
Содержание
Реферат на тему Приспособление растений к водному режиму
Содержание:
- Характеристика основных групп растений по отношению к воде
- Анатомо-морфологические приспособления растений к водному режиму
- Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности
- Заключение
| Предмет: | Ботаника |
| Тип работы: | Реферат |
| Язык: | Русский |
| Дата добавления: | 04.01.2019 |
- Данный тип работы не является научным трудом, не является готовой работой!
- Данный тип работы представляет собой готовый результат обработки, структурирования и форматирования собранной информации, предназначенной для использования в качестве источника материала для самостоятельной подготовки учебной работы.


Если вам тяжело разобраться в данной теме напишите мне в whatsapp разберём вашу тему, согласуем сроки и я вам помогу!
По этой ссылке вы сможете найти рефераты по биологии на любые темы и посмотреть как они написаны:
Посмотрите похожие темы возможно они вам могут быть полезны:
Введение:
Вода является самым важным экологическим фактором для всей жизни на Земле. Для осуществления обменных процессов с окружающей средой, составляющей основу жизни, необходимо участие воды как растворителя и продукта обмена веществ. Так, в растениях вода участвует в реакции фотосинтеза, а неорганические соли поступают в растение только в виде раствора воды из почвы.
Вода является основным компонентом растительного тела. Даже в состоянии анабиоза растения содержат воду. Особую роль воды наземных растений играет постоянное пополнение больших затрат на испарение за счет развития больших фотосинтетических поверхностей. Вода, вызывающая необходимый тургор, определенным образом участвует в поддержании формы наземных растений как живых организмов, не имеющих опорного скелета. А для большой группы растений, населяющих водоемы, моря и океаны, вода является непосредственной средой обитания.
Вода, вызывающая необходимый тургор, определенным образом участвует в поддержании формы наземных растений как живых организмов, не имеющих опорного скелета. А для большой группы растений, населяющих водоемы, моря и океаны, вода является непосредственной средой обитания.
Характеристика основных групп растений по отношению к воде
Среди наземных растений различают три основных экологических типа по различным условиям увлажнения и развитию соответствующих приспособлений, а также по близости к месту обитания: гигрофиты, мезофиты и ксерофиты.
Растения, вода которых является не только необходимым экологическим фактором, но и непосредственным местом обитания, относятся к водным растениям и называются гидрофобными.
Гигрофиты. Это растения в чрезмерно влажных местообитаниях с повышенной влажностью воздуха и почвы. Благодаря достаточно широкому разнообразию местообитаний, особенностям водоемов, анатомо-морфологическим особенностям, все гигрофиты являются травянистыми растениями и эпифитами влажных тропических лесов, которые не переносят заметного снижения влажности воздуха, а также отсутствия устройств, ограничивающих водопотребление.

Даже в разгар сезона дождей мелкие эпифиты на стволах деревьев теряют свой тургор и высыхают, когда на них в течение 2-3 часов падают солнечные лучи. Отличительной особенностью Гигрофитов являются травянистые хвойные леса (щавель, майника двулопастный, двулопастный Альпийский). К гигрофитам относятся также виды, произрастающие в открытых и светлых местообитаниях, но в местах, где грунтовые воды выходят наружу, вблизи водоема, в дельте реки наступает состояние избыточной влажности почвы и в наших широтах примером могут служить прибрежные виды: Caltha palaris, Плакучая трава-Lyustrum salikaria, Ясима, в жарком климате–папирус, яшии. Из культурных растений можно включить рис, выращенный на полях, заполненных водой.
Ксерофиты. Это растения засушливой среды обитания, способные выдержать значительный дефицит воды в почве и атмосферную засуху. Они распространены, многочисленны и разнообразны в районах с жарким и сухим климатом. К этой группе относятся пустыни, засушливые степи, саванны, колючие леса и сухие субтропические виды. В более влажных районах ксерофиты участвуют в растительном покрове только в наиболее нагретых и наименее влажных местообитаниях (например, на склонах южной экспозиции).
В более влажных районах ксерофиты участвуют в растительном покрове только в наиболее нагретых и наименее влажных местообитаниях (например, на склонах южной экспозиции).
Неблагоприятный водный режим растения в засушливой среде обитания, во-первых, при недостатке влаги в почве, а во-вторых, при высокой сухости воздуха и транспирации при высоких температурах, поэтому для преодоления недостатка влаги возможны различные способы: увеличить ее поглощение, уменьшить расход и, кроме того, способность переносить большие потери воды. Все это используется ксерофитами при адаптации к сухости, но разные растения имеют разную степень, поэтому некоторые авторы предполагают, что засуха может быть вызвана ксерофитами.
В зависимости от особенностей строения и способов регулирования водного режима ксерофиты (Хенкель П. А. выделяют несколько типов): эвксениты, ремиксирующие, Поэсилотерии. В группу ксерофитов также входят суккуленты с сочными листьями и стеблями. Есть листовые суккуленты (агавы, алоэ) и стеблевые, листья редуцированы,а наземная часть представлена мясистыми стеблями (кактус, некоторые молочаи).
Вид с наиболее выраженными гетероморфными особенностями строения листьев имел уникальный внешний вид, за что и получил название склеродермии. Появление типичной склеродермии легко представить на примере чертополоха-Carduus crispus и пустынной полыни, ковыля, саксаула.
К этой группе относятся растения, произрастающие в условиях умеренной влажности, в том числе луговые растения, лесные травы, лиственные и кустарниковые виды из районов с умеренной влажностью климата, а также большинство культурных растений.
Мезофиты очень разнообразны не только с точки зрения видового состава, но и с точки зрения различных экологических оттенков, обусловленных различным сочетанием факторов в их естественной среде обитания. есть виды с повышенной влажностью, предпочитающие влажные или временно затопляемые участки (степной лисохвост-alopecurus pratensis, бекмания обыкновенная-beckmannia eruciformis).
