
Содержание
Ароморфоз | Дистанционные уроки
31-Окт-2012 | Нет комментариев | Лолита Окольнова
Ароморфоз — прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов. Ароморфоз — это расширение жизненных условий, связанное с усложнением организации и повышением жизнедеятельности
Много умных слов, поэтому давайте разбирать на примерах.
Во-первых, это эволюционное изменение. Это означает, что это не просто небольшое изменение дочернего организма по отношению к родительскому. Это серьезное изменение в строении, которое привело к появлению абсолютно новых признаков.
Во-вторых, когда мы говорим об изменениях, то важно, с чем мы сравниваем. Так вот, в школьном курсе биологии когда речь идет об ароморфозах какого-то отдела ( у растений) или класса ( у животных), то сравнивают с тем, от кого этот класс или отдел произошел. Например, если говорят об ароморфозах млекопитающих, то сравнивают с рептилиями.
Например, если говорят об ароморфозах млекопитающих, то сравнивают с рептилиями.
Примеры ароморфозов растений:
От водорослей к наземным растениям (псилофитам и риниофитам или к мхам):
- появление тканей как таковых , а конкретнее, проводящих тканей, покровных и механических;
- появление вегетативных органов ( стебля, подобия листьев)
- размножение в наземной среде, но все еще требуется присутствие воды;
Ароморфозы хвощей и плаунов
- усовершенствование проводящей ткани, механической, появляется запасающая ткань;
- переход (ароморфоз) от ризоидов к корням и корневищам;
- размножение хотя еще и спорами, но уже без участия воды.
Ароморфозы папоротников
- появление сосудистой системы;
- более развитая корневая система;
- появление более развитых листьев (вайя), а соответственно, улучшение интенсивности фотосинтеза;
- преобладание в цикле развития спорофита над гаметофитом
Ароморфозы голосеменных растений
- здесь у нас глобальный «скачок» — переход от размножения спорами к размножению семенами, если подробнее, то получаем вот такой список:
- усовершенствование сосудистой системы (появление трахеид)
- бОльшее количество жизненных форм ( в основном, древесные)
Ароморфозы покрытосеменных
- У покрытосеменных самая развитая проводящая система
Примеры ароморфозов животных:
Здесь посложнее будет — надо рассмотреть все критерии: и внешнее, и внутреннее строение, дыхание, способы размножения и т. д.
д.
Кишечнополостные | Ароморфозов будет много, т.к. эволюционно это был огромный шаг — переход от одноклеточности к многоклеточности
|
Плоские черви | Считается, что они произошли от кишечнополостных, поэтому сравнивать будем с ними
|
Круглые черви | Теорий происхождения круглых червей несколько, но в школьном курсе считаем, что они произошли от плоских.
|
Кольчатые черви | Произошли от червей, похожих на плоских, поэтому сравнивать будем с ними
Как видите, ароморфозов много и по сравнению с круглыми червями, но если в открытом вопросе написать неправильно происхождение кольчатых, то за это снимут балл — это будет биологическая ошибка! |
Членистоногие | Их предками считаются кольчатые черви
|
Моллюски | От кого они произошли — ученые до сих пор яростно спорят…
|
Хордовые | |
Подтип бесчерепные (ланцетник) |
|
Рыбы |
|
Земноводные (амфибии) | Земноводные произошли от кистеперых рыб,
|
Рептилии (пресмыкающиеся) | Предками рептилий считают древних земноводных
|
Птицы | “ у каждой птицы есть гены динозавров”
|
Млекопитающие | Предполагается, что предком были рептилии
|

 — головоногие)
— головоногие) к. сердце и система кровообращения головастика — как у рыб)
к. сердце и система кровообращения головастика — как у рыб) к. у них появилась полная перегородка в сердце, которое, кстати, четырехкамерное, а следовательно, разделение артериальной и венозной крови
к. у них появилась полная перегородка в сердце, которое, кстати, четырехкамерное, а следовательно, разделение артериальной и венозной крови
- Подписка на рассылку
Обсуждение: «Примеры ароморфозов»
(Правила комментирования)
По поводу ароморфозов, идиоадаптаций и дегенераций
Здравствуйте, уважаемые читатели блога репетитора биологии по Скайпу biorepet-ufa. ru. Продолжаю рубрику «Из диалогов в комментариях». Накопилось много комментариев к статье «Биологический прогресс».
ru. Продолжаю рубрику «Из диалогов в комментариях». Накопилось много комментариев к статье «Биологический прогресс».
Биологический прогресс, как одно из направлений эволюции, достигается тремя путями (или характеризуется тремя формами): ароморфозом, идиоадаптацией и общей (филогенетической) дегенерацией.
В КИМах ОГЭ и ЕГЭ по биологии встречается много заданий по выявлению знаний учащимися этих трех характеристик биологического прогресса.
Очень любят авторы-составители КИМов такие задания. Любят, не смотря на то, что в пособиях разных авторов одни и те же примеры трактуются по-разному. Поэтому и накопилось довольно много вопросов у преподавателей и учащихся к моей статье «Биологический прогресс».
Светлана: Всегда считала эту тему не сложной. Но, оказывается всё не так просто… Встретился вопрос, какие из примеров иллюстрируют общую дегенерацию (три верных ответа из шести):
1) сокращение числа пальцев до двух у страусов; 2) упрощение нервной системы у ленточных червей; 3) превращение корней у повилики в присоски; 4) развитие детёнышей млекопитающих в матке; 5) редукция околоцветника, листьев, сосудистой системы у ряски; 6) отсутствие конечностей у змей.
Ответы 2 и 3 для меня очевидны, точно знаю что не 4, а остальное ?
Б.Ф.: Для характеристики общей дегенерации, вероятно, хорошими можно считать такие правила:
1) переход от активного образа жизни к более пассивному; 2) переход к паразитическому образу жизни; 3) возможности для большего использования готовых энергетических материалов.
