
Содержание
Признаки избытка и недостатка света для растений. Светолюбивые, теневыносливые, тене индифферентные растения
Признаки избытка и недостатка света
Если рассматривать растения с точки зрения их «отношения» к свету, то их принято делить на три категории:
— светолюбивые растения
— теневыносливые растения
— тене индифферентные растения.
Комнатные растения, как правило, являются светолюбивыми, поэтому наиболее оптимально они развиваются, если в помещении имеет место полное освещение. Кроме того, растения различаются еще и разной теневыносливостью.
Все растения в определенной степени могут адаптироваться к условиям, изменяющимся на протяжении их жизни. Так, отдельные виды растений отлично приспосабливаются к большому количеству света либо к его недостатку. Однако есть много видов растений, для которых очень важно обеспечить именно четко определенные параметры освещения.
Адаптируясь к слишком низкому количеству света, растение постепенно меняет свой облик. Его листья приобретают темно-зеленый оттенок, их размер становится больше. Междоузлия стебля вытягиваются и становятся менее прочными, а некоторые растения без достаточного количества света вообще перестают цвести. Все эти явления – следствие снижения производства продуктов фотосинтеза, которые необходимы для построения тела растения.
Его листья приобретают темно-зеленый оттенок, их размер становится больше. Междоузлия стебля вытягиваются и становятся менее прочными, а некоторые растения без достаточного количества света вообще перестают цвести. Все эти явления – следствие снижения производства продуктов фотосинтеза, которые необходимы для построения тела растения.
В то же время избыток света может послужить причиной частичного разрушения хлорофилла. В результате листья приобретают желто-зеленый оттенок. Если света слишком много, растения развиваются медленнее, и в итоге их отличают короткие междоузлия, а также короткие и широкие листья. В подобных обстоятельствах важно вовремя принять необходимые меры и обеспечить правильное, подходящее освещение для растений.
Растения, которые воспринимают свет нейтрально, будут цвести в том случае, если они длительное время росли под достаточным освещением. Для таких растений важно, чтобы помещение было освещено, по крайней мере, около восьми часов, в идеале же яркий свет должен присутствовать от двенадцати до шестнадцати часов. Каждый вид растений имеет собственные особенности и, соответственно, собственное «отношение» к свету.
Каждый вид растений имеет собственные особенности и, соответственно, собственное «отношение» к свету.
Если вы выращиваете светолюбивые растения, то учтите, что длина светового дня для них должна составлять от 13 до 15 часов. Только получая достаточное количество света, такое растение образует завязь и позже зацветет. При этом освещение может быть как природным, так и качественно обустроенным искусственным. Важно учесть, что и избытка освещения допускать в данном случае нельзя. К растениям светолюбивым относятся бальзамин, сенполия, пеларгония, кальцеолярия, эпифиллюм, глоксиния, примула, колеус, цинерария, колокольчик, равнолистный, стефанотис.
Для растений тенелюбивых в полнее достаточно обеспечить световой день длительностью от 12 до 14 часов света. Если соблюдать такой подход в течение 8-10 недель, то на растениях сначала появится завязь, а потом они зацветут. Среди тенелюбивых растений наиболее часто любители комнатных цветов выращивают каланхое, традесканцию, бегонии, пуансеттию, азалии, зигокактус.
Для того чтобы растения росли и развивались в нормальном режиме, важно обеспечить им дополнительные источники света в зимний период. Растения, которые растут в полутени, требуют дополнительного освещения исключительно тогда, если они располагаются слишком далеко от окон и, следовательно, от природного освещения. В данном случае рекомендуется освещенность от 1000 до 3000 лк.
Что такое люмены и люксы?
Растениям, которые оптимально чувствуют себя при рассеянном свете, подойдет освещение в пределах 3000 — 4000 лк.
Те растения, которым по нраву прямые лучи солнца, требуют, чтобы им было обеспечено освещение на уровне 4000 — 6000 лк.
|
А вот при выращивании экзотических растений освещение необходимо особенно мощное: 6000 – 12000 лк. Особенно важен свет для тех растений, которые дают плоды. Однако даже самые неприхотливые растения могут со временем погибнуть, если продолжительность светового дня будет постоянно нарушаться.
|
 Без света, единственного источника энергии для растения, растение не сможет развиваться и функционировать правильно. Поэтому если вы используете искусственное освещение, то желательно использовать электронный таймер, который будет включать и выключать свет для растений в одно и то же время. При недостаточном освещении изначально будет меняться окраска листьев, теряться яркость их рисунка. Постепенно отпадают нижние листья растения, цветы у таких экземпляров меньшего размера. Как следствие всех этих явлений, рост растения прекращается полностью, и наступает его гибель. Необходимо отметить, что более восприимчивыми к недостаточному освещению всегда будут молодые растения, ведь более зрелые экземпляры имеют развитую систему корней, в которой хранятся определенные запасы питательных веществ. Поэтому такое растение может выдержать несколько месяцев не подходящих для него условий.
Без света, единственного источника энергии для растения, растение не сможет развиваться и функционировать правильно. Поэтому если вы используете искусственное освещение, то желательно использовать электронный таймер, который будет включать и выключать свет для растений в одно и то же время. При недостаточном освещении изначально будет меняться окраска листьев, теряться яркость их рисунка. Постепенно отпадают нижние листья растения, цветы у таких экземпляров меньшего размера. Как следствие всех этих явлений, рост растения прекращается полностью, и наступает его гибель. Необходимо отметить, что более восприимчивыми к недостаточному освещению всегда будут молодые растения, ведь более зрелые экземпляры имеют развитую систему корней, в которой хранятся определенные запасы питательных веществ. Поэтому такое растение может выдержать несколько месяцев не подходящих для него условий.При недостаточном освещении изначально будет меняться окраска листьев, теряться яркость их рисунка. Постепенно отпадают нижние листья растения, цветы у таких экземпляров меньшего размера. Как следствие всех этих явлений, рост растения прекращается полностью, и наступает его гибель. Необходимо отметить, что более восприимчивыми к недостаточному освещению всегда будут молодые растения, ведь более зрелые экземпляры имеют развитую систему корней, в которой хранятся определенные запасы питательных веществ. Поэтому такое растение может выдержать несколько месяцев не подходящих для него условий.
