
Содержание
Мир дикой природы на wwlife.ru
Оглавление
| |
1.
|
|
1.1.
|
|
2.
|
|
2.1.
|
|
2.2.
|
|
2.3.
|
|
2.4.
|
|
2.5.
|
|
2.6.
|
|
2.7.
|
|
2.8.
|
|
2.9.
|
|
2.
|
|
2.11.
|
|
2.12.
|
|
2.13.
|
|
 10.
10.2.1 Животный мир протерозоя. Довендская биота. Животный мир вендского периода (эдикария)
Считается, что первые простейшие животные возникли в конце протерозойской эры — 700 млн лет назад (в некоторых публикациях указывается дата 1,4 млрд. лет назад или даже 2 млрд.лет назад).
В следствии большой распростроненности цианобактерий и водорослей резко возростает содержание кислорода в атмосфере Земли, что приводит к возможности появления таких существ, как животные. Безконтрольный рост кислорода и уменьшение парниковых газов в криогеновом периоде приводит к череде глобальных похолоданий (в период с 750 до 580 млн.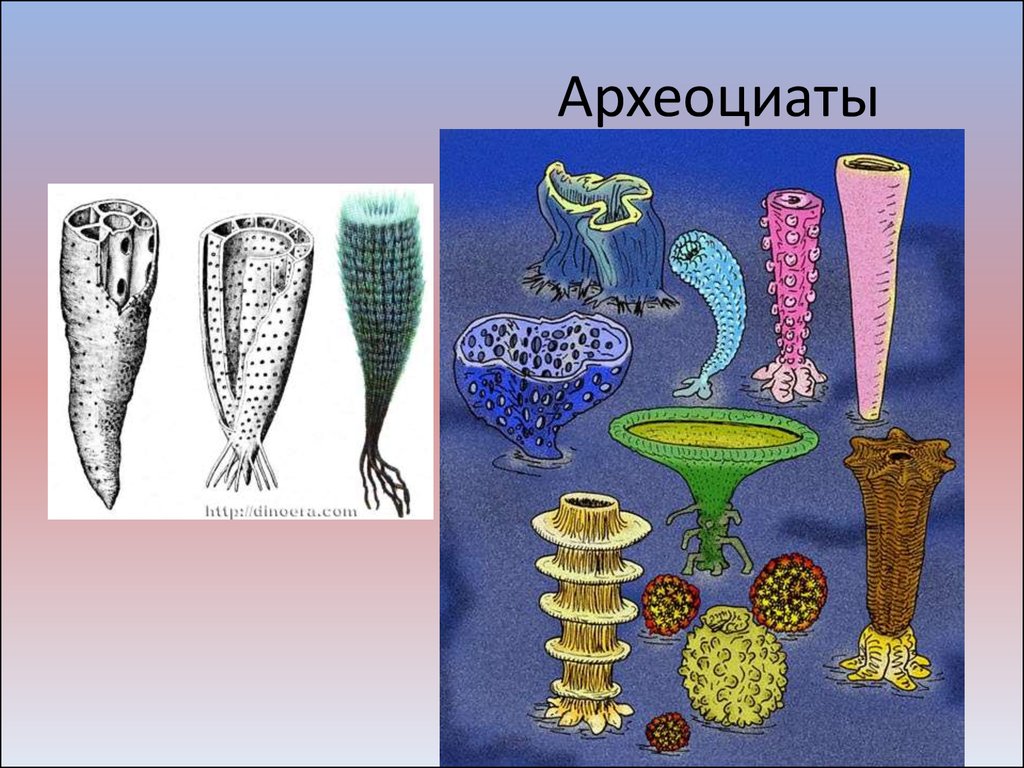 лет назад) покрывших землю слоем льда толщиной до двух километров. Каждое оледениние могло длиться от 4 до 30 млн. лет. Оледенения заканчивались катастрофически быстро, когда благодаря наземному вулканизму в атмосфере накапливалось высокое содержание углекислого газа, более чем в триста раз превышающее его современный уровень.
лет назад) покрывших землю слоем льда толщиной до двух километров. Каждое оледениние могло длиться от 4 до 30 млн. лет. Оледенения заканчивались катастрофически быстро, когда благодаря наземному вулканизму в атмосфере накапливалось высокое содержание углекислого газа, более чем в триста раз превышающее его современный уровень.
Рис. 2.1 Колония хоанофлагелляты SphaeroecaПредположительно первоначально в многоклеточные структуры объединялись простейшие хоанофлагеллаты (рис. 2.1), которые, как полагают, стоят на грани между одноклеточностью и многоклеточностью, образуют зародышеобразные колонии только с помощью бактериального липида, который получают из съеденных бактерий (прокариот). Следующим щагом было появление в этом же периоде первых настоящих многоклеточных макроогранизмов — эти организмы появились на Земле сразу после Мариноанского оледенения – одной из стадий глобального оледенения, когда нашу планету в течение многих миллионов лет сплошь покрывали льды. Первые многоклеточные существа были мягкотелыми организмами, состоящими из отдельных фракталов.
Одни из самых первых появившихся на Земле животных относятся к криогеновому периоду. Эти организмы по размерам были меньше эдикарских и являются не лентовидными, а червеобразными (иногда похожи на членистых). Многие из них строили из органики сегментированные трубки бакаловидной формы. Среди этих организмов нет ни медузоподобных «дисков» как в эдикаре, так и форм похожих на губки (примитивнейших из ныне живущих групп животных). Судя по всему, довендская хайнаньская биота не может считаться предковой ни для эдикарской, ни тем более для современной — фанерозойской [1].
Рис.2.2 Этапы происхождения многоклеточности: I, II—сферические колонии жгутиковых, III—V—фагоцителлы разной степени сложности; 1—кинобласт, 2—рыхлый фагоцитобласт, 3—скопление чувствительных клеток на переднем конце тела, 4—ротовое отверстие, 5—половые клетки, 6—эпителизованный фагоцитобласт Родоначальником многоклеточных в настоящее время считают шаровидную колонию жгутиковых, половые клетки которых перемещались в глубь колонии, а соматические первично выполняли как функцию перемещения всей колонии в пространстве, так и пищеварения за счет переваривания фагоцитированных пищевых частиц, захваченных из воды.
Осуществление одной и той же клеткой функций движения и пищеварения малоэффективно. С этим связана последующая специализация клеток в направлении преимущественно пищеварения или обеспечения движения. Результатом является возникновение фагоцитобласта (внутреннего слоя амебовидных клеток, занимающихся пищеварением) и кинобласта (наружного слоя клеток со жгутиками, обеспечивающими движение).
Стойкая дифференцировка соматических клеток по функциям и строению, возникшая первоначально на фоне выделения двух клеточных слоев, явилась ключевым моментом в происхождении многоклеточных. Именно с двуслойностью связано появление жидкой внутренней среды, через которую клетки обмениваются химическими сигналами, а также дальнейшее обособление и специализация части поверхностных клеток в направлении восприятия внешних раздражителей и передача возбуждения на другие клетки, располагающиеся в отдалении от них. Таким образом возникают предпосылки к формированию нервной системы.
Рис. 2.3. Трихоплакс — Самое примитивное животное на свете похоже на медленно ползающую тонкуюбесформенную пластинку. Гипотетический предок многоклеточных животных назван фагоцителлой (рис. 2.2). Он плавал в толще воды за счет биения ресничек кинобласта, а питался, захватывая взвешенные в среде частички пищи и переваривая их клетками фагоцитобласта. На более поздних этапах эволюции происходили многочисленные адаптации потомков фагоцителлы к многообразным условиям существования при оседании их на дно или при перемещении к поверхности, а также при изменении источников питания (захват мелких или крупных, живых или мертвых пищевых частиц). [2]
Большое значение в эволюции потомков фагоцителлы имели также изменения характера движения: пассивное движение или прикрепленный образ жизни обусловливают лучевой тип симметрии, в то время как активное перемещение в определенном направлении предусматривает формирование двубоковой, или билатеральной, симметрии. В результате возникло огромное многообразие форм многоклеточных животных.[4]
В результате возникло огромное многообразие форм многоклеточных животных.[4]
По другой теории первым примитивным животным является — трихоплакс (рис. 2.3).
Это плоское создание, похожее на медленно ползающую кляксу, не имеет ни осей симметрии, ни мускулатуры, ни переднего и заднего концов, не говоря уже о таких сложных устройствах, как пищеварительная, нервная, кровеносная или выделительная система. Трихоплакс по своему строению напоминает личинок кишечнополостных, и его действительно довольно долго считали личинкой медузы. Но потом оказалось, что трихоплакс образует половые клетки и размножается половым путем.
Митохондриальный геном трихоплакса по своему строению занимает промежуточное положение между «ближайшими родственниками животных» (хоанофлагеллятами и грибами) с одной стороны и всеми остальными животными (включая губок и кишечнополостных) — с другой.[5]
Рис. 2.4. Гребневик. Следующим этапом развития животных стало появление гребневиков (рис. 2.4).[6]
2.4).[6]
Дальнейшим развитием жизни — стало появление 635 млн лет назад (по некоторым данным 850 млн. лет назад) губок (рис. 2.5) развивавшиеся на морском дне, на мелководье, а затем распространившиеся в более глубокие воды.[7]
До развития многоклеточных организмов на нашей планете повсеместно царствовали бактериальные сообщества, покрывая дно океана тонким слоем и выстраивая величественные строматолиты. Первые животные были вынуждены вести с ними жестокую борьбу за существование, получая птательные вещества с воды, им приходилось увеличивать свои габариты, что позволяло поглощать большее количества питательных веществ. [8]
Рис. 2.5. Семейство губок. Одними из наиболее древних находок многоклеточных животных являются археоциаты, а также рангеоморфы, такие, как Харния или чарния Charnia и Charnodiscus, многочисленны медузы (Beltanella, Medusinites, Cyclomedusa и проблематичные формы, близкие современным морским перьям (Rangea, Arborea) жившие в эдикарском периоде. На морском и океаническом дне в то время, обитало большое разнообразие кольчатых червей (известно 5 видов многощетинковых червей принадлежащих родам Сприггина (Spriggina) и Дикинсония (Dickinsonia), от которых в дальнейшем произошли моллюски и членистоногие. Кроме вышеперечисленных морских обитателей эдикария, встречались членистоногие-антроподы (Precambridium), являющиеся отдаленными предками ископаемых трилобитов, а также современных насекомых — пауков и скорпионов. Другими интересными животными эдикара являлись трибрахидиумы (Tribrachidium) которые до сих пор не нашли своей ниши в современой систематике. Некоторые из эдиакарских животных достигали больше метра в размере.
