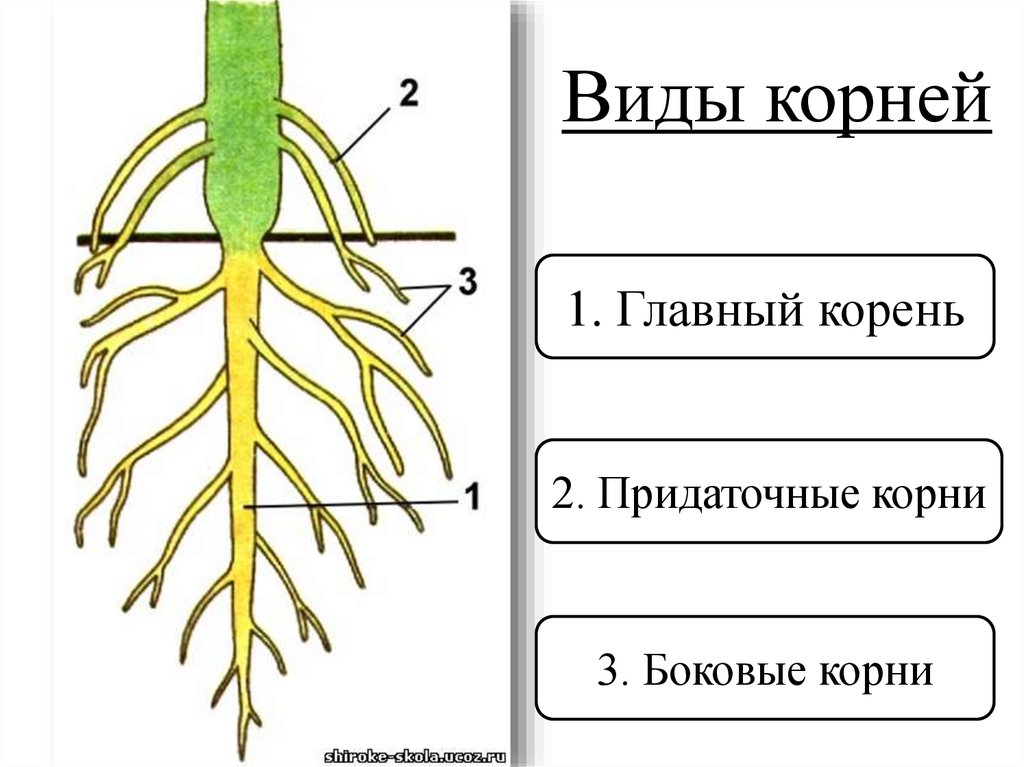 У ряда растений они откладываются в корне.
У ряда растений они откладываются в корне.
Корни имеют значение не только для самих растений, но и для остальной природы. Корни растений укрепляют почвы. Особенно это важно для рыхлых песчаных почв. Если же почва глинистая или каменистая, то корни делают их более рыхлыми. Отмирающие корни и их корневые волоски, отпавшие слизистые клетки корневого чехлика служат пищей для других организмов почвы, в том числе бактерий и грибов. Таким образом, корни делают почву более плодородной.
Кроме того сами корни могут служить пищей животным и человеку, когда в них накапливается запас питательных веществ.
Кроме перечисленных функций у некоторых растений корни могут выполнять другие специфические функции. Растения нередко вступают в симбиоз с бактериями, а также с грибами. Азотфиксирующие клубеньковые бактерии поселяются на корневой системе бобовых культур, из-за этого на корневище формируются клубеньки. При симбиозе растений, чаще деревьев, с грибами их корни сожительствуют с грибными гифами, получается микориза.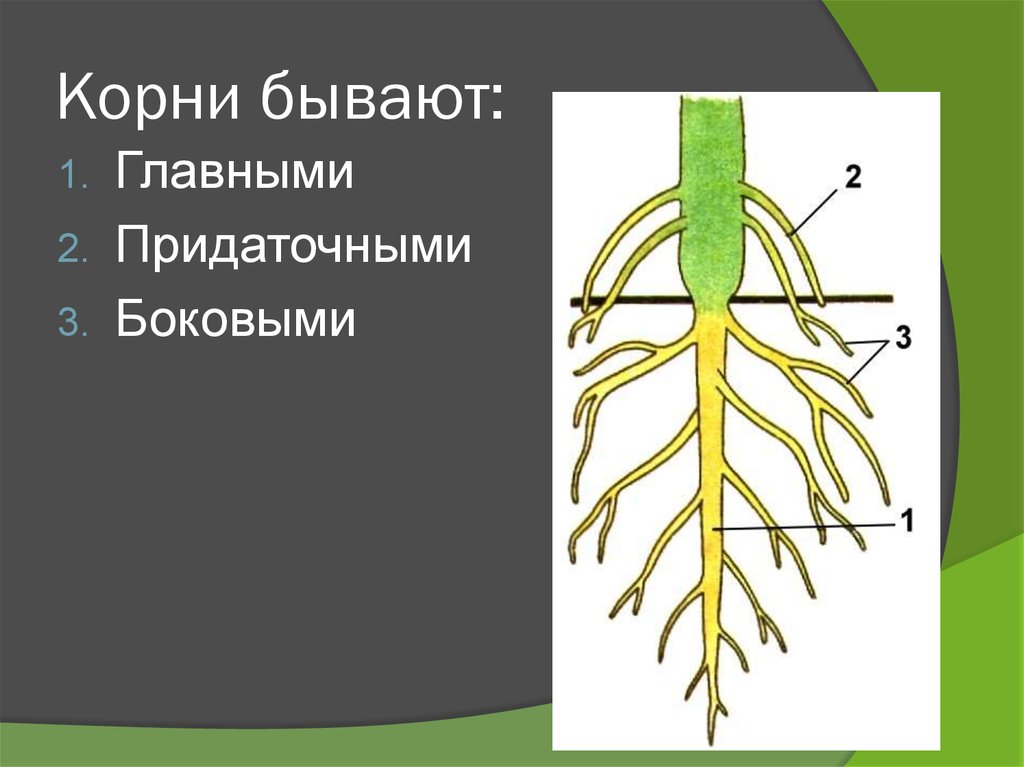 Гифы гриба оплетают корни деревьев и поставляют им минеральные элементы, а сами получают органические вещества. Корень дерева разветвляется, утолщая концы, но не образуя корневых волосков.
Гифы гриба оплетают корни деревьев и поставляют им минеральные элементы, а сами получают органические вещества. Корень дерева разветвляется, утолщая концы, но не образуя корневых волосков.
ВИДОИЗМЕНЕНИЯ КОРНЕЙ
Основными функциями корней является удерживание растения и закрепление его в почве, а также снабжение его минеральными солями и водой. Иногда у корня появляются дополнительные функции, происходит видоизменение корневища.
Когда корни начинают запасать питательные вещества и выступать их хранилищем, то главный корень сильно утолщается, разрастается. Это наиболее характерно для корнеплодов двулетних культур: моркови, свеклы, петрушки, сельдерея, цикория. Из однолетних растений корнеплод образует редис. Если начинают разрастаться боковые корни, то формируются клубни. Так запасает питательные вещества георгин, они помогают растению быстрее зацвести. Корневые клубни образует батат. При этом кроме клубней в корневой системе растения есть и обычные корни.
Лианы используют особые корни-прицепки, чтобы прикрепляться к стволам других растений, стенам. оплетая их, чтобы добраться до солнечных лучей. Плющ при помощи корней-зацепок способен оплетать любую опору. Омела своими корнями-присосками надежно закрепляется на ветвях деревьев, вытягивая из них питательные вещества и влагу. Такие же корни у цветка Иван да Марья, который растет на земле, но присасывается к другим растениям. У повилики нет листвы совсем, она обвивает своим стеблем «хозяина» и за счет видоизмененных корней в виде присосок вытягивает все соки из растения.
Обитатели тропиков — орхидеи отличаются наличием воздушных корней, которые не заглубляются в почву, они улавливают из воздуха влагу. Поэтому комнатные орхидеи так любят опрыскивание.