Постепенно отпадают нижние листья растения, цветы у таких экземпляров меньшего размера. Как следствие всех этих явлений, рост растения прекращается полностью, и наступает его гибель. Необходимо отметить, что более восприимчивыми к недостаточному освещению всегда будут молодые растения, ведь более зрелые экземпляры имеют развитую систему корней, в которой хранятся определенные запасы питательных веществ. Поэтому такое растение может выдержать несколько месяцев не подходящих для него условий.
В то же время нарушением оптимального светового режима для растений тенелюбивых будет слишком большое количество света. Так, если на листья такого растения слишком длительный период времени попадают лучи солнца, то в итоге на листьях может проявиться световой ожог, а в некоторых случаях растение гибнет.
Существуют и виды растений, которые оптимально развиваются исключительно при соблюдении периодичности светового дня. Следовательно, в качестве нарушений светового режима возможно не только избыточное либо недостаточное освещение.
Так, в широтах нашей страны световой период составляет от 12 до 16 часов в сутки. Например, для тех растений, родиной которых являются тропики, наиболее комфортным для развития будет двенадцатичасовой период светового дня.
При хроническом дефиците света у растений проявляются разные дефекты в процессе роста. Прежде всего, у растения при тотальном дефиците света появляются новые молодые побеги, листья на молодых побегах растут бледноватые, они постепенно уменьшаются в размерах, а междоузлия растений вытягиваются.
К примеру, монстера деликатесная в подобной ситуации образует на этапе раннего развития листья, не разрезанные до конца. Через определенное время у растения уже вырастают крупные листья и в итоге ее декоративный вид ухудшается.
У колеуса может пострадать степень насыщенности окраски листьев, если качество освещения будет неудовлетворительным. При плохом, недостаточном освещении стебель растения оголяется снизу, яркость окраски становится намного хуже. Последнее характерно также для эписции, драцены, кордилины, каладиума. В данном случае оптимальным вариантом будет свет рассеянного характера, умеренно яркий.
Последнее характерно также для эписции, драцены, кордилины, каладиума. В данном случае оптимальным вариантом будет свет рассеянного характера, умеренно яркий.
|
Чтобы защитить растения от преждевременной гибели и обеспечить им комфортный режим, важно позаботиться о дополнительном освещении. Дополнительное освещение необходимо устанавливать, придерживаясь ряда правил. Большая часть растений достаточно комфортно себя чувствуют и, соответственно, хорошо развиваются при освещении от 500 до 2000лк. Поэтому при выборе лампы нужно учесть эти требования. При грамотном подходе даже в тех местах, где освещение недостаточное, можно создавать настоящие живые уголки с помощью качественного искусственного освещения. При искусственном освещении очень важен такой показатель, как люмен. Люмен определяет, сколько световой энергии выходит из проектора. Эта цифра расскажет вам про потенциал проектора, однако, о яркости изображения нечего известно не будет.
|
 Число люмен крайне трудно измерить. Большее число способов дают возможность измерения освещенности экрана (в люксах). Достаточный световой поток можно рассчитать по определенной формуле:
Число люмен крайне трудно измерить. Большее число способов дают возможность измерения освещенности экрана (в люксах). Достаточный световой поток можно рассчитать по определенной формуле:Создавая дополнительную подсветку для растений, следует обязательно обратить внимание на несколько важнейших факторов.
|
Прежде всего, важно знать, что для растений свойственен фототропизм. Этот термин обозначает реакцию растения на то, в каком направлении падает свет. Искусственное освещение, как и естественное, должно обязательно падать сверху. Если данная условность соблюдена, то растения не будут тратить дополнительную энергию на то, чтобы изменить положение листьев по отношению к источнику света. В отличие от растений, свет на которые падает сбоку, в случае источника света сверху растения будут менее искривленными. Для взрослых, хорошо развитых растений световой день должен составлять не больше двенадцати часов в сутки.
|
 Если световой день будет более длинным, то цветение растений может нарушиться, соответственно, полноценно цвести и плодоносить растение уже не будет. Если вы выращиваете сеянцы, то освещение в данном случае должно быть одинаково интенсивным целые сутки. Подобный режим важен до тех пор, пока растение не начнет прорастать и немного не вытянется. Далее постепенно световой день сокращается: сначала свет остается на 16 часов в сутки, позже — до 14 часов.
Если световой день будет более длинным, то цветение растений может нарушиться, соответственно, полноценно цвести и плодоносить растение уже не будет. Если вы выращиваете сеянцы, то освещение в данном случае должно быть одинаково интенсивным целые сутки. Подобный режим важен до тех пор, пока растение не начнет прорастать и немного не вытянется. Далее постепенно световой день сокращается: сначала свет остается на 16 часов в сутки, позже — до 14 часов.Если вы выращиваете сеянцы, то освещение в данном случае должно быть одинаково интенсивным целые сутки. Подобный режим важен до тех пор, пока растение не начнет прорастать и немного не вытянется. Далее постепенно световой день сокращается: сначала свет остается на 16 часов в сутки, позже — до 14 часов.
Для правильного выбора освещения зимой важно учесть температурный режим в помещении. Так, растения из тропиков, которые являются теплолюбивыми, зимой требуют лишь небольшого понижения температуры и незначительного уменьшения интенсивности света. Все остальные растения зимой требуют уменьшения интенсивности освещения только при условии прохладных температур (5-15 градусов С). А цветы, полностью теряющие листья, могут зимовать в темноте и при температуре 0-5 градусов.
Все остальные растения зимой требуют уменьшения интенсивности освещения только при условии прохладных температур (5-15 градусов С). А цветы, полностью теряющие листья, могут зимовать в темноте и при температуре 0-5 градусов.
Признаки избытка света
Растение ослаблено: бледные листья, почернение листьев.
Листья покрываются желтыми пятнами, либо точечными, либо обширными
Макушка становится уплотненной, жесткой, появление ожогов на листьях, замедленный рост растения
Листья становятся хрупкими, закручиваются, срок жизни растения сократится
Черешки листьев становятся короткими, изгибаются
Растение отклоняется от источника света
Признаки недостатка света
Ряды листьев будут расти широко, с большими промежутками, почва будет видна
Черенки листьев непропорционально длинные, изгибаются, поворачиваются, чтобы быть ближе к свету.