На морском и океаническом дне в то время, обитало большое разнообразие кольчатых червей (известно 5 видов многощетинковых червей принадлежащих родам Сприггина (Spriggina) и Дикинсония (Dickinsonia), от которых в дальнейшем произошли моллюски и членистоногие. Кроме вышеперечисленных морских обитателей эдикария, встречались членистоногие-антроподы (Precambridium), являющиеся отдаленными предками ископаемых трилобитов, а также современных насекомых — пауков и скорпионов. Другими интересными животными эдикара являлись трибрахидиумы (Tribrachidium) которые до сих пор не нашли своей ниши в современой систематике. Некоторые из эдиакарских животных достигали больше метра в размере.
Рис. 2.6. Вендский период (Эдиакарийская биотика). Вообще, в вендский период (рис. 2.6) образовалось большое количество мягкотелых животных не имеющих минерального скелета, останки которых, как уже говорилось, не дошли до наших дней. Тогда же появились первые кишечнополосные хищники.
Животные Эдиакар жили преимущественно на морском дне. Они кормились в слое органического вещества (детрита), который покрывал донный ил, образованный останками множества одноклеточных организмов, населявших толщу воды над ними. Плоские и кольчатые черви плавали над самым дном или ползали среди осадков. Спешить им было некуда, ибо хищников (животных, питающихся другими животными) здесь было очень мало.
Они кормились в слое органического вещества (детрита), который покрывал донный ил, образованный останками множества одноклеточных организмов, населявших толщу воды над ними. Плоские и кольчатые черви плавали над самым дном или ползали среди осадков. Спешить им было некуда, ибо хищников (животных, питающихся другими животными) здесь было очень мало.
Рис. 2.7. Животные Эдиакар (Вендский период). Морские перья поднимались с морского дна (рис. 2.7), подобно неким перообразным цветкам, тщательно отфильтровывая воду в поисках пищи. Трубчатые черви лежали среди донных отложений, шевеля своими щупальцами в насыщенной детритом воде. Примитивные иглокожие, родичи современных морских звезд и морских ежей, всю свою жизнь проводили в толстом слое ила. Было там и множество крупных плоских животных в форме блина; эти похожие на медуз создания также, судя по всему, обитали на илистом дне. А над ними в морской воде медленно проплывали настоящие медузы.
В Эдиакарских отложениях встречаются многочисленные окаменевшие отпечатки мягкотелых животных, ползавших когда-то по морскому дну. В некоторых местах в иле запечатлелись парные V-образные отметины, похожие на царапины, оставленные парами крохотных ножек. Возможно, это следы вышеупомянутых примитивных артропод, или членистоногих, — отдаленных предков ископаемых трилобитов, а также современных нам насекомых — пауков и скорпионов. Правда, твердых останков этих животных пока не обнаружено: по всей видимости, они еще не обзавелись твердым панцирем. [9]
В некоторых местах в иле запечатлелись парные V-образные отметины, похожие на царапины, оставленные парами крохотных ножек. Возможно, это следы вышеупомянутых примитивных артропод, или членистоногих, — отдаленных предков ископаемых трилобитов, а также современных нам насекомых — пауков и скорпионов. Правда, твердых останков этих животных пока не обнаружено: по всей видимости, они еще не обзавелись твердым панцирем. [9]
Самые первые животные возникали в холодных водах, т.к. теплые мелководные бассейны, в частности, обширные моря покрывавшие континенты в рифее, контролировались архаичной прокариотной биотой вплоть до конца венда. Древние цианобактерии, как и современные, были способны защищать себя ядами, которые угнетают рост и размножение эукариот, а в ряде случаев приводят к гибели последних. Так что, колонизация высшими организмами тепловодных бассейнов была непростой задачей.
Первую попытку животных колонизовать тепловодный карбонатный бассейн мы наблюдаем на примере карбонатных отложений Оленекского поднятия (север Якутии). Когда по окончании Варангерского оледенения морские воды начали затапливать континент, животные быстро заняли теплые мелководные обитания. Вендские беспозвоночные довольно долго «удерживали свои позиции» – остатки мягкотелых беспозвоночных, преимущественно, кишечнополостных, в изобилии встречаются в битуминозных тонкослоистых известняках хатыспытской свиты в интервале более 100 метров. Трудно сказать точно, сколько длился этот эпизод, но цианобактериальные сообщества «взяли реванш» и надолго: толща строматолитовых пород туркутской свиты имеет мощность более 200 м. Судя по современным аналогам, строматолиты растут крайне медленно. Лишь в самом конце венда (542±1 млн. лет) и, особенно, в начале кембрийского периода сообщества животных получили возможность вернуться в свободные от строматолитов обитания.
Когда по окончании Варангерского оледенения морские воды начали затапливать континент, животные быстро заняли теплые мелководные обитания. Вендские беспозвоночные довольно долго «удерживали свои позиции» – остатки мягкотелых беспозвоночных, преимущественно, кишечнополостных, в изобилии встречаются в битуминозных тонкослоистых известняках хатыспытской свиты в интервале более 100 метров. Трудно сказать точно, сколько длился этот эпизод, но цианобактериальные сообщества «взяли реванш» и надолго: толща строматолитовых пород туркутской свиты имеет мощность более 200 м. Судя по современным аналогам, строматолиты растут крайне медленно. Лишь в самом конце венда (542±1 млн. лет) и, особенно, в начале кембрийского периода сообщества животных получили возможность вернуться в свободные от строматолитов обитания.
Сезонность питания, характерную для высоких широт, можно рассматривать как фактор отбора в пользу форм с большей массой. Так называемая «резервная биомасса» нужна, чтобы переживать неблагоприятные периоды. Однако рост и размеры тела ограничиваются возможностью обменных процессов – прежде всего дыханием. Развитие гетеротрофии и эффективных способов сбора пищи могло реализоваться в создание резервной биомассы (больших размеров тела) только при условии достаточно высокой концентрации кислорода в воде. Холодноводные бассейны давали такое преимущество.
Однако рост и размеры тела ограничиваются возможностью обменных процессов – прежде всего дыханием. Развитие гетеротрофии и эффективных способов сбора пищи могло реализоваться в создание резервной биомассы (больших размеров тела) только при условии достаточно высокой концентрации кислорода в воде. Холодноводные бассейны давали такое преимущество.
Путь из холодных вод, богатых кислородом, в теплые стал возможным в связи с резким ростом содержания свободного кислорода в атмосфере. Данные изотопного анализа углерода из позднего докембрия показывают, что это событие произошло в самом конце протерозоя.
Специалистам по кораллам известна одна замечательная закономерность: виды, имеющие симбиотические водоросли (их собирательное название – зооксантеллы) формируют прочный массивный скелет, и наоборот – виды без симбиотических водорослей имеют весьма слабую минерализацию скелета или не имеют минерального скелета вовсе. Как любая закономерность в мире живого, эта имеет массу исключений. Но представим вендскую фауну холодных вод, и станет ясно, что там не могло быть мощного минерального скелета по двум причинам: одна из них – низкая эффективности ферментов, ответственных за биоминерализацию, из-за низких температур; другая связана с высокой растворимостью карбоната в холодных водах, его труднее концентрировать и сохранять. Но, возможно была и третья причина – отсутствие зооксантелл у животных, обитающих в высоких широтах – там, где существуют долгие зимние ночи одноклеточным водорослям внутри живого тела выжить трудно. Колонизация тропиков и гарантированный световой день сделал симбиоз более эффективным в двух аспектах: снабжение кислородом хозяина и расширение возможностей биоминерализации.
Но представим вендскую фауну холодных вод, и станет ясно, что там не могло быть мощного минерального скелета по двум причинам: одна из них – низкая эффективности ферментов, ответственных за биоминерализацию, из-за низких температур; другая связана с высокой растворимостью карбоната в холодных водах, его труднее концентрировать и сохранять. Но, возможно была и третья причина – отсутствие зооксантелл у животных, обитающих в высоких широтах – там, где существуют долгие зимние ночи одноклеточным водорослям внутри живого тела выжить трудно. Колонизация тропиков и гарантированный световой день сделал симбиоз более эффективным в двух аспектах: снабжение кислородом хозяина и расширение возможностей биоминерализации.
Животные появились в относительно холодных водах вне карбонатного пояса планеты, который контролировался прокариотами. Эра великих оледенений давала большее преимущество именно эукариотам, в том числе, животным, хотя это было время их трудной эволюции. В эту холодную пору площади карбонатных бассейнов и ареалы прокариотных сообществ резко сократились. Высшие организмы, пережившие 200 млн. лет преимущественно холодной биосферы, по окончании ледниковой эры оказались способными бросить вызов архаичной бактериальной биоте и с начала кембрия прочно заняли тепловодные бассейны карбонатного пояса планеты, колонизировав тепловодные бассейноы карбонатного пояса планеты и постепенно заменяя карбонатные постройки цианобактерий рифами. Это обстоятельство резко ускорило эволюционные процессы, в том числе – на основе сформированного минерального скелета.
Высшие организмы, пережившие 200 млн. лет преимущественно холодной биосферы, по окончании ледниковой эры оказались способными бросить вызов архаичной бактериальной биоте и с начала кембрия прочно заняли тепловодные бассейны карбонатного пояса планеты, колонизировав тепловодные бассейноы карбонатного пояса планеты и постепенно заменяя карбонатные постройки цианобактерий рифами. Это обстоятельство резко ускорило эволюционные процессы, в том числе – на основе сформированного минерального скелета.