Ходульные корни – это придаточные корни, которые отрастают от ствола, а когда достигают земли, прорастают в нее. Они выступаю дополнительной опорой, поддерживают растения. У мангровых деревьев они кроме поддержания растения дополнительно снабжают его воздухом.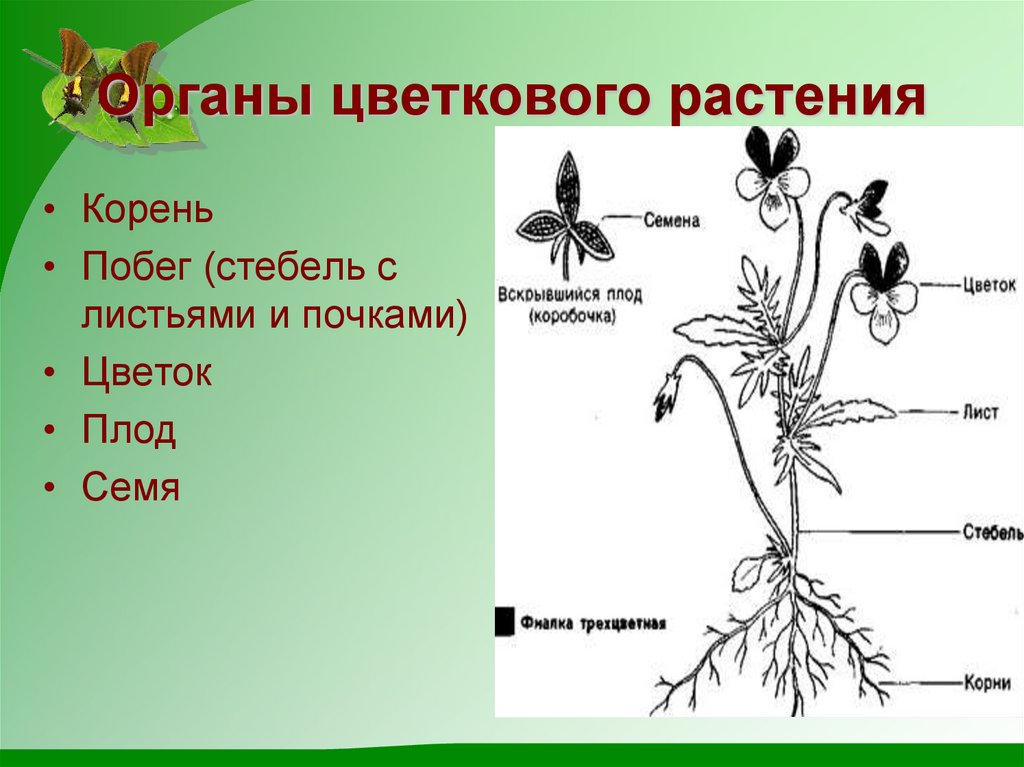 Это очень распространенное явления, когда корни одновременно воздушные и опорные. Похожие на диск они у каучуконосного фикуса, крепкие и похожие на столбы у баньяна. Дыхательные корни начинают образовываться у растений, когда им в почве не хватает кислорода. У кукурузы они напоминают ходули и помогают кукурузе противостоять ветру. Нередкое явление для растений, произрастающих в наводненной или заболоченной местности. Вырастают выше уровня поверхности почвы или воды. Образуются, чтобы дать дополнительный источник кислорода, которого в воде содержится недостаточно для полноценной жизни растения. Можно встретить у ивы ломкой, кипариса болотного.
Это очень распространенное явления, когда корни одновременно воздушные и опорные. Похожие на диск они у каучуконосного фикуса, крепкие и похожие на столбы у баньяна. Дыхательные корни начинают образовываться у растений, когда им в почве не хватает кислорода. У кукурузы они напоминают ходули и помогают кукурузе противостоять ветру. Нередкое явление для растений, произрастающих в наводненной или заболоченной местности. Вырастают выше уровня поверхности почвы или воды. Образуются, чтобы дать дополнительный источник кислорода, которого в воде содержится недостаточно для полноценной жизни растения. Можно встретить у ивы ломкой, кипариса болотного.
1.(1,5 балла). Корнеплод имеют:
а).Георгин б). Репа в). Турнепс г). Плющ д). Баньян е). Брюква.
2. (1 балл)Какую из перечисленных функций корни не выполняют
а) закрепление растений в почве
б) всасывание воды и минеральных солей из почвы
в) запасание питательных веществ
г) образование питательных веществ в растении
3. (1 балл) В результате утолщения боковых и придаточных корней образуются
(1 балл) В результате утолщения боковых и придаточных корней образуются
а) воздушные корни
б) корневые клубни
в) корнеплоды
г) корни-присоски
1.(1,5 балла). Корнеплод имеют:
а).Георгин б). Репа в). Турнепс г). Плющ д). Баньян е). Брюква.
2. (1 балл)Какую из перечисленных функций корни не выполняют
а) закрепление растений в почве
б) всасывание воды и минеральных солей из почвы
в) запасание питательных веществ
г) образование питательных веществ в растении
3. (1 балл) В результате утолщения боковых и придаточных корней образуются
а) воздушные корни
б) корневые клубни
в) корнеплоды
г) корни-присоски
1.(1,5 балла). Корнеплод имеют:
а).Георгин б). Репа в). Турнепс г). Плющ д). Баньян е). Брюква.
2. (1 балл)Какую из перечисленных функций корни не выполняют
а) закрепление растений в почве
б) всасывание воды и минеральных солей из почвы
в) запасание питательных веществ
г) образование питательных веществ в растении
3. (1 балл) В результате утолщения боковых и придаточных корней образуются
(1 балл) В результате утолщения боковых и придаточных корней образуются
а) воздушные корни
б) корневые клубни
в) корнеплоды
г) корни-присоски
1.(1,5 балла). Корнеплод имеют:
а).Георгин б). Репа в). Турнепс г). Плющ д). Баньян е). Брюква.
2. (1 балл)Какую из перечисленных функций корни не выполняют
а) закрепление растений в почве
б) всасывание воды и минеральных солей из почвы
в) запасание питательных веществ
г) образование питательных веществ в растении
3. (1 балл) В результате утолщения боковых и придаточных корней образуются
а) воздушные корни
б) корневые клубни
в) корнеплоды
г) корни-присоски
Анатомия растений
Эта страница создана Джимом Бидлаком, Университет Центральной Оклахомы, на основе оригинальной деятельности Пола Шульте, Университет Невады, Лас-Вегас, Джейн Элизабет Краус и Джулиана Писанески, Университет Сан-Паулу, и Джеймса Д.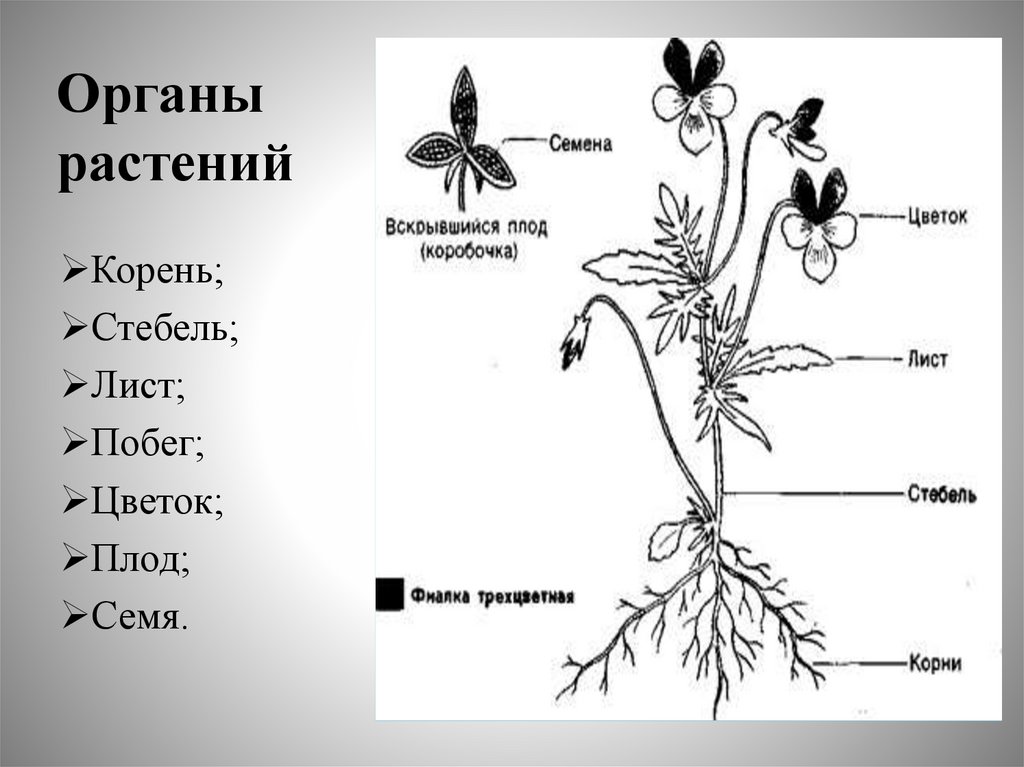 Мосета, Техасский университет.
Мосета, Техасский университет.
Профиль автора
Это задание было проверено двумя процессами проверки
Скрыть
Это задание прошло экспертную оценку.
Это задание прошло рецензирование, в ходе которого отправленные действия сравниваются с набором критериев. В коллекцию добавлены действия, соответствующие или измененные для соответствия этим критериям. Чтобы узнать больше о критериях проверки, см. [http://taste.merlot.org/evaluationcriteria.html]. Дополнительную информацию о процессе рецензирования можно найти по адресу [http://taste.merlot.org/peerreviewprocess.html].
Эта страница впервые опубликована: 17 августа 2010 г.
Данный материал тиражирован на ряде сайтов
как часть
SERC Pedagogic Service Project
Резюме
Обсуждение вместе с рисунками, образцами и визуализациями помогает участникам понять различия между двудольными и однодольными растениями, особенно в корнях, стеблях и листьях.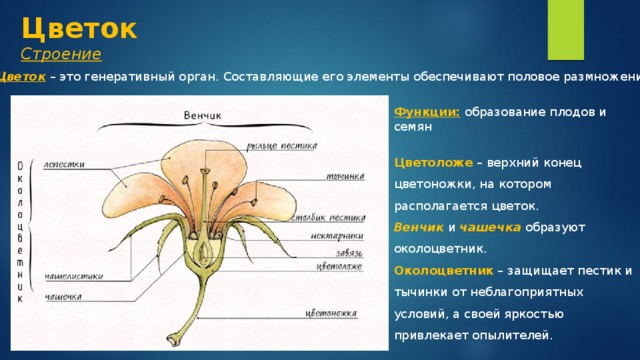
Использовали это занятие? Поделитесь своим опытом и изменениями
Цели обучения
- Поймите, что анатомия растений — это больше, чем просто корни, стебли, листья и цветы, это скорее изучение внутренних тканей и структур внутри органов.
- Знайте общие различия между двудольными и однодольными растениями.
- Нарисуйте и подпишите поперечные сечения корней двудольных и однодольных растений.
- Нарисуйте и подпишите поперечные сечения стеблей двудольных и однодольных растений.
- Нарисуйте и подпишите поперечные сечения листьев двудольных и однодольных растений.