Листья будут разворачиваться веером, стебель оголяется, уменьшается яркость
Наклон растения к источнику света
Растения будут стараться тянуться вверх, будет отсутствовать цветение
Растение на вид слабое, бледное.
Быстрое отмирание нижних листьев
Урок № 5.1 Тема. Свет и адаптация к нему живых организмов
Методическое пособие уроков с видеоматериалами
Тип урока — комбинированный
Методы: частично-поисковый, проблемного изложения, репродуктивный, объяснительно-иллюстративный.
Цель:
— осознание учащимися значимости всех обсуждаемых вопросов, умение строить свои отношения с природой и обществом на основе уважения к жизни, ко всему живому как уникальной и бесценной части биосферы;
Задачи:
Образовательные: показать множественность факторов, действующих на организмы в природе, относительность понятия «вредные и полезные факторы», многообразие жизни на планете Земля и варианты адаптаций живых существ ко всему спектру условий среды обитания.
Развивающие: развивать коммуникативные навыки, умения самостоятельно добывать знания и стимулировать свою познавательную активность; умения анализировать информацию, выделять главное в изучаемом материале.
Воспитательные:
Воспитывать культуру поведения в природе, качества толерантной личности, прививать интерес и любовь к живой природе, формировать устойчивое положительное отношение к каждому живому организму на Земле, формировать умение видеть прекрасное.
УУД
Личностные: познавательный интерес к экологии.. Понимание необходимости получения знаний о многообразии биотических связей в природных сообществах для сохранения естественных биоценозов. Способность выбирать целевые и смысловые установки в своих действиях и поступках по отношению к живой природе. Потребность в справедливом оценивании своей работы и работы одноклассников
Познавательные: умение работать с различными источниками информации, преобразовывать её из одной формы в другую, сравнивать и анализировать информацию, делать выводы, готовить сообщения и презентации.
Регулятивные: умение организовать самостоятельно выполнение заданий, оценивать правильность выполнения работы, рефлексию своей деятельности.
Коммуникативные: участвовать в диалоге на уроке; отвечать на вопросы учителя, товарищей по классу, выступать перед аудиторией, используя мультимедийное оборудование или другие средства демонстрации
Планируемые результаты
Предметные: знать — понятия «среда обитания», «экология», «экологические факторы» их влияние на живые организмы, «связи живого и неживого»;. Уметь — определять понятие «биотические факторы»; характеризовать биотические факторы, приводить примеры.
Личностные: высказывать суждения, осуществлять поиск и отбор информации; анализировать связи, сопоставлять, находить ответ на проблемный вопрос
Метапредметные: связи с такими учебными дисциплинами как биология, химия, физика, география. Планировать действия с поставленной целью; находить необходимую информацию в учебнике и справочной литературе; осуществлять анализ объектов природы; делать выводы; сформулировать собственное мнение.
Форма организации учебной деятельности – индивидуальная, групповая
Методы обучения: наглядно-иллюстративный, объяснительно-иллюстративный, частично-поисковый, самостоятельная работа с дополнительной литературой и учебником, с ЦОР.
Приемы: анализ, синтез, умозаключение, перевод информации с одного вида в другой, обобщение.
Изучение нового материала
Свет и адаптация к нему живых организмов
Среди основных абиотических факторов рассмотрим свет, температуру и влажность.
Свет.
В свое время французский астроном Камиль Фламмарион (1842-1925) написал: «Мы об этом не думаем, но все, что ходит, двигается, живет на нашей планете, есть дитя Солнца».
Действительно, только на свету осуществляется важнейший в биосфере процесс фотосинтеза, который в общем виде может быть представлен следующим образом:
У зеленых растений (высших растений и водорослей) донором
электронов является вода (кислород), поэтому в результате фотосинтеза образуется кислород.
У бактерий роль донора электронов могут выполнять, например, серо-водород (сера), органические вещества.
В отношении света организмы стоят перед дилеммой: с одной стороны, прямое воздействие света на протоплазму клетки смертельно для организма, с другой — свет служит первичным источником энергии, без которого невозможна жизнь.
Видимый свет оказывает на организмы смешанное действие: красные лучи — тепловое воздействие; синие и фиолетовые лучи — изменяют скорость и направление биохимических реакций. В целом свет влияет на скорость роста и развития растений, на интенсивность фотосинтеза, на активность животных, вызывает изменение влажности и температуры среды, является важным фактором, обеспечивающим суточные и сезонные биологические циклы. Каждое местообитание характеризуется определенным световым режимом, определяемым интенсивностью (силой), количеством и качеством света.
Интенсивность (сила) света измеряется энергией, приходящейся на единицу площади в единицу времени: Дж/м2-с; Дж/см’ с. На этот фактор сильно влияют особенности рельефа. Самым интенсивным является прямой свет, однако более полно растениями используется рассеянный свет.
На этот фактор сильно влияют особенности рельефа. Самым интенсивным является прямой свет, однако более полно растениями используется рассеянный свет.
Количество света определяется суммарной радиацией. От полюсов к экватору количество света увеличивается. Для определения светового режима необходимо учитывать и количество отраженного света, так называемое альбедо (А). Альбедо (от лат. а1Ьиз — белый) — отражающая способность поверхностей различных тел — выражается в процентах от общей радиации и зависит от угла падения лучей и свойств отражающей поверхности. Например, альбедо чистого снега — 85%, загрязненного — 40-50%, черноземной почвы — 5-14%, светлого песка — 35-45%, полога леса — 10-18%, зеленых листьев клена — 10%, осенних пожелтевших листьев — 28%.
По отношению к свету как экологическому фактору различают следующие группы растений: гелиофиты, сциофиты и теневыносливые растения (факультативные гелиофиты).
Световые виды (гелиофиты) — обитают на открытых местах с хорошей освещенностью и в лесной зоне встречаются редко. Процесс фотосинтеза начинает преобладать над процессом дыхания только при высокой освещенности (пшеница, сосна, лиственница). Цветки таких светолюбивых растений, как подсолнечник, козлобородник, череда, поворачиваются за солнцем.
