
Найдите ошибки в приведённом ниже тексте. Укажите в ответе номера предложений, в которых были сделаны ошибки, и после исправьте эти ошибки.1. Основными, классическими методами селекции растений были и остаются гибридизация и отбор. 2. Массовый отбор применяют при селекции перекрестноопыляемых растений. 3. Многие культурные растения представляют собой полиплоидные организмы. 4. Г.Д. Карпеченко открыл явление полиплоидии у культурных растений. 5. Мул (гибрид осла и кобылы) получен методом мутагенеза. 6. В селекции домашних животных в случае индивидуального отбора важна оценка производителей
Найдите ошибки в приведённом ниже тексте. Укажите в ответе номера предложений, в которых были сделаны ошибки, и после исправьте эти ошибки.1. Основными, классическими методами селекции растений были и остаются гибридизация и отбор. 2. Массовый отбор применяют при селекции перекрестноопыляемых растений. 3. Многие культурные растения представляют собой полиплоидные организмы. 4. Г.Д. Карпеченко открыл явление полиплоидии у культурных растений. 5. Мул (гибрид осла и кобылы) получен методом мутагенеза. 6. В селекции домашних животных в случае индивидуального отбора важна оценка производителей — племенных животных, используемых для размножения. 7. Для выявления генов хозяйственно ценных признаков животных используется метод испытания производителей по потомству.
5. Мул (гибрид осла и кобылы) получен методом мутагенеза. 6. В селекции домашних животных в случае индивидуального отбора важна оценка производителей — племенных животных, используемых для размножения. 7. Для выявления генов хозяйственно ценных признаков животных используется метод испытания производителей по потомству.
Учебник Курсы Книги Тесты Вопросы Личный кабинет
Учебник Курсы Книги Тесты Вопросы
Личный кабинет
Задание ЕГЭ по биологии
Линия заданий — 24
Наслаждайтесь
интересным учебником
и решайте
десятки тестов
на Studarium,
мы всегда рады вам! =)
3511. Найдите ошибки в приведённом ниже тексте. Укажите в ответе номера предложений, в которых были сделаны ошибки, и после исправьте эти ошибки.
Найдите ошибки в приведённом ниже тексте. Укажите в ответе номера предложений, в которых были сделаны ошибки, и после исправьте эти ошибки.
1. Основными, классическими методами селекции растений были и остаются гибридизация и отбор. 2. Массовый отбор применяют при селекции перекрестноопыляемых растений. 3. Многие культурные растения представляют собой полиплоидные организмы. 4. Г.Д. Карпеченко открыл явление полиплоидии у культурных растений. 5. Мул (гибрид осла и кобылы) получен методом мутагенеза. 6. В селекции домашних животных в случае индивидуального отбора важна оценка производителей — племенных животных, используемых для размножения. 7. Для выявления генов хозяйственно ценных признаков животных используется метод испытания производителей по потомству.
Ошибки допущены в предложениях 4, 5, 6:
4) Г.Д. Карпеченко смог получить при помощи полиплоидии межвидовые гибриды культурных растений, которые давали плодовитое потомство
5) Мул — гибрид осла и кобылы — получен в результате межвидового скрещивания (результат скрещивания осла и кобылы)
6) В селекции домашних животных в случае индивидуального отбора одним из способов является оценка производителя по потомству
P. S. Нашли ошибку в задании? Пожалуйста, сообщите о вашей находке 😉
S. Нашли ошибку в задании? Пожалуйста, сообщите о вашей находке 😉
При обращении указывайте id этого вопроса — 3511.
P.S. Мы нашли статью, которая относится к данной теме, изучите ее — Селекция 😉
P.S.S. Для вас готово следующее случайное задание. Мы сами не знаем, но вас ждет что-то интересное!
Селекция растений: основные методы и роль естественного отбора. Самоопыление перекрестноопыляемых растений. Гетерозис. Полиплоидия и отдаленная гибридизация. Достижения селекции растений — Генетика — ОБЩАЯ БИОЛОГИЯ
В основе селекции растений лежит искусственный отбор, когда человек отбирает растения с интересующими его признаками. До XVI-XVII вв. отбор происходил бессознательно, то есть человек, например, отбирал для посева лучшие, самые крупные семена пшеницы, не задумываясь о том, что он изменяет растения в нужном ему направлении.
Только в последние столетия человек, еще не зная законов генетики, стал использовать отбор сознательно или целенаправленно, скрещивая те растения, которые удовлетворяли его в наибольшей степени.
Однако методом отбора человек не может получить принципиально новых свойств у разводимых организмов, так как при отборе можно выделить только те генотипы, которые уже существуют в популяции. Поэтому для получения новых пород и сортов животных и растений применяют гибридизацию, скрещивая растения с желательными признаками и в дальнейшем отбирая из потомства те особи, у которых полезные свойства выражены наиболее сильно. Например, один сорт пшеницы отличается прочным стеблем и устойчив к полеганию, а другой сорт с тонкой соломиной не заражается стеблевой ржавчиной. При скрещивании растений из двух сортов в потомстве возникают различные комбинации признаков. Но отбирают именно те растения, которые одновременно имеют прочную соломину и не болеют стеблевой ржавчиной. Так создается новый сорт.
Основные методы селекции вообще и селекции растений в частности — отбор и гибридизация.
Для перекрестноопыляемых растений применяют массовый отбор особей с желаемыми свойствами. В противном случае невозможно получить материал для дальнейшего скрещивания. Таким образом получают, например, новые сорта ржи. Эти сорта не являются генетически однородными.
Если же желательно получение «чистой линии», то есть генетически однородного сорта, то применяют индивидуальный отбор, при котором путем самоопыления получают потомство от одной единственной особи с желательными признаками. Таким методом были получены многие сорта пшеницы, капусты и др.
Для закрепления полезных наследственных свойств необходимо повысить гомозиготность нового сорта. Иногда для этого применяют самоопыление перекрестноопыляемых растений. При этом могут фенотипически проявиться неблагоприятные воздействия рецессивных генов.
Основная причина этого — переход многих генов в гомозиготное состояние. У любого организма в генотипе постепенно накапливаются неблагоприятные мутантные гены. Они чаще всего рецессивны и фенотипически не проявляются. Но при самоопылении они переходят в гомозиготное состояние, и возникает неблагоприятное наследственное изменение. В природе у самоопыляемых растений рецессивные мутантные гены быстро переходят в гомозиготное состояние, и такие растения погибают, выбраковываясь естественным отбором.
Они чаще всего рецессивны и фенотипически не проявляются. Но при самоопылении они переходят в гомозиготное состояние, и возникает неблагоприятное наследственное изменение. В природе у самоопыляемых растений рецессивные мутантные гены быстро переходят в гомозиготное состояние, и такие растения погибают, выбраковываясь естественным отбором.