Контекст для использования
Эта стратегия обучения анатомии растений представляет собой двухчасовую презентацию с рисунками, образцами и визуализациями для изучения корней, стеблей и листьев двудольных и однодольных растений.
Описание и учебные материалы
- Расскажите об анатомии растений, объяснив, что этот предмет включает в себя не только корни, стебли, листья и цветы, но и внутренние ткани и структуры этих органов растений. В этом учебном материале сосредоточьтесь на корнях, стеблях и листьях.
- Попросите участников предположить, какие типы тканей и структур находятся в корнях, стеблях и листьях. Раздайте образцы корней моркови, люцерны или других стеблей и листьев шпината. Укажите основные функции этих органов (корни – поглощение воды и питательных веществ; стебли – проведение питательных веществ и связь с другими органами; листья – фотосинтез).
- Начав с корня моркови, попросите участников изучить образец невооруженным глазом и начать обсуждение того, какие ткани обнаружены от внешней части до внутренней части моркови.
- Познакомить с терминами эпидермис, кора, эндодерма и стела (с ксилемой и флоэмой). Нарисуйте поперечный разрез корня и обозначьте эти ткани. Обратите внимание, что это поперечное сечение корня двудольных растений. См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial. htm?id=80115 для получения дополнительной информации. подробные визуализации.
- Объясните, чем корни однодольных отличаются от корней двудольных, и нарисуйте поперечный разрез корня однодольного растения и обозначьте эпидермис, кору, эндодерму и стелу (с ксилемой и флоэмой). См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации.
- Используйте люцерну или другой стебель в качестве примера двудольного растения и попросите участников изучить образец невооруженным глазом и обсудить, какие ткани находятся от внешней части стебля до внутренней части стебля.
- Нарисуйте поперечное сечение стебля двудольного растения. См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации.
- Нарисуйте поперечное сечение стебля однодольного растения. См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации.
- Используйте лист шпината в качестве примера двудольного растения и попросите участников изучить образец невооруженным глазом и обсудить, какие ткани находятся в очень тонком слое листа.
- Нарисуйте лист двудольного растения в поперечном сечении. См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации.
- Нарисуйте поперечный разрез листа однодольного растения. См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации.
- Кратко изложите эту тему анатомии растений, в которой рассматриваются корни, стебли и листья. Предложите участникам подумать о том, какую часть растения они едят и какие ткани потребляют в следующий раз, когда они будут за обеденным столом.
 В этом учебном материале сосредоточьтесь на корнях, стеблях и листьях.
В этом учебном материале сосредоточьтесь на корнях, стеблях и листьях.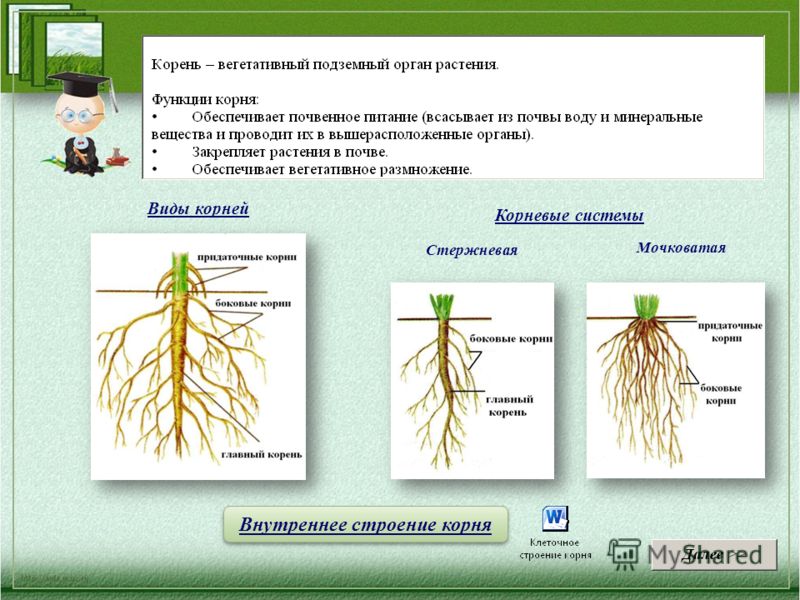 htm?id=80115 для получения дополнительной информации. подробные визуализации.
htm?id=80115 для получения дополнительной информации. подробные визуализации.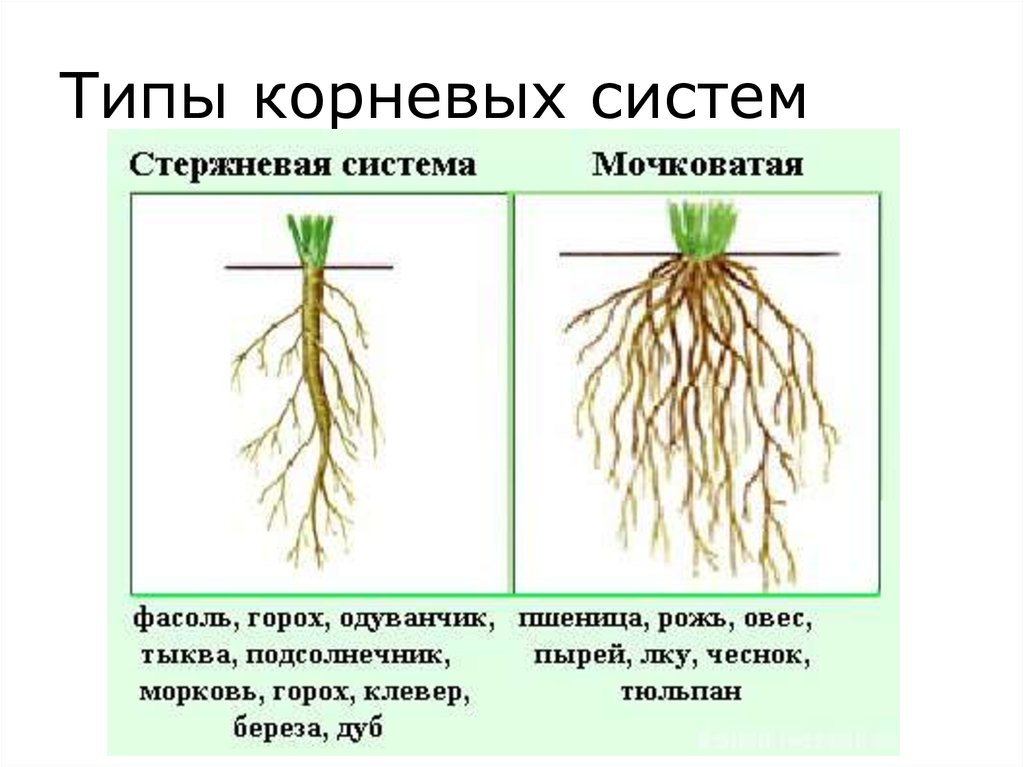 См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации.
См. http://www.merlot.org/merlot/viewMaterial.htm?id=80106 для упрощенной версии этого поперечного сечения и http://www.merlot.org/merlot/viewMaterial.htm?id=80115 для более подробные визуализации. Предложите участникам подумать о том, какую часть растения они едят и какие ткани потребляют в следующий раз, когда они будут за обеденным столом.
Предложите участникам подумать о том, какую часть растения они едят и какие ткани потребляют в следующий раз, когда они будут за обеденным столом.Вспомогательные файлы:
- Outline — Plant Anatomy (Microsoft Word 37kB Aug12 10)
- Вопросы — Анатомия растений (Microsoft Word, 33kB, 12 августа, 10)
Учебные заметки и советы
Эта стратегия обучения предусматривает обсуждение, а также образцы и визуализации для изучения анатомии двудольных и однодольных растений. Участники узнают о поперечных срезах корней, стеблей и листьев, чтобы лучше понять ткани, обнаруженные в этих органах, их организацию и различия, наблюдаемые в каждом из них. Включение образцов и визуализаций поддерживает интерес учащихся к теме. Когда участники рисуют структуры, которые они действительно могут видеть и держать, это помогает им оставаться активными в процессе обучения.
Оценка
Участники могут быть проверены на понимание этого учебного материала с помощью экзаменов с несколькими вариантами ответов, кратких ответов или эссе.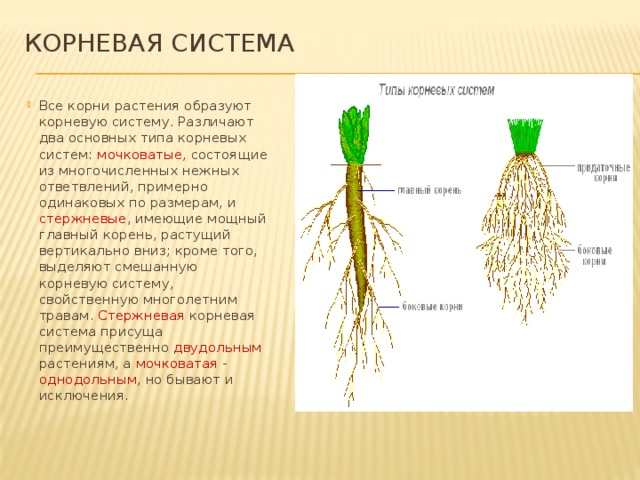 Несколько примеров вопросов включены в этот рабочий лист под названием «Вопросы — анатомия растений».
Несколько примеров вопросов включены в этот рабочий лист под названием «Вопросы — анатомия растений».