Процесс фотосинтеза начинает преобладать над процессом дыхания только при высокой освещенности (пшеница, сосна, лиственница). Цветки таких светолюбивых растений, как подсолнечник, козлобородник, череда, поворачиваются за солнцем.
Теневые растения (сциофиты) — не выносят сильного освещения и живут под пологом леса в постоянной тени (это в основном лесные травы, папоротники, мхи, кислица). На вырубках при сильном освещении они проявляют явные признаки угнетения и часто погибают.
Теневыносливые растения (факультативные гелиофиты) — могут жить при хорошем освещении, но легко переносят и затемненные места (большинство растений лесов, луговые растения, лесные травы и кустарники).
Теневыносливые древесные породы и теневые травянистые
растения отличаются мозаичным расположением листьев. У эвкалиптов листья обращены к свету ребром. У деревьев световые и
У деревьев световые и
теневые листья (располагаются соответственно по поверхности и
внутри кроны) — хорошо освещаемые и затененные — имеют анатомические различия. Световые листья толще и грубее, иногда
они блестящие, что способствует отражению света. Теневые листья обычно матовые, не опущенные, тонкие, с очень нежной кутикулой или вовсе без нее (кутикула — наружная пленка, покрывающая эпидермис).
В лесу теневыносливые деревья образуют густо сомкнутые насаждения. Под их пологом растут еще более теневыносливые деревья и кустарники, а ниже — теневые кустарнички и травы.
Наибольшее значение свет как средство ориентации имеет в
жизни животных. Уже у простейших появляются светочувствительные органеллы. Так, эвглена зеленая с помощью светочувствительного «глазка» реагирует на степень освещенности среды. Начиная с кишечнополостных, практически у всех животных
развиваются светочувствительные органы — глаза, имеющие то или иное строение.
Среди животных различают дневные, ночные и сумеречные
виды. Имеются также виды, живущие в постоянной темноте и
не выносящие яркого солнечного света (почвенные животные, обитатели пещер и больших глубин, внутренние паразиты животных и растений).
Происходит это в результате окисления сложных органических соединений при участии катализаторов обычно в ответ на раздражения, поступающие из внешней среды. Световые сигналы, испускаемые рыбами, головоногими моллюсками и другими гидробионтами, а также некоторыми организмами наземно-воздушной среды (например, жуками семейства светляков), служат для привлечения особей противоположного пола, приманивания добычи или отпугивания хищников, ориентации в стае и др
Это свечение вызвано живыми существами.
Вопросы и задания
Дайте характеристику света как абиотического фактора. Приведите классификацию экологических классов растений по отношению к свету.
Приведите классификацию экологических классов растений по отношению к свету.
Свет в жизни живых организмов.
Абиотические факторы: освещенность
Как недостаток солнечного света влияет на растение.
Для чего свет нужен растениям
 youtube.com/embed/ofqhi_V7so4″>
youtube.com/embed/ofqhi_V7so4″>
Ресурсы:
С. В. Алексеев. Экология: Учебное пособие для учащихся 9 класса общеобразовательных учреждений разных видов. СМИО Пресс, 1997. — 320 с
Сайт YouTube: https://www.youtube.com /
Хостинг презентаций
— http://ppt4web.ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
Опубликовано в группе «Учебный фильм — окружающий мир, естествознание, краеведение»
Адаптация растений к динамично меняющейся среде: реакция избегания тени
Обзор
. 2012 сен-октябрь;30(5):1047-58.
doi: 10.1016/j.biotechadv.2011.08.014.
Epub 2011 24 августа.
Я Руберти
1
, G Sessa, A Ciolfi, M Possenti, M Carabelli, G Morelli
принадлежность
- 1 Институт молекулярной биологии и патологии, Национальный исследовательский совет, Пьяццале Альдо Моро 5, Рим, Италия. [email protected]
PMID:
21888962
DOI:
10.1016/j.biotechadv.2011.08.014
Обзор
Я Ruberti et al.
Биотехнология Adv.
2012 сен-окт.
. 2012 сен-октябрь;30(5):1047-58.
doi: 10. 1016/j.biotechadv.2011.08.014.
1016/j.biotechadv.2011.08.014.
Epub 2011 24 августа.
Авторы
Я Руберти
1
, Г. Сесса, А. Чолфи, М. Поссенти, М. Карабелли, Г. Морелли
принадлежность
- 1 Институт молекулярной биологии и патологии, Национальный исследовательский совет, Пьяццале Альдо Моро 5, Рим, Италия. [email protected]
PMID:
21888962
DOI:
10.1016/j.biotechadv.2011.08.014
Абстрактный
Успех конкурентных взаимодействий между растениями определяет шансы на выживание отдельных особей и, в конечном счете, целых видов растений. Теневыносливые растения адаптировали свой фотосинтез для оптимального функционирования в условиях низкой освещенности. Поэтому эти растения способны к длительному выживанию в тени навеса. Напротив, растения, избегающие тени, приспосабливают свой рост к максимальному восприятию солнечного света и поэтому быстро занимают промежутки в кроне. Дневной свет содержит примерно равные пропорции красного и дальнего красного света, но в растительности это соотношение снижается в результате поглощения красного цвета фотосинтезирующими пигментами. Это изменение качества света воспринимается системой фитохромов как недвусмысленный сигнал о близости соседей, что приводит к набору реакций развития (называемых реакцией избегания тени), которые в случае успеха приводят к чрезмерному росту этих соседей. Удлинение побегов, вызванное слабым красным/дальним красным светом, может обеспечить высокую относительную приспособленность в естественных плотных сообществах. Однако, поскольку удлинение часто достигается за счет роста листьев и корней, избегание затенения может привести к снижению продуктивности культурных растений.