Рост разнообразия животных и эвкариот в целом способствовал удлинению пищевых цепей. Однако, в тканях животных, находящихся на вершине трофической пирамиды, могли накапливаться высокие концентрации ряда элементов, в частности, Ca, P, Si. Выведение минеральных солей или детоксикация стали необходимостью. Возможность строить минеральный скелет у части беспозвоночных была следствием детоксикации в условиях тепловодных местообитаний, где растворимость биоминералов ниже и энергетические затраты на биоминерализацию не так высоки, как в холодных водах. [10]
[10]
Животный мир протерозоя. Довендская биота. Животный мир вендского периода (эдикария)
А.С.Антоненко
| Источники: | 1. | Хайнаньская биота |
| 2. | Фагоцителла/ Fagocitella (Паренхимелла) | |
| 4. | Экологический портал | |
| 5. | Элементы | |
| 6. | ScienceBlog.ru | |
| 7. | PrimeInfo | |
8. | Размер имеет значение. | |
| 9. | Теория эволюции как она есть. Эдикар | |
| 10. | Теория эволюции как она есть. Протерозой |
Архейская и протерозойская эры | Статья в журнале «Юный ученый»
Рис. 1
В
сем прекрасно известно о том, что история Земли подразделена на эоны, эры и периоды. Что же из себя представляют архейская и протерозойская эры?
Жизнь на Земле зародилась около 3,8 млрд. лет назад. Деление истории Земли на эры и периоды помогает нам понять особенности развития жизни в разных промежутках времени. Учёные выделяют эры, которые делятся на периоды. В этих отдельных отрезках времени происходят особо значимые события в формировании жизни на нашей планете.
Существует пять эр:
- Архейская;
- протерозойская;
- палеозойская;
- мезозойская;
- кайнозойская. Таблица развития жизни на Земле:
Таблица 1
| | | |
Архейская эра (древняя жизнь) | 3,5 млрд лет | Появление сине-зеленых водорослей, фотосинтез. Гетеротрофы | Преобладание суши над океаном, минимальное количество кислорода в атмосфере. |
Протерозойская эра (ранняя жизнь) | 2,7 млрд лет | Появление червей, моллюсков, первых хордовых, почвообразование. | Суша — каменная пустыня. Накапливание кислорода в атмосфере. |

Всего выделяют два эона-криптозой (скрытая жизнь) и фанерозой (явная жизнь). Фанерозой включает в себя три эры: палеозойскую, мезозойскую и кайнозойскую. Криптозой-две эры: архейскую и протерозойскую.
Архейская эра (от греческого «археос» — начало) -самый древний этап жизни на планете. Данная эра началась с раскалённых вулканов 4 млрд. лет назад, на Землю постоянно падали метеориты. Планета только стала формироваться, следовательно, признаков жизни на ней не было. В воздухе содержался хлор, водород, аммиак, температура достигала 80°, уровень радиации превосходил допускаемые пределы, такие условия препятствовали возникновению жизни. Архей продлился 1,5 млрд. лет. Термин «архей» предложил американский геолог Джеймс Дана в 1872 году. Именно после этого периода зародились первые живые организмы.
В архейской эре геологи выделяют четыре эры:
– Эоархей
– Палеоархей
– Мезоархей
– Неоархей
Эоархей был 4–3,6 млрд лет назад. В этот период формировалась земная кора, кратеры вулканов. Здесь появились цианобактерии.
В этот период формировалась земная кора, кратеры вулканов. Здесь появились цианобактерии.
Палеоархей был 3,6–3,2 млрд лет назад. В этом периоде формировались кратеры вулканов, шло образование мирового океана. Действующие вулканы являлись рельефом поверхности.
Мезоархей был 3,2–2,8 лет назад. Масштабы материка достигли до площади нынешнего Мадагаскара. Именно в этом периоде происходит охлаждение планеты и формируется ледниковое образование.
Неоархей был 2,8–2,5 млрд лет назад. Появляются первые многоклеточные организмы. Формируется второй материк-Ур и образуются полезные ископаемые (золото, гранит, сера).
На период эоархей приходится так называемая “Поздняя тяжёлая бомбардировка” — время формирования кратеров на Луне и Марсе.
В данный период на планете существовал только один вид растений — одноклеточные нитчатые водоросли (сфероморфид). В дальнейшем водоросли привели к формированию лишайников. В архее появились одноклеточные безъядерные организмы, которые называются прокариоты.
К основным событиям завершающего этапа этого эона (неоархею) следует отнести превращение материка Ур в суперконтинент Кенорленд. Это произошло около 2,7 млрд лет назад. Позже, около 2,6 млрд лет назад, Кенорленд присоединился к суперконтиненту Арктида.
Без сомнений, условия раннего архея отличались от современных:
Температура достигала от 95 до 140 °C и при этом отсутствовал кислород.
Несомненно, знакомые и привычные для нас формы жизни не смогли бы появится в подобных ситуациях, где вместо воздуха ядовитые газы.
НО.
Несмотря на это, жизнь уже существовала, и она была бескислородной.
Бескислородные или анаэробные (от греческих слов «ан» — отрицательная частица, «аер» — воздух и «биос» — жизнь) организмы существуют и по сей день.
Первым термин «анаэробные» стал использовать великий французский микробиолог Луи Пастер. В 1861 г. он открыл бактерии, которые вызывали брожение (прокисание) масла. Ученый был поражен тем, что эти странные организмы умели обходиться без кислорода, — необходимую для жизни энергию они получали из таких химических реакций, в которых он не участвует. Для всех анаэробов кислород — опасный яд!
Для всех анаэробов кислород — опасный яд!
Рис. 2. Луи Пастер, 1822–1895 гг.
Имеются весомые причины считать, что они были первыми обитателями нашей Земли. Из-за отличий бескислородных организмов от прочих форм жизни, биологи выделяют анаэробов в отдельное царство-архебактерии.
Как же произошло зарождение жизни — важнейшее событие в истории не только нашей планеты, но и всей Солнечной системы?
Наиболее убедительной гипотезой на данный момент является версия биохимической эволюции, предложенная еще в 1924 г. русским ученым, академиком Александром Ивановичем Опариным в книге «Происхождение жизни».
Рис. 3. Александр Иванович Опарин, 1894–1980гг.
Ученый предложил объяснение того, как под воздействием химических и физических факторов первые одноклеточные формы жизни могли появиться из неживой материи. Как мы знаем, атмосфера архейской Земли была богата аммиаком, оксидами углерода и водяным паром. В более низких концентрациях в ней также присутствовали водород, азот и кислород. Таким образом, основные химические элементы, необходимые для сборки биологически активных молекул, к тому времени уже были доступны, а ультрафиолетовое излучение Солнца могло служить неисчерпаемым источником энергии для химических превращений. Энергия внутреннего тепла Земли (вулканических извержений), могучих грозовых разрядов и радиоактивного распада также, вероятно, участвовала в синтезе сложных молекул из более простых.
Таким образом, основные химические элементы, необходимые для сборки биологически активных молекул, к тому времени уже были доступны, а ультрафиолетовое излучение Солнца могло служить неисчерпаемым источником энергии для химических превращений. Энергия внутреннего тепла Земли (вулканических извержений), могучих грозовых разрядов и радиоактивного распада также, вероятно, участвовала в синтезе сложных молекул из более простых.
По мнению Опарина, биохимическая эволюция могла протекать в три этапа. На первом этапе происходил интенсивный синтез органических (то есть основанных на цепочках углерода) веществ из неорганических предшественников. Соли, растворенные в архейском океане, и атмосферные газы служили реагентами в гигантском химическом реакторе — литосфере древней Земли.
Следующей геологической эрой стала протерозойская (от греческих слов «протерос» — более ранний и «зоя» — жизнь). Ее считают самой длительной в истории Земли. (2,5 млрд — 540 млн лет назад). Он продолжался 2 млрд лет и включал в себя: палеопротерозой, мезопротерозой и неопротерозой. (табл.2). В начале протерозойской эры складывались ядра будущих континентов — древние платформы, или кратоны (от греческого «кратос» — сила, крепость). Самые первые части нынешней Евразии — Восточно-Европейская и Сибирская платформы родом именно из тех времен.
(табл.2). В начале протерозойской эры складывались ядра будущих континентов — древние платформы, или кратоны (от греческого «кратос» — сила, крепость). Самые первые части нынешней Евразии — Восточно-Европейская и Сибирская платформы родом именно из тех времен.
Таблица 2
Деление протерозоя на отдельные периоды не случайно и базируется на стратиграфических исследованиях. Так, палеопротерозой — время достижения кислородом «точки Пастера» — 1 % от его содержания в атмосфере, современной нам. Произошло это событие 2 млрд. лет назад и носит название кислородной катастрофы. Учёные считают, что такая концентрация кислорода достаточна для того, чтобы обеспечить устойчивую жизнедеятельность одноклеточных аэробных организмов, т. е. организмов, нуждающихся в свободном молекулярном кислороде для поддержания своей жизнедеятельности. В отличии от аэробных, анаэробные организмы, разнообразие которых в то время было существенно больше, в большинстве своём вымерли. Для них молекулярный кислород был губительным.
Кроме кислородной катастрофы в эпоху палеопротерозоя наступает первая стабилизация континентов.
Следующий за палеопротерозоем мезопротерозой — время формирования суперконтинента.
Наконец, последний из протерозойских периодов — неопротерозой характеризуется распадом первого суперконтинента и масштабным оледенением, охватившим практически всю поверхность суши. Кроме того, именно к этому временному интервалу относятся древнейшие ископаемые останки животных, что связано с формированием у них твёрдого скелета.
Вулканы продолжали извергаться так же бурно, как и в архейский период. На их активность влияли движения земной коры. Когда материки перемещаются, океаническая кора (та, которая находится под дном океана) «задвигается» под материковую и буквально выдавливает на поверхность магму из земных недр.
Судя по найденным на сегодняшней суше протерозойским отложениям явно морской природы, водная и земная стихии в те времена довольно часто менялись местами: из океанских глубин вздымались юные горы, а более старые скальные хребты уходили под воду.
Протерозойские отложения также показывают, что на Земле в то время уже существовали и пустыни, и ледники — климат был разнообразен.
В этот период появились отложения, ставшие в будущем полезными ископаемыми. Например, месторождения железных руд возникли в результате работы железобактерий (они были открыты в начале ХХ в. русским ученым Сергеем Николаевичем Виноградским). Так появились отложения железистых кварцитов (чередование слоев кварца и железосодержащего магнетита).