Они объединяются в переходную группу гигромезофитов вместе с некоторыми влаголюбивыми лесными травами, которые предпочитают наиболее сырые леса, лесные ущелья (Impatiens nolitangere). С другой стороны, в местообитаниях с перемежающимся или стойким (малым) недостатком влаги многие группы мезодермы со специфической атипией с повышенной физиологической устойчивостью к засухе мигрировали между промежуточными остистыми клетками и пигментными колюче-пластиформными остистыми. Примерами могут служить многие виды северных степей, сухих сосновых лесов и песчаных местообитаний: Клевер белоголовый-Trifolium montanum, подмаренник желтый-Galium verum и др.
С другой стороны, в местообитаниях с перемежающимся или стойким (малым) недостатком влаги многие группы мезодермы со специфической атипией с повышенной физиологической устойчивостью к засухе мигрировали между промежуточными остистыми клетками и пигментными колюче-пластиформными остистыми. Примерами могут служить многие виды северных степей, сухих сосновых лесов и песчаных местообитаний: Клевер белоголовый-Trifolium montanum, подмаренник желтый-Galium verum и др.
Особое место среди Мезофитов занимают эфемериды и эфемероиды весной в степях и пустынях. К этой группе относятся растения, покрывающие Луга и пустыни красочными цветочными коврами (многолетники-тюльпаны, гуси-Луки, однолетники — маки, вероники) ранней весной. Это виды с очень короткими вегетационными периодами и длительными периодами покоя, имеющие опыт однолетних эфемероидов, таких как семена, и многолетних эфемероидов — в виде спящих луковиц, клубней, кроме весенних есть еще и осенние эфемероиды, произрастающие в регионах со средиземноморским климатическим ритмом. Это включает в себя род Крокус, Сцилла, и других видов.
Это включает в себя род Крокус, Сцилла, и других видов.
По многим особенностям строения и физиологии растения испытывают недостаток влаги, который близок к нулевому Фитоценозу и связан по каким-то причинам с действием низких температур. Иногда такие виды включаются как особое подразделение ксерофитной группы, а иногда их выделяют в самостоятельные экологические типы (сапрофиты и мерзлые грибы), также Психрофиты-в прохладную среду обитания, влажные, холодные почвы растений высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, им часто не хватает влаги (из-за физиологической сухости, вызванной низкими температурами, или из-за землистых травянистых растений, таких как торфяные почвы (например, северные луговые травы: Белоус-Nardus strikta; высокогорные травы белые), высокогорные, Фестовые и тундровые кустарники и вечнозеленые кустарники (call-evergreen). к психрофилам относятся хвойные породы умеренных и северных широт.
Экологически очень близок к психотическим грибам, связан с ними переходной формой. Это растения сухой и холодной среды обитания, сухие участки тундры, скалы и чешуи. Как обычно и предполагают, он характеризуется психрофитом не из-за его многочисленных сходных морфологических и физиологических особенностей. Однако некоторые холодные растения также имеют очень своеобразную форму.
Это растения сухой и холодной среды обитания, сухие участки тундры, скалы и чешуи. Как обычно и предполагают, он характеризуется психрофитом не из-за его многочисленных сходных морфологических и физиологических особенностей. Однако некоторые холодные растения также имеют очень своеобразную форму.
Это гидрофильное вещество. Это водные растения. Согласно их образу жизни и структуре, они содержат погруженные в воду растения и растения с плавающими листьями. Погруженные растения делятся на укореняющиеся в донной почве и подвешенные в толще воды. Из высших растений первым является телорез — Stratiotes aloides, а водным шильником — Subularia aquatika. В эту группу также входят водоросли, которые прилипают к Земле. Среди растений, подвешенных в толще воды, можно упомянуть погруженные в воду роговые водоросли — ceratophyllum demelthum, обыкновенную пузырчатку — Utrikularia vulgaris и многочисленные виды плавающих водорослей.
Растения с плавающими листьями частично используют воду, частично — воздух. Из них в землю пускают корни водяные лилии рода Nymphaea, стручки рода Nuphar, rdesta и водяные орехи Trapa natans.
Из них в землю пускают корни водяные лилии рода Nymphaea, стручки рода Nuphar, rdesta и водяные орехи Trapa natans.
Многие виды, наряду с плавающими листьями на поверхности воды, также встречаются в воде. Ряска и водокрас плавают на поверхности воды, не приживаясь.
Гелофиты или амфибии — группа растений земноводных, которые очень тесно связаны с реальными водными растениями, обычно рассматриваются вместе с ними. Это типы прибрежных и прибрежных местообитаний с избыточным или изменчивым увлажнением. Они могут расти как в воздухе, так и частично в воде, выдерживая полное временное наводнение. Как и в природе, здесь нет резкой границы между водными и сухопутными местообитаниями для растений, поэтому группа герофитов, с одной стороны, имеет реальные гидрофильные, а с другой-примеры наземных Гелофитов-растения прибрежной зоны пресноводных водоемов и рек: Стрелец тонтифолия, ежеголовка-сагоголовка-Sparganium ramosum.
Анатомо-морфологические приспособления растений к водному режиму
Растения, ограниченные ареалом обитания в различных увлажнительных условиях, выработали соответствующую адаптацию к водному режиму.
Характерной структурной особенностью Гигрофитов является тонкая и мягкая листовая пластинка с небольшим количеством пор, не имеющих толстой кутикулы, и рыхлый слой листовой ткани с большим межклеточным пространством.
Для ксерофитов большое значение имеют различные структурные приспособления к состоянию недостатка влаги.