Теневыносливые растения адаптировали свой фотосинтез для оптимального функционирования в условиях низкой освещенности. Поэтому эти растения способны к длительному выживанию в тени навеса. Напротив, растения, избегающие тени, приспосабливают свой рост к максимальному восприятию солнечного света и поэтому быстро занимают промежутки в кроне. Дневной свет содержит примерно равные пропорции красного и дальнего красного света, но в растительности это соотношение снижается в результате поглощения красного цвета фотосинтезирующими пигментами. Это изменение качества света воспринимается системой фитохромов как недвусмысленный сигнал о близости соседей, что приводит к набору реакций развития (называемых реакцией избегания тени), которые в случае успеха приводят к чрезмерному росту этих соседей. Удлинение побегов, вызванное слабым красным/дальним красным светом, может обеспечить высокую относительную приспособленность в естественных плотных сообществах. Однако, поскольку удлинение часто достигается за счет роста листьев и корней, избегание затенения может привести к снижению продуктивности культурных растений. За последнее десятилетие был достигнут значительный прогресс в понимании молекулярных основ избегания тени. Однако выявление механизмов, лежащих в основе реакции растений и адаптации к изменениям соотношения красного и дальнего красного света, является ключом к разработке новых стратегий для точного регулирования уклонения от тени во времени и пространстве без ухудшения общей способности растений конкурировать за свет.
За последнее десятилетие был достигнут значительный прогресс в понимании молекулярных основ избегания тени. Однако выявление механизмов, лежащих в основе реакции растений и адаптации к изменениям соотношения красного и дальнего красного света, является ключом к разработке новых стратегий для точного регулирования уклонения от тени во времени и пространстве без ухудшения общей способности растений конкурировать за свет.
Copyright © 2011 Elsevier Inc. Все права защищены.
Похожие статьи
Теневыносливость: при выращивании в высоту не вариант.
Gommers CM, Visser EJ, St Onge KR, Voesenek LA, Pierik R.
Гоммерс С.М. и соавт.
Тенденции Растениевод. 2013 Февраль;18(2):65-71. doi: 10.1016/j.tplants.2012.09.008. Epub 2012 17 октября.
Тенденции Растениевод. 2013.PMID: 23084466
Различия в сигнале близости и избегании тени между ранними и поздними сукцессионными деревьями.

Гилберт И.Р., Джарвис П.Г., Смит Х.
Гилберт И.Р. и соавт.
Природа. 2001 14 июня; 411 (6839): 792-5. дои: 10.1038/35081062.
Природа. 2001.PMID: 11459056
Выход из тени.
Vandenbussche F, Pierik R, Millenaar FF, Voesenek LA, Van Der Straeten D.
Vandenbussche F, et al.
Curr Opin Plant Biol. 2005 г., октябрь; 8 (5): 462-8. doi: 10.1016/j.pbi.2005.07.007.
Curr Opin Plant Biol. 2005.PMID: 16040269
Обзор.
Функция белка DELLA в реакциях роста на сигналы кроны.
Джакович-Петрович Т., де Вит М., Воесенек Л.А., Пиерик Р.
Джакович-Петрович Т. и соавт.
Плант Дж. 2007 г., июль; 51 (1): 117–26. doi: 10.1111/j.1365-313X.2007.03122.x. Epub 2007 3 мая.
Завод Дж. 2007.PMID: 17488236
Избегание тени: сигнализация фитохромами и другие сигналы обнаружения соседей над землей.
Пьерик Р., де Вит М.
Пьерик Р. и соавт.
J Опытный бот. 2014 июнь; 65 (11): 2815-24. дои: 10.1093/jxb/ert389. Epub 2013 9 декабря.
J Опытный бот. 2014.PMID: 24323503
Обзор.


Посмотреть все похожие статьи
Цитируется
Влияние дальнего красного света на рост и морфологию салата при выращивании в помещении зависит от сорта.
Лю Дж., ван Ирсел М.В.
Лю Дж. и др.
Растения (Базель). 2022 14 октября; 11 (20): 2714. doi: 10.3390/plants11202714.
Растения (Базель). 2022.PMID: 36297739
Бесплатная статья ЧВК.Фактор транскрипции CmbHLh26 регулирует гомеостаз лепестков антоцианов при различном освещении у хризантемы.
Чжоу Л.Дж., Ван Ю., Ван Ю., Сун А., Цзян Дж., Чен С., Дин Б., Гуань З., Чен Ф.
Чжоу Л.Дж. и др.
Завод Физиол. 2022, 28 сентября; 190(2):1134-1152. doi: 10.1093/plphys/kiac342.
Завод Физиол. 2022.PMID: 35876821
Рапс ( Brassica napus ) Митоген-активируемая протеинкиназа 1 повышает устойчивость к затенению, регулируя способность фотосинтеза фотосистемы II.
Ван З., Лю М., Яо М., Чжан С., Цюй С., Ду Х., Лу К., Ли Дж., Вэй Л., Лян Ю.
Ван Цзи и др.
Фронт завод науч. 2022 2 июня; 13:902989. doi: 10.3389/fpls.2022.902989. Электронная коллекция 2022.
Фронт завод науч. 2022.PMID: 35720537
Бесплатная статья ЧВК.Летучие вещества растений и индуцированные травоядными летучие вещества из перца чили действуют как аттрактанты паразитоида тли Aphelinus varipes (Hymenoptera: Aphelinidae).
Али М.Ю., Насим Т., Чжан Дж., Пан М., Чжан Ф., Лю Т.Х.
Али М.И. и др.
Растения (Базель). 2022 19 мая; 11 (10): 1350. doi: 10.3390/plants11101350.
Растения (Базель). 2022.PMID: 35631774
Бесплатная статья ЧВК.Анатомия проростков березы повислой ( Betula pendula Roth.) и осины гибридной ( Populus tremuloides Michx. × Populus tremula L.) демонстрирует внутривидовые реакции на освещение In Vitro .
Зепс М., Кондратович Т., Григжде Э., Янсонс А., Зелтиньш П., Самсоне И., Матисонс Р.
Зепс М. и соавт.
Растения (Базель). 2022 18 апреля; 11 (8): 1097. doi: 10.3390/plants11081097.
Растения (Базель). 2022.PMID: 35448825
Бесплатная статья ЧВК.


Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
Растения на свету — PMC
1. Андерсон Дж. М., Осмонд Б. Реакция на солнце: компромиссы между акклиматизацией и фотоингибированием. В: Kyle DJ, Osmond B, Arntzen CJ, редакторы. Фотоингибирование. Амстердам: Эльзевир; 2001. С. 1–38. [Google Scholar]
Андерсон Дж. М., Осмонд Б. Реакция на солнце: компромиссы между акклиматизацией и фотоингибированием. В: Kyle DJ, Osmond B, Arntzen CJ, редакторы. Фотоингибирование. Амстердам: Эльзевир; 2001. С. 1–38. [Google Scholar]
2. Бьоркман О., Паулз С.Б. Движение листьев в тени видов Oxalis oregana L I. Реакция на уровень и качество света. Carneg Inst Wash Yearb. 1987; 80: 59–62. [Google Scholar]
3. Коллер Д. Световые движения листьев. Окружающая среда растительной клетки. 1990;13:615–632. [Google Scholar]
4. Чоу В.С., Андерсон Дж.М., Хоуп А.Б. Переменные стехиометрии от фотосистемы-II до реакционных центров фотосистемы-I. Фотосинтез рез. 1988; 17: 277–281. [PubMed] [Google Scholar]
5. Бруньоли Э., Бьоркман О. Рост хлопчатника в условиях постоянного стресса от засоления — влияние на схему распределения, устьичные и неустьичные компоненты фотосинтеза и рассеивание избыточной световой энергии. Планта. 1992; 187: 335–347. [PubMed] [Google Scholar]
6. Kloppstech K. Световая регуляция фотосинтетических генов. Завод Физиол. 1997;100:739–747. [Google Scholar]
Kloppstech K. Световая регуляция фотосинтетических генов. Завод Физиол. 1997;100:739–747. [Google Scholar]
7. Андерсон Дж.М., Чоу В.С., Пак Й.И. Великий замысел фотосинтеза: акклиматизация фотосинтетического аппарата к сигналам окружающей среды. Фотосинтез рез. 1995; 46: 129–139. [PubMed] [Google Scholar]
8. Андерссон Б., Аро Э.М. Протеолитическая активность и протеазы хлоропластов растений. Завод Физиол. 1997; 100:780–793. [Google Scholar]
9. Андерсон Дж.М., Чоу В.С., Гудчайлд Д.Дж. Организация тилакоидной мембраны при акклиматизации к солнцу/тени. Aust J Plant Physiol. 1988;15:11–26. [Google Scholar]
10. Мелис А. Динамика состава и функции фотосинтетической мембраны. Биохим Биофиз Акта. 1991; 1058: 87–106. [Google Scholar]
11. Бонавентура С., Майерс Дж. Флуоресценция и выделение кислорода из Chlorella pyrenoidosa . Биохим Биофиз Акта. 1969; 189: 366–383. [PubMed] [Google Scholar]
12. Horton P, Hague A. Исследования индукции флуоресценции хлорофилла в изолированных протопластах ячменя 4. Разрешение нефотохимического тушения. Биохим Биофиз Акта. 1988;932:107–115. [Google Scholar]
Разрешение нефотохимического тушения. Биохим Биофиз Акта. 1988;932:107–115. [Google Scholar]
13. Хортон П. Влияние окислительно-восстановительного потенциала на кинетику индукции флуоресценции в пехлоропластах 2. Сигмоидность. Биохим Биофиз Акта. 1981; 637: 152–158. [PubMed] [Google Scholar]
14. Allen JF, Bennett J, Steinback KE, Arntzen CJ. Фосфорилирование белков хлоропластов связывает окислительно-восстановительное состояние пластохинона с распределением энергии возбуждения между фотосистемами. Природа. 1981; 291:1–5. [Google Scholar]
15. Gal A, Shahak Y, Schuster G, Ohad I. Специфическая потеря фосфорилирования LHCII в Lemna , мутант-1073, лишенный комплекса цитохром-b6/f. ФЭБС латыши. 1987; 221: 205–210. [Google Scholar]
16. Bennett J. Фосфорилирование мембранных белков хлоропластов. Природа. 1977; 269: 344–346. [Google Scholar]
17. Bennett J. Фосфопротеины хлоропластов. Протеинкиназа мембран тилакоидов является светозависимой. ФЭБС латыши. 1979; 103: 342–344. [PubMed] [Google Scholar]
ФЭБС латыши. 1979; 103: 342–344. [PubMed] [Google Scholar]
18. Кефаль JE. Аминокислотная последовательность полипептидного сегмента, который регулирует адгезию мембран (укладку гран) в хлоропластах. Дж. Биол. Хим. 1983;258:9941–9948. [PubMed] [Google Scholar]
19. Барбер Дж. Свойства и организация фотосинтетических пигментов. Symp Soc Exp Biol. 1983; 36: 19–52. [PubMed] [Google Scholar]
20. Kyle DJ, Arntzen CJ. Фосфорилирование белка мембраны тилакоидов избирательно изменяет локальные поверхностные заряды мембран вблизи первичного акцептора фотосистемы II. Фотобиохимия Фотобиофиз. 1983; 5:11–25. [Google Scholar]
21. Паулз С.Б. Фотоингибирование фотосинтеза, вызванное видимым светом. Ann Rev Plant Physiol Plant Molec Biol. 1984;35:15–44. [Google Scholar]
22. Барбер Дж. Молекулярные основы уязвимости фотосистемы-II к повреждению светом. Aust J Plant Physiol. 1995; 22: 201–208. [Google Scholar]