Рис. 4. Сергей Николаевич Виноградский
Одно из крупнейших таких месторождений — Курская магнитная аномалия в России. Колоссальные залежи железа в тех местах отклоняют стрелку компаса от ее обычного положения. Эти многокилометровые залежи руды появились 2,5 млрд лет назад на дне протерозойских морей.
В те времена большую территорию Земли охватывало море, а потому вся флора и фауна непосредственно зависела от воды. На суше обитали лишь бактерии, поскольку только они удачно пережили процесс акклиматизации.
Протерозой — это эра водорослей и бактерий. Лишь к концу ее возникли самые ранние представители многоклеточных животных — черви, губки и археоциаты. Это была также эра одноклеточных простейших животных, пока слабо вскрываемых при исследовательских работах. Но главнейшими видимыми проявлениями жизни в протерозое были водоросли типа пресноводных. Водные растения создавали очень своеобразные накопления известняка и даже древнейшие рифы и банки. Исследователь далеко не всегда отличит небольшие водорослевые скопления карбонатной породы от вмещающих отложений. Но повторяемость рисунка поверхности породы иногда подсказывает, что найдены остатки ископаемых древнейших водорослей.
Рис. 5. Остатки ископаемых древнейших водорослей
Температура воздуха у земной поверхности приближалась к 15 °С — не особо жарко, но и не так уж холодно. Из сконденсировавшегося водяного пара складывалась гидросфера планеты — запасы жидкой воды. В нее переходила часть атмосферных газов. До определенного времени в первичной атмосфере кислорода было совсем мало — намного меньше 0,001 его массовой доли в сравнении с нынешним уровнем. Почти каждая вновь образовывавшаяся молекула O2 тратилась на различные реакции окисления. В какой-то момент грянула «кислородная катастрофа» — так называют событие, которое произошло 2,4 млрд лет назад и направило в новое русло ход земной истории. В то время кислород буквально заполнил собой атмосферу планеты. Разумеется, катастрофой это обернулось лишь для анаэробных археобактерий. Об изменениях в атмосфере мы знаем по тому, что характер минеральных отложений с какого-то момента резко преобразился.
До определенного времени в первичной атмосфере кислорода было совсем мало — намного меньше 0,001 его массовой доли в сравнении с нынешним уровнем. Почти каждая вновь образовывавшаяся молекула O2 тратилась на различные реакции окисления. В какой-то момент грянула «кислородная катастрофа» — так называют событие, которое произошло 2,4 млрд лет назад и направило в новое русло ход земной истории. В то время кислород буквально заполнил собой атмосферу планеты. Разумеется, катастрофой это обернулось лишь для анаэробных археобактерий. Об изменениях в атмосфере мы знаем по тому, что характер минеральных отложений с какого-то момента резко преобразился.
Наконец-то в воздухе появилось много свободного кислорода — менее чем за 200 млн лет его концентрация выросла в 15 раз, атмосфера стала не восстановительной, а окислительной.
Для того чтобы возникли анаэробные формы жизни, необходимо было, чтобы концентрация кислорода в атмосфере составляла около 1 % от современной — так называемая точка Пастера. В протерозое этот биологический рубеж был преодолен «с перевыполнением», что стало толчком к бурному развитию и совершенствованию разнообразных форм новой жизни.
В протерозое этот биологический рубеж был преодолен «с перевыполнением», что стало толчком к бурному развитию и совершенствованию разнообразных форм новой жизни.
Представьте себе, что во времена архея железный гвоздь мог лежать на земле миллионы лет, не покрываясь ржавчиной!
Откуда же кислород возник в таком количестве? Вспомните про сине-зеленые водоросли, которые появились еще в архее. Миллионы лет они исправно выделяли кислород в качестве побочного продукта фотосинтеза. Однако он тут же уходил на окисление минералов и газов. В условиях восстановительной атмосферы кислород был настоящим дефицитом и мгновенно расходовался во множестве химических реакций. Когда же все, что можно было окислить, оказалось уже окисленным, кислород стал накапливаться в виде газа. Одновременно с этим падала концентрация углекислого газа в атмосфере, ведь он был так необходим сине-зеленым водорослям для фотосинтеза. Парниковый эффект благодаря этому стал уменьшаться и перед земной жизнью открылись новые заманчивые перспективы.
Основная причина криогенского ледникового периода была той же, что и причина наступления первого ледникового периода: снижение концентрации в атмосфере диоксида углерода или углекислого газа.
Так почему же верхнепротерозойский ледниковый период был гораздо более «мощным»? Этому способствовало расположение материков на время криогения в непосредственной близости от экватора. Когда Земля охлаждается, в силу каких-либо причин, реакция извлечения углекислого газа из атмосферы замедляется, тем самым препятствуя дальнейшему охлаждению. Но расположение материков в тропических широтах не позволяло замедлить темпы удаления углекислого газа, в результате чего ледники продвинулись до экватора, а высокая отражательная способность льда способствовала дальнейшему охлаждению.
Криогенский ледниковый период закончился примерно 635 млн. лет назад, причём очень внезапно. За то время, пока Земля была скована многометровым слоем льда, в её атмосфере, благодаря вулканическим взрывам, постепенно росло количество углекислого газа, ведь механизм его удаления уже не работал.
Таяние льдов, в результате которого в воду попали многие биогены, а также практически полное исчезновение металлического железа из мантии, способствовали быстрому росту парциального давления кислорода: во-первых — произошёл взрывной рост популяций цианобионтов, а во-вторых — перестал действовать механизм связывания атмосферного кислорода железом. Наступил всплеск биоразнообразия, на основании чего ранее выделялся даже особый период протерозойской эры — венд.
Остатки животных в отложениях протерозоя очень редки, но нет сомнения в том, что основы животного мира были заложены одновременно с возникновением мира бактерий и фотосинтезирующих растений. Животные представлены в протерозое мелкими формами, не получившими массового развития и не принимавшими участия в породообразовании.
Таким образом, протерозойская эра истории нашей планеты была в основном временем исключительного господства бактерий и водорослей в водных средах. За этот этап времени, длившийся, по данным абсолютной геохронологии, около 1200 млн. лет, упомянутые группы организмов выполнили огромную геологическую работу по образованию ряда типов осадочных пород и руд, а также по переработке вещественного состава самой биосферы и атмосферы Земли. Протерозойская эра на самом деле послужила основой для формирования всего того, что мы имеем сейчас в окружающей нас действительности.
лет, упомянутые группы организмов выполнили огромную геологическую работу по образованию ряда типов осадочных пород и руд, а также по переработке вещественного состава самой биосферы и атмосферы Земли. Протерозойская эра на самом деле послужила основой для формирования всего того, что мы имеем сейчас в окружающей нас действительности.
Литература:
- https://obrazovaka.ru/istoriya/arheyskaya-era-periody.html
- https://yandex.ru/turbo/sitekid.ru/s/planeta_zemlya/arhejskaya_era_425_mlrd_let_nazad.html
- https://animals-world.ru/delenie-istorii-zemli-na-ery-i-periody/
- https://nsportal.ru/shkola/biologiya/library/2020/03/26/arheyskaya-i-proterozoyskaya-ery
- https://www.sites.google.com/a/tl-2.ru/bionicka2/8-klass-1/razvitie-zizni-na-zemle-v-arhejskuu-i-proterozojskuu-eru
- https://sprint-olympic.ru/uroki/istorija/52817-arheiskaia-era-periody-osobennosti-klimat-kratko.html
- https://yandex.
 ru/turbo/sitekid.ru/s/planeta_zemlya/proterozojskaya_era_25_mlrd__540_mln_let_nazad.html
ru/turbo/sitekid.ru/s/planeta_zemlya/proterozojskaya_era_25_mlrd__540_mln_let_nazad.html - https://yandex.ru/turbo/xn--e1adcaacuhnujm.xn--p1ai/s/proterozojskaya-era.html
 ru/turbo/sitekid.ru/s/planeta_zemlya/proterozojskaya_era_25_mlrd__540_mln_let_nazad.html
ru/turbo/sitekid.ru/s/planeta_zemlya/proterozojskaya_era_25_mlrd__540_mln_let_nazad.htmlПротерозойский эон
Протерозойский эонОнлайн экспонаты: Геологическая шкала времени
Сначала несколько слов о докембрии, неофициальном названии обширного пространства времени до фанерозойского эона (включающего палеозойскую, мезозойскую и кайнозойскую эры). Земля сформировалась. Затем потребовалось почти четыре миллиарда лет, прежде чем первые животные оставили свои следы на планете. Этот промежуток времени составляет примерно семь восьмых истории Земли. В докембрии произошли важнейшие события биологической истории. Учтите, что сформировалась Земля, возникла жизнь, возникли и начали двигаться первые тектонические плиты, эволюционировали эукариотические клетки, атмосфера обогащалась кислородом и незадолго до конца докембрия возникли сложные многоклеточные организмы, в том числе первые животные.
Докембрий делится на три эона. От самого молодого к самому старому: протерозойский, архейский и гадейский (последнее — неофициальное название).
Протерозойский эон
Период истории Земли, начавшийся 2,5 млрд лет назад и закончившийся 542,0 млн лет назад, известен как протерозой, который подразделяется на три эры: палеопротерозойскую (2,5–1,6 млрд лет назад), мезопротерозойскую (1,6–1 млрд лет назад). ) и неопротерозой (от 1 миллиарда до 542,0 миллионов лет назад). Многие из самых захватывающих событий в истории Земли и жизни произошли в течение протерозоя впервые появились и начали срастаться стабильные континенты, причем этот длительный процесс занял около миллиард лет. Также в это время появились первые обильные окаменелости живых организмов, в основном бактерии и археи, но примерно 1,8 миллиарда лет назад эукариотические клетки также появляются в виде окаменелостей.
С началом мезопротерозоя появляются первые свидетельства накопления кислорода в атмосфере. Эта глобальная катастрофа погубила многие группы бактерий, но сделала возможным взрыв эукариотических форм. К ним относятся многоклеточные водоросли, а к концу протерозоя — первые животные.
Эта глобальная катастрофа погубила многие группы бактерий, но сделала возможным взрыв эукариотических форм. К ним относятся многоклеточные водоросли, а к концу протерозоя — первые животные.