Считаю, как и Вы, ответы 2 и 3 очевидными — характеризуют переход организмов к паразитизму. Наверное еще и 5, так как 1 и 6 — идиоадаптации, 4 — ароморфоз.
Светлана: Я так же думаю, как и вы. Но прочитала в пособии под редакцией Гусева, Каменского издательство Московского университета, что дегенерация рассматривается как переход к оседлому образу жизни, или жизни в экстремальных условиях (я поэтому и ряску отнесла к дегенерации, потому что для покрытосеменного растения вода это экстремальные условия). Всё вроде бы ничего.
Но там приводятся примеры дегенераций: превращение листьев кактуса в колючки, редукция задних конечностей китообразных и ещё ряд примеров, которые я всегда относила к идиоадаптациям… Короче я в расстройстве. Я никудышный биолог…
Я никудышный биолог…
Б.Ф.: Светлана, «не надо спешить с выводАми». Вы отличный биолог хотя бы уже потому, что находите множество всякого несоответствия не только в тестовых заданиях, но и в учебниках. Вот что посоветовать школьникам, как им готовиться к сдаче экзаменов, если даже учебники «врут»?
Светлана: Приведите, пожалуйста, примеры дегенераций с возможностью использования готовых энергетических материалов.
Б.Ф.: Растения паразиты утратили хлоропласты и стали питаться готовыми органическими веществами растения-хозяина, а не синтезировать их сами. (Не путать этот пример дегенерации с хищными растениями, как пример идиоадаптации, которые лишь частично утратили способность к фотосинтезу).
Сюда следует отнести и паразитических червей, утративших пищеварительную систему, перешедших фактически на осмотрофное питание всей поверхностью тела.
Светлана: Вот что сегодня обнаружила в книжице Калиновой, Петросовой, Никишовой отличник ЕГЭ по биологии 2010 (стр. 41):
41):
«следует помнить, что редукция органов возможна и при идиоадаптациях, и дегенерациях, но в первом случае изменения носят частный характер, а во втором кардинальный. Если у животных утрачиваются важные системы органов, то эволюция происходит по пути дегенерации, в противоположность этому отсутствие конечностей у змей, это изменение способа передвижения, свойственное небольшой группе животных в связи с образом жизни, и оценивать его следует как идиоадаптацию».
Б.Ф.: Только теперь остаётся сообразить какие изменения носят частный характер, а какие кардинальный?
Марина: как Вы можете объяснить, почему утрата зрения у крота — это общая дегенерация, а отсутствие конечностей у китов — это идиоадаптация. Общая дегенерация возникает при переходе организма в более простую среду обитания, а по сравнению с наземно-воздушной — водная, более простая среда; сопровождается утратой органов — так и есть утратили. В чем разница?
Б. Ф.: утрата зрения кротами и всеми остальными «земельными слепышами» только в половине руководств определяется как основной пример общей дегенерации, а в половине — как идиоадаптации.
Ф.: утрата зрения кротами и всеми остальными «земельными слепышами» только в половине руководств определяется как основной пример общей дегенерации, а в половине — как идиоадаптации.
Я считаю, что утрата зрения — это серьёзно, потеря эволюционно очень важного анализатора. К тому же это явно привело и к менее подвижному существованию (значит дегенерация).
А утрата за ненадобностью задних конечностей у китообразных (только задних) никак не может быть примером дегенерации. Бывшие парнокопытные идеально приспособились к жизни в водной среде, видоизменив конечности, способны активно передвигаться в ней — это бесспорно пример идиоадаптации.
К тому же, основных критериев для дегенерации все же два: переход организмов к паразитическому или к малоподвижному образу жизни.
Если Вы, Марина, репетитор по биологии, то Ваш вопрос очень злободневный. Как мы должны готовить своих учеников для получения высоких баллов на ЕГЭ? Не можем же мы знать заранее, какой ответ «заложили» как правильный авторы вопроса экзаменационных заданий.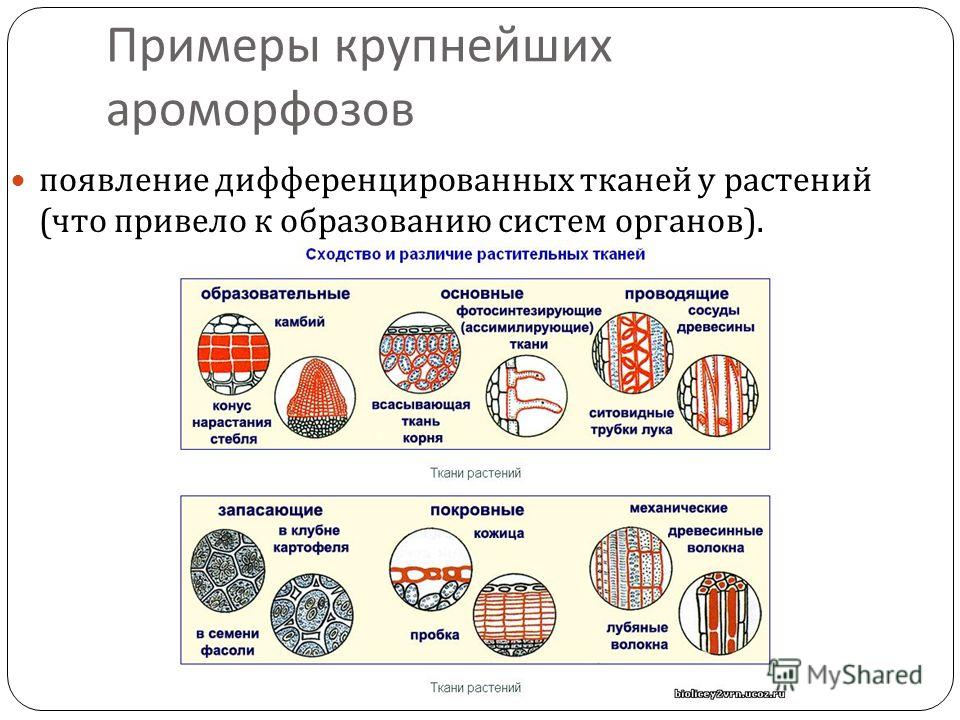 По честному, никакие спорные моменты (как, например, утрата зрения у кротов) вообще не должны попадать в вопросы экзаменационных работ, но они, к сожалению, продолжают попадать!