23. Telfer A, He WZ, Barber J. Спектральное разрешение более чем одного донора электронов хлорофилла в изолированном комплексе фотосистема-II реакционный центр. Биохим Биофиз Акта. 1990;1017:143–151. [Google Scholar]
Биохим Биофиз Акта. 1990;1017:143–151. [Google Scholar]
24. Охад И., Кайл Д.Дж., Арнтцен С.Дж. Повреждение и восстановление мембранных белков — удаление и замена инактивированных 32-килодальтонных полипептидов в мембранах хлоропластов. Джей Селл Биол. 1984;99:481–485. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Weis E, Berry JA. Квантовая эффективность Фотосистемы-II в отношении энергозависимого тушения флуоресценции хлорофилла. Биохим Биофиз Акта. 1987; 894: 198–208. [Google Scholar]
26. Genty B, Briantais JM, Baker NR. Взаимосвязь между квантовым выходом фотосинтетического транспорта электронов и тушением флуоресценции хлорофилла. Биохим Биофиз Акта. 1989; 990: 87–92. [Google Scholar]
27. Schreiber U. Обнаружение кинетики быстрой индукции с помощью нового типа высокочастотно-модулированного флуориметра хлорофилла. Фотосинтез рез. 1986;9:261–272. [PubMed] [Google Scholar]
28. Оксборо К., Хортон П. Исследование регуляции и функции энергозависимого тушения в хлоропластах гороха. Биохим Биофиз Акта. 1988; 934: 135–143. [Google Scholar]
Биохим Биофиз Акта. 1988; 934: 135–143. [Google Scholar]
29. Хортон П., Рубан А.В., Уолтерс Р.Г. Регулирование светоуборки зеленых растений. Annu Rev Plant Physiol Plant Mol Biol. 1996; 47: 665–684. [PubMed] [Google Scholar]
30. Хортон П., Рубан А.В. Зависимое от дельта-pH тушение Fo-уровня флуоресценции хлорофилла в листьях шпината. Биохим Биофиз Акта. 1993;1142:203–206. [Google Scholar]
31. Рубан А.В., Рис Д., Ноктор Г.Д., Янг А., Хортон П. Длинноволновые виды хлорофилла связаны с усилением подавления высокоэнергетического возбуждения у высших растений. Биохим Биофиз Акта. 1991; 1059: 355–360. [Google Scholar]
32. Genty B, Goulas Y, Dimon B, Peltier G, Briantais JM, Moya I. Модуляция эффективности первичной конверсии в листьях. Фотосинтез рез. 1992; 34:106. [Google Scholar]
33. Mullineaux CW, Ruban AV, Horton P. Быстрое выделение тепла, связанное с дельта-pH-зависимым гашением в мембранах тилакоидов шпината. Биохим Биофиз Акта. 1994;1185:119–123. [Google Scholar]
[Google Scholar]
34. Илиоая С., Джонсон М., Хортон П., Рубан А.В. Индукция эффективного рассеяния энергии в изолированном светособирающем комплексе фотосистемы II в отсутствие агрегации белков. Дж. Биол. Хим. 2008; 283:29505–29512. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Noctor G, Ruban AV, Horton P. Модуляция дельта-pH-зависимого нефотохимического тушения флуоресценции хлорофилла в хлоропластах шпината. Биохим Биофиз Акта. 1993;1183:339–344. [Google Scholar]
36. Рубан А.В., Янг А., Хортон П. Модуляция тушения флуоресценции хлорофилла в изолированном светособирающем комплексе фотосистемы-II. Биохим Биофиз Акта. 1994; 1186:3–127. [Google Scholar]
37. Niyogi KK, Shih C, Chow WS, Pogson BJ, DellaPenna D, Björkman O. Фотозащита у двойного мутанта арабидопсиса с дефицитом зеаксантина и лютеина. Фотосинтез Рез. 2001; 67: 139–145. [PubMed] [Google Scholar]
38. Рубан А.В., Уолтерс Р.Г., Хортон П. Молекулярный механизм контроля диссипации энергии возбуждения в мембранах хлоропластов — ингибирование дельта-pH-зависимого тушения флуоресценции хлорофилла дициклогексилкарбодиимидом. ФЭБС латыши. 1992;309:175–179. [PubMed] [Google Scholar]
ФЭБС латыши. 1992;309:175–179. [PubMed] [Google Scholar]
39. Уолтерс Р.Г., Рубан А.В., Хортон П. Идентификация протонно-активных остатков в светособирающем комплексе высших растений. Proc Nat Acad Sci USA. 93:14204–14209. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Li X-P, Bjorkman O, Shih C, Grossman AR, Rosenquist M, Jansson S, Niyogi KK. Пигментсвязывающий белок, необходимый для регуляции фотосинтетического сбора света. Природа. 2000; 403:391–395. [PubMed] [Академия Google]
41. Bonente G, Howes BD, Caffarri S, Smulevich G, Bassi R. Взаимодействия между субъединицей фотосистемы II PsbS и ксантофиллами, изученными in vivo и in vitro. Дж. Биол. Хим. 2008; 283:8434–8445. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Li XP, Gilmore AM, Caffari S, Bassi R, Golan T, Kramer D, Niyogi KK. Регуляция сбора света включает определение pH внутритилакоидного просвета белком PsbS. Дж. Биол. Хим. 2004; 279:22866–22874. [PubMed] [Google Scholar]