Корневая система обычно сильно развита, что помогает растению увеличить поступление почвенной влаги. По общей массе корневая система ксерофитов часто превышает надземную часть, иногда очень крупную. Например, многие виды трав и кустарников в пустынях Средней Азии имеют подземные массы в 9-10 раз больше, чем наземные массы, в то время как корневая система кеслофитов в холодных пустынях Памирского нагорья часто является типом, то есть растениями с длинными корнями, которые распространяются по большим массам почвы, но относительно далеко расходятся. При более глубоком проникновении таких корней сухие грибы могут использовать влагу в глубоких слоях почвы, а в некоторых случаях и грунтовые воды.
У других видов корневая система имеет интенсивный тип: они покрывают сравнительно небольшое количество почвы, но за счет очень плотного ветвления максимизируют влажность почвы. Корни многих аэробных видов имеют специальные приспособления для хранения влаги.
Наземные органы ксерофитов также отличаются уникальным (так называемым гетеротипическим признаком), который несет на себе отпечаток сложных условий водоснабжения. Они имеют высокоразвитую систему водоснабжения, что хорошо видно по плотности венозной сети листьев, которые снабжают водой ткани. Эта особенность позволяет легко пополнять запасы влаги, которые ксерофиты используют для испарения.
Различные конструктивные устройства защитного характера, направленные на снижение водопотребления, сводятся в основном к следующему:
- Общее уменьшение поверхности транспирации. Многие ксерофиты имеют небольшие, узкие и очень редуцированные листовые пластинки. В особо сухих пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцируются до едва заметных чешуек. У таких видов фотосинтез осуществляется зелеными ветвями.
- Уменьшение листовой поверхности в самые жаркие и засушливые периоды вегетации. Для многих кустарников Средней Азии, Северной Африки и других пустынь, а также для некоторых видов средиземноморских сухих субтропиков характерны сезонные варианты листьев.
- Защита листьев от больших потерь влаги при транспирации. Это достигается за счет развития мощной покровной ткани (толстостенного, иногда многослойного эпидермиса), а в различных выростах и других видах, которые часто образуют толстые «войлочные» удары по поверхности листа, поверхность кутикулы покрывается толстым слоем или восковым гидроизоляционным покрытием.
 У таких видов фотосинтез осуществляется зелеными ветвями.
У таких видов фотосинтез осуществляется зелеными ветвями.Развитие защитного покрова листьев является причиной того, что луговая трава имеет тусклый сероватый оттенок, который значительно отличается от ярко-зеленого луга. Поры ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальном углублении ткани листа, иногда с помощью волос и других дополнительных защитных приспособлений.
Ковили и другие степные травы обладают интересным механизмом защиты устьиц в самое жаркое и сухое время суток:при большой потере воды, во влажную погоду эпидермиса клетки эпидермиса восстанавливают выпуклость, и лопасти листьев снова разворачиваются.
Повышенное развитие механической ткани. Клетки ткани листа ксерофитов имеют небольшие размеры и очень плотную упаковку, то есть малую генерацию межклеточных клеток, что значительно уменьшает внутреннее испарение поверхности листа. Ксерофиты обычно живут в открытых, светлых местообитаниях, поэтому многие особенности ксероморфного строения листьев являются также особенностями светлого строения. Так, у многих видов листья часто имеют мощную, иногда многорядную частоколообразную паренхиму, расположенную с обеих сторон.
Основными морфологическими и эктопическими признаками Мезофитов были средние значения между гигрофитами и ксофитами. Мезофиты умеренно развивают широкий и интенсивный тип корневой системы и имеют все переходы между ними. Листья характеризуются дифференцировкой тканей на рыхлую губчатую паренхиму с системой более или менее плотных палисадных паренхим и просветов. Сеть вен относительно разрежена. Покровная ткань имеет признаки отдельной атипичной формы, но не выражена, как при сухом дерматите.
Листья характеризуются дифференцировкой тканей на рыхлую губчатую паренхиму с системой более или менее плотных палисадных паренхим и просветов. Сеть вен относительно разрежена. Покровная ткань имеет признаки отдельной атипичной формы, но не выражена, как при сухом дерматите.
Ментальная зародышевая клетка имеет сильную гетерогенную структуру листа. Таким образом, кишечнополостные являются узколистными, хорошо развитыми проводящими и механическими тканями. Вечнозеленые кустарники имеют плотные кожистые листья, иногда с очень сильной кутикулой, плотный частокол паренхимы. Нижняя сторона, которая поддерживает поры, часто покрыта слоем полового созревания или воска. Однако некоторые влажные особенности отчетливо прослеживаются в строении листьев, главным образом в развитии больших межклеточных пространств в больших клеточных размерах и губчатых тканях.
Анатомо-морфологические особенности водных растений существенно отличают их от наземных растений. Уменьшение механической ткани заметно. Проводящие системы уменьшаются так же сильно. Если земля мезодермы имеет длину жилок на 1 кв. из см листьев около 100 мм и более, у сухих грибов она достигает 300, А у водных и прибрежных растений в несколько раз меньше. Вот некоторые примеры.
Проводящие системы уменьшаются так же сильно. Если земля мезодермы имеет длину жилок на 1 кв. из см листьев около 100 мм и более, у сухих грибов она достигает 300, А у водных и прибрежных растений в несколько раз меньше. Вот некоторые примеры.
У некоторых погруженных растений, не прикрепленных к земле, корни полностью редуцируются, остальные корни сохраняются, но отдельные плавающие части растения можно обойтись и без них. Однако многие виды имеют толстые и крепкие корневища, которые играют роль якорей, резервов и вегетативных органов.
Листья погруженных в воду водных растений очень тонкие и нежные и имеют упрощенную мезофильную структуру без выраженной дифференциации на частокол и губчатую паренхиму. Листья в воде без устьиц. В некоторых местах имеется группа клеток эпидермиса с тонкими стенками. Считается, что они играют большую роль в усвоении воды и растворенных минеральных солей.