По честному, никакие спорные моменты (как, например, утрата зрения у кротов) вообще не должны попадать в вопросы экзаменационных работ, но они, к сожалению, продолжают попадать!
Марина: Большое спасибо за ответ. Будем надеяться, что спорных вопросов в этом году будет меньше.
Людмила: Возникли сомнения в следующем вопросе. Укажите направление эволюции, которое привело к возрастанию численности многих видов костных рыб?
1) ароморфоз; 2) дегенерация; 3) биологический прогресс; 4) биологический регресс.
Мне кажется, если говорить о том, что приводит к возрастанию численности, как одному из проявлений биопрогресса, то в данном случае это ароморфоз. Но, учитывая, что это часть А, авторы вполне могут иметь в виду сам биопрогресс?
Б.Ф.: Здесь и сомневаться не стоит. Направления эволюции: биологический прогресс, биологический регресс, стабилизация. А вот пути достижения (или формы) биологического прогресса: ароморфоз, идиоадаптация, общая дегенерация. Поэтому здесь правильный ответ 3.
Поэтому здесь правильный ответ 3.
Ира: В чем заключается разница между понятиями «ароморфоз» и «алломорфоз» ?
Б.Ф.: Ароморфоз, алломорфоз (или идиоадаптация) и общая дегенерация — это три формы (три пути) биологического прогресса.
Ароморфозами называют крупные эволюционные достижения организмов, приводящие к формированию новых крупных таксонов: царств, типов, классов. Это, как правило, усложнения строения и функций организмов не являющиеся приспособлением к конкретным условиям среды, но носящие самый общий характер, с явным повышением уровня организации и возможным выходом в другую адаптивную зону (возникновение эукариотической клетки, многоклеточности, автотрофности).
Алломорфозами (идиоадаптациями) называют приспособления организмов к определенным условиям существования — адаптации в узком смысле, или идиоадаптации, посредством которых организмы из поколения в поколение приспосабливаются к разнообразным условиям жизни, причем сложность их
организации и интенсивность жизнедеятельности в общем не повышаются. Алломорфозы протекают на уровне семейств, родов или видов. Например, покровительственная окраска у животных, видоизменения вегетативных органов растений.
Алломорфозы протекают на уровне семейств, родов или видов. Например, покровительственная окраска у животных, видоизменения вегетативных органов растений.
Катерина: Вот раньше постоянно путалась с регрессом и дегенерацией…они почему-то упорно в голове были соединены в одно целое. Теперь поняла. Дегенерация — это прогресс…просто он достигается путём упрощения общей организации, исчезновения чего-то НЕнужного…так? Тогда сюда же можно отнести редукцию мышц ушных раковин у человека, третье веко и копчик (это всё рудименты) , но они относятся к дегенерации или нет?
Б.Ф.: Нет, эти примеры не являются результатом общей дегенерации — это примеры идиоадаптаций, то есть «идиоадаптация» здесь должна рассматриваться как термин, характеризующий само явление, сам процесс. А названия этих рудиментарных органов — это результат идиоадаптации.
Под «дегенерацией» лучше всего запоминаются примеры приспособленности паразитических организмов. У них произошла утрата (дегенерация) некоторых органов наряду с развитием каких-либо дополнительных приспособлений. Ведь все признаки биологического прогресса у них «налицо». Высокие показатели и скорости размножения, и плодовитости, и численности.
Ведь все признаки биологического прогресса у них «налицо». Высокие показатели и скорости размножения, и плодовитости, и численности.
Марина: Почему всё же утрату зрения у крота относят к дегенерации? Ведь это не связано с паразитизмом или переходом к неподвижному образу жизни.
Б.Ф.: В третьей колонке таблицы Т.Л. Богдановой приведены лишь примеры дегенерации, связанные с переходом организмов к паразитическому существованию.
Но, если обратить внимание на первую строку колонки «Признаки»: «Какими преобразованиями сопровождаются», мы видим, что в целом, филогенетическая (общая) дегенерация связана с «возникновением приспособлений к более простым условиям жизни». Поэтому редукция зрения и даже почти полное отсутствие глаз у самых различных животных, обитающих в темноте: подземные животные (крот), пещерные (протей), глубоководные — это наглядные примеры перехода к менее подвижному образу жизни, значит это примеры общей дегенерации.
Марина: Спасибо за ответ!
Может быть полезным сравнение понятий биологический и морфофизиологический прогресс, а также биологический и морфофизиологический регресс. Биологический прогресс может быть достигнут с помощью морфофизиологического регресса (общей дегенерации — упрощения строения).
Дарья: Хищные растения — это результат ароморфоза или идиоадаптации?
Б.Ф.: По определению ароморфоз — это прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов. Идиоадаптация — это наиболее распространенный путь эволюции, при котором у организмов развиваются приспособления к определенным, частным условиям среды.
У насекомоядных (хищных) растений нет какого-то крупного «общего повышения уровня организации», то есть переход к хищничеству — это явно не ароморфоз.
Живя в болотистой местности, при дефиците азота и фосфора, они, не утратив полностью способности к фотосинтезу (производству углеводов из углекислого газа воздуха и воды), выработали особенный (хищный) способ дополнительного обеспечения себя дефицитными элементами питания — значит это пример идиоадаптации.
Алексей: Помогите, пожалуйста, разобраться. Специализация — это тип приспособления или путь эволюции?
Я считал, что специализация это крайняя степень проявления идиоадаптации. Отвечал на вопрос, где было изображение колибри на цветке. Надо было определить тип приспособления, форму естественного отбора и путь эволюции. В ответах: тип приспособления — специализация, а путь — идиоадаптация, форма отбора — движущий (тут мне все понятно).