Жизнь
Первые следы жизни появляются почти 3,5 миллиарда лет назад, в раннем архее. Однако четко идентифицируемые окаменелости остаются редкими до позднего архея, когда строматолиты, слоистые насыпи, образовавшиеся в результате роста микробных матов, стали обычным явлением в летописи горных пород. Разнообразие строматолитов увеличивалось на протяжении большей части протерозоя. Примерно миллиард лет назад они процветали на мелководье по всему миру. Их значение для понимания протерозойской жизни огромно; окремненные строматолиты (образующие породу, известную как строматолитовые кремни) часто сохраняют изысканные микроокаменелости микробов, которые их создали (см. две фотографии внизу слева).
Слева направо: (1) Образец строматолитового кремня из формации Биттер-Спрингс в центральной Австралии, возраст около 850 миллионов лет. Обратите внимание на типичные тонкие узоры полос. (2) Цианобактерии, вероятно, Myxococcoides minor , колониальная хроококковая форма из кремня Биттер-Спрингс. (3) Кимберелла был билатерально-симметричным эдиакарским животным с жесткими частями — обратите внимание на глубокую впадину, образовавшуюся в иле при захоронении из-за твердого панцирного покрытия. Этот экземпляр из Беломорского региона России имеет диаметр около 1,5 см и длину 2,5-3 см. (4) Два экземпляра Charniodiscus из Мистейкен-Пойнт, Ньюфаундленд. У этих эдиакарских организмов есть «упор» в форме луковицы или диска, который был прикреплен к морскому дну, и основное тело в форме листа, которое, вероятно, удерживалось в толще воды. Обратите внимание на типичные тонкие узоры полос. (2) Цианобактерии, вероятно, Myxococcoides minor , колониальная хроококковая форма из кремня Биттер-Спрингс. (3) Кимберелла был билатерально-симметричным эдиакарским животным с жесткими частями — обратите внимание на глубокую впадину, образовавшуюся в иле при захоронении из-за твердого панцирного покрытия. Этот экземпляр из Беломорского региона России имеет диаметр около 1,5 см и длину 2,5-3 см. (4) Два экземпляра Charniodiscus из Мистейкен-Пойнт, Ньюфаундленд. У этих эдиакарских организмов есть «упор» в форме луковицы или диска, который был прикреплен к морскому дну, и основное тело в форме листа, которое, вероятно, удерживалось в толще воды. | ||||||
Численность и разнообразие строматолитов начали сокращаться около 700 миллионов лет назад. Популярная теория их упадка (хотя, конечно, не единственное возможное объяснение) состоит в том, что травоядные эукариоты, возможно, включая первых животных, эволюционировали примерно в это время и начали активно питаться растущими строматолитами.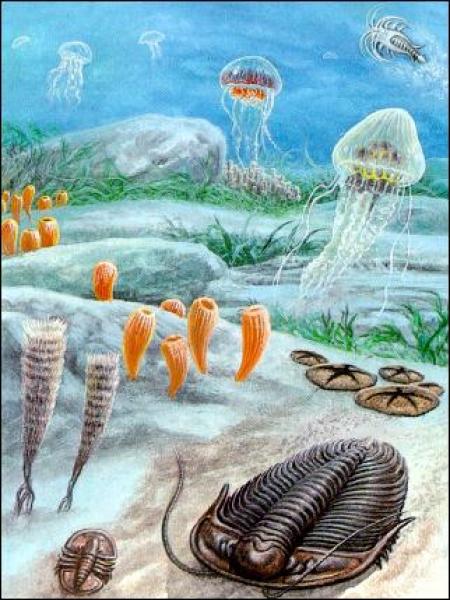 Строматолиты — редкие окаменелости возрастом около 450 миллионов лет. Сегодня они встречаются только в ограниченных местах обитания с низким уровнем выпаса скота, таких как мелководные соленые воды залива Шарк в Австралии.
Строматолиты — редкие окаменелости возрастом около 450 миллионов лет. Сегодня они встречаются только в ограниченных местах обитания с низким уровнем выпаса скота, таких как мелководные соленые воды залива Шарк в Австралии.
Возраст самой старой окаменелости, которая может представлять собой макроскопический организм, составляет около 2,1 миллиарда лет. К концу палеопротерозоя было обнаружено несколько типов окаменелостей, которые, по-видимому, представляют собой простые многоклеточные формы жизни. Эти окаменелости, известные как углеродные пленки, на самом деле таковы: маленькие, темные уплотнения, больше всего напоминающие круги, ленты или листья; они наиболее обычны и широко распространены в неопротерозое. Некоторые напоминают морские водоросли и могут представлять собой эукариотические водоросли; из независимых источников мы знаем, что красные и зеленые водоросли появились в протерозое, вероятно, более миллиарда лет назад.
Следы окаменелостей и молекулярная биология дают дразнящие намеки на то, что животные могли появиться миллиард лет назад. Однако самые древние относительно не вызывающие споров, хорошо изученные окаменелости животных появляются в последние сто миллионов лет протерозоя, как раз перед кембрийским излучением таксонов. Время от 635 миллионов лет назад до 542 миллионов лет назад, известное как эдиакарский период (иногда называемый вендским), стало свидетелем происхождения и первой диверсификации мягкотелых организмов (см. две фотографии вверху справа). Период и фауна названы в честь Эдиакарских холмов на юге Австралии, где были обнаружены первые обильные и разнообразные окаменелости этого вида.
Однако самые древние относительно не вызывающие споров, хорошо изученные окаменелости животных появляются в последние сто миллионов лет протерозоя, как раз перед кембрийским излучением таксонов. Время от 635 миллионов лет назад до 542 миллионов лет назад, известное как эдиакарский период (иногда называемый вендским), стало свидетелем происхождения и первой диверсификации мягкотелых организмов (см. две фотографии вверху справа). Период и фауна названы в честь Эдиакарских холмов на юге Австралии, где были обнаружены первые обильные и разнообразные окаменелости этого вида.
Древнее глобальное загрязнение
Первый «кризис загрязнения» обрушился на Землю около 2,2 миллиарда лет назад. Несколько фактов — наличие оксидов железа в палеопочве (ископаемых почвах), появление «красных пластов», содержащих оксиды металлов, и другие — указывают на довольно быстрое повышение уровня кислорода в атмосфере примерно в это время. Уровень кислорода в атмосфере в архее был менее 1% от нынешнего уровня, но примерно 1,8 миллиарда лет назад уровень кислорода превышал 15% от нынешнего уровня и продолжал расти. Может показаться странным называть это «кризисом загрязнения», поскольку большинство известных нам организмов не только переносят кислород, но и нуждаются в нем для жизни. Однако кислород является мощным разрушителем органических соединений. Даже сегодня многие бактерии и простейшие погибают от кислорода. Организмы должны были разработать биохимические методы обезвреживания кислорода; один из этих методов, окислительное дыхание, имел то преимущество, что производил большое количество энергии для клетки, и в настоящее время обнаружен у большинства эукариот.
Может показаться странным называть это «кризисом загрязнения», поскольку большинство известных нам организмов не только переносят кислород, но и нуждаются в нем для жизни. Однако кислород является мощным разрушителем органических соединений. Даже сегодня многие бактерии и простейшие погибают от кислорода. Организмы должны были разработать биохимические методы обезвреживания кислорода; один из этих методов, окислительное дыхание, имел то преимущество, что производил большое количество энергии для клетки, и в настоящее время обнаружен у большинства эукариот.
Откуда взялся кислород? Цианобактерии, фотосинтезирующие организмы, производящие кислород в качестве побочного продукта, впервые появились 3,5 миллиарда лет назад, но стали обычным явлением и широко распространились в протерозое. Их фотосинтетическая активность была в первую очередь ответственна за повышение содержания кислорода в атмосфере.
местонахождения протерозойских ископаемых
Формирование Биттер-Спрингс: Самые древние из известных окаменелостей эукариот происходят из этого позднепротерозойского доломита в центральной Австралии.
Эдиакарские холмы: В этом австралийском районе в 1946 году были обнаружены останки некоторых из древнейших известных животных.
Мистейкен Пойнт, Ньюфаундленд: Таинственные окаменелости с побережья Ньюфаундленда.
Нопах Диапазон: Древнейшим осадочным породам в этом регионе Южной Калифорнии 1,5 миллиарда лет; более молодые отложения богаты строматолитами.
Белое море: Расположенный на северном побережье России, он был местом активных исследований UCMP вендской фауны.

Ресурсы и ссылки
- Hofmann, HJ 1994. Протерозойские углеродистые уплотнения («метафиты» и «черви»). Стр. 342-357 в С. Бенгтсон (ред.), Ранняя жизнь на Земле. Издательство Колумбийского университета, Нью-Йорк.
- Голландия, Х.Д. 1994. Атмосферные изменения в раннем протерозое. Стр. 237-244 в С. Бенгтсон (ред.), Ранняя жизнь на Земле. Издательство Колумбийского университета, Нью-Йорк.
- Узнайте больше об эдиакарском периоде и эдиакарской биоте в Википедии.
- Узнайте больше о докембрийской палеонтологии и геологии Северной Америки на Палеонтологическом портале.
- См. страницу Википедии о протерозое.

* Даты из Международной стратиграфической таблицы Международной комиссии по стратиграфии, 2009 г.
Бен М. Вагонер создал исходную страницу 20 февраля 1996 г. и начал дополнять и исправлять 28 февраля 2019 г.97; Брайан Р. Спир предоставил новое введение и графику, 08.03.1997; Сара Рибольдт обновила страницы, чтобы отразить геологическую шкалу времени Геологического общества Америки (GSA) 1999 г., 11/2002; Дэйв Смит повторно объединил содержимое в одну страницу, адаптировал его к новому формату сайта и внес некоторые изменения в содержимое, 7 июля 2011 г .; Кремень Биттер-Спрингс, Kimberella и Charniodiscus , фотографии Бена М. Ваггонера; Myxococcoides фото Дж. Уильяма Шопфа
Палеос Протерозой: Протерозойская эра
Палеос Протерозой: Протерозойская эра
| Протерозойский эон | ||
| Протерозой | Протерозой 1 |
Протерозойский эон докембрия Время: от 2500 до 542 миллионов лет назад
Палеопротерозой
Мезопротерозой
Неопротерозой
Фанерозой
Введение
География
Временная шкала
Жизнь
Акритархи
Животные
Животные до Эдиакара?