Ссылки и ресурсы
Описание MERLOT и ссылка на «Атлас анатомии растений», в котором представлены хорошие микрофотографии со слайдов структур растений. См. http://www.merlot.org/merlot/viewMaterial.htm?id=80115
Описание MERLOT и ссылка на «Атлас анатомии растений», который представляет собой очень простой и понятный сайт, на котором представлены микрофотографии корней, стеблей и листьев двудольных и однодольных растений. См. описание http://www.merlot.org/merlot/viewMaterial.htm?id=80106
MERLOT и ссылку на «Лабораторию анатомии растений», веб-сайт, сопровождающий учебник по анатомии растений. Микрофотографии типов клеток и тканей можно найти на этом сайте. См. http://www.merlot.org/merlot/viewMaterial.htm?id=76843
Подробнее Обучение с помощью визуализаций »
Гипотеза «корневого мозга» Чарльза и Фрэнсиса Дарвинов
1. Балушка Ф., Главацка А., Манкузо С., Барлоу П.В. Нейробиологический взгляд на растения и их строение тела. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений: нейронные аспекты жизни растений. Берлин-Гейдельберг-Нью-Йорк: Springer Verlag; 2006. С. 19–35. [Google Scholar]
Балушка Ф., Главацка А., Манкузо С., Барлоу П.В. Нейробиологический взгляд на растения и их строение тела. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений: нейронные аспекты жизни растений. Берлин-Гейдельберг-Нью-Йорк: Springer Verlag; 2006. С. 19–35. [Google Scholar]
2. Бреннер Э.Д., Штальберг Р., Манкузо С., Виванко Дж., Балушка Ф., Ван Волкенбург Э. Нейробиология растений: интегрированный взгляд на сигнализацию растений. Тенденции Растениевод. 2006; 11: 413–419.. [PubMed] [Google Scholar]
3. Кальво Гарсон Ф., Кейзер Ф. Познание у растений. В: Балушка Ф, редактор. Взаимодействие растений и окружающей среды с поведенческой точки зрения. Берлин; Гейдельберг; Нью-Йорк: Springer Verlag; 2009. стр. 247–266. [Google Scholar]
4. Тревавас А. Интеллект растений. Натурвиссеншафтен. 2005; 92: 401–413. [PubMed] [Google Scholar]
5. Тревавас А. Ответ Альпи и др.: Нейробиология растений — все метафоры имеют значение. Тенденции Растениевод. 2007; 12: 231–233. [PubMed] [Академия Google]
2007; 12: 231–233. [PubMed] [Академия Google]
6. Балушка Ф., Манкузо С. Нейробиология растений: от восприятия стимулов до адаптивного поведения растений с помощью интегрированной химической и электрической сигнализации. Поведение сигналов растений. 2009; 4: 475–476. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Alpi A, Amrhein N, Bertl A, Blatt MR, Blumwald E, Cervone F, et al. Нейробиология растений: нет мозга — нет прибыли? Тенденции Растениевод. 2007; 12: 135–136. [PubMed] [Google Scholar]
8. Tapken D, Hollmann M. Arabidopsis thaliana Функция ионных каналов рецептора глутамата продемонстрирована трансплантацией ионных пор. Дж Мол Биол. 2008; 383:36–48. [PubMed] [Академия Google]
9. Masi E, Ciszak M, Stefano G, Renna L, Azzarello E, Pandolfi C, et al. Пространственно-временная динамика активности электрической сети в верхушке корня: исследование многоэлектродной решетки (МЭА). Proc Natl Acad Sci USA. 2008; 106:4048–4053. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Karban R, Shiojiri K. Самопознание влияет на коммуникацию и защиту растений. Эколь латыши. 2009; 12: 502–506. [PubMed] [Google Scholar]
Karban R, Shiojiri K. Самопознание влияет на коммуникацию и защиту растений. Эколь латыши. 2009; 12: 502–506. [PubMed] [Google Scholar]
11. Biedrzycki ML, Jilany TA, Dudley SA, Bais HP. Корневые экссудаты опосредуют узнавание родства у растений. Коммун Интегр Биол. 2010;3 В печати. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Тревавас А. Интеллект растений. Натурвиссеншафтен. 2005; 92: 401–413. [PubMed] [Google Scholar]
13. Тревавас А. Что такое поведение растений? Окружающая среда растительной клетки. 2009; 32: 606–616. [PubMed] [Google Scholar]
14. Айрес П. Живые растения: Дарвины на заре наук о растениях. Лондон: Пикеринг и Чатто; 2008. [Google Scholar]
15. Kutschera U, Briggs WR. От ботанических исследований Чарльза Дарвина в загородных домах до современной физиологии растений. биол. растений 2009 г.;11:785–795. [PubMed] [Google Scholar]
16. Kutschera U, Nicklas KJ. Эволюционная физиология растений: забытый синтез Чарльза Дарвина.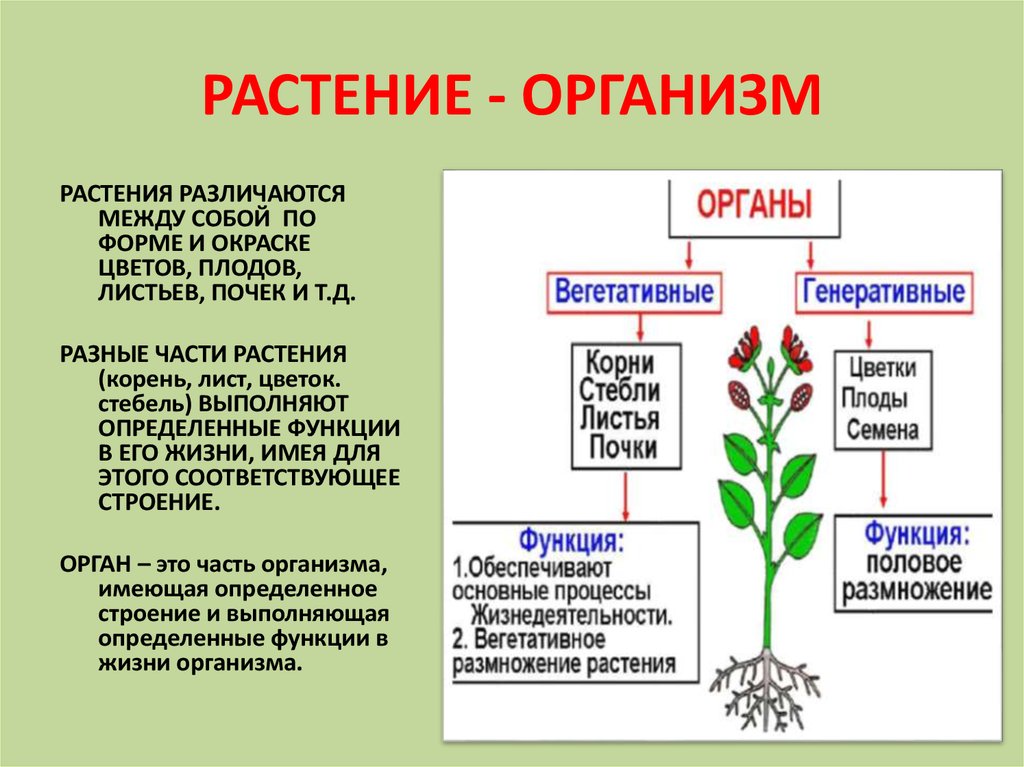 Натурвиссеншафтен. 2009; 96: 1339–1354. [PubMed] [Google Scholar]
Натурвиссеншафтен. 2009; 96: 1339–1354. [PubMed] [Google Scholar]
17. Darwin CR. Сила движений в растениях. Лондон: Джон Мюррей; 1880. (http://darwin-online.org.uk/) [Google Scholar]
18. Пенни Д. Чарльз Дарвин как биолог-теоретик в механистической традиции. Тенденции Эвол Биол. 2009;1:e1. [Академия Google]
19. Барлоу П.В. Чарльз Дарвин и верхушка корня растения: ликвидация пробела в теории живых систем применительно к растениям. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений: нейронные аспекты жизни растений. Берлин-Гейдельберг-Нью-Йорк: Springer; 2006. С. 37–51. [Google Scholar]
20. Хеслоп Харрисон Дж. Дарвин и движение растений: ретроспектива. В: Скуг Ф., редактор. Вещества для роста растений 1979. 1980. стр. 3–14. [Google Scholar]