Несмотря на неблагоприятные последствия самоопыления, его часто применяют у перекрестноопыляемых растений для получения гомозиготных («чистых») линий с нужными признаками. Это приводит к снижению урожайности. Однако затем проводят перекрестное опыление между разными самоопыляющимися линиями и в результате в ряде случаев получают высокоурожайные гибриды, обладающие нужными селекционеру свойствами. Это метод межлинейной гибридизации, при котором часто наблюдается эффект гетерозиса: гибриды первого поколения обладают высокой урожайностью и устойчивостью к неблагоприятным воздействиям. Гетерозис характерен для гибридов первого поколения, которые получаются при скрещивании не только разных линий, но и разных сортов и даже видов. К сожалению, эффект гетерозисной (или гибридной) мощности бывает сильным только в первом гибридном поколении, а в следующих поколениях постепенно снижается.
К сожалению, эффект гетерозисной (или гибридной) мощности бывает сильным только в первом гибридном поколении, а в следующих поколениях постепенно снижается.
Основная причина гетерозиса заключается в устранении в гибридах вредного проявления накопившихся рецессивных генов. Другая причина — объединение в гибридах доминантных генов родительских особей и взаимное усиление их эффектов. В селекции растений широко применяется экспериментальная полиплоидия, так как полиплоиды отличаются быстрым ростом, крупными размерами и высокой урожайностью.
В основе явления полиплоидии лежат следующие причины: каждому виду живых организмов присущ строго определенный набор хромосом. В половых клетках все хромосомы различны. Такой набор называется гаплоидным и обозначается буквой п. Клетки тела (соматические) обычно содержат двойной набор хромосом, называемый диплоидным (2n). Если хромосомы, удвоившиеся в процессе деления, не разойдутся в дочерние клетки, а останутся в одном ядре, то возникает явление кратного увеличения числа хромосом, называемое полиплоидией.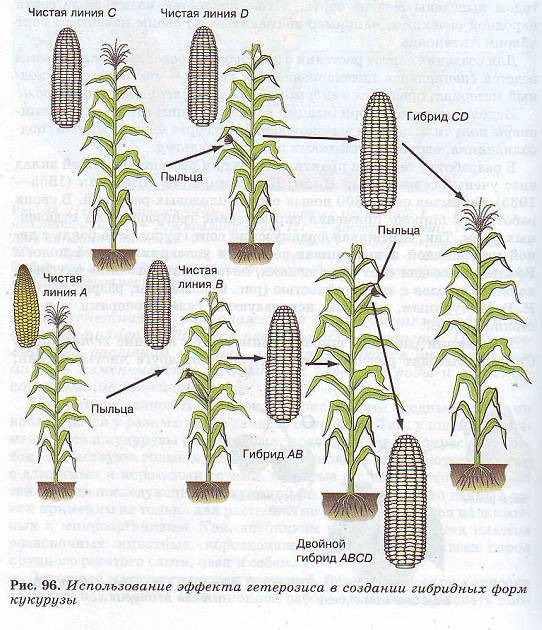 В сельскохозяйственной практике широко используются триплоидная сахарная свекла, четырехплоидные клевер, рожь и твердая пшеница, а также шестиплоидная мягкая пшеница.
В сельскохозяйственной практике широко используются триплоидная сахарная свекла, четырехплоидные клевер, рожь и твердая пшеница, а также шестиплоидная мягкая пшеница.
Получают искусственные полиплоиды при помощи химических веществ, которые разрушают веретено деления, в результате чего удвоившиеся хромосомы не могут разойтись и остаются в одном ядре. Одно из таких веществ — колхицин. Применение колхицина для получения искусственных полиплоидов является примером искусственного мутагенеза, применяемого при селекции растений.
Путем искусственного мутагенеза и последующего отбора мутантов были получены новые высокоурожайные сорта ячменя и пшеницы. Этими же методами удалось получить новые штаммы грибов, выделяющие в 20 раз больше антибиотиков, чем исходные формы.
Сейчас в мире культивируют более 250 сортов сельскохозяйственных растений, созданных при помощи физического и химического мутагенеза. Это сорта кукурузы, ячменя, сои, риса, томатов, подсолнечника, хлопчатника, декоративных растений.
При создании новых сортов при помощи искусственного мутагенеза исследователи используют закон гомологических рядов Н. И. Вавилова. Организм, получивший в результате мутации новые свойства, называют мутантом. Большинство мутантов имеет сниженную жизнеспособность и отсеивается в процессе естественного отбора. Для эволюции или селекции новых пород и сортов необходимы те редкие особи, которые имеют благоприятные или нейтральные мутации.
Отдаленная гибридизация — это метод, при котором производят скрещивание растений разных видов одного рода или даже разных родов. Существуют гибриды ржи и пшеницы, пшеницы и дикого злака эгилопс. Однако отдаленные гибриды, как правило, бесплодны, так как у них невозможен нормальный процесс созревания гамет из-за неспособности хромосом конъюгировать в профазе I мейоза.
К одному из достижений современной генетики и селекции относится преодоление бесплодия межвидовых гибридов. Впервые это удалось сделать Г. Д. Карпеченко при получении капустно-редечного гибрида. В результате отдаленной гибридизации было получено новое культурное растение — тритикале — гибрид пшеницы с рожью (лат. Triticum — пшеница и Secale — рожь). Отдаленная гибридизация широко применяется в плодоводстве.
В результате отдаленной гибридизации было получено новое культурное растение — тритикале — гибрид пшеницы с рожью (лат. Triticum — пшеница и Secale — рожь). Отдаленная гибридизация широко применяется в плодоводстве.
перекрестное опыление | Описание, механизмы и факты
перекрестное опыление
Просмотреть все средства массовой информации
- Похожие темы:
- перекрестное опыление
дихогамия
протогиния
протандрия
гетеростилия
Просмотреть все связанные материалы →
перекрестное опыление , также называемое гетерогамией , тип опыления, при котором пыльцевые зерна, содержащие сперму, переносятся с шишек или цветков одного растения на яйценосные шишки или цветки другого. — опыление наблюдается как у покрытосеменных (цветковые растения), так и у голосеменных (шишконосные растения) и способствует перекрестному опылению и аутбридингу. Это перемещение пыльцы может происходить ветром, как у хвойных, или посредством симбиотических отношений с различными животными (например, пчелами и некоторыми птицами и летучими мышами), которые переносят пыльцу с растения на растение, питаясь нектаром.
Преимущества перекрестного опыления
По сравнению с самоопылением (перенос пыльцы внутри цветка или между цветками одного и того же растения) перекрестное опыление явно имеет определенные эволюционные преимущества. Семена, образующиеся в результате аутбридинга, могут сочетать в себе наследственные признаки обоих родителей, и получающееся в результате потомство обычно бывает более разнообразным, чем в случае самоопыления. В изменяющейся среде генетическая изменчивость в популяции с перекрестным опылением может позволить некоторым особям адаптироваться к их новой ситуации, обеспечивая выживание вида, тогда как особи, появившиеся в результате самоопыления, могут оказаться неспособными приспособиться. Самоопыление или самоопыление, хотя и надежное в стабильной среде, таким образом, является эволюционным тупиком. Существует также более прямая, заметная разница между самоопылением и аутбридингом: у тех видов, где работают оба метода, перекрестное опыление обычно дает больше семян лучшего качества. Яркая демонстрация этого эффекта была обнаружена в гибридной кукурузе (маисе), превосходном продукте, полученном в результате скрещивания нескольких специально выведенных линий. Около половины наиболее важных культурных растений естественным образом перекрестно опыляются.