У растений, которые лишь частично погружены в воду, гетерофилл хорошо выражен разница в строении листьев на поверхности одна и та же особь и в воде. Первый имеет общую черту в листьях второго наземного растения — очень тонкую или рассеченную листовую пластинку. Гетерофилия наблюдалась у водяных кимбана, водяных лилий и стручков, стреловидных и других видов. Интересным примером является поручейник, где можно увидеть несколько форм листьев, представляющих все переходы от типичной суши к воде.
Первый имеет общую черту в листьях второго наземного растения — очень тонкую или рассеченную листовую пластинку. Гетерофилия наблюдалась у водяных кимбана, водяных лилий и стручков, стреловидных и других видов. Интересным примером является поручейник, где можно увидеть несколько форм листьев, представляющих все переходы от типичной суши к воде.
Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности
Помимо морфологических особенностей растения, оно ограничено, развито и физиологично в местах с повышенной влажностью в различных условиях.
Способность гигроскопичности регулировать режим воды ограничена:устьица в основном широко открыты, поэтому транспирация мало отличается от физического испарения. Благодаря беспрепятственному поступлению воды и отсутствию защитных приспособлений интенсивность транспирации очень высока:у легких гигрофитов в дневное время листья могут терять в весе в 4-5 раз больше, чем за время полива, а высокое содержание воды в тканях Гигрофитов поддерживается главным образом за счет постоянного поступления влаги из окружающей среды.
Другой характерной физиологической особенностью гигрофитов, из-за ее легкого использования влаги, является низкое осмотическое давление в клеточном соке, что приводит к быстрой потере водоподготовки, низкой водоудерживающей способности. Например, в случае кислицы и майника потеря от 15% до 20% подачи воды уже необратима и приводит к гибели людей. В некоторых случаях растения в очень влажной среде обитания нуждаются в удалении лишней влаги. Обычно это происходит, когда почва теплая,корни активно впитывают воду,а транспирации нет (например, утро или туман с влажностью 100%). Избыток влаги удаляется путем выделения воды через специальные выделительные ячейки, расположенные по краям или на кончиках листьев.
Ксерофиты имеют много различных физиологических приспособлений, которые могут успешно противостоять недостатку влаги.
Ксерофиты обычно повышают осмотическое давление клеточных соков, что позволяет им поглощать воду даже при большом водоотводящем усилии в почве.
 Она измеряется в тысячах кПа, а некоторые пустынные кустарники регистрируют регистрационный номер, который достигает от 10000 до 30000 кПа.
Она измеряется в тысячах кПа, а некоторые пустынные кустарники регистрируют регистрационный номер, который достигает от 10000 до 30000 кПа.В течение длительного времени внимание было привлечено к вопросу потребления воды сухими батареями для испарения. Представляется, что значительное уменьшение транспирации должно способствовать большому количеству анатомических приспособлений, надежно защищающих наземную часть ксерофитов от сильного испарения. Но оказывается, что это не совсем так. При достаточном запасе воды большинство сухих грибов обладают достаточно высокой транспирацией, но с наступлением сухих условий они значительно ее снижают. При этом играет роль и закрытие пор, и начало сильного обезвоживания подводнии листа. Несомненно, анатомо-морфологические приспособления имеют определенное значение, но главная роль в устойчивости к высыханию сухих грибов в настоящее время отводится физиологическим механизмам.
Эти механизмы включают высокую водоудерживающую способность тканей и клеток, обусловленную многими физиологическими и биохимическими особенностями.
Большое значение для выживания ксерофитов при недостатке влаги имеет способность переносить глубокое обезвоживание тканей без потери жизнеспособности, а при возобновлении благоприятных условий растения — ксерофиты могут потерять 75% общего запаса воды, но все же жить.
Другая адаптивная система, обеспечивающая выживание ксерофитов в засушливых условиях, заключается в использовании растениями наиболее благоприятных периодов года для вегетации, а в районах с ярко выраженным средиземноморским климатом с сухим периодом лета за счет жизнедеятельности во время засух многие виды имеют «двухтактный» ритм аэробного сезонного развития: во время весенних дождей; в течение лета происходит регенерация растительности; в течение лета следует зимний отдых. Аналогичное явление наблюдается у сухих луговых растений в середине и конце лета: потеря части листовой поверхности, приостановка развития, сильное обезвоживание тканей и др.
Физиологическая адаптация суккулентов очень своеобразна и требует отдельного рассмотрения.
Основным способом преодоления условий высыхания суккулентов является накопление большого количества воды в ткани и использование ее очень экономно. В жарком и сухом климате весь запас воды может быть быстро израсходован, но растение имеет защитное устройство для уменьшения испарения. Одна из них — своеобразная форма надземной части суккулентов. Кроме того, многие суккуленты имеют поверхность, защищенную восковым покрытием витилиго, но суккуленты с тонким незащищенным эпидермисом также имеют очень мало пор и часто погружаются в ткани листьев или стеблей. В течение дня устьица обычно закрыты, и потеря воды проходит главным образом через покровную ткань.
Транспирация суккулентов очень низкая. Его трудно поймать за короткий промежуток времени и необходимо определить расход воды. Водоудерживающая способность мясистых тканей была значительно выше, чем у других растений экологической группы, за счет содержания в клетках гидрофильных веществ. Поэтому, даже не имея доступа к влаге, суккуленты потребляют воду очень медленно и продолжают жить в течение длительного времени даже в растительных образцах.