21. Балушка Ф., Манкузо С. Растения и животные: конвергентная эволюция в действии? В: Балушка Ф, редактор. Взаимодействие растений и окружающей среды с поведенческой точки зрения. Берлин-Гейдельберг-Нью-Йорк: Springer Verlag; 2009 г. . стр. 285–301. [Google Scholar]
. стр. 285–301. [Google Scholar]
22. Балушка Ф., Шлихт М., Ван Ю.-Л., Бурбах С., Фолькманн Д. Внутриклеточные домены и полярность в верхушках корней: от синаптических доменов до нейробиологии растений. Нова Акта Леопольд. 2009;96:103–122. [Google Scholar]
23. Барлоу П.В. Регенерация чехлика первичных корней Zea mays . Новый Фитол. 1974; 73: 937–954. [Google Scholar]
24. Балушка Ф., Манкузо С., Фолькманн Д., Барлоу П.В. Верхушки корней как командные центры растений: уникальный «мозгоподобный» статус переходной зоны верхушек корней. Биология. 2004; 59(Приложение 13): 9–17. [Google Scholar]
25. Baluška F, Kubica Š, Hauskrecht M. Постмитотический «изодиаметрический» рост клеток в верхушке корня кукурузы. Планта. 1990; 181: 269–274. [PubMed] [Google Scholar]
26. Балушка Ф., Барлоу П.В., Кубица Ш. Важность области постмитотического «изодиаметрического» роста (PIG) для роста и развития корней. Растение и почва. 1994; 167:31–42. [Google Scholar]
27. Балушка Ф., Фолькманн Д., Барлоу П.В. Специализированные зоны развития в корнях: взгляд с клеточного уровня. Завод Физиол. 112:3–4. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Балушка Ф., Фолькманн Д., Барлоу П.В. Специализированные зоны развития в корнях: взгляд с клеточного уровня. Завод Физиол. 112:3–4. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Исикава Х., Эванс М.Л. Индукция искривления корней кукурузы кальцием или тигмостимуляцией: роль постмитотической изодиаметрической зоны роста. Завод Физиол. 1993; 100:762–768. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Исикава Х., Эванс М.Л. Роль дистальной зоны растяжения в реакции корней кукурузы на ауксин и гравитацию. Завод Физиол. 1993; 102:1203–1210. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Балушка Ф., Вита С., Барлоу П.В., Фолькманн Д. Перестройки массивов F-актина в растущих клетках интактных тканей верхушки корня кукурузы: основное переключение в развитии происходит в область постмитотического перехода. Eur J Cell Biol. 1997;72:113–121. [PubMed] [Google Scholar]
31. Балушка Ф., Фолькманн Д., Барлоу П.В. Перекресток полярности в переходной зоне роста верхушек корней кукурузы: последствия для цитоскелета и развития. J Регулятор роста растений. 2001; 20: 170–181. [Google Scholar]
J Регулятор роста растений. 2001; 20: 170–181. [Google Scholar]
32. De Smet I, Tetsumura T, De Rybel B, Frey NF, Laplaze L, Casimiro I, et al. Ауксин-зависимая регуляция положения боковых корней в базальной меристеме арабидопсиса. Разработка. 2007; 134: 681–690. [PubMed] [Академия Google]
33. Де Смет И., Юргенс Г. Паттернирование оси у растений — контроль над ауксином. Curr Opin Genet Dev. 2007; 17: 337–343. [PubMed] [Google Scholar]
34. Перет Б., Де Рибель Б., Касимиро И., Бенкова Э., Сваруп Р., Лаплаз Л. и др. Развитие боковых корней арабидопсиса: новая история. Тенденции Растениевод. 2009; 14: 399–408. [PubMed] [Google Scholar]
35. Verbelen J-P, De Cnodder T, Le J, Vissenberg K, Baluška F. Верхушка корня Arabidopsis thaliana состоит из четырех отдельных зон клеточной активности: меристематическая зона, переходная зона, зона быстрого удлинения и зона прекращения роста. Поведение сигналов растений. 2006;1:296–304. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Baluška F, Hlavacka A. Растительные формины стареют: что-то особенное в поперечных стенах. Новый Фитол. 2005; 168: 499–503. [PubMed] [Google Scholar]
Baluška F, Hlavacka A. Растительные формины стареют: что-то особенное в поперечных стенах. Новый Фитол. 2005; 168: 499–503. [PubMed] [Google Scholar]
37. Фолькманн Д., Балушка Ф. Актиновый цитоскелет растений: от транспортных сетей к сигнальным сетям. Микроск Рес Тех. 1999; 47: 135–154. [PubMed] [Google Scholar]
38. Балушка Ф., Барлоу П.В., Фолькманн Д. Актин и миозин VIII в развивающихся клетках корня. В: Staiger CJ, Baluška F, Volkmann D, Barlow PW, редакторы. Актин: динамическая структура для нескольких функций растительных клеток. Дордрехт, Нидерланды: Kluwer Academic Publishers; 2000. С. 457–476. [Академия Google]
39. Балушка Ф., Фолькманн Д., Барлоу П.В. Основанные на актине домены «комплекса периферии клетки» и их ассоциации с поляризованными «телами клеток» у высших растений. биол. растений 2000; 2: 253–267. [Google Scholar]
40. Барлоу П.В., Балушка Ф. Цитоскелетный взгляд на рост и морфогенез корней. Annu Rev Plant Physiol Plant Mol Biol. 2000; 51: 289–322. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
41. Балушка Ф., Войташек П., Фолькманн Д., Барлоу П.В. Архитектура роста поляризованных клеток: уникальный статус удлиняющихся растительных клеток. Биоэссе. 2003;25:569–576. [PubMed] [Google Scholar]