Б.Ф.: Посмотрите табличку с определением ароморфоза. Понятно, что поскольку ароморфозы, идиоадаптации (алломорфозы) и общая дегенерация являются путями биологического прогресса, то термин «специализация» надо рассматривать как тип приспособления.
***************************************
У кого есть вопросы по статье к репетитору биологии по Скайпу, замечания, пожелания — прошу в комментарии.
Для подготовки к сдаче ЕГЭ или ОГЭ, у меня на блоге вы можете приобрести ответы на все тесты Открытого Банка Заданий ФИПИ за все годы проведения экзаменов по ЕГЭ и ОГЭ (ГИА).
SCIRP Открытый доступ
Издательство научных исследований
Журналы от A до Z
Журналы по темам
- Биомедицинские и биологические науки.
- Бизнес и экономика
- Химия и материаловедение.
- Информатика. и общ.
- Науки о Земле и окружающей среде.
- Машиностроение
- Медицина и здравоохранение
- Физика и математика
- Социальные науки. и гуманитарные науки
Журналы по тематике
- Биомедицина и науки о жизни
- Бизнес и экономика
- Химия и материаловедение
- Информатика и связь
- Науки о Земле и окружающей среде
- Машиностроение
- Медицина и здравоохранение
- Физика и математика
- Социальные и гуманитарные науки
Публикация у нас
- Представление статьи
- Информация для авторов
- Ресурсы для экспертной оценки
- Открытые специальные выпуски
- Заявление об открытом доступе
- Часто задаваемые вопросы
Публикуйте у нас
- Представление статьи
- Информация для авторов
- Ресурсы для экспертной оценки
- Открытые специальные выпуски
- Заявление об открытом доступе
- Часто задаваемые вопросы
Подпишитесь на SCIRP
Свяжитесь с нами
| клиент@scirp. org | |
| +86 18163351462 (WhatsApp) | |
| 1655362766 | |
| Публикация бумаги WeChat |
| Недавно опубликованные статьи |
| Недавно опубликованные статьи |
Подпишитесь на SCIRP
Свяжитесь с нами
клиент@scirp. org org | |
| +86 18163351462 (WhatsApp) | |
| 1655362766 | |
| Публикация бумаги WeChat |
Бесплатные информационные бюллетени SCIRP
Copyright © 2006-2022 Scientific Research Publishing Inc. Все права защищены.
верхний
Эволюция хордовых и система трех типов
1. Дарвин К.
1859 г.
О происхождении видов. Лондон, Великобритания: Джон Мюррей. [Google Scholar]
2. Геккель Э.
1874 г.
Антропогения oder Entwickelungsgeschichte des Menschen. Лейпциг, Германия: Verlag von Wilhelm Engelmann. [Академия Google]
3. Бейтсон В.
1886 г.
Родословная хордовых. Q. J. Microsc. науч.
науч.
26, 535–571. [Google Scholar]
4. Гарстанг В.
1928 год.
Происхождение и эволюция личиночных форм. Представитель бр. доц. Доп. науч. №, 1928, 77–98. [Google Scholar]
5. Беррилл, Нью-Джерси.
1955.
Происхождение позвоночных. Оксфорд, Великобритания: Издательство Оксфордского университета. [Google Scholar]
6. Джеффрис RPS.
1986 год.
Родословная позвоночных. Лондон, Великобритания: Британский музей естественной истории. [Академия Google]
7. Шеффер Б.
1987.
Монофилия и филогения вторичноротых. Эвол. биол.
21, 179–235. ( 10.1007/978-1-4615-6986-2_8) [CrossRef] [Google Scholar]
8. Gee H.
1996.
До позвоночника: взгляды на происхождение позвоночных. Лондон, Великобритания: Чепмен и Холл. [Google Scholar]
9. Холл Б.К.
1999.
Эволюционная биология развития, 2-е изд.
Берлин, Германия: Springer. [Google Scholar]
10. Нильсен К.
1999.
Происхождение центральной нервной системы хордовых и происхождение хордовых. Дев. Гены Эвол.
209, 198–205. ( 10.1007/s004270050244) [PubMed] [CrossRef] [Google Scholar]
( 10.1007/s004270050244) [PubMed] [CrossRef] [Google Scholar]
11. Shimeld SM, Holland PW.
2000.
Позвоночные инновации. проц. Натл акад. науч. США
97, 4449–4452. ( 10.1073/pnas.97.9.4449) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Cameron CB, Garey JR, Swalla BJ.
2000.
Эволюция плана тела хордовых: новые данные филогенетического анализа типов вторичноротых. проц. Натл акад. науч. США
97, 4469–4474. ( 10.1073/pnas.97.9.4469) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Сато Н.
2003.
Личинка головастика асцидии: сравнительное молекулярное развитие и геномика. Нац. Преподобный Жене.
4, 285–295. ( 10.1038/nrg1042) [PubMed] [CrossRef] [Google Scholar]
14. Zeng L, Swalla BJ.
2005.
Молекулярная филогения протохордовых: эволюция хордовых. Можно. Дж. Зул.
83, 24–33. ( 10.1139/z05-010) [CrossRef] [Google Scholar]
15. Lacalli TC.
2005.
План тела протохордовых и эволюционная роль личинок: старые споры разрешены?
Можно. Дж. Зул.
Дж. Зул.
83, 216–224. ( 10.1139/z04-162) [CrossRef] [Google Scholar]
16. Герхарт Дж.
2006.
Предок вторичноротых. Дж. Селл. Физиол.
209, 677–685. ( 10.1002/jcp.20803) [PubMed] [CrossRef] [Google Scholar]
17. Браун Ф.Д., Прендергаст А., Свалла Б.Дж.
2008.
Человек всего лишь червь: происхождение хордовых. Бытие
46, 605–613. ( 10.1002/dvg.20471) [PubMed] [CrossRef] [Google Scholar]
18. Сато Н.
2008.