Яркая демонстрация этого эффекта была обнаружена в гибридной кукурузе (маисе), превосходном продукте, полученном в результате скрещивания нескольких специально выведенных линий. Около половины наиболее важных культурных растений естественным образом перекрестно опыляются.
Подробнее по этой теме
опыление: Типы: самоопыление и перекрестное опыление
Яйцеклетка в семязачатке цветка может быть оплодотворена сперматозоидом, полученным из пыльцевого зерна, произведенного тем же или другим цветком …
Механизмы, препятствующие самоопылению
В ходе эволюции многие цветковые растения ограничивали самоопыление и облегчали перекрестное опыление. Самоопыление может быть уменьшено или почти исключено за счет строения цветка, самонесовместимости и сроков созревания тычинок и пестиков одного и того же цветка или растения. Тогда перекрестное опыление может быть вызвано рядом агентов, главным образом насекомыми и ветром. Цветы, опыляемые ветром, обычно можно узнать по отсутствию цвета, запаха или нектара, а также по их рыльцам, которые расположены таким образом, чтобы оптимизировать захват переносимой по воздуху пыльцы. Цветы, опыляемые животными, бросаются в глаза благодаря своей структуре, цвету или производству аромата или нектара, и все это эволюционировало в тандеме с животными-опылителями.
Цветы, опыляемые животными, бросаются в глаза благодаря своей структуре, цвету или производству аромата или нектара, и все это эволюционировало в тандеме с животными-опылителями.
Структурные
Многие виды растений развили физические и структурные механизмы, предотвращающие самоопыление. Некоторые — например, финиковые пальмы ( Phoenix dactylifera ) и ивы ( Salix видов) — стали раздельнополыми; то есть некоторые растения производят только «мужские» (тычиночные) цветки, а остальные — только «женские» (пестичные или семязачаточные). У видов, у которых тычиночные и пестичные цветки встречаются на одной и той же особи (однодомные растения), а также у видов с гермафродитными цветками (цветки, обладающие как тычинками, так и пестиками), обычным способом предотвращения самоопыления является сброс пыльцы либо до, либо после периода, в течение которого рыльца на одном и том же растении восприимчивы, ситуация, известная как дихогамия. Более обычная форма дихогамии, особенно встречающаяся у таких опыляемых насекомыми цветов, как кипрей (9). 0037 Epilobium angustifolium ) и сальвии ( Salvia видов), является протандрией, у которой тычинки созревают раньше пестиков. Протогиния, ситуация, при которой пестики созревают первыми, встречается у арумовых лилий и многих ветроопыляемых растений, таких как травы, хотя некоторые травы самоопыляются, включая распространенные разновидности пшеницы, ячменя и овса. У авокадо есть как протогинные, так и протандрические сорта, и их часто выращивают вместе, чтобы стимулировать перекрестное опыление.
0037 Epilobium angustifolium ) и сальвии ( Salvia видов), является протандрией, у которой тычинки созревают раньше пестиков. Протогиния, ситуация, при которой пестики созревают первыми, встречается у арумовых лилий и многих ветроопыляемых растений, таких как травы, хотя некоторые травы самоопыляются, включая распространенные разновидности пшеницы, ячменя и овса. У авокадо есть как протогинные, так и протандрические сорта, и их часто выращивают вместе, чтобы стимулировать перекрестное опыление.
Особенностью строения цветков, препятствующей самоопылению, является гетеростилия, или вариация длины столбика (шейки пестика). Это происходит у примулы обыкновенной ( Primula vulgaris ) и видов кислицы ( Oxalis ) и льна ( Linum usitatissimum ). В большинстве популяций британской примулы, например, примерно половина особей имеет так называемые «булавочные» цветки, которые обладают короткими тычинками и длинным столбиком, придающим рыльцу положение во рту цветка, тогда как у другой половины есть цветки «барабанные». , у которых столбик короткий, а тычинки длинные, образующие «головку» при раскрытии цветка. Пчелы не могут не откладывать пыльцу, которую они получают от одного типа цветов, на рыльца другого типа. Генетическая система, которая регулирует структуру цветка у этих первоцветов, устроена таким образом, что перекрестное опыление автоматически поддерживает соотношение 50:50 между булавками и барабанчиками. В цветках вербейника пурпурного ( Lythrum salicaria ), тычинки и столбики имеют три разные длины, чтобы ограничить самоопыление.
, у которых столбик короткий, а тычинки длинные, образующие «головку» при раскрытии цветка. Пчелы не могут не откладывать пыльцу, которую они получают от одного типа цветов, на рыльца другого типа. Генетическая система, которая регулирует структуру цветка у этих первоцветов, устроена таким образом, что перекрестное опыление автоматически поддерживает соотношение 50:50 между булавками и барабанчиками. В цветках вербейника пурпурного ( Lythrum salicaria ), тычинки и столбики имеют три разные длины, чтобы ограничить самоопыление.
Химическая
Химическая самонесовместимость является еще одним средством предотвращения самооплодотворения. В этом явлении, которое зависит от химических веществ в растении, пыльца может не вырасти на рыльце того же цветка, который ее произвел, или после прорастания пыльцевая трубка может не расти нормально вниз по столбику, чтобы произвести оплодотворение. Это происходит у белого клевера ( Trifolium repens ), капуста ( Brassica oleracea ) и многие другие виды. Процесс контролируется генетически; он не обязательно должен быть абсолютным и может изменяться в степени в течение сезона цветения. Неудивительно, что химическая несовместимость обычно не обнаруживается у тех растений, которые имеют сильные структурные или временные барьеры против самоопыления. Формирования одного такого механизма в ходе эволюции, по-видимому, было достаточно для большинства видов растений.
Процесс контролируется генетически; он не обязательно должен быть абсолютным и может изменяться в степени в течение сезона цветения. Неудивительно, что химическая несовместимость обычно не обнаруживается у тех растений, которые имеют сильные структурные или временные барьеры против самоопыления. Формирования одного такого механизма в ходе эволюции, по-видимому, было достаточно для большинства видов растений.
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Подпишитесь сейчас
Редакторы Британской энциклопедии
Эта статья была недавно отредактирована и обновлена Мелиссой Петруцелло. . Некоторые растения размножаются путем самооплодотворения, когда пыльца растения может оплодотворять репродуктивную клетку или яйцеклетку того же растения. Другие растения допускают только перекрестное опыление, когда пыльца одного растения может оплодотворять только другое растение. Бесполое размножение также может происходить у растений (например, у побегов земляники), что дает новое растение, генетически идентичное своему родительскому растению. Все эти различия меняют способ работы селекционеров.
Все эти различия меняют способ работы селекционеров.