42. Небенфюр А., Ритценталер С., Робинсон Д.Г. Брефельдин А: расшифровка загадочного ингибитора секреции. Завод Физиол. 2002; 130:1102–1108. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Baluška F, Hlavacka A, Šamaj J, Palme K, Robinson DG, Matoh T, et al. F-актин-зависимый эндоцитоз пектинов клеточной стенки в меристематических клетках корня: информация из компартментов, индуцированных брефельдином А. Завод Физиол. 2002; 130:422–431. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Hause G, Šamaj J, Menzel D, Baluška F. Тонкий структурный анализ формирования компартмента, вызванного брефельдином А, после фиксации замораживанием эпидермиса корня кукурузы под высоким давлением: сложный экзоцитоз, напоминающий образование клеточной пластинки во время цитокинеза.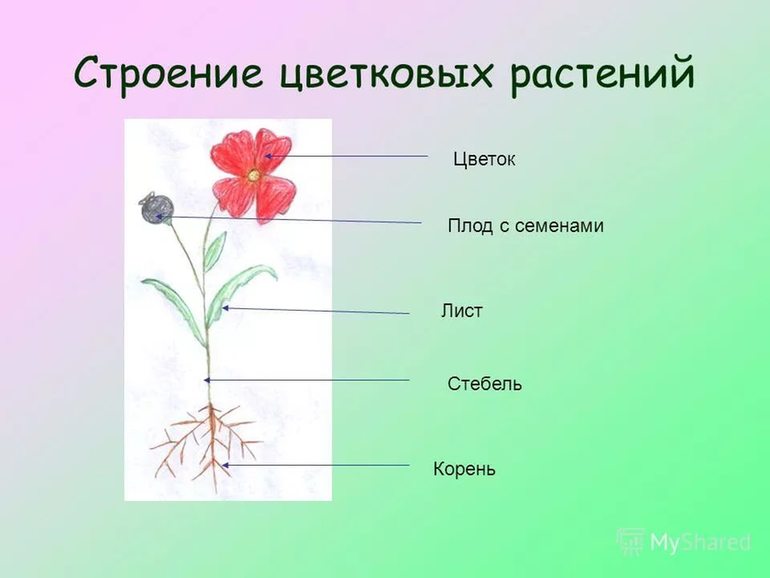 Поведение сигналов растений. 2006; 1: 134–139. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Поведение сигналов растений. 2006; 1: 134–139. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Robinson DG, Langhans M, Saint-Jore-Dupas C, Hawes C. Эффекты BFA зависят от ткани, а не только от растения. Тенденции Растениевод. 2008; 13: 405–408. [PubMed] [Академия Google]
46. Baluška F, Šamaj J, Wojtaszek P, Volkmann D, Menzel D. Континуум цитоскелет-плазматическая мембрана-клеточная стенка у растений: новый взгляд на возникающие связи. Завод Физиол. 2003; 133:482–491. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Гельднер Н., Фримл Дж., Штирхоф Ю.Д., Юргенс Г., Пальме К. Ингибиторы транспорта ауксина блокируют циклирование PIN1 и транспортировку везикул. Природа. 2001; 413:425–428. [PubMed] [Google Scholar]
48. Тил В. Д., Папонов И. А., Пальме К. Ауксин в действии: сигнализация, транспорт и контроль роста и развития растений. Nat Rev Mol Cell Biol. 2006; 7: 847–859.. [PubMed] [Google Scholar]
49. Пальме К., Довженко А., Дитенгу Ф.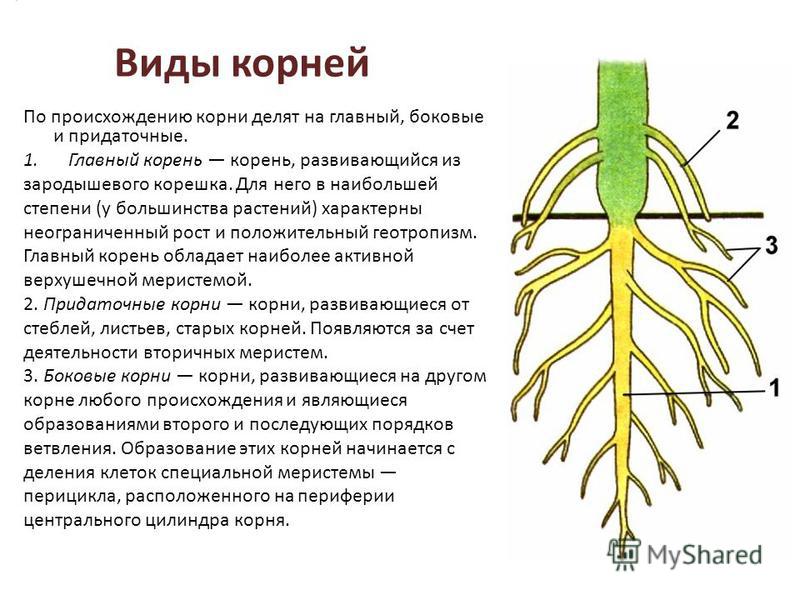 А. Транспорт ауксинов и гравитационные исследования: перспективы. Протоплазма. 2006; 229:175–181. [PubMed] [Google Scholar]
А. Транспорт ауксинов и гравитационные исследования: перспективы. Протоплазма. 2006; 229:175–181. [PubMed] [Google Scholar]
50. Kleine-Vehn J, Friml J. Полярное нацеливание и рециркуляция эндоцитов в развитии ауксин-зависимых растений. Annu Rev Cell Dev Biol. 2008; 24:447–473. [PubMed] [Google Scholar]
51. Петрасек Дж., Фримл Дж. Транспортные пути ауксина в развитии растений. Разработка. 2009; 136: 2675–2688. [PubMed] [Академия Google]
52. Дхонукше П., Григорьев И., Фишер Р., Томинага М., Робинсон Д.Г., Хасек Дж. и соавт. Ингибиторы транспорта ауксина нарушают подвижность пузырьков и динамику актинового цитоскелета у различных эукариот. Proc Natl Acad Sci USA. 2008; 105:4489–4494. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Манкузо С., Маррас А.М., Волкер М., Балушка Ф. Неинвазивные и непрерывные записи потоков ауксина в интактной верхушке корня с модифицированной углеродной нанотрубкой и самореферентной микроэлектрод. Анальная биохимия. 2005; 341:344–351.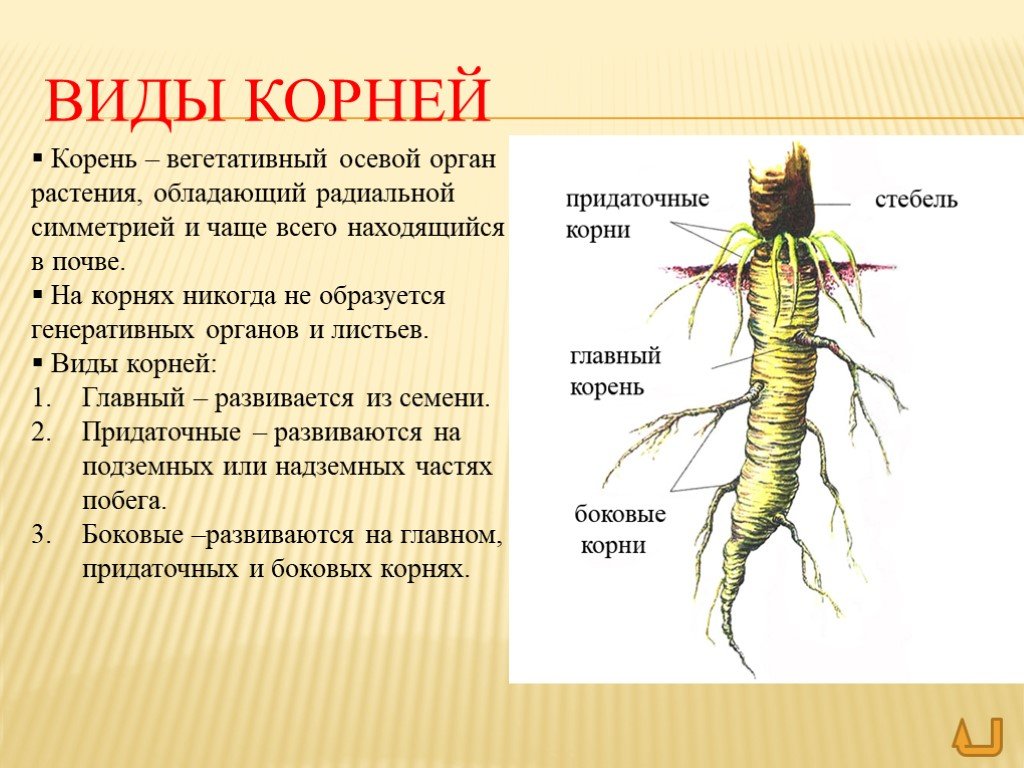 [PubMed] [Академия Google]
[PubMed] [Академия Google]
54. Schlicht M, Strnad M, Scanlon MJ, Mancuso S, Hochholdinger F, Palme K, et al. Иммунолокализация ауксина подразумевает везикулярный нейротрансмиттерный способ полярного транспорта ауксина в верхушках корней. Поведение сигналов растений. 2006; 1: 122–133. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Mancuso S, Marras AM, Mugnai S, Schlicht M, Zarsky V, Li G, et al. Фосфолипаза Dξ2 управляет везикулярной секрецией ауксина для его полярного межклеточного транспорта в переходной зоне верхушки корня. Поведение сигналов растений. 2007; 2: 240–244. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Уилкинсон С., Моррис Д.А. Нацеливание переносчиков ауксина на плазматическую мембрану: влияние монензина на трансмембранный транспорт ауксина в ткани Cucurbita pepo L.. Планта. 1994; 193:194–202. [Google Scholar]
57. Фудзи К., Мива К., Фудзивара Т. Внутриклеточный транспорт переносчиков: мембранный транспорт переносчиков минералов.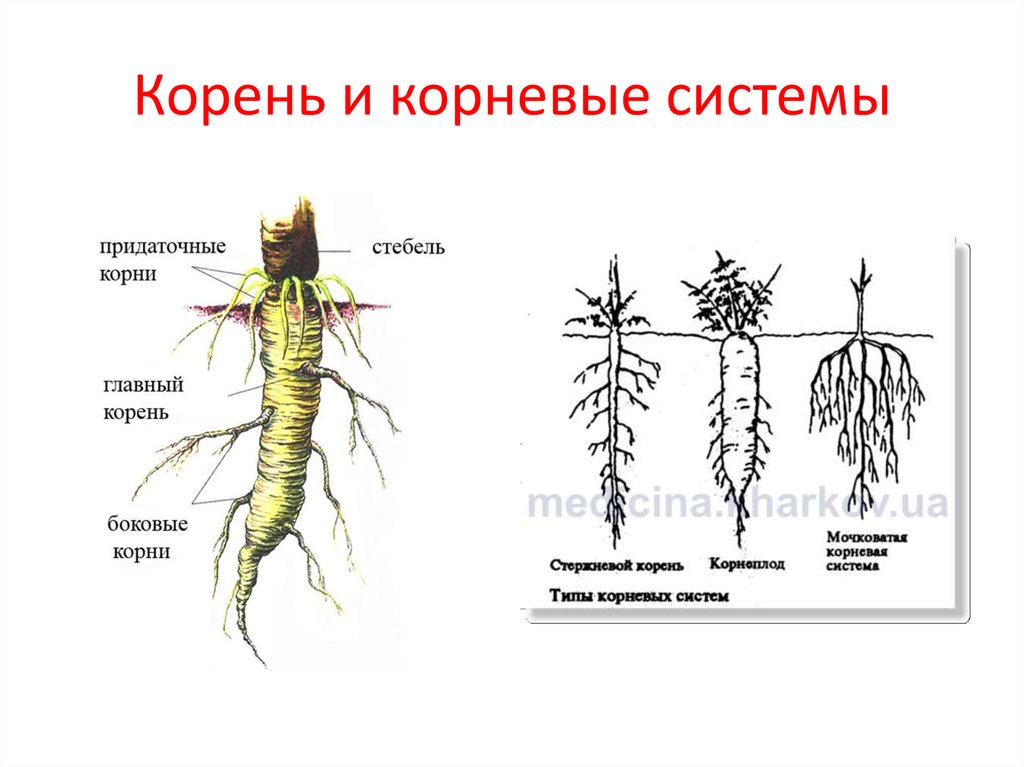 Curr Opin Plant Biol. 2009;12 В печати. [PubMed] [Google Scholar]
Curr Opin Plant Biol. 2009;12 В печати. [PubMed] [Google Scholar]
58. Baluška F, Liners F, Hlavacka A, Schlicht M, Van Cutsem P, McCurdy D, et al. Пектины клеточной стенки и ксилоглюканы интернализуются в делящиеся клетки корня и накапливаются внутри клеточных пластинок во время цитокинеза. Протоплазма. 2005; 225:141–155. [PubMed] [Академия Google]
59. Дхонукше П., Балушка Ф., Шлихт М., Главацка А., Шамай Дж., Фримл Дж. и др. Эндоцитоз материала клеточной поверхности опосредует образование клеточной пластинки во время цитокинеза растений. Ячейка Дев. 2006; 10: 137–150. [PubMed] [Google Scholar]
60. Шамай Дж., Балушка Ф., Фойгт Б., Шлихт М., Фолькманн Д., Менцель Д. Эндоцитоз, актиновый цитоскелет и передача сигналов. Завод Физиол. 2004; 135:1150–1161. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Шамай Дж., Рид Н.Д., Фолькманн Д., Мензель Д., Балушка Ф. Эндоцитарная сеть у растений. Тенденции клеточной биологии. 2005; 15: 425–433. [PubMed] [Академия Google]