Гипотеза аборальной дорсализации происхождения хордовых. Бытие
46, 614–622. ( 10.1002/dvg.20416) [PubMed] [CrossRef] [Google Scholar]
19. Свалла Б.Дж., Смит А.Б.
2008.
Расшифровка филогении вторичноротых: молекулярная, морфологическая и палеонтологическая перспективы. Фил. Транс. Р. Соц. Б
363, 1557–1568. ( 10.1098/rstb.2007.2246) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Nielsen C.
2012.
Эволюция животных: взаимосвязь живых типов, 3-е изд.
Нью-Йорк, штат Нью-Йорк: Издательство Оксфордского университета. [Google Scholar]
21. Линней К.
Линней К.
1766–1767 гг.
Systema naturae, 12-е изд., том. 1
Holmiae, Швеция: Salvius. [Академия Google]
22. Ламарк Дж.Б.
1794.
Recherches sur les причины des principaux faits physiques. Париж, Франция: Марадан. [Google Scholar]
23. Кювье Г.
1815.
Mémoire sur les Ascidies et sur leur anatomie. Мем. Мус. История Нац. Париж
2, 10–39. [Google Scholar]
24. Ламарк Дж.Б.
1816.
Histoire naturelle des animaux sans vertebres, vol. III: Тунисьеры
Париж, Франция: Детервиль. [Google Scholar]
25. Яррелл В.
1836.
История британских рыб, том. 2
Лондон, Великобритания: Джон Ван Ворст. [Академия Google]
26. Ковалевский А.
1866 г.
Entwicklungsgeschichte дер einfachen Ascidien. Мем. л’акад. Санкт-Петербург.
Сер. . 7,
10, 11–19. [Google Scholar]
27. Ковалевский А.
1867 г.
Entwicklungsgeschichte des Amphioxus lanceolatus . Мем. л’акад. Санкт-Петербург.
Сер. . 7,
4, 1–17. [Google Scholar]
28. Геккель Э.
1866 г.
Общая морфология организмов. Берлин, Германия: Verlag von Georg Reimer. [Google Scholar]
Берлин, Германия: Verlag von Georg Reimer. [Google Scholar]
29. Геккель Э.
1874 г.
Die Gastrea-Theorie, die phylogenetische Classification des Thierreichs und die Homologie der Keimblatter. Jenaische Zischr Naturw
8, 1–55. [Академия Google]
30. Геккель Э.
1894 г.
Систематическая филогения. Берлин, Германия: Verlag von Georg Reimer. [Google Scholar]
31. Lankester ER.
1877 г.
Заметки об эмбриологии и классификации животного мира: пересмотр предположений о происхождении и значении зародышевых листков. Q. J. Microsc. науч.
17, 399–454. [Google Scholar]
32. Balfour FM.
1880.
Трактат по сравнительной эмбриологии. Лондон, Великобритания: Макмиллан. [Google Scholar]
33. Гроббен К.
1908.
Die systematische Einteilung des Tierreiches. Вер. Зоол. Бот. Гэс. Вена
58, 491–511. [Google Scholar]
34. Агинальдо А.М., Турбевилль Дж.М., Линфорд Л.С., Ривера М.С., Гэри Дж.Р., Рафф Р.А., Лейк Дж.А.
1997.
Доказательства наличия клады нематод, членистоногих и других линяющих животных. Природа
Природа
387, 489–493. ( 10.1038/387489a0) [PubMed] [CrossRef] [Google Scholar]
35. Adoutte A, Balavoine G, Lartillot N, Lespinet O, Prud’homme B, de Rosa R.
2000.
Новая филогения животных: достоверность и значение. проц. Натл акад. науч. США
97, 4453–4456. ( 10.1073/pnas.97.9.4453) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Dunn MP, et al.
2008.
Широкая филогеномная выборка улучшает разрешение дерева жизни животных. Природа
452, 745–749. ( 10.1038/nature06614) [PubMed] [CrossRef] [Google Scholar]
37. Philippe H, et al.
2009.
Филогеномика возрождает традиционные взгляды на глубинные взаимоотношения животных. Курс. биол.
19, 706–712. ( 10.1016/j.cub.2009.02.052) [PubMed] [CrossRef] [Google Scholar]
38. Вада Х, Сато Н.
1994.
Подробности истории эволюции от беспозвоночных к позвоночным, полученные из последовательностей 18S рДНК. проц. Натл акад. науч. США
91, 1801–1804 гг. ( 10.1073/pnas.91.5.1801) [бесплатная статья ЧВК] [PubMed] [CrossRef] [Google Scholar]
39. Галаныч К.М.
Галаныч К.М.
1995.
Филогенетическое положение крыложаберных полухордовых на основе данных о последовательности 18S рДНК. Мол. Филогенет. Эвол.
4, 72–76. ( 10.1006/mpev.1995.1007) [PubMed] [CrossRef] [Google Scholar]
40. Персеке М., Голомбек А., Шлегель М., Струк Т.Х.
2013.
Влияние анализа митохондриального генома на понимание филогении вторичноротых. Мол. Филогенет. Эвол.
66, 898–905. ( 10.1016/j.ympev.2012.11.019) [PubMed] [CrossRef] [Google Scholar]
41. Мечников Е.
1881.
Über die systematische Stellung von Balanoglossus . Зоол. Анц.
4, 153–157. [Google Scholar]
42. Jefferies RPS, et al.
1996.
Ранняя филогения хордовых и иглокожих и происхождение хордовых лево-правой асимметрии и двусторонней симметрии. Акта Зоолог. (Стокгольм)
77, 101–122. ( 10.1111/j.1463-6395.1996.tb01256.x) [CrossRef] [Google Scholar]
43. Delsuc F, Brinkmann H, Chourrout D, Philippe H.
2006.
Оболочечные, а не головохордовые, являются ближайшими живыми родственниками позвоночных. Природа
Природа
439, 965–968. ( 10.1038/nature04336) [PubMed] [CrossRef] [Google Scholar]
44. Bourlat SJ, et al.
2006.