Дополнительные рекомендуемые знания
Содержимое
|
 5 Выборки с проверкой перекрестной производительности
5 Выборки с проверкой перекрестной производительностиВажность способа воспроизведения
Способ воспроизводства сельскохозяйственных культур определяет их генетический состав, который, в свою очередь, является решающим фактором для разработки подходящих методов селекции и селекции. Знание способа воспроизводства также необходимо для его искусственного манипулирования с целью получения улучшенных типов. Для культуры подходят только те методы селекции и селекции, которые не нарушают ее естественного состояния и не обеспечивают поддержание такого состояния. Именно по таким причинам навязывание самоопыления перекрестноопыляемым культурам приводит к резкому снижению их продуктивности.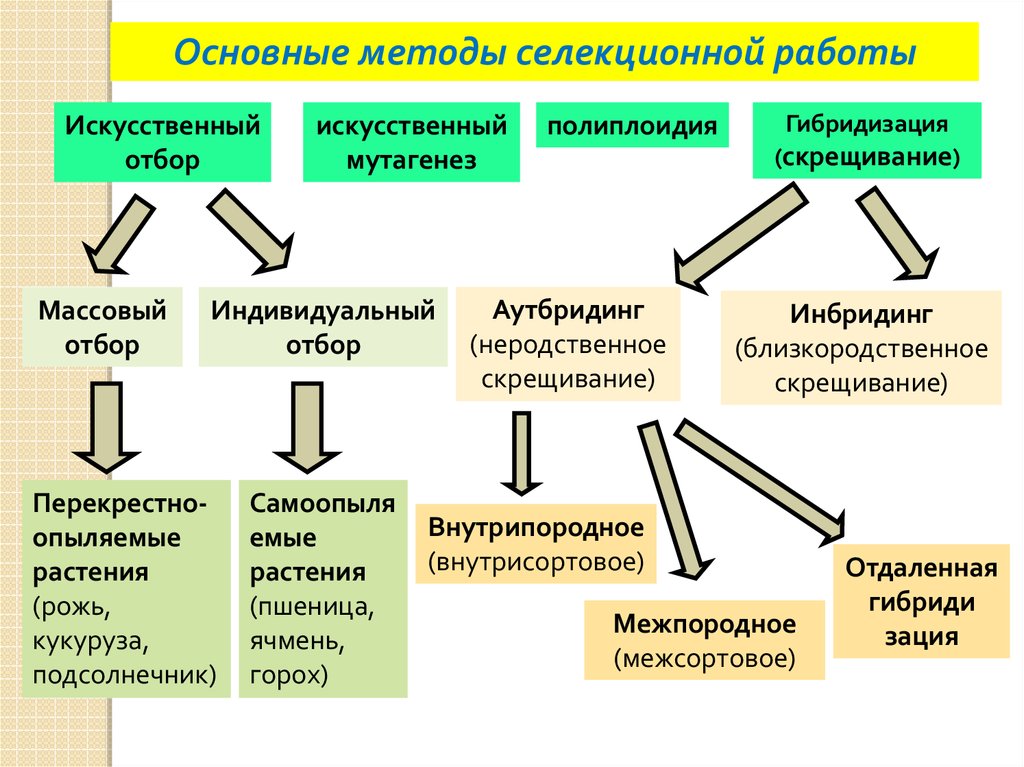 Точно так же практически невозможно поддерживать постоянную гетерозиготность у самоопыляющихся культур, что делает создание гибридов непривлекательным селекционным подходом. Бесполое размножение — еще один тип размножения, при котором любое растение или его часть можно использовать для размножения даже без незначительных генетических изменений от поколения к поколению. Таким образом, методы размножения и размножения таких культур полностью отличаются от методов размножения культур половым путем.
Точно так же практически невозможно поддерживать постоянную гетерозиготность у самоопыляющихся культур, что делает создание гибридов непривлекательным селекционным подходом. Бесполое размножение — еще один тип размножения, при котором любое растение или его часть можно использовать для размножения даже без незначительных генетических изменений от поколения к поколению. Таким образом, методы размножения и размножения таких культур полностью отличаются от методов размножения культур половым путем.
Самоопыляющиеся культуры (автогамные культуры)
Определенные ограничения привели к развитию механизмов самооплодотворения (частичного и полного самооплодотворения) у ряда видов растений.
Некоторые из причин, по которым самооплодотворяющийся метод воспроизводства настолько эффективен, заключаются в эффективности воспроизводства, а также в уменьшении генетической изменчивости и, таким образом, закреплении высоко адаптированных генотипов. Большинство локусов фиксируются с высокой скоростью; это можно объяснить тем, что с каждым поколением самооплодотворения доля гетерозигот уменьшается на 50%. Таким образом, гомозиготность будет получена в 5-8 поколениях. Третья причина эффективности самоопыления заключается в том, что в смешанных насаждениях самоопыляющихся и перекрестноопыляемых культур самоопыляющиеся растения могут отдавать пыльцу обоим типам растений, где перекрестноопыляемые растения ограничены в отношении вклада, который они могут внести в популяцию с помощью в отношении донорства пыльцы.
Таким образом, гомозиготность будет получена в 5-8 поколениях. Третья причина эффективности самоопыления заключается в том, что в смешанных насаждениях самоопыляющихся и перекрестноопыляемых культур самоопыляющиеся растения могут отдавать пыльцу обоим типам растений, где перекрестноопыляемые растения ограничены в отношении вклада, который они могут внести в популяцию с помощью в отношении донорства пыльцы.
Депрессия инбридинга почти не возникает у самоопыляющихся растений, потому что способ размножения позволяет естественному отбору иметь место в диких популяциях таких растений. Таким образом, генетически не превосходящие или неустойчивые растения удаляются из популяции на ранней эволюционной стадии. Популяции, полученные в результате самоопыления, иногда не так эволюционно адаптируются, как при использовании других методов воспроизводства, но, как известно, более эффективно используют определенные экологические ниши.
Важнейшими шагами в улучшении самоопыляющихся культур являются выбор родителей и идентификация лучших растений в сегрегационных поколениях. У заводчика также должны быть определенные цели при выборе родителей. Самоопыляющиеся сорта легче поддерживать, но это может привести к неправильному использованию семян.
У заводчика также должны быть определенные цели при выборе родителей. Самоопыляющиеся сорта легче поддерживать, но это может привести к неправильному использованию семян.
Некоторые из важных с агрономической точки зрения самоопыляющихся культур включают пшеницу, рис, ячмень, сухие бобы, соевые бобы, арахис, хлопок, томаты и т. д. интродукции, селекции и гибридизации.
К самоопыляемым культурам применяется несколько различных методов селекции, таких как массовая селекция, единичная селекция, родословная, массовая популяционная селекция, обратное скрещивание, рекуррентная селекция, гибрид F1, а также селекция по потомству одного семени (SSD) . В большинстве программ разведения применяется комбинация этих методов. Различные методы отбора можно резюмировать следующим образом:
Массовый отбор
Этот метод селекции зависит главным образом от отбора растений по их фенотипу и продуктивности. Семена отобранных растений собираются для следующего поколения. Этот метод используется для улучшения общей популяции путем положительного или отрицательного массового отбора. Массовая селекция лишь в ограниченной степени применяется к самоопыляющимся растениям и является эффективным методом улучшения местных сортов. Этот метод отбора будет эффективен только для высоко наследуемых признаков. Некоторые из недостатков массового отбора заключаются в том, что невозможно узнать, являются ли выбранные растения гетеро- или гомозиготными. Гетерозиготные растения будут сегрегировать в следующем поколении, что может потребовать повторения фенотипического отбора. Во-вторых, влияние окружающей среды на рост растений включает развитие, фенотип и производительность. Не всегда ясно, являются ли фенотипически превосходные растения также генотипически превосходными, и сильные различия в окружающей среде могут привести к низкой эффективности селекции.