62. Балушка Ф., Фолькманн Д., Менцель Д. Синапсы растений: домены адгезии на основе актина для межклеточной коммуникации. Тенденции Растениевод. 2005; 10:106–111. [PubMed] [Google Scholar]
Балушка Ф., Фолькманн Д., Менцель Д. Синапсы растений: домены адгезии на основе актина для межклеточной коммуникации. Тенденции Растениевод. 2005; 10:106–111. [PubMed] [Google Scholar]
63. Shen H, Hou NY, Schlicht M, Wan Y, Mancuso S, Baluška F. Токсичность алюминия нацелена на PIN2 в верхушках корней арабидопсиса: влияние на эндоцитоз PIN2, везикулярную рециркуляцию и транспорт полярного ауксина. Чин Научный Бык. 2008; 53: 2480–2487. [Google Scholar]
64. Балушка Ф., Шлихт М., Фолькманн Д., Манкузо С. Везикулярная секреция ауксина: доказательства и последствия. Поведение сигналов растений. 2008; 3: 254–256. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Ли Г., Сюэ Х.В. PLDzeta2 Arabidopsis регулирует транспортировку везикул и необходим для реакции ауксина. Растительная клетка. 2007; 19: 281–295. [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Yang X, Song L, Xue HW. Мембранный стероид-связывающий белок 1 (MSBP1) стимулирует тропизм, регулируя перенос везикул и перераспределение ауксина. Мол завод. 2008; 1:1077–1088. [PubMed] [Google Scholar]
Мол завод. 2008; 1:1077–1088. [PubMed] [Google Scholar]
67. Ван И, Линь ВХ, Чен Х, Сюэ ХВ. Роль 5PTase13 арабидопсиса в корневом гравитропизме посредством модуляции транспорта везикул. Сотовый рез. 2009 г.;19:1191–1204. [PubMed] [Google Scholar]
68. Балушка Ф., Барлоу П.В., Фолькманн Д., Манкузо С. Парадоксы растений, связанные с гравитацией: нейробиология растений предоставляет средства для их разрешения. В: Witzany G, редактор. Биосемиотика в междисциплинарном контексте, Материалы собрания по биосемиотике 6, Зальцбург, 2006 г. Хельсинки: Umweb; 2007. С. 9–35. [Google Scholar]
69. Siechen S, Yang S, Chiba A, Saif T. Механическое напряжение способствует скоплению нейротрансмиттерных везикул на пресинаптических окончаниях. Proc Natl Acad Sci USA. 2009 г.;106:12611–12616. [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Wolverton C, Mullen JL, Ishikawa H, Evans ML. Корневой гравитропизм в ответ на сигнал, исходящий за пределами шапки. Планта. 2002: 153–157.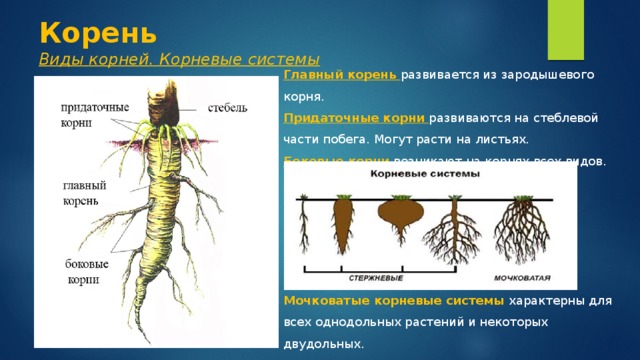 [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
71. Манкузо С., Барлоу П.В., Фолькманн Д., Балушка Ф. Реакция силы тяжести, опосредованная оборотом актина, в верхушках корней кукурузы: гравитропизм корней без колпачка подразумевает грависенсор вне корневого чехлика. Поведение сигналов растений. 2006; 1:52–58. [Бесплатная статья PMC] [PubMed] [Google Scholar]
72. Staves MP, Wayne R, Leopold AC. Влияние внешней среды на гравитропную кривизну корней риса ( Oryza sativa , Poaceae). Эм Джей Бот. 1997; 84: 1522–1529. [PubMed] [Google Scholar]
73. Wayne R, Staves MP, Leopold AC. Вклад внеклеточного матрикса в грависенсинг в клетках харовых. Дж. Клеточные науки. 1992; 101: 611–623. [PubMed] [Google Scholar]
74. Staves MP. Цитоплазматический поток и гравитационное зондирование в межузловых клетках Chara. Планта. 1997;203:79–84. [PubMed] [Google Scholar]
75. Nemec B. Über die Art der Wahrnehmung des Schwerkraftreizes bei den Pflanzen. Бер дтч бот Гэс. 1900; 18: 241–245.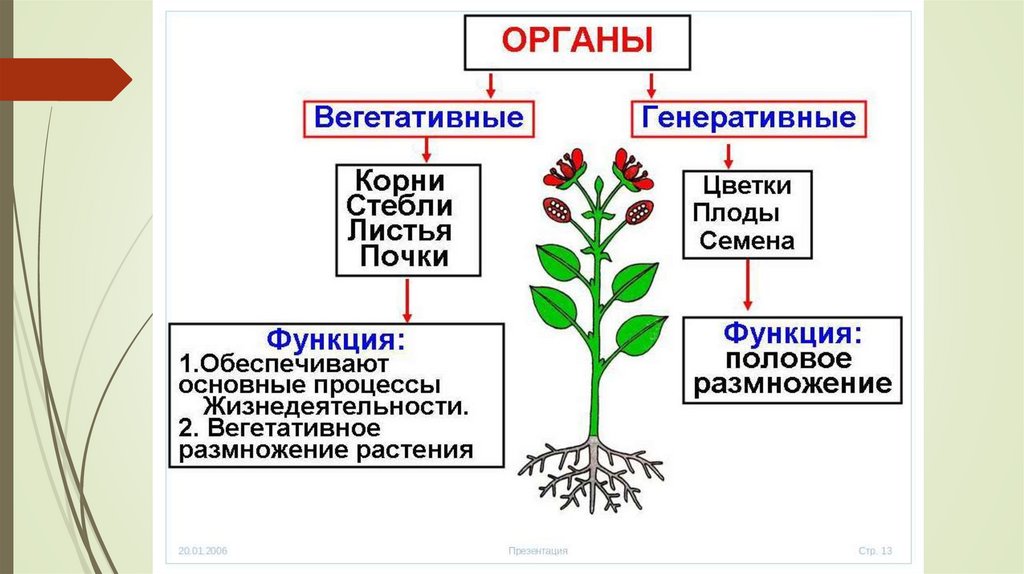 (Гер). [Google Scholar]
(Гер). [Google Scholar]
76. Сак Ф. Пластиды и гравитропное зондирование. Планта. 1997; 203: 63–68. [PubMed] [Google Scholar]
77. Friml J, Wisniewska J, Benková E, Mendgen K, Palme K. Боковое перемещение регулятора оттока ауксина PIN3 опосредует тропизм у арабидопсиса. Природа. 2002; 415:806–809. [PubMed] [Академия Google]
78. Гилрой С. Тропизмы растений. Карр Биол. 2008; 18: 275–277. [PubMed] [Google Scholar]
79. Ходжес А. Корневые решения. Окружающая среда растительной клетки. 2009; 32: 628–640. [PubMed] [Google Scholar]
80. Монсхаузен Г.Б., Гилрой С. Изучение реакции корневого роста на местные условия окружающей среды. Curr Opin Plant Biol. 2009; 12: 766–772. [PubMed] [Google Scholar]
81. Whippo CW, Hangarter RP. Фототропизм: стремление к просветлению. Растительная клетка. 2006; 18:1110–1119. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Балушка Ф., Манкузо С., Фолькманн Д. Коммуникация у растений: нейронные аспекты жизни растений.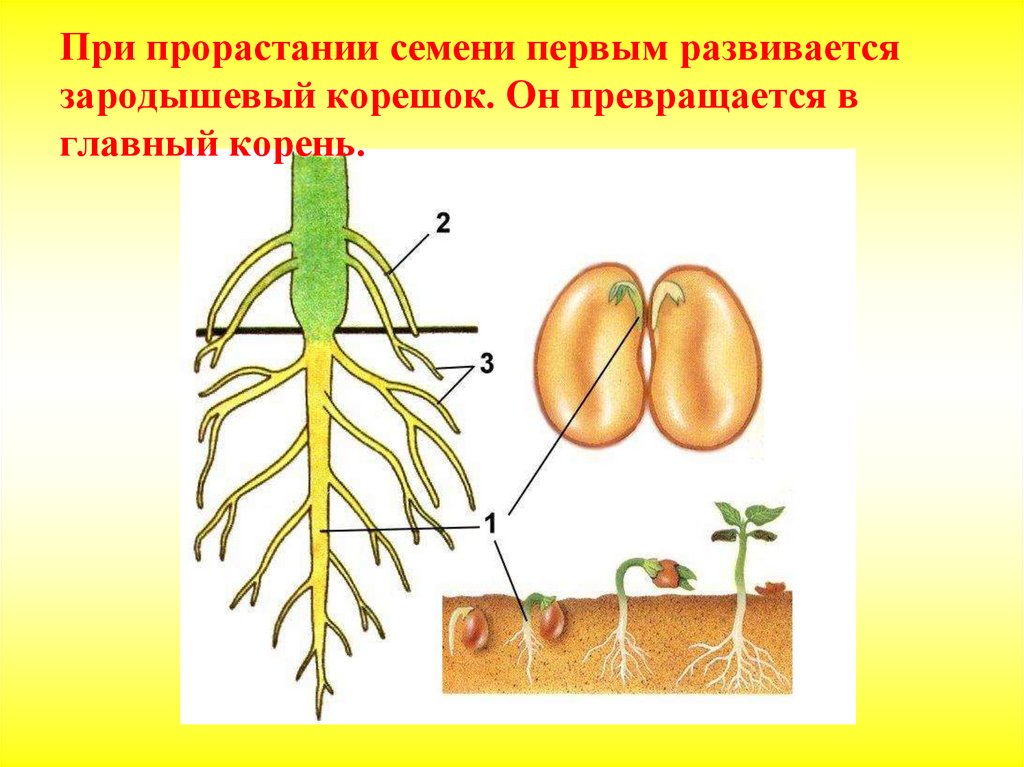 Берлин; Гейдельберг; Нью-Йорк: Springer Verlag; 2006. [Google Scholar]
Берлин; Гейдельберг; Нью-Йорк: Springer Verlag; 2006. [Google Scholar]
83. Хименес-Гомес Дж.М., Малуф Дж.Н. Исследования растений ускоряются на супермагистрали (био)информатики: симпозиум по восприятию растений, реакции и адаптации к окружающей среде. EMBO Rep. 2009; 10: 568–572. [Бесплатная статья PMC] [PubMed] [Google Scholar]
84. Wolverton C, Ishikawa H, Evans ML. Кинетика корневого гравитропизма: двойные моторы и датчики. J Регулятор роста растений. 2002; 21: 102–112. [PubMed] [Академия Google]