Филогения вторичноротых выявляет монофилетических хордовых и новый тип Xenoturbellida. Природа
444, 85–88. ( 10.1038/nature05241) [PubMed] [CrossRef] [Google Scholar]
45. Putnam NH, et al.
2008.
Геном амфиокса и эволюция кариотипа хордовых. Природа
453, 1064–1071. ( 10.1038/nature06967) [PubMed] [CrossRef] [Google Scholar]
46. Горман А.Л., Макрейнольдс Дж.С., Барнс С.Н.
1971.
Фоторецепторы примитивных хордовых: тонкая структура, гиперполяризующие рецепторные потенциалы и эволюция. Наука
172, 1052–1054. ( 10.1126/science.172.3987.1052) [PubMed] [CrossRef] [Google Scholar]
47. Vandekerckhove J, Weber K.
1984.
Актины хордовых мышц отчетливо отличаются от актинов мышц беспозвоночных. Эволюция различных мышечных актинов позвоночных. Дж. Мол. биол.
179, 391–413. ( 10.1016/0022-2836(84)
-X) [PubMed] [CrossRef] [Google Scholar]
48. Кусакабе Т. , Араки И., Сато Н., Джеффри В.Р.
1997.
Эволюция генов актина хордовых: данные геномной организации и аминокислотных последовательностей. Дж. Мол. Эвол.
44, 289–298. ( 10.1007/PL00006146) [PubMed] [CrossRef] [Google Scholar]
49. Oda H, et al.
2002.
Новый кадгерин amphioxus, локализующийся в эпителиальных слипчивых соединениях, имеет необычную доменную организацию, имеющую значение для филогении хордовых. Эвол. Дев.
4, 426–434. ( 10.1046/j.1525-142X.2002.02031.x) [PubMed] [CrossRef] [Google Scholar]
50. Филипп Х., Бринкманн Х., Копли Р.Р., Мороз Л.Л., Накано Х., Пустка А.Дж., Уоллберг А., Петерсон К.Дж., Телфорд М.Дж.
2011.
Ацеломорфные плоские черви — вторичноротые, родственные Xenoturbella . Природа
470, 255–258. ( 10.1038/nature09676) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Romer AS.
1967.
Основные этапы эволюции позвоночных. Наука
158, 1629 (10.1126/science.158.3809.1629) [PubMed] [CrossRef] [Google Scholar]
52. Токиока Т.
1971.
Филогенетические предположения Tunicata. Опубл. Сето Мар Биол. лаборатория
19, 43–63. [Google Scholar]
53. Сальвини-Плавен Л.
1999.
О филогенетическом значении нервно-кишечного канала (хордовые). Зоол. Анальный. Комплекс Сист.
102, 175–183. [Google Scholar]
54. Holland PW.
2000.
Эмбриональное развитие головы, скелетов и амфиокса: новый взгляд на Эдвина С. Гудрича. Междунар. Дж. Дев. биол.
44, 29–34. [PubMed] [Google Scholar]
55. Сато Н.
2009.
Усовершенствованная гипотеза фильтратора эволюции урохордовых. Зоолог. науч.
26, 97–111. ( 10.2108/zsj.26.97) [PubMed] [CrossRef] [Google Scholar]
56. Takacs CM, Moy VN, Peterson KJ.
2002.
Тестирование предполагаемых полухордовых гомологов спинной нервной системы и эндостиля хордовых: экспрессия NK2.1 ( TTF-1 ) у желудевого червя Ptychodera flava (Hemichordata, Ptychoderidae). Эвол. Дев.
4, 405–417. ( 10.1046/j.1525-142X.2002.02029.x) [PubMed] [CrossRef] [Google Scholar]
57. Holland ND.
Holland ND.
2003.
Ранняя эволюция центральной нервной системы: эпоха кожных мозгов?
Нац. Преподобный Нейроски.
4, 617–627. ( 10.1038/nrn1175) [PubMed] [CrossRef] [Google Scholar]
58. Герхарт Дж., Лоу К., Киршнер М.
2005.
Полухордовые и происхождение хордовых. Курс. мнение Жене. Дев.
15, 461–467. ( 10.1016/j.gde.2005.06.004) [PubMed] [CrossRef] [Google Scholar]
59. Ромер А.С.
1960.
Биология позвоночных, 2-е изд.
Филадельфия, Пенсильвания: Sanders & Co. [Google Scholar]
60. Арендт Д., Нублер-Юнг К.
1994.
Инверсия дорсовентральной оси?
Природа
371, 26 (10.1038/371026a0) [PubMed] [CrossRef] [Google Scholar]
61. De Robertis EM, Sasai Y.
1996.
Общий план формирования дорсовентрального паттерна у Bilateria. Природа
380, 37–40. ( 10.1038/380037a0) [PubMed] [CrossRef] [Google Scholar]
62. Дубок В., Роттингер Э., Беснардо Л., Лепаж Т.
2004.
Передача сигналов Nodal и BMP2/4 организует орально-аборальную ось эмбрионов морского ежа. Дев. Клетка
6, 397–410. (10.1016/S1534-5807(04)00056-5) [PubMed] [CrossRef] [Google Scholar]
(10.1016/S1534-5807(04)00056-5) [PubMed] [CrossRef] [Google Scholar]
63. Lowe CJ, et al.
2006.
Дорсовентральные узоры у полухордовых: взгляд на раннюю эволюцию хордовых. PLoS биол.
4, е291 ( 10.1371/journal.pbio.0040291) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
64. Yu JK, Satou Y, Holland ND, Shin-I T, Kohara Y, Satoh N, Bronner -Фрейзер М., Голландия Л.З.
2007.
Осевое формирование у головохордовых и эволюция организатора. Природа
445, 613–617. ( 10.1038/nature05472) [PubMed] [CrossRef] [Google Scholar]
65. Каул С., Стах Т.
2010.