Массовая селекция лишь в ограниченной степени применяется к самоопыляющимся растениям и является эффективным методом улучшения местных сортов. Этот метод отбора будет эффективен только для высоко наследуемых признаков. Некоторые из недостатков массового отбора заключаются в том, что невозможно узнать, являются ли выбранные растения гетеро- или гомозиготными. Гетерозиготные растения будут сегрегировать в следующем поколении, что может потребовать повторения фенотипического отбора. Во-вторых, влияние окружающей среды на рост растений включает развитие, фенотип и производительность. Не всегда ясно, являются ли фенотипически превосходные растения также генотипически превосходными, и сильные различия в окружающей среде могут привести к низкой эффективности селекции.
Выбор одного растения (чистый выбор линии)
Сорт, выведенный этим методом, будет более однородным, чем сорт, выведенный путем массовой селекции, потому что все растения в таком сорте будут иметь одинаковый генотип. Семена от выбранных растений не добавляют вместе, а хранят отдельно и используют для проведения тестов потомства. Это делается для изучения поведения размножения выбранных растений. В прошлом подчеркивалась высокая однородность роста и производительности, но риск развития узкоспециализированных патогенов очень высок. Большая генетическая изменчивость могла бы защитить культуру от таких патогенов, а также обеспечить стабильность производства в различных условиях окружающей среды.
Семена от выбранных растений не добавляют вместе, а хранят отдельно и используют для проведения тестов потомства. Это делается для изучения поведения размножения выбранных растений. В прошлом подчеркивалась высокая однородность роста и производительности, но риск развития узкоспециализированных патогенов очень высок. Большая генетическая изменчивость могла бы защитить культуру от таких патогенов, а также обеспечить стабильность производства в различных условиях окружающей среды.
Методы селекции для создания чистопородных сортов путем скрещивания
Скрещивания между сортами, введение зародышевой плазмы и линии разведения осуществляются для создания новых комбинаций генов. В последующих поколениях лучшие комбинации генов отбираются и фиксируются в гомозиготном состоянии посредством самооплодотворения и отбора. Эти отборы тщательно тестируются с целью выпуска их для выращивания.
Выбор родословной
Скрещивание родительских линий и отбор растений с новыми комбинациями генов происходит уже в поколении F2 (поколении растений, образованных от скрещивания гибридов F1). Потомство отобранных популяций в последующих поколениях многократно подвергается отбору до тех пор, пока не будет достигнуто генетическое единообразие. Записи ведутся о происхождении выбранных особей или линий. Количество поколений отдельных растений и линий селекции, а также интенсивность селекции на практике можно варьировать в зависимости от культуры и наличия средств.
Потомство отобранных популяций в последующих поколениях многократно подвергается отбору до тех пор, пока не будет достигнуто генетическое единообразие. Записи ведутся о происхождении выбранных особей или линий. Количество поколений отдельных растений и линий селекции, а также интенсивность селекции на практике можно варьировать в зависимости от культуры и наличия средств.
Обычно при использовании этого метода селекции основное внимание уделяется признакам с высокой наследуемостью, которые можно быстро и легко измерить. Одно из главных возражений против этого метода состоит в том, что генетическая изменчивость, доступная для отбора количественных признаков, резко снижается в последующих поколениях. Очистка семян и размножение обычно включаются в одно из последних поколений племенной селекции. Этот метод очень трудоемкий.
Эффективность селекции является одной из целей тестирования раннего поколения. Это делается путем раннего выявления высших гетерогенных популяций. Предполагается, что раннее устранение низших популяций и последующая концентрация селекционных усилий в пределах высших популяций приведет к повышению эффективности. Точная оценка гетерогенных популяций необходима для успеха этого метода и предполагает, что трансгрессивные сегреганты из низших популяций не будут превосходить по производительности выборки из высших популяций.
Точная оценка гетерогенных популяций необходима для успеха этого метода и предполагает, что трансгрессивные сегреганты из низших популяций не будут превосходить по производительности выборки из высших популяций.
Массовый выбор популяции
При этом методе селекции потомство от скрещивания высаживают с густотой посадки, равной плотности посадки товарной. В этот период, который может включать в себя ряд поколений, повышается уровень гомозиготности в основной популяции.
Этот метод прост и дешев и требует меньше усилий, чем родословная в предыдущих поколениях. Необходимо высаживать большие популяции, чтобы к началу селекции были отобраны лучшие сегрегации. Разделяющиеся поколения подвергаются еще одному этапу отбора растений. В более ранних поколениях ведется меньше записей, чем при отборе по родословной. Этот тип селекции особенно применим к культурам, которые обычно сажают с высокой плотностью посадки, например, мелкие зерновые культуры.
Выбор одного семени
Этот метод был введен, чтобы сохранить как можно больше растений F2 нетронутыми и, таким образом, предотвратить потерю изменчивости признаков с низкой наследуемостью в более ранних поколениях.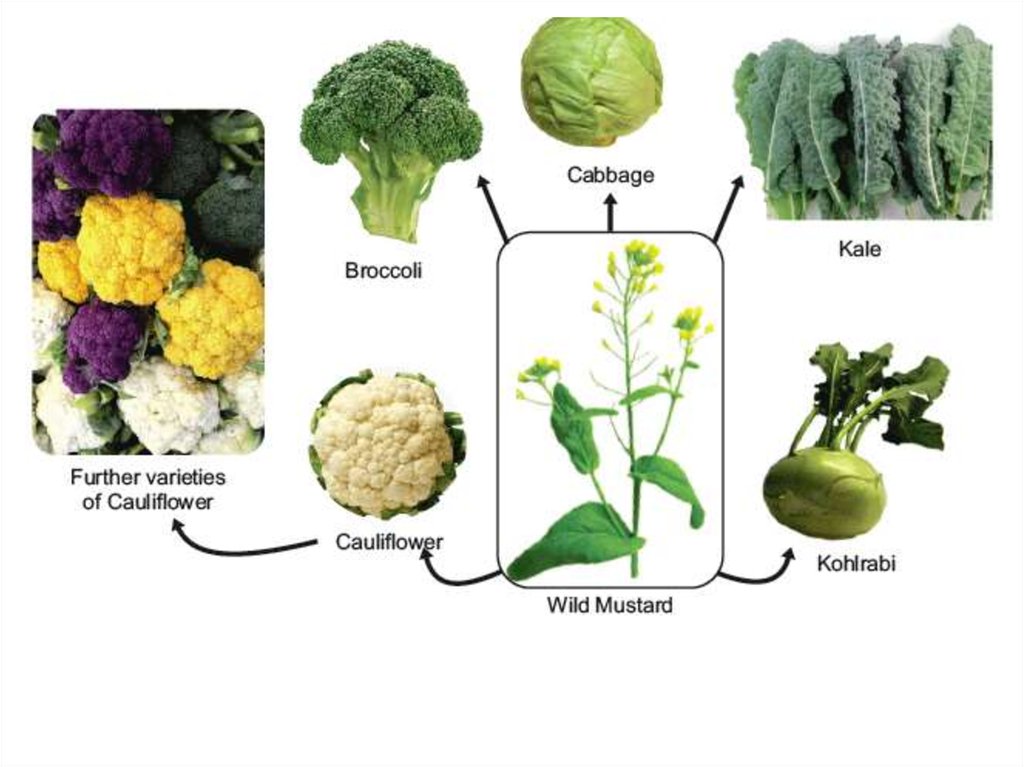 Этот метод также используется для уменьшения времени, необходимого для выращивания отдельных поколений. Поскольку с одного растения собирают только одно семя, в этом нет необходимости для оптимального развития растений, и можно манипулировать условиями, чтобы можно было собирать 2-3 поколения в год. Это может привести к сокращению времени процесса от начала до конца на 1-3 года.