85. Эдельманн Х.Г., Рот У. Гравитропная регуляция роста растений и этилен: неожиданная кардинальная координата для вышедшей из употребления модели. Протоплазма. 2006; 229:183–191. [PubMed] [Google Scholar]
86. Хан А., Фирн Р., Эдельманн Х.Г. Взаимодействующие цепи передачи сигнала в корнях кукурузы, стимулированных гравитацией. Сигнальный преобразователь. 2006; 6: 449–455. [Google Scholar]
87. Хан А., Циммерманн Р., Ванке Д., Хартер К., Эдельманн Х.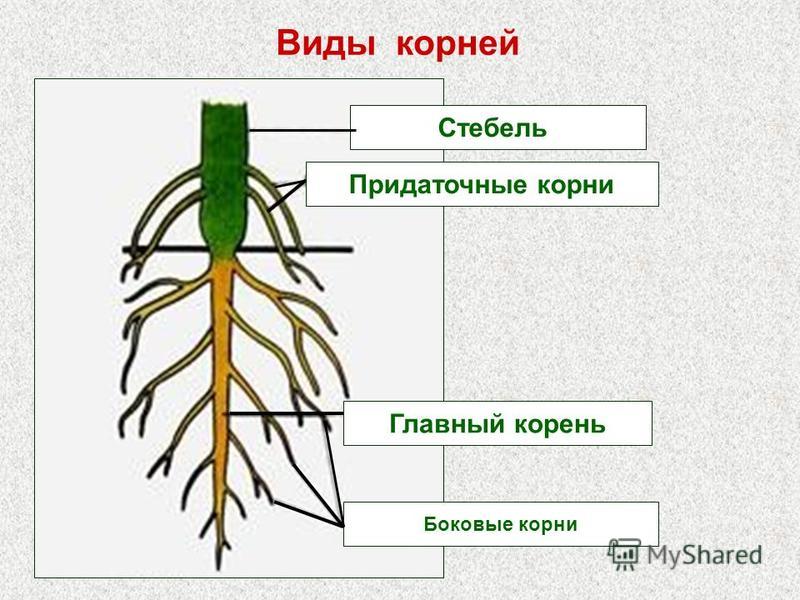 Г. Корневой чехлик определяет зависимый от этилена рост и развитие корней кукурузы. Мол завод. 2008;1:359–367. [PubMed] [Google Scholar]
Г. Корневой чехлик определяет зависимый от этилена рост и развитие корней кукурузы. Мол завод. 2008;1:359–367. [PubMed] [Google Scholar]
88. Darwin F. Über das Wachstum negativ heliotropischer Wurzeln im Licht und im Finster. Arbeiten des Botanisches Instituts в Вюрцбурге. 1880; 2: 521–528. (Гер). [Google Scholar]
89. Ван Ю., Эйзингер В., Эрхардт Д., Балушка Ф., Бриггс В. Субклеточная локализация и индуцированное синим светом движение фототропина 1-GFP в этиолированных проростках Arabidopsis thaliana . Мол завод. 2008; 1:103–117. [PubMed] [Академия Google]
90. Кассаб Г. Другие тропизмы и их связь с гравитропизмом. В: Gilroy S, Masson PH, редакторы. Тропизмы растений. 2008. стр. 123–139. [Google Scholar]
91. Laxmi A, Pan J, Morsy M, Chen R. Свет играет важную роль во внутриклеточном распределении переносчика оттока ауксина PIN2 в Arabidopsis thaliana . ПЛОС ОДИН. 2008;3:1510. [Бесплатная статья PMC] [PubMed] [Google Scholar]
92. Li X, Zhang WS. Тропизм избегания соли у Arabidopsis thaliana . Поведение сигналов растений. 2008;3:351–353. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Li X, Zhang WS. Тропизм избегания соли у Arabidopsis thaliana . Поведение сигналов растений. 2008;3:351–353. [Бесплатная статья PMC] [PubMed] [Google Scholar]
93. Hawes MC, Gunawardena U, Miyasaka SC, Zhao X. Роль пограничных клеток корня в защите растений. Тенденции Растениевод. 2000;5:123–133. [Google Scholar]
94. Балушка Ф., Барлоу П., Баскин Т., Чен Р., Фельдман Л., Форде Б.Г. и др. Что такое апикальное и что такое базальное в развитии корня растения? Тенденции Растениевод. 2005; 10:409–411. [PubMed] [Google Scholar]
95. Friml J, Benfey P, Benková E, Bennett M, Berleth T, Geldner N, et al. Апикально-базальная полярность: почему растительные клетки не стоят на голове. Тенденции Растениевод. 2005; 11:12–14. [PubMed] [Академия Google]
96. Бозе Дж.К. Единство жизни. Президентское обращение на 13-м -м -м Индийском научном конгрессе в Лахоре. 1927 г. Перепечатано в Everyman’s Science 2004; 34: 206–223. [Google Scholar]
97. Пастух В.А. От полупроводников к ритмам чувствительных растений: исследования Дж. К. Бозе. Селл Мол Биол. 2005; 51: 607–619. [PubMed] [Google Scholar]
Пастух В.А. От полупроводников к ритмам чувствительных растений: исследования Дж. К. Бозе. Селл Мол Биол. 2005; 51: 607–619. [PubMed] [Google Scholar]
98. Барлоу П.В. Размышления о биосистемах «нейробиологии растений». 2008; 92: 132–147. [PubMed] [Google Scholar]
99. Struik PC, Yin X, Meinke H. Нейробиология растений и интеллект зеленых растений: наука, метафоры и бессмыслица. J Sci Food Agric. 2008; 88: 363–370. [Академия Google]
100. Бреннер С. В теории. Карр Биол. 1997;7:202. [PubMed] [Google Scholar]
101. Метла Робертсона М. Оккама. Дж. Биол. 2009; 8:79. [Google Scholar]
102. Брюин Дж., Дике М. Химическая передача информации между ранеными и неповрежденными растениями: резервное копирование будущего. Биохим Сист Экол. 2001; 29:1103–1113. [Google Scholar]
103. Pierik R, Whitelam GC, Voesenek LA, de Kroon H, Visser EJ. Исследования листьев табака, нечувствительного к этилену, выявили этилен как новый элемент синего света и передачи сигналов между растениями.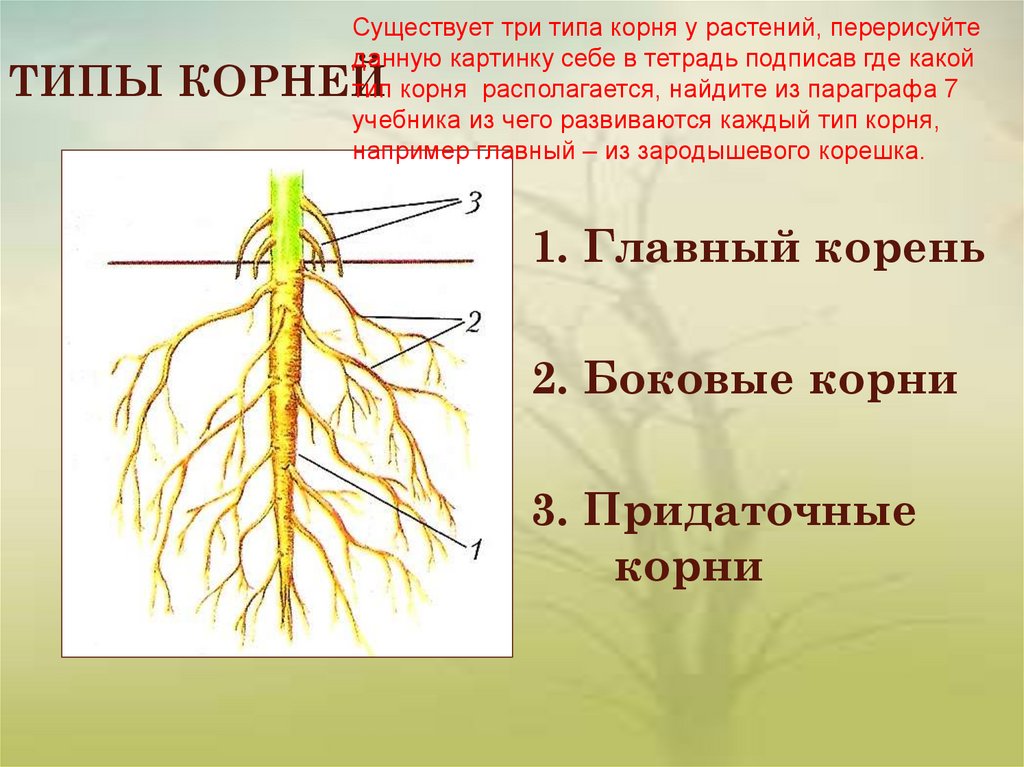 Плант Дж. 2004; 38: 310–319.. [PubMed] [Google Scholar]
Плант Дж. 2004; 38: 310–319.. [PubMed] [Google Scholar]
104. Karban R, Shiojiri K, Huntzinger M, McCall AC. Устойчивость полыни к повреждениям: летучие вещества играют ключевую роль во внутри- и межрастительной коммуникации. Экология. 2006; 87: 922–930. [PubMed] [Google Scholar]
105. Аримура Г., Мацуи К., Такабаяси Дж. Химическая и молекулярная экология летучих веществ растений, вызываемых травоядными: непосредственные факторы и их конечные функции. Физиология клеток растений. 2009;50:911–923. [PubMed] [Google Scholar]
106. Dicke M, Van Loon JJ, Soler R. Химическая сложность летучих веществ растений, вызванная множественной атакой. Nat Chem Biol. 2009 г.;5:317–324. [PubMed] [Google Scholar]
107. Балушка Ф., редактор. Взаимодействие растений и окружающей среды: от сенсорной биологии растений к активному поведению растений. Берлин-Гейдельберг-Нью-Йорк: Springer Verlag; [Google Scholar]
108. Хейл М., Карбан Р. Объяснение эволюции связи растений с помощью сигналов, передаваемых по воздуху.