Ограничения, обусловленные особенностями водного режима суккулентов, вызывают и другие трудности для жизни этих растений в сухих условиях. Слабая транспирация сводит к минимуму возможность терморегуляции, которая связана с сильным нагреванием огромных наземных органов суккулентов. Поскольку устьица обычно закрыты днем и открыты ночью, близость углекислого газа и света не совпадает во времени, возникают также трудности для фотосинтеза. Поэтому суккуленты разработали особый метод фотосинтеза, при котором продукты дыхания частично используются в качестве источника углекислого газа. Иными словами, в экстремальных ситуациях растения частично используют принцип замкнутой системы с повторным использованием метаболических отходов. Из-за всех этих ограничений интенсивность фотосинтеза у суккулентов низкая, их рост и накопление массы очень медленные,поэтому они не обладают высокой биологической продуктивностью и могут быть использованы для закрытых растений.
Физиологические показатели подтверждают их промежуточное положение: они характеризуются умеренным значением осмотического давления, количеством воды в листьях и маргинальным дефицитом воды. Что касается транспирации, то ее величина в большей степени зависит от условий освещения и других элементов микроклимата.
Что касается транспирации, то ее величина в большей степени зависит от условий освещения и других элементов микроклимата.
Одни и те же среднетемпературные виды, попадая в различные условия водоснабжения, проявляют определенную пластичность, приобретают более глубокие свойства в сухих условиях, обладают более влажными свойствами во влажных условиях.
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одних и тех же особей. Например, у дерева на опушке леса листья на стороне, обращенной к лесу, имеют более умеренный и затененный характер, по сравнению со слегка деформированными листьями на внешней стороне дерева. Разные высотные слои листьев одного и того же растения находятся в разных условиях полива, так как приток воды к вершине связан с преодолением большого сопротивления. Кроме того, верхние листья дерева обычно находятся в другом микроклимате.
Различие анатомо-физиологических показателей листьев на разных сторонах кроны дерева, растущего на опушке леса.
Поскольку водная среда существенно отличается от воздушной, водные растения обладают многими уникальными физиологическими адаптивными функциями. Интенсивность света в воде сильно ослаблена, так как часть падающего излучения отражается от поверхности воды, а другие поглощаются ее толщей. Из-за ослабления света фотосинтез у растений в воде значительно уменьшается с увеличением глубины. Выживание глубоководного фитопланктона в зоне, где освещенность ниже точки коррекции, обеспечивается периодическим вертикальным перемещением в Верхнюю зону, где происходит интенсивный фотосинтез и пополнение органического вещества.
В воде, помимо недостатка света, растения могут испытывать еще одну трудность, необходимую для фотосинтеза-недостаток доступного CO2. Углекислый газ поступает в воду в результате растворения СО2, содержащегося в воздухе,дыхания водных организмов, разложения органических остатков и выделения из них карбонатов. Вот почему при интенсивном фотосинтезе растений потребление CO2 увеличивается, и его легко не хватает.
Гидрофиты реагируют на увеличение содержания СО2 в воде значительным увеличением фотосинтеза.
В погруженном растении нет испарения, поэтому нет «верхнего двигателя», который поддерживает поток воды растения. Однако этот поток, снабжающий ткани питательными веществами, присутствует с ежедневной частотой. Активная роль в его поддержании принадлежит корневому давлению и активности особых клеток, выделяющих воду-водяных устьиц.
Заключение
Плавающие или выступающие листья обычно имеют сильную транспирацию, но они непосредственно соприкасаются с водой и располагаются в воздушном слое с высокой влажностью. Поры широко открыты и полностью закрываются только ночью.
Транспирация одинаково высока у прибрежных растений, у которых значительное количество воды потребляется не только листьями, но и стеблями.
Осмотическое давление в водных и прибрежных растениях очень низкое, так как нет необходимости преодолевать водоудерживающую способность почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959).
7) Приспособления животных и растений к недостатку воды
1.Суккуленты —
очень стойкие к перегреву и устойчивые
к обезвоживанию, во время засухи они не
испытывают недостатка воды, потому что
содержат большое количество ее и медленно
расходуют. Корневая система у них
разветвлена во все стороны в верхних
слоях почвы, благодаря чему в дождливые
периоды растения быстро всасывают воду.
Это кактусы, алоэ, молодило.
2.Эвксерофиты —
жаростойкие растения, которые хорошо
переносят засуху. К этой группе относятся
степные растения:полынь , верблюжья
колючка. У них корневая система очень
разветвлена, и основная ее масса размещена
в верхнем слое почвы (50-60 см). Они способны
сбрасывать листья и даже целые ветви.
3.
Стипаксерофиты —
это ковыль, тырса и другие узколистные
степные злаки. Они устойчивы к перегреву,
хорошо используют влагу кратковременных
дождей. Выдерживают лишь кратковременную
Выдерживают лишь кратковременную
нехватку воды в почве.
4.
Пойкилоксерофиты —
растения, не регулирующие своего водного
режима. Это в основном лишайники, которые
могут высыхать до воздушно-сухого
состояния и снова проявлять жизнедеятельность
после дождей.
У
животных также выработался ряд
приспособлений к недостатку влаги.
Мелкие животные (грызуны, пресмыкающиеся,
членистоногие) довольствуются водой,
поступающей вместе с пищей.
Хранением
воды для ряда животных засушливых
районов служат отложения жира (горб у
верблюда, курдюк у овец), при окислении
которого образуется необходимое
количество воды. Ряд животных пустынных
районов обладают способностью к
длительному быстрому бегу (антилопы,
куланы, сайгаки), позволяющему им
совершать дальние миграции на водопой.
Некоторые виды (преимущественно грызуны)
перешли к ночному образу жизни, тем
самым избегая перегрева и большого,
испарения воды
8)
Биологические ритмы или биоритмы
– это более или менее регулярные
изменения характера и интенсивности
биологических процессов. Способность
Способность
к таким изменениям жизнедеятельности
передается по наследству и обнаружена
практически у всех живых организмов.