Этот метод также используется для уменьшения времени, необходимого для выращивания отдельных поколений. Поскольку с одного растения собирают только одно семя, в этом нет необходимости для оптимального развития растений, и можно манипулировать условиями, чтобы можно было собирать 2-3 поколения в год. Это может привести к сокращению времени процесса от начала до конца на 1-3 года.
Этот метод не устраняет слабые растения, как в других методах, и также не предусматривается отбор лучших растений в поколении F2. Возможна модификация этого метода, и в ранних поколениях нет необходимости вести учет.
Двойной гаплоидный метод
Гаплоидные растения могут быть получены путем элиминации хромосом при широком скрещивании, культуре семяпочек или культуре пыльников. Культура пыльников, однако, в основном используется из-за ее способности производить гаплоидные растения в гораздо большем количестве по сравнению с двумя другими методами. Стрессы обычно необходимы, чтобы изменить пути развития микроспор от производства пыльцы к формированию гаплоидных растений.
Число хромосом гаплоидных растений затем удваивается с использованием колхицина. Однако спонтанно удвоенные гаплоидные растения также могут быть получены непосредственно тремя методами. Можно использовать методы спасения эмбрионов, чтобы гарантировать, что семена от этих широких скрещиваний или культурных растений стигмы выживут и не будут абортированы. Этот метод может сократить циклы генетического улучшения по сравнению с родословными или массовыми методами. Как и при методе односеменной селекции, ранние поколения не подлежат отбору, но большинство линий отсеивается в ходе испытаний по оценке земель.
Этот метод очень трудоемкий и самый дорогой из процедур, который увеличивает количество поколений в год. Чтобы этот метод был успешным, растения должны быть генетически стабильными.
Обратное скрещивание: метод селекции для улучшения генотипов
Это тип повторной селекции, при которой определенный ген может быть включен в превосходные сорта. Один из родительских сортов высокопродуктивен и коммерчески успешен, но ему не хватает определенного гена (например, устойчивости к болезням). Эта черта обычно присутствует у другого родительского сорта. После каждого обратного скрещивания гибридные растения идентифицируют с рассматриваемым геном и снова подвергают обратному скрещиванию с повторяющимся родителем.
Эта черта обычно присутствует у другого родительского сорта. После каждого обратного скрещивания гибридные растения идентифицируют с рассматриваемым геном и снова подвергают обратному скрещиванию с повторяющимся родителем.
Этот метод удобен, когда добавляются признаки, которые легко наследуются, доминируют и легко идентифицируются у гибридных растений. Если нежелательные гены тесно связаны с полезными генами, нежелательные гены могут быть переданы вместе, и потомство может оказаться менее продуктивным. Одним из преимуществ метода обратного скрещивания является то, что нет необходимости в обширных испытаниях. Этот метод используется для создания гибридных сортов в самоопыляемых культурах и для установления мужской стерильности в родительских линиях.
Обратное скрещивание с использованием маркеров обычно применяется в программах разведения для генной интрогрессии.
Традиционные методы отбора
A Относительно небольшой генофонд создается с помощью традиционных методов селекции, не так много возможностей для рекомбинации генов и мало возможностей для разрыва блоков сцепления. Селекционеры стремятся преодолеть эти недостатки, увеличив количество производимых скрещиваний, а также начав тестирование продуктивности в более ранних поколениях и внедрив системы улучшения популяций (например, повторный отбор).
Селекционеры стремятся преодолеть эти недостатки, увеличив количество производимых скрещиваний, а также начав тестирование продуктивности в более ранних поколениях и внедрив системы улучшения популяций (например, повторный отбор).
Другие стратегии селекции включают мультилинии (сочетание генетически идентичных линий, которые имеют разные гены, например устойчивость к стеблевой ржавчине). Линии, которые в основном генетически идентичны, за исключением одного гена, называются изогенными линиями. Также используются смешанные сорта (композиты), которые менее однородны, чем чистые сорта, и их следует регулярно пересматривать, чтобы не допустить сдвигов в соотношении компонентов. Рекуррентная селекция чаще всего используется в культурах с перекрестным опылением, но может быть успешно использована в самоудобрении только при наличии гена мужской стерильности. Гибридные сорта являются очень важным методом самоопыления культур и имеют следующие преимущества:
- Гетерозис
- Легче создавать сорта с множественными генами устойчивости
- Привлекательный метод для частных селекционеров благодаря встроенным механизмам защиты сортов.


Выращивание гибридных семян было одной из основных целей садоводческой и сельскохозяйственной практики, поскольку гибридные растения более продуктивны (благодаря гибридной силе) и более однородны по качеству, чем растения, полученные в результате самоопыления или случайного опыления. Чтобы вырастить гибридные семена, необходимо избежать самоопыления и сибопыления (опыление растением одного и того же гибрида). Одним из методов является кастрация вручную линии, используемой в качестве родителя женского пола, которая затем естественным образом перекрестно опыляется пыльцой линии, выступающей в качестве родителя мужского пола, и высаживается в соседний ряд. Однако этот процесс является очень трудоемким и неизменно дорогостоящим. Если сельскохозяйственные растения можно сделать самонесовместимыми путем введения генов, контролирующих самонесовместимость, то все произведенные семена будут гибридами, полученными в результате перекрестного опыления между двумя разными линиями. Это облегчит производство и повысит урожай гибридных семян и в то же время снизит трудозатраты.
Селекция перекрестноопыляемых культур
Естественным состоянием самоопыляющихся культур является гомозиготность и генетическая однородность, тогда как перекрестноопыляемые культуры характеризуются высокой степенью гетерозиготности. Виды растений, у которых нормальным способом завязывания семян является высокая степень перекрестного опыления, имеют характерные репродуктивные особенности и структуру популяции. Существование самостерильности, самонесовместимости, несовершенных цветков и механических препятствий делает растение зависимым от чужеродной пыльцы для нормального развития. набор семян. Каждое растение получает смесь пыльцы от большого количества особей, каждая из которых имеет разные генотипы. Такие популяции характеризуются высокой степенью гетерозиготности с огромной свободной и потенциальной генетической изменчивостью, которая поддерживается в устойчивом состоянии за счет свободного потока генов между особями внутри популяции. Неуместно и может быть довольно опасно брать одного или нескольких человек для исследования или улучшения этих популяций. Повышенная пригодность гетерозигот по сравнению с гомозиготами перекрестно опыляемых культур использовалась в форме двух различных подходов к селекции, а именно улучшения популяции и гибридной селекции таких культур.