Эдиакарская среда
Вендобионта
Эмбрионы Доушантуо
Ссылки
Протерозой — это, грубо говоря, время, когда более или менее «современная» тектоника плит стала господствовать над другими процессами в определении формы земной коры. Хотя континенты были небольшими, они состояли из устойчивых кратонов. Срединно-океанические спрединговые хребты, как и сегодня, в значительной степени двигались. Однако все произошло гораздо быстрее. Магма, на которой плавали континенты, была более горячей, менее вязкой и располагалась ближе к поверхности. Горячие точки были, вероятно, более горячими. Континенты двигались быстрее, чаще сталкивались и чаще ломались или сшивались.
Хотя континенты были небольшими, они состояли из устойчивых кратонов. Срединно-океанические спрединговые хребты, как и сегодня, в значительной степени двигались. Однако все произошло гораздо быстрее. Магма, на которой плавали континенты, была более горячей, менее вязкой и располагалась ближе к поверхности. Горячие точки были, вероятно, более горячими. Континенты двигались быстрее, чаще сталкивались и чаще ломались или сшивались.
Жизнь развивалась от младенческой стадии одноклеточных организмов до юности эукариот и ранних растений, грибов, животных. Возможно, возникли и другие формы, о которых мы знаем меньше, потому что они не взорвались во время кембрийского взрыва. Как и все другие подростки, Лайф стал намного больше, открыл для себя секс и часто менял свое мнение о том, чем он станет, когда вырастет. Несомненно, он испробовал множество форм и образов жизни, которые, если бы мы узнали о них в то время, мы бы категорически не одобряли. Жизнь вовлекалась в рискованное поведение, например, небрежно выбрасывала в атмосферу столько кислорода, что чуть не отравила себя, пока не научилась приспосабливаться. Он вышел из теплых геотермальных источников, где, возможно, был поднят, и чуть не замерз один или два раза, забредая в очень серьезные ледниковые периоды без рукавиц. Каким-то образом, несмотря на ряд этих очень близких вызовов, она переросла в ту взрослую Жизнь, которую мы знаем сегодня.
Он вышел из теплых геотермальных источников, где, возможно, был поднят, и чуть не замерз один или два раза, забредая в очень серьезные ледниковые периоды без рукавиц. Каким-то образом, несмотря на ряд этих очень близких вызовов, она переросла в ту взрослую Жизнь, которую мы знаем сегодня.
ATW040930. Всеобщее достояние. Права не защищены.
Мы очень мало знаем о географии протерозоя. Как отмечает доктор Кристофер Скотезе на своем сайте палеокарты: «По имеющимся данным, 650 миллионов лет — это самое большое расстояние, которое мы можем проследить». Конечно, это не помешало ему (или многим другим) расширить границы палеокарты примерно до 750 млн лет назад. Однако, помимо этого, есть только обоснованные догадки.
Теперь один факт кажется достаточно ясным. Около 1100 или 1200 млн лет назад большая часть суши Земли была заперта на континенте под названием Родиния. На данный момент вопрос о том, как собиралась Родиния, является спекулятивным. Поскольку доктор Скотезе, несомненно, является самым образованным из образованных гадателей, вам может быть интересно увидеть его концепцию Родинии в ее наибольшей степени (возможно, 1100 млн лет назад).
Приведенная выше карта взята из Геологическая история Джеймстауна, Род-Айленд . Она представляет собой одну из популярных интерпретаций состава Родинии примерно 900–1000 млн лет назад. Большинство континентов представляют собой грубые капли. Однако мы знаем, что Лаврентия была примерно вверх ногами по сравнению с сегодняшним днем. Следовательно, центр Земли в то время находился где-то в северной части штата Миннесота — этого достаточно, чтобы доказать, что это был действительно мир, очень отличный от нашего.
Около 900 млн лет назад Родиния начала разрушаться; и модель фрагментации стала довольно хорошо известна. На этой карте показана очень красивая структура вокруг угла Амазония — Балтика — Лаврентия, известная как трехрукий грабен . Это не какая-то мошенническая игра или высокотехнологичный игровой автомат. Это настоящее разрушающее континент слияние двух разломов точно такого же типа, который разорвал Пангею на части 800 млн лет назад и который пытается, но, вероятно, безуспешно, разорвать Восточную Африку в наши дни. На самом деле угол Калахари — Восточная Антарктида — Лаврентия может быть местом второго грабена, точно так же, как на самом деле было две таких структуры, причастных к гибели Пангеи. Скотезе, кажется, уделяет больше всего внимания разлому с северо-запада на юго-восток, разделяющему Родинию на части. Огромная масса Лаврентии и Сибири затем поворачивается по часовой стрелке примерно на 120° по часовой стрелке, двигаясь вниз почти к Южному полюсу и снова поднимаясь, ударяя по Западной Африке с Юг точно так же, как Восточная Гондвана (т.е. Северная Родиния), немного повернувшись по часовой стрелке, попадает в нее с северо-востока.
На самом деле угол Калахари — Восточная Антарктида — Лаврентия может быть местом второго грабена, точно так же, как на самом деле было две таких структуры, причастных к гибели Пангеи. Скотезе, кажется, уделяет больше всего внимания разлому с северо-запада на юго-восток, разделяющему Родинию на части. Огромная масса Лаврентии и Сибири затем поворачивается по часовой стрелке примерно на 120° по часовой стрелке, двигаясь вниз почти к Южному полюсу и снова поднимаясь, ударяя по Западной Африке с Юг точно так же, как Восточная Гондвана (т.е. Северная Родиния), немного повернувшись по часовой стрелке, попадает в нее с северо-востока.
Можно представить себе более мирный сценарий, в котором Лаврентия и Балтика просто пересекают «пролив» между Калахари и Восточной Антарктидой, вращаясь (в любом направлении), чтобы достичь своих кембрийских позиций и ориентаций. Есть веские причины, почему это не может быть так, не последней из которых является крайняя южная палеоширота Северной Америки (Лаврентия) в позднем неопротерозое. Тем не менее, в духе несносности мы предлагаем эту дополнительную карту от Стивена Датча из Университета Висконсина. Большое темно-зеленое пятно на западе карты Датча — это блок Нигера, на самом деле части Африки и Южной Америки. См. Johnson & Rivers (2004), который поддерживает это размещение. Если эта версия — не так уж сильно отличающаяся от карты выше — верна, Лаврентии будет трудно вращаться на юг вокруг этих препятствий, чтобы достичь кембрийского положения примерно там, где показан блок Нигера, всего за 230 млн лет. . Не исключено, заметьте, но не самая экономная интерпретация с геометрической точки зрения.
Тем не менее, в духе несносности мы предлагаем эту дополнительную карту от Стивена Датча из Университета Висконсина. Большое темно-зеленое пятно на западе карты Датча — это блок Нигера, на самом деле части Африки и Южной Америки. См. Johnson & Rivers (2004), который поддерживает это размещение. Если эта версия — не так уж сильно отличающаяся от карты выше — верна, Лаврентии будет трудно вращаться на юг вокруг этих препятствий, чтобы достичь кембрийского положения примерно там, где показан блок Нигера, всего за 230 млн лет. . Не исключено, заметьте, но не самая экономная интерпретация с геометрической точки зрения.
ATW040930. Текст общественное достояние. Права не защищены.
Эон | Эра | Период | когда началось Мой назад | продолжительность Моя |
| Протерозой | Неопротерозой 1000 млн лет назад | Эдиакарский | 630 | 88 |
| Криогенный | 850 | 250 | ||
| Тониан | 1000 | 150 | ||
| Мезопротерозой 1600 млн лет назад | Стениан | 1200 | 200 | |
| Эктазиан | 1400 | 200 | ||
| Калимийцы | 1600 | 200 | ||
| Палеопротерозой 2500 млн лет назад | Статерианский | 1800 | 200 | |
| Оросириан | 2050 | 250 | ||
| Райский | 2300 | 250 | ||
| Сидериан | 2500 | 200 |
Протерозой включает более половины истории жизни на этой планете, но доступные данные о протерозойских организмах чрезвычайно скудны. Обычно мы немного обсуждаем планктон примерно в этот момент на страницах Time, но мы практически ничего не знаем о планктоне в протерозое. Следовательно, вместо этого мы будем говорить в основном об акритархах. А что именно это акритарх? Никто не уверен. Акритархи подобны зубам акул из фанерозоя или окуркам из
Обычно мы немного обсуждаем планктон примерно в этот момент на страницах Time, но мы практически ничего не знаем о планктоне в протерозое. Следовательно, вместо этого мы будем говорить в основном об акритархах. А что именно это акритарх? Никто не уверен. Акритархи подобны зубам акул из фанерозоя или окуркам из
1960-е годы. То есть они встречаются повсюду, практически не поддаются разрушению и бывают самых разных интересных форм и размеров. Они кажутся информативными, но в конечном итоге довольно мало говорят нам об организмах или экосистеме, которые их произвели, кроме того факта, что в ней обитали акулы или курильщики, в зависимости от обстоятельств. Акритархи говорят нам еще меньше, потому что мы не знаем, что их породило. Типичное утверждение в литературе звучит так: «Акритархи [являются] группой устойчивых к разложению везикулярных микрофоссилий с органическими стенками… Большинство акритарх протерозоя и палеозоя интерпретируются как одноклеточные фотосинтезирующие протисты…». Хантли и др. . 2006).
Хантли и др. . 2006).
Несмотря на то, что акритархов исследовали и подвергали практически всем видам устройств, известных человеку, они дали относительно мало подробной структурной информации. Это явно неминерализованные структуры с органическими стенками. После простоя в течение одного или двух миллиардов лет почти вся молекулярная организация наномасштаба была утеряна. Остается кероген , аморфные пластинки полициклических ароматических углеводородов, не имеющие явного сходства с каким-либо знакомым материалом клеточных стенок. Кемпе и др. . 2002).
Акритархи раннего палеопротерозоя пока не известны. Данные биомаркеров (, т.е. наличие длинноцепочечных 2-метилгопанов) позволяют предположить, что присутствовала популяция фитопланктона, состоящая в основном/полностью из цианобактерий. Вызывает и др. . (1999 г.); Фальковски и др. . (2004 г.); Кэнфилд (2005).