Онтогенез воротникового канатика: нейруляция полухордовых Saccoglossus kowalevskii . Дж. Морфол.
271, 1240–1259 гг.. ( 10.1002/jmor.10868) [PubMed] [CrossRef] [Google Scholar]
66. Luttrell S, Konikoff C, Byrne A, Bengtsson B, Swalla BJ.
2012.
Птиходеридная полухордовая нейруляция без хорды. интегр. Комп. биол.
52, 829–834. ( 10.1093/icb/ics117) [PubMed] [CrossRef] [Google Scholar]
67. Миямото Н. , Вада Х.
, Вада Х.
2013.
Полухордовая нейруляция и происхождение нервной трубки. Нац. коммун.
4, 2713 (10.1038/ncomms3713) [PubMed] [CrossRef] [Google Scholar]
68. Spemann H, Mangold H.
1924.
Индукция эмбриональных зачатков имплантацией организаторов другого вида. Арка Ру. Entw. мех.
100, 599–638. [PubMed] [Google Scholar]
69. Огасавара М., Вада Х., Питерс Х., Сато Н.
1999.
Экспрессия генов Pax1/9 в жабрах урохордовых и полухордовых: понимание функции и эволюции эпителия глотки. Разработка
126, 2539–2550. [PubMed] [Google Scholar]
70. Гиллис Дж. А., Фритценванкер Дж. Х., Лоу С. Дж.
2012.
Стволово-второротовое происхождение транскрипционной сети глотки позвоночных. проц. Р. Соц. Б
279, 237–246. ( 10.1098/rspb.2011.0599) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
71. Satoh N, et al.
Поданный. О возможной эволюционной связи хорды желудка полухордовых с глоточными органами хордовых. [Бесплатная статья PMC] [PubMed]
72. Гонт Л. К., Штайнбайссер Х., Блумберг Б., де Робертис Э.М.
К., Штайнбайссер Х., Блумберг Б., де Робертис Э.М.
1993.
Формирование хвоста как продолжение гаструляции: множественные клеточные популяции хвостовой почки Xenopus происходят из губы позднего бластопора. Разработка
119, 991–1004. [PubMed] [Google Scholar]
73. Маргулис Л., Шварц К.В.
1998.
Пять королевств, 3-е изд.
Нью-Йорк, штат Нью-Йорк: Freeman & Co. [Google Scholar]
74. Brusca RC, Brusca GJ.
2003.
Беспозвоночные, 2-е изд.
Сандерленд, Массачусетс: Sinauer Associates. [Google Scholar]
75. Рупперт Э.Э., Фокс Р.С., Барнс Р.Д.
2004.
Зоология беспозвоночных: функциональный эволюционный подход, 7-е изд.
Бельмонт, Калифорния: Томсон-Брукс/Коул. [Google Scholar]
76. Вада Х., Сайга Х., Сатох Н., Холланд П.В.
1998.
Трехсторонняя организация мозга предковых хордовых и древность плакод: выводы из генов асцидий Pax-2/5/8, Hox и Otx . Разработка
125, 1113–1122. [PubMed] [Google Scholar]
77. Бертран С., Эскрива Х.
2011.
Эволюционный перекресток в биологии развития: амфиокс. Разработка
Разработка
138, 4819–4830. ( 10.1242/dev.066720) [PubMed] [CrossRef] [Google Scholar]
78. Holland LZ.
2013.
Эволюция новых признаков после дупликации всего генома: взгляды амфиокса. Семин. Сотовый Дев. биол.
24, 101–109. ( 10.1016/j.semcdb.2012.12.007) [PubMed] [CrossRef] [Google Scholar]
79. Нохара М., Нисида М., Мия М., Нисикава Т.
2005.
Эволюция митохондорального генома у головоногих, как следует из полных нуклеотидных последовательностей двух видов Epigonichthys . Дж. Мол. Эвол.
60, 526–537. ( 10.1007/s00239-004-0238-x) [PubMed] [CrossRef] [Google Scholar]
80. Kon T, Nohara M, Yamanoue Y, Fujiwara Y, Nishida M, Nishikawa T.
2007.
Филогенетическое положение китового ланцетника (Cephalochordata), выведенное из полных последовательностей митохондриального генома. БМС Эвол. биол.
7, 127 ( 10.1186/1471-2148-7-127) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
81. Шу Д., Моррис С., Чжан С.
1996.
Хордовое животное, похожее на Pikaia , из нижнего кембрия Китая. Природа
Природа
348, 157–158. ( 10.1038/384157a0) [CrossRef] [Google Scholar]
82. Конвей Моррис С.
1982.
Атлас сланцев Берджесс. Лондон, Великобритания: Палеонтологическая ассоциация. [Google Scholar]
83. Somorjai I, Bertrand S, Camasses A, Haguenauer A, Escriva H.
2008.
Доказательства застоя, а не генетического пиратства в паттернах экспрессии развития Branchiostoma lanceolatum и Branchiostoma floridae , два вида амфиоксусов, которые развивались независимо в течение 200 млн лет. Дев. Гены Эвол.
218, 703–713. ( 10.1007/s00427-008-0256-6) [PubMed] [CrossRef] [Google Scholar]
84. Ruppert EE.
1997.
Головохордовые (Акрании) . В Микроскопическая анатомия беспозвоночных, том. 15: Hemichordata, Chaetognatha и беспозвоночные хордовые (ред. Harrison FW, Ruppert EE.), стр. 349–504. Нью-Йорк, штат Нью-Йорк: Уайли-Лисс. [Академия Google]
85. Судзуки М.М., Сато Н.
2000.
Гены, экспрессированные в хорде амфиоксуса, выявленные с помощью анализа EST. Дев. биол.
Дев. биол.
224, 168–177. ( 10.1006/dbio.2000.9796) [PubMed] [CrossRef] [Google Scholar]
86. Сато Н.
2014.
Геномика развития асцидий. Нью-Йорк, штат Нью-Йорк: Уайли Брэквелл. [Google Scholar]
87. Swalla BJ, Cameron CB, Corley LS, Garey JR.
2000.