Повышенная пригодность гетерозигот по сравнению с гомозиготами перекрестно опыляемых культур использовалась в форме двух различных подходов к селекции, а именно улучшения популяции и гибридной селекции таких культур.
При создании гибридных сортов ставится цель выявить из популяции наиболее продуктивную гетерозиготу, которую затем производят с исключением других представителей популяции. Напротив, улучшение популяции предусматривает поэтапную элиминацию вредных и менее продуктивных аллелей посредством повторяющихся циклов селективного скрещивания более продуктивных генотипов. Улучшение популяции — это медленная, неуклонная и долгосрочная программа, в то время как производство гибридов направлено на максимизацию генетического прироста за гораздо меньшее время. Оба этих подхода к разведению дополняют друг друга, а не исключают друг друга, и основаны на надежной генетической теории. Различные методы отбора можно резюмировать следующим образом:
Массовый отбор
Это самый простой, легкий и самый старый метод селекции, при котором отдельные растения отбираются на основе их фенотипических характеристик, а для получения следующего поколения используется масса семян. Массовая селекция оказалась достаточно эффективной для улучшения кукурузы на начальных этапах, но ее эффективность, особенно для повышения урожайности, вскоре подверглась резкой критике, завершившейся усовершенствованием метода массовой селекции. Отбор после опыления не обеспечивает какого-либо контроля над родительской пыльцой, в результате чего эффективный отбор ограничивается только родителями-женщинами. Оценки наследуемости уменьшаются вдвое, поскольку для сбора семян используются только родители, тогда как источник пыльцы после этого неизвестен. произошло перекрестное опыление.
Массовая селекция оказалась достаточно эффективной для улучшения кукурузы на начальных этапах, но ее эффективность, особенно для повышения урожайности, вскоре подверглась резкой критике, завершившейся усовершенствованием метода массовой селекции. Отбор после опыления не обеспечивает какого-либо контроля над родительской пыльцой, в результате чего эффективный отбор ограничивается только родителями-женщинами. Оценки наследуемости уменьшаются вдвое, поскольку для сбора семян используются только родители, тогда как источник пыльцы после этого неизвестен. произошло перекрестное опыление.
Повторяющийся выбор
Этот тип отбора является усовершенствованной версией процедуры массового отбора и отличается следующим:
- Визуально отобранные особи из базовой популяции проходят тестирование потомства
- Особи, отобранные на основе данных испытаний потомства, скрещиваются друг с другом всеми возможными способами для получения семян для формирования новой базовой популяции.

Отбор полусибсов с тестированием потомства
Селекция производится на основе результатов испытаний потомства, а не фенотипического внешнего вида родительских растений. Семена от отобранных полусибсов, которые были опылены случайной пыльцой из популяции, выращивают в нереплицированных рядах потомства с целью селекции. Часть семян высаживают, чтобы определить урожайность или племенную ценность любого признака каждого растения. Семена из наиболее продуктивных рядов или остатки семян из выдающихся полусибсов объединяются для завершения одного цикла селекции.
Отбор полных сибсов с тестированием потомства
Ряд полнородственных семейств, каждое из которых получен путем скрещивания двух растений из базовой популяции, оценивается в повторных испытаниях. Часть каждого полного родственного семейства сохраняется для рекомбинации. На основе оценки оставшееся семя выбранных полнородных семей используется для рекомбинации лучших семей.
Отборы с тестовой перекрестной производительностью
Целью этого типа отбора является небольшое отклонение от концепции улучшения внутри популяции в том смысле, что популяция улучшается не только с точки зрения производительности, но и в отношении комбинирования способностей с конкретной эталонной популяцией. Он предполагает генетические модификации популяции с целью ее лучшего использования для эксплуатации гетерозиса. Он включает в себя три шага:
Он предполагает генетические модификации популяции с целью ее лучшего использования для эксплуатации гетерозиса. Он включает в себя три шага:
- Самоопыление и тестовое скрещивание особей
- Оценка тестовых скрещиваний в повторных испытаниях
- Рекомбинация из самоопыленных остатков семян отдельных растений
Выбор семейства самоопыляемых
Растения исходной базовой популяции самоопыляются для получения потомства S1, которое оценивается в следующем сезоне в повторных испытаниях в нескольких средах для выявления многообещающих семейств S1. Остаток семян S1 таких отобранных семейств затем рекомбинируют в третьем сезоне, в результате чего за три сезона завершается один цикл. Следовательно, единицами отбора и рекомбинации являются потомки S1.
Селекция культур, размножаемых бесполым путем
Бесполое размножение охватывает все те способы размножения растений, при которых не происходит нормального образования гамет и оплодотворения, что делает их явно отличными от нормальных семенных культур. При отсутствии полового размножения генетический состав размножаемого растительного материала остается практически таким же, как у исходного растения.
При отсутствии полового размножения генетический состав размножаемого растительного материала остается практически таким же, как у исходного растения.
Клоны материнских растений могут быть получены с точным генетическим составом материнского растения. Высшие растения отбираются и размножаются вегетативно; вегетативно размноженное потомство используется для создания стабильных сортов без каких-либо ухудшений из-за сегрегации комбинаций генов. Эта уникальная характеристика бесполого размножения помогла вывести ряд сортов фруктов и овощей, включая виноград, яблоки, груши и персики.
Улучшение бесполого растительного материала путем селекции
Отбор этих культур ограничивается материалом, завезенным из других источников, таких как полевые плантации. Многообещающие варианты проверяются в крупномасштабных испытаниях, которые, в случае успеха, могут быть размножены и выпущены для коммерческого выращивания. Улучшение растений, размножаемых бесполым путем, посредством индуцированных мутаций имеет явные преимущества и ограничения. Любая вегетативная посадка может быть обработана мутагенами, и даже отдельный желательный мутант или часть мутировавшей побеги (химеры) могут быть размножены как улучшенный тип исходного сорта.
Любая вегетативная посадка может быть обработана мутагенами, и даже отдельный желательный мутант или часть мутировавшей побеги (химеры) могут быть размножены как улучшенный тип исходного сорта.
Селекция бесполых растений
Отбор в случае бесполых растений можно определить как отбор наиболее продуктивного растения и его вегетативное размножение. Поскольку растения не являются полностью генетически стабильными, можно ожидать, что с годами будут происходить отклонения. Таким образом, отбор представляет собой непрерывный процесс, в ходе которого отклоняются или исключаются из программы отбора. Основной целью селекции является улучшение качества и урожайности будущих насаждений. Как и любой селекционер, селекционер должен хорошо знать характеристики рассматриваемого сорта. В процессе отбора бесполых растений можно использовать различные подходы, такие как массовый отбор и отбор клонов из блоков клонов.
При массовом отборе необходимо учитывать некоторые факторы при выборе растений в материнском блоке, например. виноградник. Время выбора является важным фактором, потому что вы должны выбирать, когда ясно проявляются большинство характеристик растения. Лучшее время для бесполых многолетников — непосредственно перед сбором урожая. Для достижения наилучших результатов выбранное растение необходимо оценить в течение следующего сезона, когда лучше всего визуализируются аномалии роста, деформация листьев и симптомы вируса. Массовый отбор проводится ежегодно на одном и том же растении в течение как минимум трех лет. Растение, не соответствующее требованиям в какой-либо год селекционного цикла, исключается из программы.