Акритархи присутствуют в позднем палеопротерозое; но они редки и состоят из морфологически простых сфер (т. е. сфероморфных акритархов). Javaux и др. . 2004). Иногда они очень похожи на празинофиты (базальные зеленые водоросли), и, возможно, они таковыми и являются. Скорее всего, они находятся ниже разделения между красными и зелеными водорослями, такими как глаукофиты, или даже базальнее всех коронных эукариот. Многие или большинство акритарх палеопротерозоя и раннего мезопротерозоя, вероятно, являются акинетами или другими бактериальными остатками. Голубкова и Раевская (2005). Акинеты — это неактивные покоящиеся стадии цианобактерий, вызванные холодом, нехваткой пищи или подобными условиями окружающей среды, неблагоприятными для роста — виды стимулов, которые обычно заставляют организмы искать метаболический застой и/или поступление в аспирантуру. Микс и др. . 2002) (обзор).
е. сфероморфных акритархов). Javaux и др. . 2004). Иногда они очень похожи на празинофиты (базальные зеленые водоросли), и, возможно, они таковыми и являются. Скорее всего, они находятся ниже разделения между красными и зелеными водорослями, такими как глаукофиты, или даже базальнее всех коронных эукариот. Многие или большинство акритарх палеопротерозоя и раннего мезопротерозоя, вероятно, являются акинетами или другими бактериальными остатками. Голубкова и Раевская (2005). Акинеты — это неактивные покоящиеся стадии цианобактерий, вызванные холодом, нехваткой пищи или подобными условиями окружающей среды, неблагоприятными для роста — виды стимулов, которые обычно заставляют организмы искать метаболический застой и/или поступление в аспирантуру. Микс и др. . 2002) (обзор).
Другие типы окаменелостей вблизи границы палеопротерозоя и мезопротерозоя включают спиральные или червеобразные формы, такие как Grypania, и, возможно, родственные структуры, напоминающие бусы на нитке. Портер (2004). Некоторые из Grypania -подобных окаменелостей достигают 1 мм в ширину, что кажется неоправданно большим для бактерии или даже колониальной бактериальной структуры. Тем не менее, самым ранним Grypania примерно на миллиард лет больше, чем настоящим червям. Так что, возможно — из-за отсутствия какой-либо другой гипотезы — это очень ранние эукариоты, возможно, вне коронной группы.
Портер (2004). Некоторые из Grypania -подобных окаменелостей достигают 1 мм в ширину, что кажется неоправданно большим для бактерии или даже колониальной бактериальной структуры. Тем не менее, самым ранним Grypania примерно на миллиард лет больше, чем настоящим червям. Так что, возможно — из-за отсутствия какой-либо другой гипотезы — это очень ранние эукариоты, возможно, вне коронной группы.
К раннему мезопротерозою (калимскому периоду) свидетельства эукариотической степени организации становятся более определенными. Это суждение основано на: «(1) структуре стенок и орнаментации поверхности (2) процессах, которые распространяются от стенок пузырьков (3) эксцистозных структурах (отверстиях, через которые кисты высвобождают свое клеточное содержимое) (4) ультраструктуре стенок и (5) химическом составе стенок .» Javaux и др. . 2004). В частности, крупные клетки с отростками, выходящими за пределы стенки (т.е. акантоморфные acritarchs) считается невозможным без эукариотического цитоскелета. Идентификатор . Как ни странно, это довольно хорошо согласуется с недавней работой «молекулярных часов», которая также относит первичное излучение Plantae к калимийскому периоду. Юн и др. . 2004). Мезопротерозойские акритархи включают образцы с новыми морфологическими признаками: эллипсоидальной формой, порами пузырьков, многоклеточным или колониальным видом. Хантли и др. . 2006).
Идентификатор . Как ни странно, это довольно хорошо согласуется с недавней работой «молекулярных часов», которая также относит первичное излучение Plantae к калимийскому периоду. Юн и др. . 2004). Мезопротерозойские акритархи включают образцы с новыми морфологическими признаками: эллипсоидальной формой, порами пузырьков, многоклеточным или колониальным видом. Хантли и др. . 2006).
Жаво и др. . 2001) изучал хорошо датированные образцы из группы Роупер в северной Австралии. Они обнаружили образцы спорного, но почти наверняка эукариотического вида Tappania . Обратите внимание на относительно большой размер и длинные неправильные отростки, проникающие во внешнюю стенку. Бактериальное происхождение не является полностью невозможным, но более вероятным объяснением является эукариот с хорошо развитым цитоскелетом. В любом случае, некоторые из акритархов, известных из этой эпохи, относительно огромны, например, Chuaria circleis (более известный из неопротерозоя), который может достигать 1 мм в диаметре. Голубкова и Раевская (2005).
Голубкова и Раевская (2005).
Еще один ключевой вывод этого исследования заключался в том, что акритархи Ропера демонстрировали четкую экологическую зональность с различными популяциями, характерными для прибрежной, прибрежной и дальней шельфовой среды. Авторы предполагают, что сообщества были ограничены стоком питательных минералов, поскольку изобилие и разнообразие кажутся выше в маргинальных морских условиях. Однако нет никакой гарантии, что разнообразие акритархов отражает биотическое разнообразие в целом.
Чуть более поздние (эктазианские) сообщества из группы Руян Северного Китая во власти Dictyosphaera , акритарха, также обнаруженного в группе Ропера. Кауфман и Сяо (2003). Такое космополитическое распространение типично для протерозойских акритарх. Авторы выполнили ионный микрозондовый изотопный анализ отдельных образцов и смогли сделать грубую оценку того, что мезопротерозойские уровни CO 2 в настоящее время были в 10–1000 раз выше.
Неясно, являются ли эти окаменелости коронными эукариотами ( , то есть потомки последнего общего предка всех ныне живущих эукариот). Это кажется вероятным хотя бы потому, что предполагается, что все акритархи были фотосинтезаторами; но остается много места для сомнений. Однако существует скромное мнение, что коронные эукариоты появились Стенианом, если не раньше, поскольку эти поздние мезопротерозойские акритархи больше напоминают современные водоросли. Javaux и др. . (2004 г.); Портер (2004). В частности, Bangiomorpha выглядит как современная водоросль и демонстрирует очень необычный «интеркалярный» паттерн клеточного деления, характерный только для современных красных водорослей из семейства банговых. Портер (2004). Некоторые акритархи этого периода можно встретить с поверхностным орнаментом, а не просто отростками. Стениан также стал первым
Это кажется вероятным хотя бы потому, что предполагается, что все акритархи были фотосинтезаторами; но остается много места для сомнений. Однако существует скромное мнение, что коронные эукариоты появились Стенианом, если не раньше, поскольку эти поздние мезопротерозойские акритархи больше напоминают современные водоросли. Javaux и др. . (2004 г.); Портер (2004). В частности, Bangiomorpha выглядит как современная водоросль и демонстрирует очень необычный «интеркалярный» паттерн клеточного деления, характерный только для современных красных водорослей из семейства банговых. Портер (2004). Некоторые акритархи этого периода можно встретить с поверхностным орнаментом, а не просто отростками. Стениан также стал первым
появление стебельчатых цианобактерий. Голубкова и Раевская (2005).
Окаменелости даже самого раннего неопротерозоя (тонского периода) включают формы, обычно идентифицируемые как грибы, и известные современные отряды зеленых водорослей. Фальковски и др. . (2004 г.); Голубкова и Раевская (2005). (Мы опускаем подробное рассмотрение многоклеточных животных, включая эдиакарские формы, и метафитов, появившихся в позднем криогене и эдиакаре. Peterson & Butterfield (2005). Они подробно обсуждаются ниже и в разделах, посвященных этим эпохам). Морфологические особенности, новые для неопротерозоя, включают «многогранные везикулы, пузыревидные везикулы, бочкообразные везикулы, треугольные и волосовидные отростки, отростки с воронкообразными наконечниками, отростки, которые сливаются на концах, и фланцевый орнамент вокруг экватора везикул». Хантли и др. . 2006) (примеры и внутренние ссылки опущены).
. (2004 г.); Голубкова и Раевская (2005). (Мы опускаем подробное рассмотрение многоклеточных животных, включая эдиакарские формы, и метафитов, появившихся в позднем криогене и эдиакаре. Peterson & Butterfield (2005). Они подробно обсуждаются ниже и в разделах, посвященных этим эпохам). Морфологические особенности, новые для неопротерозоя, включают «многогранные везикулы, пузыревидные везикулы, бочкообразные везикулы, треугольные и волосовидные отростки, отростки с воронкообразными наконечниками, отростки, которые сливаются на концах, и фланцевый орнамент вокруг экватора везикул». Хантли и др. . 2006) (примеры и внутренние ссылки опущены).
Вообще говоря, криогенные данные скудны, но предполагают преемственность с тонскими формами без резких изменений в морфологии или даже разнообразия. Портер 2004). Первые ископаемые останки панцирных амеб появляются в конце криогенного периода — первое достоверное свидетельство существования гетеротрофных эукариот. Портер (2004). Эдиакарцы представили два новых сообщества, одно из которых связано с эдиакарскими животными (или кем бы они ни были), за которым следует другое, связанное с переходными многоклеточными животными типа доушантуо. К последним относятся узнаваемые современные отряды красных водорослей. Сяо и др. . 2004). Многие акритархи этого периода имели регулярные отростки и орнамент на поверхности, например, . Appendisphaera , Ericasphaera . Голубкова и Раевская (2005).
К последним относятся узнаваемые современные отряды красных водорослей. Сяо и др. . 2004). Многие акритархи этого периода имели регулярные отростки и орнамент на поверхности, например, . Appendisphaera , Ericasphaera . Голубкова и Раевская (2005).
Был предпринят ряд усилий для количественной оценки характера разнообразия акритархов в протерозое. Нолл (1994); Портер, 2004 г.); Хантли и др. . (2006). В целом они подтверждают, что после первоначального всплеска диверсификации в позднем палеопротерозое развитие было постепенным или даже статичным до неопротерозоя. Удивительно, но доказательства узкого места разнообразия или даже изобилия в результате криогенных эпизодов «земли-снежного кома» слабы или почти отсутствуют. См. также Olcott et al . 2005) (доказательства биомаркеров). Было бы правильнее сказать, что кривая разнообразия начала подниматься в тониане и, самое большее, замедлилась в криогене.