Их можно наблюдать в отдельных клетках,
тканях и органах, в целых организмах и
в популяциях.
Выделим
следующие важные достижения биоритмологии:
1.
Биологические ритмы обнаружены на всех
уровнях организации живой природы –
от одноклеточных до биосферы. Это
свидетельствует о том, что биоритмика
– одно из наиболее общих свойств живых
систем.
2.
Биологические ритмы признаны важнейшим
механизмом регуляции функций организма,
обеспечивающим гомеостаз, динамическое
равновесие и процессы адаптации в
биологических системах.
3.
Установлено, что биологические ритмы,
с одной стороны, имеют эндогенную природу
и генетическую регуляцию, с другой, их
осуществление тесно связано с
модифицирующим фактором внешней среды,
так называемых датчиков времени. Эта
связь в основе единства организма со
средой во многом определяет экологические
закономерности.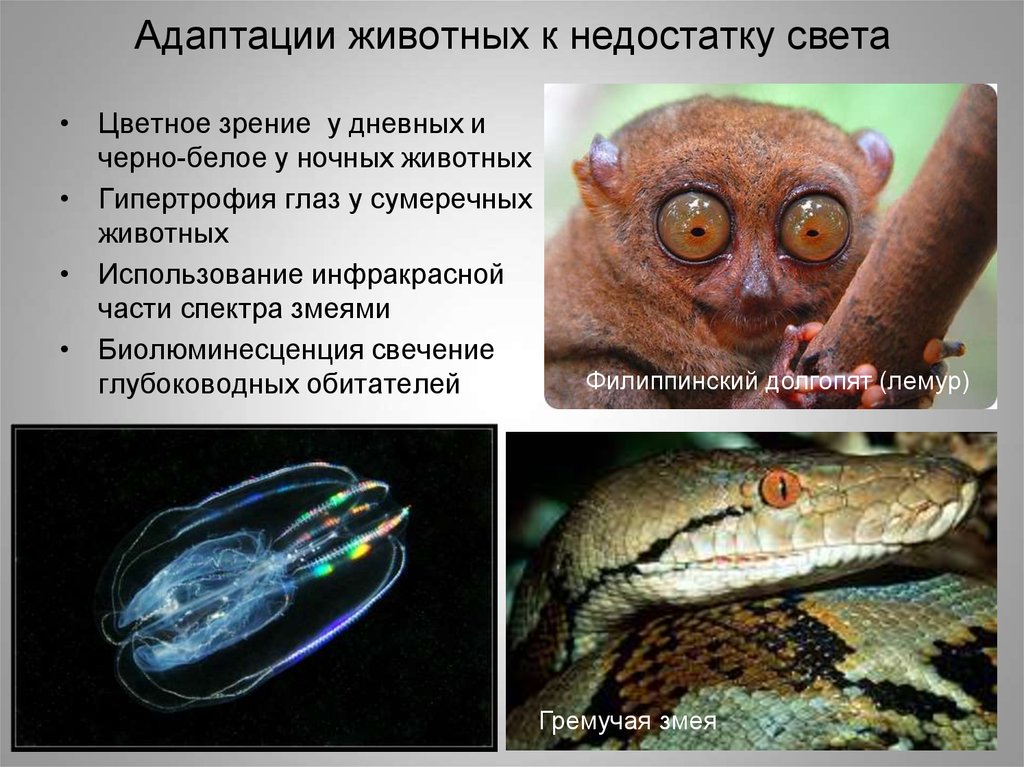
4.
Сформулированы положения о временной
организации живых систем, в том числе
– человека – одним из основных принципов
биологической организации. Развитие
этих положений очень важно для анализа
патологических состояний живых систем.
5.
Обнаружены биологические ритмы
чувствительности организмов к действию
факторов химической (среди них
лекарственные средства) и физической
природы. Это стало основой для развития
хронофармакологии, т.е. способов
применения лекарств с учетом зависимости
их действия от фаз биологических ритмов
функционирования организма и от состояния
его временной организации, изменяющейся
при развитии болезни.
6.
Закономерности биологических ритмов
учитывают при профилактике, диагностике
и лечении заболеваний.
Биоритмы
подразделяются на физиологические и
экологические. Физиологическиеритмы,
как правило, имеют периоды от долей
секунды до нескольких минут. Это,
например, ритмы давления, биения сердца
и артериального давления. Имеются данные
о влиянии, например, магнитного поля
Земли на период и амплитуду энцефалограммы
человека.
Экологические ритмы
по длительности совпадают с каким-либо
естественным ритмом окружающей среды.
К ним относятся суточные, сезонные
(годовые), приливные и лунные ритмы.
Благодаря экологическим ритмам, организм
ориентируется во времени и заранее
готовится к ожидаемым условиям
существования. Так, некоторые цветки
раскрываются незадолго до рассвета,
как будто зная, что скоро взойдет солнце.
Многие животные еще до наступления
холодов впадают в зимнюю спячку или
мигрируют. Таким образом, экологические
ритмы служат организму как биологические
часы.
Фотопериодическая
реакция —
ответная реакция живых организмов на
относительную продолжительность дня
и ночи, наблюдаемая в годовом цикле.
И
в мире растений, и в мире животных есть
великое множество событий, происходящих
всего один раз в году. В зонах умеренного
климата такие события так или иначе
связаны со сменой времен года, но
аналогичные явления наблюдаются и в
тропиках, расположенных недалеко от
экватора, где эта смена не столь заметна.
Ни для кого, хоть сколько-нибудь
внимательно наблюдающего за жизнью
окружающей его природы, не секрет, что
разные растения цветут в разное время.
К концу зимы, как только появляются
первые проталинки, расцветает подснежник.