Урохордовые монофилетичны среди вторичноротых. Сист. биол.
49, 122–134. [PubMed] [Академия Google]
88. Нишино А., Сато Н.
2001.
Простой хвост хордовых: филогенетическое значение аппендикулярий. Бытие
29, 36–45. ( 10.1002/1526-968X(200101)29:1<36::AID-GENE1003>3.0.CO;2-J) [PubMed] [CrossRef] [Google Scholar]
89. Стач Т., Винтер Дж., Букет Дж.М. , Чуррут Д., Шнабель Р.
2008.
Эмбриология планктонных оболочников обнаруживает следы сидячей формы. проц. Натл акад. науч. США
105, 7229–7234. ( 10.1073/pnas.0710196105) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
90. Шу Д.Г., Чен Л., Хань Дж., Чжан XL.
2001.
Раннекембрийский оболочник из Китая. Природа
411, 472–473. ( 10.1038/35078069) [PubMed] [CrossRef] [Google Scholar]
91. Nakashima K, Yamada L, Satou Y, Azuma J, Satoh N.
Nakashima K, Yamada L, Satou Y, Azuma J, Satoh N.
2004.
Эволюционное происхождение синтазы целлюлозы животных. Дев. Гены Эвол.
214, 81–88. ( 10.1007/s00427-003-0379-8) [PubMed] [CrossRef] [Google Scholar]
92. Matthysse AG, Deschet K, Williams M, Marry M, White AR, Smith WC.
2004.
Функциональная синтаза целлюлозы из эпидермиса асцидий. проц. Натл акад. науч. США
101, 986–991. ( 10.1073/pnas.0303623101) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
93. Sasakura Y, Nakashima K, Awazu S, Matsuoka T, Nakayama A, Azuma J-I, Satoh N.
2005.
Опосредованный транспозонами инсерционный мутагенез выявил функции синтазы целлюлозы животных у асцидий Ciona кишечная . проц. Натл акад. науч. США
102, 15 134–15 139. ( 10.1073/pnas.0503640102) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
94. Ahlberg PE.
2001.
Основные события ранней эволюции позвоночных. Лондон, Великобритания: Тейлор и Фрэнсис. [Академия Google]
95. Кардонг К.В.
2009.
Позвоночные: сравнительная анатомия, функции, эволюция, 5-е изд.
Бостон, Массачусетс: Высшее образование McGraw-Hill. [Google Scholar]
96. Venkatesh B, et al.
2014.
Геном слоновой акулы дает уникальное представление об эволюции челюстноротых. Природа
505, 174–179. ( 10.1038/nature12826) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
97. Yu JK.
2010.
Эволюционное происхождение нервного гребня позвоночных и регуляторной сети его генов развития — взгляды амфиоксуса. Зоология (Йена)
113, 1–9. ( 10.1016/j.zool.2009.06.001) [PubMed] [CrossRef] [Google Scholar]
98. Simakov O, et al.
2013.
Взгляд на билатеральную эволюцию из трех спиралевидных геномов. Природа
493, 526–531. ( 10.1038/nature11696) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
99. Gans C, Northcutt RG.
1983.
Нервный гребень и происхождение позвоночных: новая голова. Наука
220, 268–273. ( 10.1126/science.220.4594.268) [PubMed] [CrossRef] [Google Scholar]
100. Шигетани Ю., Сугахара Ф., Куратани С.
Шигетани Ю., Сугахара Ф., Куратани С.
2005.
Новый эволюционный сценарий челюсти позвоночных. Биоэссе
27, 331–338. ( 10.1002/bies.20182) [PubMed] [CrossRef] [Google Scholar]
101. Ле Дуарен Н., Кальхейм К.
1999.
Нервный гребень, 2-е изд.
Кембридж, Великобритания: Издательство Кембриджского университета. [Google Scholar]
102. Меулеманс Д., Броннер-Фрейзер М.
2004.
Генно-регуляторные взаимодействия в эволюции и развитии нервного гребня. Дев. Клетка
7, 291–299. (10.1016/j.devcel.2004.08.007) [PubMed] [CrossRef] [Google Scholar]
103. Jeffery WR, Strickler AG, Yamamoto Y.
2004.
Мигрирующие клетки, похожие на нервный гребень, образуют пигментацию тела у урохордовых эмбрионов. Природа
431, 696–699. ( 10.1038/nature02975) [PubMed] [CrossRef] [Google Scholar]
104. Abitua PB, Wagner E, Navarrete IA, Levine M.
2012.
Идентификация рудиментарного нервного гребня у беспозвоночных хордовых. Природа
492, 104–107. ( 10.1038/nature11589) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
105. Boehm T.
Boehm T.
2012.
Эволюция иммунитета позвоночных. Курс. биол.
22, Р722–Р732. (10.1016/j.cub.2012.07.003) [PubMed] [CrossRef] [Google Scholar]
106. Azumi K, et al.
2003.
Геномный анализ иммунитета у урохордовых и появление иммунной системы у позвоночных: «в ожидании Годо». Иммуногенетика
55, 570–581. ( 10.1007/s00251-003-0606-5) [PubMed] [CrossRef] [Google Scholar]
107. Holland LZ, et al.
2008.
Геном амфиокса освещает происхождение позвоночных и биологию головохордовых. Геном Res.
18, 11:00–11:11. ( 10.1101/gr.073676.107) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
108. Pancer Z, Amemiya CT, Ehrhardt GR, Ceitlin J, Gartland GL, Cooper MD.
2004.
Соматическая диверсификация вариабельных рецепторов лимфоцитов бесчелюстной морской миноги. Природа
430, 174–180. ( 10.1038/nature02740) [PubMed] [CrossRef] [Google Scholar]
109. Guo P, Hirano M, Herrin BR, Li J, Yu C, Sadlonova A, Cooper MD.
2009.
Двойственный характер адаптивной иммунной системы у миног.