ATW061208. Текст общественное достояние. Права не защищены.
Права не защищены.
В долгосрочной перспективе самой успешной группой животных протерозоя были губки. Мы несколько неохотно много говорим о протерозойских губках, потому что у нас возникли очень своеобразные и идиосинкразические представления об эволюции губок в другой части Палеоса. Эти предположения были обобщены в другом месте, возможно, в самом плохо написанном эссе на этом сайте. Мы ориентировочно придерживаемся того, что было сказано, если не того, как мы это сказали.
Здесь мы будем более консервативны, поскольку мы подводим итоги текущего состояния огромной темы, по которой нет единого мнения между двумя учеными. Мы представим только две самодельные идеи. Первый касается филогенетического положения Kimberella . Второй — это еще один взгляд на знаменитых эмбрионов Доушантуо, упомянутых в предыдущем разделе. План (если мы сможем его придерживаться) состоит в том, чтобы избегать слишком тяжелых размышлений и концентрироваться на мыслях более квалифицированных людей.
Мы начнем с краткой ориентации, которая охватывает первые 95% протерозоя в паре абзацев. Затем мы рассмотрим эдиакарскую фауну, которую назовем Vendobionta, следуя терминологии, введенной Зейлахером. Особое внимание мы уделим «рангеоморфным» вендобионтам и некоторым идеям, развитым Нарбонном (2004). Мы кратко применим эти идеи к Kimberella , вендобионту, которого часто сравнивают с моллюсками. Короче говоря, мы подозреваем, что независимо от того, Kimberella на самом деле является базальным моллюском, его также можно интерпретировать как рангеоморфный вендобионт, не имеющий особого происхождения. После этого мы немного поговорим о докембрийских губках в качестве своеобразной разминки перед демилитаризованной зоной Доушаньто. Наконец, мы осторожно пройдем через само минное поле Доушантуо. Местонахождение Weng’an в формации Doushantuo было местом ожесточенных научных сражений с 6 февраля 1998 года, когда Li et al . (1998) объявили об открытии окаменелостей и эмбрионов маленьких губок из Weng’an. Мы не принимаем ничьей стороны в этом открытом для всех, но мы вводим возможность, которую (насколько нам известно) никто из сражающихся, по-видимому, не рассматривал: что зародыши Weng’an являются зародышами вендобионтов. ATW081205
Мы не принимаем ничьей стороны в этом открытом для всех, но мы вводим возможность, которую (насколько нам известно) никто из сражающихся, по-видимому, не рассматривал: что зародыши Weng’an являются зародышами вендобионтов. ATW081205
Животные до Эдиакара?
В старые времена молекулярной филогении, грубо говоря, в десятилетие между 1995 и 2005 годами, было обычным делом читать статьи о «молекулярных часах», в которых уверенно объявлялось, что метазои эволюционировали в мезопротерозое, если не раньше. К счастью, нам больше не нужно тратить страницы на эти разногласия. В более поздних работах с использованием гораздо более сложных методов были получены результаты, значительно более согласующиеся с летописью окаменелостей. Петерсон и др. . (2008) — хороший пример и хорошее исследование.
Группа Петерсона уделяет большое внимание калибровке своего молекулярного анализа с использованием нескольких временных точек из летописи окаменелостей. Они обнаруживают (с большими погрешностями), что грибы отделились от Metazoa в какой-то момент стенианского или тонианского времени. Мы используем «Metazoa» для обозначения жаб > поганок. Это слишком много, и мы продолжаем использовать его только для согласованности с другими частями Палеоса. Когда большинство людей используют термин «животное», они обычно имеют в виду что-то вроде губки + Спиноза. В этом смысле животные начали развиваться в криогене, с развитием основных клад губок. Петерсон и др. . обнаруживают, что большинство клад животных высокого уровня затем разошлись в эдиакарском периоде и разошлись в кембрии. При оценке этих результатов важно помнить, что мы говорим о временах генетической дивергенции. Окаменелости с поддающимися обнаружению специфическими для клады характеристиками («апоморфиями») могут появиться только спустя долгое время после того, как клада генетически отличается от всех остальных. При таком понимании результаты, о которых сообщила группа Петерсона, вполне заслуживают доверия. Поскольку они тесно связаны с летописью окаменелостей, такие исследования представляют собой синтез имеющихся в настоящее время молекулярных и палеонтологических данных.
Мы используем «Metazoa» для обозначения жаб > поганок. Это слишком много, и мы продолжаем использовать его только для согласованности с другими частями Палеоса. Когда большинство людей используют термин «животное», они обычно имеют в виду что-то вроде губки + Спиноза. В этом смысле животные начали развиваться в криогене, с развитием основных клад губок. Петерсон и др. . обнаруживают, что большинство клад животных высокого уровня затем разошлись в эдиакарском периоде и разошлись в кембрии. При оценке этих результатов важно помнить, что мы говорим о временах генетической дивергенции. Окаменелости с поддающимися обнаружению специфическими для клады характеристиками («апоморфиями») могут появиться только спустя долгое время после того, как клада генетически отличается от всех остальных. При таком понимании результаты, о которых сообщила группа Петерсона, вполне заслуживают доверия. Поскольку они тесно связаны с летописью окаменелостей, такие исследования представляют собой синтез имеющихся в настоящее время молекулярных и палеонтологических данных.
Некоторые результаты палеонтологии также предполагают раннее начало Metazoa. Наиболее многообещающие из этих кандидатов были рассмотрены Федонкиным (2003). Из них одним из самых интересных является очень простое существо из бусинок на нитке, Horodyskia . Федонкин и Йохельсон (2002). Horodyskia нередко встречается в донных отложениях калимского возраста (мезопротерозой, около 1500 млн лет назад) или моложе в Северной Америке и Австралии. Конические бусины, или «зооиды», явно чем-то связаны, предположительно столоном. Струны кажутся относительно одинаковой длины. Размер зооидов одинаков для каждой строки, но значительно различается между строками, отражая рост зооидов со временем. Пропорциональное расстояние между зооидами сохраняется, по-видимому, за счет резорбции чередующихся зооидов. Таким образом, предполагаемые молодые цепочки имеют много мелких зооидов, в то время как более старые цепочки имеют ту же длину, но имеют небольшое количество более крупных зооидов. Такой скоординированный рост предполагает, как отмечают Федонкин и Йохельсон, организацию «на уровне ткани», а не колонию индивидуумов.
Такой скоординированный рост предполагает, как отмечают Федонкин и Йохельсон, организацию «на уровне ткани», а не колонию индивидуумов.
Городиска не входит в наше уравнение. Федонкин (2003) в общих чертах называет Horodyskia «многоклеточным животным». Немногие всерьез утверждают, что Horodyskia происходит от последнего общего предка лягушек и грибов, а уж тем более что это «животное» в том смысле, в каком оно определено выше. Некоторые исследователи до сих пор не до конца убеждены, что Horodyskia является даже эукариотической. Нолл и др. . (2006). Кооперативная многоклеточность развилась независимо как минимум в шести живых группы эукариот (красные водоросли, зеленые водоросли, 2-3 группы хромистов, слизевики, грибы, метазои). Удивительно, что известно так мало вымерших многоклеточных клад.
Федонкин (2003) упоминает также червеобразные Parmia и Sinosabellidites , два червеобразных организма раннего неопротерозоя. Parmia выглядит как нечестивая помесь морского огурца (Holothuroidea) и кольчатого червя. Известен только по тониану Русской платформы. Sinosabellidites и несколько подобных форм — несколько более молодые организмы из Китая. Все показывают близко расположенные кольца, на расстоянии порядка 200 микрон друг от друга. Однако Донг и др. . (2007) интерпретируют организмы этого типа как водоросли, а любой видимый хоботок интерпретируется как опора. Короче говоря, доказательства существования передовых многоклеточных животных до эдиакара неубедительны. ATW081205.
Parmia выглядит как нечестивая помесь морского огурца (Holothuroidea) и кольчатого червя. Известен только по тониану Русской платформы. Sinosabellidites и несколько подобных форм — несколько более молодые организмы из Китая. Все показывают близко расположенные кольца, на расстоянии порядка 200 микрон друг от друга. Однако Донг и др. . (2007) интерпретируют организмы этого типа как водоросли, а любой видимый хоботок интерпретируется как опора. Короче говоря, доказательства существования передовых многоклеточных животных до эдиакара неубедительны. ATW081205.
Окружающая среда Эдиакара
Окружающая среда, в которой эволюционировали многоклеточные эдиакарские животные, лишь немногим менее противоречива. Некоторые вопросы обсуждались выше, в разделе об акритархах. Эдиакарский период начинается с окончания эпизода Маринойского оледенения «Земля-снежок», примерно 630 млн лет назад. В настоящее время кажется, что за этим последовало увеличение содержания кислорода в атмосфере или, по крайней мере, насыщение кислородом океанов. Главный предмет разногласий заключается в том, происходило ли насыщение кислородом каким-то ступенчатым образом — через серию быстрых «событий» (Fike и др. , 2006; 2007 г.; Скотт и др. , 2008) или более постепенно (Grey & Calver, 2007; Shen et al. , 2008).
Главный предмет разногласий заключается в том, происходило ли насыщение кислородом каким-то ступенчатым образом — через серию быстрых «событий» (Fike и др. , 2006; 2007 г.; Скотт и др. , 2008) или более постепенно (Grey & Calver, 2007; Shen et al. , 2008).
Океанское дно, на котором развивались животные, было совсем не похоже на дно фанерозоя. Подвижные травоядные были чрезвычайно редки или отсутствовали, как и роющие черви. Следовательно, морское дно часто было покрыто водорослевым/бактериальным матом. Большая часть ила и донных илов фанерозоя является продуктом постоянного вспенивания животной жизнью. Точно так же многие из твердых субстратов были созданы кораллами и другими кальцифицирующими животными. До появления животных обычное дно представляло собой прочный живой бактериальный покров с редкими, обычно небольшими выходами карбоната, построенными кальцифицирующими бактериями. Дорнбос и др. (2005).
У первых многоклеточных животных отсутствовали какие-либо очевидные способы передвижения, а также отсутствовали очевидные рот, кишечник или центральная полость.