Позднее, когда основательно потеплеет,
луга покрываются желтыми цветками
лютичного чистяка, весеннего адониса.
Среди лета цветков самой различной
окраски столько, что и не сосчитать,
зато в конце осени прощальный привет
теплому времени года шлют немногие
растения: безвременник, борец, астры.
К
чередованиям тепла и холода на протяжении
года приспособились не только растения,
но и животные. С наступлением весны
вылезают из своих нор и берлог те из
них, кто провел зиму в спячке, возвращаются
перелетные птицы. В жизни многих животных
с весенними месяцами совпадает брачный
период, а лето уходит на то, чтобы
вырастить потомство.
Наступление
осени также знаменует большие перемены:
перелетные птицы улетают, животные,
впадающие в зимнюю спячку, нагуливают
жир перед тем, как залечь в берлоги и
норы, грызуны делают запасы на зиму. У
У
насекомых замедляются все жизненные
процессы, они скрываются в укромных
местах, чтобы там на стадии покоя
перезимовать и дождаться прихода весны.
Долгое
время значение в регуляции процессов
с годичным ритмом придавали температуре,
влажности и скорости созревания растений.
Много гипотез было выдвинуто и опровергнуто
прежде, чем удалось доказать, что решающим
фактором в наступлении той или иной
стадии в репродуктивном цикле у растений
служит относительная продолжительность
дня и ночи.
Лицом к солнцу: адаптация растений
10 марта 2017 г. Фото: Дебора Смолл Для всех живых существ адаптация имеет решающее значение для обеспечения выживания в заданных условиях. Чтобы выжить, организм должен собирать достаточно пищи или энергии для роста, защищать себя от вреда и размножаться. Чтобы получить или сохранить достаточно воды для роста, в сухом климате, таком как Сан-Диего, растения выработали различные приспособления. Фото: Дебора Смолл Еще один способ борьбы с засухой – запасать воду или ограничивать испарение в засушливые периоды. Фото: Дебора Смолл У каждого из наших местных растений, найденных в Национальном памятнике Кабрильо и по всему Сан-Диего, есть своя история адаптации. Адаптация растений, растительность, природные ресурсы |
 Некоторые из них имеют длинную или сложную корневую систему, которая подключается к грунтовым водам, хранящимся глубоко под землей. Другие хранят воду в тканях листьев для использования в условиях засухи. Различные приспособления листьев, такие как окраска, форма и жесткость листьев, помогают ограничить транспирацию в течение дня. Эти же приспособления помогают собирать и направлять воду к растению. В средиземноморском климате Сан-Диего, характеризующемся коротким сезоном дождей, сменяющимся продолжительным засушливым периодом, местные растительные сообщества прибрежных кустарников шалфея и южного морского чапараля адаптировались разными способами, чтобы обеспечить выживание в эти ежегодные засушливые периоды. Кроме того, каждое растение, найденное в Национальном памятнике Кабрильо, адаптировалось одним или несколькими способами, чтобы обеспечить его выживание в течение всего года.
Некоторые из них имеют длинную или сложную корневую систему, которая подключается к грунтовым водам, хранящимся глубоко под землей. Другие хранят воду в тканях листьев для использования в условиях засухи. Различные приспособления листьев, такие как окраска, форма и жесткость листьев, помогают ограничить транспирацию в течение дня. Эти же приспособления помогают собирать и направлять воду к растению. В средиземноморском климате Сан-Диего, характеризующемся коротким сезоном дождей, сменяющимся продолжительным засушливым периодом, местные растительные сообщества прибрежных кустарников шалфея и южного морского чапараля адаптировались разными способами, чтобы обеспечить выживание в эти ежегодные засушливые периоды. Кроме того, каждое растение, найденное в Национальном памятнике Кабрильо, адаптировалось одним или несколькими способами, чтобы обеспечить его выживание в течение всего года.  В среднем в этом районе выпадает менее десяти дюймов дождя в год, а сухой период обычно бывает с апреля по октябрь. Поскольку вода является жизненно важным ресурсом для жизни, как же выживают эти местные растения? Большинство местных растений, произрастающих в парке, относятся к засухо-лиственным. Эти растения сбрасывают листья в сухой сезон и впадают в состояние покоя до следующего дождя. Например, Ecelia californica, Калифорнийский подсолнух, быстро цветущий в сезон дождей, раскрывающий свои красивые желтые цветы. Эти цветы дают семена для будущих поколений. В засушливый сезон Ecelia californica сбрасывает листья и для неподготовленного глаза выглядит мертвой. Тем не менее, эта зависимость от воды сохранится на долгие годы вперед. В следующий раз, когда вы будете в парке, осмотритесь, чтобы увидеть, какие растения могут находиться в спящем или цветущем состоянии.
В среднем в этом районе выпадает менее десяти дюймов дождя в год, а сухой период обычно бывает с апреля по октябрь. Поскольку вода является жизненно важным ресурсом для жизни, как же выживают эти местные растения? Большинство местных растений, произрастающих в парке, относятся к засухо-лиственным. Эти растения сбрасывают листья в сухой сезон и впадают в состояние покоя до следующего дождя. Например, Ecelia californica, Калифорнийский подсолнух, быстро цветущий в сезон дождей, раскрывающий свои красивые желтые цветы. Эти цветы дают семена для будущих поколений. В засушливый сезон Ecelia californica сбрасывает листья и для неподготовленного глаза выглядит мертвой. Тем не менее, эта зависимость от воды сохранится на долгие годы вперед. В следующий раз, когда вы будете в парке, осмотритесь, чтобы увидеть, какие растения могут находиться в спящем или цветущем состоянии.  У большинства суккулентов или кактусов есть утолщенные и мясистые части, которые используются для удержания воды, например, у прибрежной опунции, дамских пальчиков или агавы Шоу. Эти растения обычно легко идентифицировать. Тем не менее, присмотритесь к Rhus integrifolia (Lemonade Berry) или Dudleya pulverulenta ( Chalk Dudleya). Оба растения приспособились удерживать воду в своих листьях. Листья Lemonade Berry жесткие и восковые, что ограничивает транспирацию растения в атмосферу. Тогда как Мел Дудлея своими широкими мясистыми зелеными листьями выделяет белое «известковое» вещество, которое использовалось для отражения солнечного света. Точно так же, как вы наносите солнцезащитный крем перед тем, как долго стоять перед солнцем, меловая дудлея адаптировалась аналогичным образом.