43. Bergantino E, Segalia A, Brunetta A, Teardo E, Rogoni F, Giacometti GM, Szabo I. Свето- и pH-зависимые структурные изменения в белке PsbS хотосистемы II. Proc Nat Acad Sci USA. 2003; 100:15265–15270. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Bergantino E, Segalia A, Brunetta A, Teardo E, Rogoni F, Giacometti GM, Szabo I. Свето- и pH-зависимые структурные изменения в белке PsbS хотосистемы II. Proc Nat Acad Sci USA. 2003; 100:15265–15270. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Тирдо Э., Полверино де Лаурето П., Бергантино Э., Далла Веккиа Ф., Ригони Ф., Сабо И., Джакометти Г.М. Доказательства взаимодействия PsbS с фотосинтетическими комплексами тилакоидов кукурузы. Биохим Биофиз Акта. 2007; 1767: 703–711. [PubMed] [Google Scholar]
45. Kiss A, Crouchman S, Ruban AV, Horton P. Белок PsbS контролирует организацию антенны фотосистемы II в мембранах тилакоидов высших растений. Дж. Биол. Хим. 2008; 283:3972–3978. [PubMed] [Академия Google]
46. Li XP, Muller-Moule P, Gilmore AM, Niyogi KK. PsbS-зависимое усиление девозбуждения по обратной связи защищает фотосистему II от фотоингибирования. Proc Nat Acad Sci USA. 2002;99:15222–15227. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Кирхгоф Х. Молекулярная скученность и порядок в фотосинтетических мембранах. Тенденции Растениевод. 2008; 13: 201–207. [PubMed] [Google Scholar]
Кирхгоф Х. Молекулярная скученность и порядок в фотосинтетических мембранах. Тенденции Растениевод. 2008; 13: 201–207. [PubMed] [Google Scholar]
48. Хортон П., Рубан А.В., Рис Д., Ноктор Г., Паскаль А.А., Янг А. Контроль светособирающей функции мембран хлоропластов путем агрегации белкового комплекса хлорофилла LHCII. ФЭБС латыши. 1991;292:1–4. [PubMed] [Google Scholar]
49. Yamamoto HY, Nakayama TOM, Chichester CO. Исследования световых и темных взаимопревращений ксантофиллов листьев. Арх Биохим Биофиз. 1962; 97: 168–173. [PubMed] [Google Scholar]
50. Хортон П., Рубан А.В., Вентворт М. Аллостерическая регуляция светособирающей системы фотосистемы II. Фил Транс Рой Сок Лондон Б. 2000; 355: 1361–1370. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Хортон П., Вентворт М., Рубан А. Контроль светособирающей функции мембран хлоропластов: модель агрегации LHCII для нефотохимического тушения II. ФЭБС латыши. 2005;579: 4201–4206. [PubMed] [Google Scholar]
52. Duffy CDP, Ruban A, Barford W. Возможная роль сильно связанных димеров хлорофилла в фотозащите LHCII. J Phys Chem. 2008; 112:12508–12515. [PubMed] [Google Scholar]
Duffy CDP, Ruban A, Barford W. Возможная роль сильно связанных димеров хлорофилла в фотозащите LHCII. J Phys Chem. 2008; 112:12508–12515. [PubMed] [Google Scholar]
53. Ruban AV, Berera R, Ilioaia C, Stokkum IHM, Kennis JTM, Pascal AA, van Amerongen H, Robert B, Horton P, van Grondelle R. Идентификация механизма фотозащитной энергии диссипация у высших растений. Природа. 2007; 450: 575–578. [PubMed] [Академия Google]
54. Лю З.Ф., Ян Х.К., Ван К.Б., Куанг Т.И., Чжан Д.П., Гуй Л.Л., Ан Х.М., Чанг В.Р. Кристаллическая структура комплекса, собирающего основной свет из шпината, с разрешением 2,72 Å. Природа. 2004; 428: 287–292. [PubMed] [Google Scholar]
55. Холт Н.Е., Зигмантас Д., Валкунас Л., Ли Х.П., Нийоги К.К., Флеминг Г.Р. Образование каротиноидных катионов и регуляция фотосинтетического сбора света. Наука. 2005; 307: 433–436. [PubMed] [Google Scholar]
56. Ан Т.К., Авенсон Т., Баллоттари М., Ченг Ю.К., Нийоги К.К., Басси Р., Флеминг Г.Р. Архитектура состояния переноса заряда, регулирующего сбор света в белке антенны растения.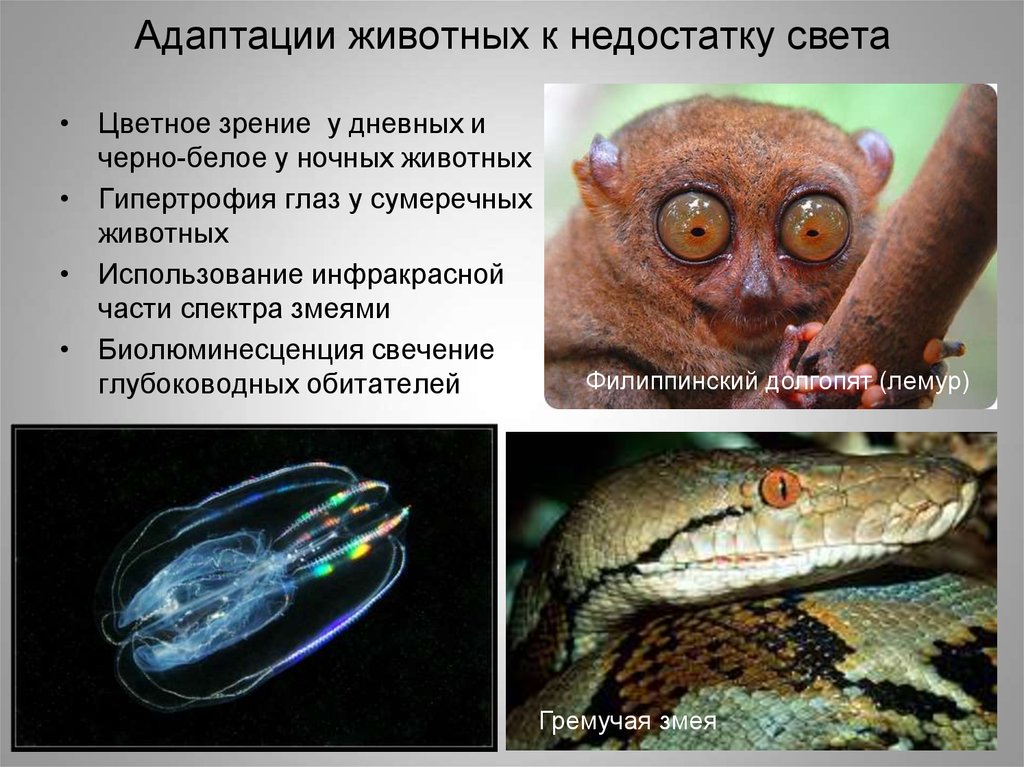 Наука. 2008;320:794–797. [PubMed] [Google Scholar]
Наука. 2008;320:794–797. [PubMed] [Google Scholar]
57. Джонсон М.П., Перес-Буэно М.Л., Зия А., Хортон П., Рубан А.В. Зеаксантин-независимый и зеаксантин-зависимый компоненты qE нефотохимического тушения включают общие конформационные изменения в антенне Photosystem II у Arabidopsis thaliana . Завод Физиол. 2008 г.: 10.1104/стр.109.129957. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
58. Джонсон Г.Н., Янг А.Дж., Скоулз Д.Д., Хортон П. Рассеивание избыточной энергии возбуждения у британских видов растений. Окружающая среда растительной клетки. 1993;16:673–679. [Google Scholar]
59. Рубан А.В., Янг А.Дж., Хортон П. Индукция нефотохимических изменений рассеивания энергии и поглощения в листьях — свидетельство изменений состояния светособирающей системы Фотосистемы-II in-vivo . Завод Физиол. 1993; 102: 741–750. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Horton P, Johnson M, Perez M, Ruban AV. Регулирует ли структура и макроорганизация фотосистемы II в мембранах гран высших растений состояния светосбора? ФЕБС Дж.