У большинства суккулентов или кактусов есть утолщенные и мясистые части, которые используются для удержания воды, например, у прибрежной опунции, дамских пальчиков или агавы Шоу. Эти растения обычно легко идентифицировать. Тем не менее, присмотритесь к Rhus integrifolia (Lemonade Berry) или Dudleya pulverulenta ( Chalk Dudleya). Оба растения приспособились удерживать воду в своих листьях. Листья Lemonade Berry жесткие и восковые, что ограничивает транспирацию растения в атмосферу. Тогда как Мел Дудлея своими широкими мясистыми зелеными листьями выделяет белое «известковое» вещество, которое использовалось для отражения солнечного света. Точно так же, как вы наносите солнцезащитный крем перед тем, как долго стоять перед солнцем, меловая дудлея адаптировалась аналогичным образом.  Исследуя парк, постарайтесь узнать больше таких историй. Как растение переживает хищничество? Как он способен размножаться? Как растение смотрит на солнце? Всегда помните, что первый шаг любого ученого — это наблюдение. Так что выходите и начинайте наблюдать!
Исследуя парк, постарайтесь узнать больше таких историй. Как растение переживает хищничество? Как он способен размножаться? Как растение смотрит на солнце? Всегда помните, что первый шаг любого ученого — это наблюдение. Так что выходите и начинайте наблюдать!Последнее обновление: 10 марта 2017 г.
Адаптации | manoa.hawaii.edu/ExploringOurFluidEarth
Световые зоны
Солнечный свет является основным источником энергии для фотосинтеза. Водоросли и сосудистые растения, живущие в воде, должны жить достаточно близко к поверхности, чтобы получать солнечный свет. Эвфотическая зона — это слой воды, в котором достаточно света для чистого фотосинтеза. Другими словами, скорость фотосинтеза выше скорости дыхания (рис. 2.42). В эвфотической зоне происходит общее чистое производство кислорода.
В чистых тропических водах эвфотическая зона может достигать глубины 80 метров (м). Ближе к полюсам энергия солнечного света не проникает так глубоко. Таким образом, эвфотическая зона может иметь глубину менее 10 м (рис. 2.42). В мутной воде взвешенный материал блокирует свет, так что проникает еще меньше света. В очень мутных водах эвфотическая зона может составлять всего несколько сантиметров.
Ближе к полюсам энергия солнечного света не проникает так глубоко. Таким образом, эвфотическая зона может иметь глубину менее 10 м (рис. 2.42). В мутной воде взвешенный материал блокирует свет, так что проникает еще меньше света. В очень мутных водах эвфотическая зона может составлять всего несколько сантиметров.
Дисфотическая зона — слой ниже эвфотической зоны. Этот слой также известен как сумеречная зона. В этой зоне достаточно света, чтобы видеть зрительных хищников, и даже для некоторого фотосинтеза. Однако скорость дыхания в дисфотической зоне больше скорости фотосинтеза. В чистой воде дисфотическая зона может простираться до 800 м. Эвфотическая и дисфотическая зоны вместе составляют фотическая зона .
Большая часть биомассы океана обитает в фотической зоне. Однако большая часть воды океана находится под фотической зоной — в области, называемой афотической зоной. Афотическая зона начинается там, где достигает менее одного процента солнечного света. Поэтому афотическая зона очень темная. Фактически, света, производимого биолюминесцентными организмами, больше, чем солнечного света. Биолюминесценция — производство и излучение света живым организмом. Биолюминесцентный свет производится в результате химических реакций.
Поэтому афотическая зона очень темная. Фактически, света, производимого биолюминесцентными организмами, больше, чем солнечного света. Биолюминесценция — производство и излучение света живым организмом. Биолюминесцентный свет производится в результате химических реакций.
Афотические зоны не ограничиваются водами океана. Некоторые озера также достаточно глубоки, чтобы иметь афотическую зону. Озеро Байкал в России, например, имеет глубину более 1600 м и имеет большую афотическую зону.
| фото — свет. Например, копировальный аппарат использует свет для копирования. |
| ЕС – хорошо. Например, человек, находящийся в эйфории, чувствует себя исключительно хорошо. |
дис – удаление. Например, разъединение означает удаление соединения. Например, разъединение означает удаление соединения. |
| и – без. Например, тот, кто аморален, лишен морали. |
Количество энергии, проникающей через поверхность воды, зависит от угла, под которым солнечный свет падает на океан. Когда энергия солнечного света попадает в океан, часть ее отражается от поверхности и возвращается в атмосферу. Поскольку Земля представляет собой шар, только области вблизи экватора в полдень почти перпендикулярны пути падающего света. В других местах свет падает под углом (рис. 2.43). Таким образом, на экваторе к поверхности вод проникает большее количество энергии, чем в полярных широтах.
Помимо отражения, энергия солнца теряется на поглощение (рис. 2.44). Фактически, вода поглощает почти все относительно низкоэнергетические инфракрасные волны, исходящие от солнечного света в пределах 0,1 м (10 сантиметров) от поверхности. В этом очень мелком слое энергия преобразуется в тепло, которое либо повышает температуру воды, либо вызывает ее испарение.