виноградник. Время выбора является важным фактором, потому что вы должны выбирать, когда ясно проявляются большинство характеристик растения. Лучшее время для бесполых многолетников — непосредственно перед сбором урожая. Для достижения наилучших результатов выбранное растение необходимо оценить в течение следующего сезона, когда лучше всего визуализируются аномалии роста, деформация листьев и симптомы вируса. Массовый отбор проводится ежегодно на одном и том же растении в течение как минимум трех лет. Растение, не соответствующее требованиям в какой-либо год селекционного цикла, исключается из программы.
Старые плантации, подвергшиеся воздействию суровых условий роста, рассматриваются как предпочтительные источники селекции. Видно, что растения, выросшие в этих условиях и хорошо зарекомендовавшие себя, обладают хорошими генетическими свойствами. На этих старых плантациях естественный отбор позаботился о самых плохих растениях.
Разработка нового клона
Разработка и регистрация новых клонов осуществляется путем местной селекции клонов на старых плантациях, а также импорта высококачественных клонов из-за рубежа для местной оценки.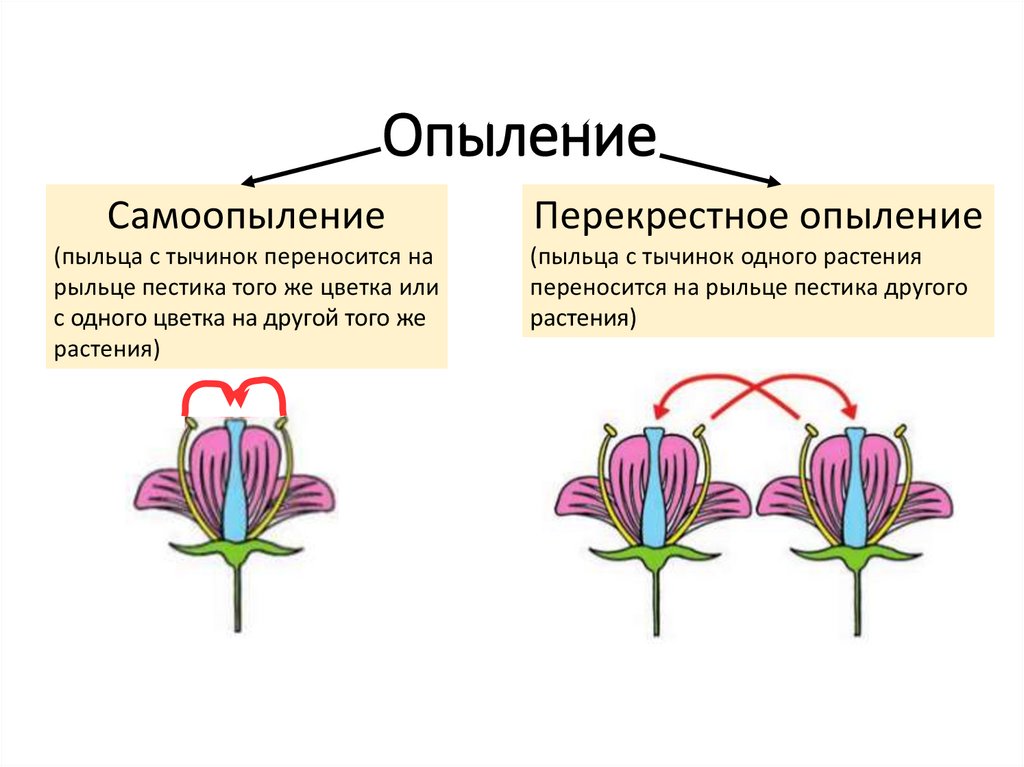
Клон – это вегетативное потомство одного определенного материнского растения; он не показывает каких-либо генетических, морфологических или физиологических отклонений от материнского растения. Оценка проводится с различными отобранными клонами после отбора. Непохожие клоны сравнивают друг с другом, чтобы определить их качество и устойчивость. Разведение не связано с отбором клонов; клон не может быть выведен на устойчивость к определенным типам вирусов, необходимо убедиться, что материал клона, покидающий питомник, не содержит вирусов. Разработаны методы проверки клонов на наличие вредоносных вирусов. Вредоносные вирусы иногда не проявляются в предварительных оценках. Таким образом, фитосанитарная разработка (выявление вирусов и уничтожение вирусов) осуществляется в лабораториях и теплицах параллельно с полевой оценкой и оценкой качества при полевых испытаниях клонов.
Когда клон соответствует минимальным стандартам качества и фитосанитарным стандартам, установленным Ассоциацией улучшения растений (PIA), он официально регистрируется для сертификации и коммерческого использования. PIA — это ассоциация, которая занимается улучшением всех растений, включая виноград, яблони, груши и персики.
PIA — это ассоциация, которая занимается улучшением всех растений, включая виноград, яблони, груши и персики.
Клоны получают из черенков выращенного в поле материнского растения. Из-за плохого управления и заражения с соседних плантаций в отборных плантациях есть только несколько свободных от вирусов материнских растений. Разработчики клонов должны были использовать такие методы, как культивирование тканей и размножение in vitro, чтобы получить клоны, свободные от вирусов, из ограниченного материнского материала. Апикальная меристема свободна от вредных вирусов. Используя апикальную меристему для культуры ткани, можно получить клон, свободный от вирусов.
Умножение
Во время первой фазы размножения вырезка ядра из каждого кандидата и/или зарегистрированного клона хранится в одобренной PIA теплице с блоком ядер без насекомых. Отсюда будут выполняться все будущие операции умножения и оценки.
На втором этапе размножения подвои и привитые черенки помещают в фундаментные блоки в помещениях, свободных от насекомых, и на изолированных участках открытого грунта, откуда проводится дальнейшая оценка.
Материал привоя и подвоя из источника второй фазы привит и окаймлен. Затем привитые растения высаживают на изолированных участках для создания материнских блоков. Эти блоки используются для умножения.
Третий этап – создание маточников привоев из вышеуказанного источника в хозяйствах договорных производителей-коллаборантов на заранее выбранной целине, из которых поддерживается около 4 км².
Каталожные номера
- ЧАХАЛ, Г.С. и ГОСАЛ, С.С., 2002. Принципы и процедуры селекции растений, Alpha Science International, Соединенное Королевство.
- Фальконер, Д.С., 1989. Введение в количественную генетику. 3-е изд. Лонгман. Сгоревшая мельница.
- FRISCH, M. & MELCHINGER, AE, 2005. Теория отбора для обратного скрещивания с помощью маркеров. Genetics: Published Articles Ahead of Print, опубликовано 31 марта 2005 г. под номером 10.1534/genetics.104.035451 [1]
- ГУССАР, П.Г. (2004) Улучшение материала виноградной лозы путем селекции. Департамент виноградарства. Стелленбосский университет. Конспект лекций.
- HOLSINGER, K.E., 2000. Репродуктивные системы и эволюция сосудистых растений. ПНАС. 20 июня, т. 1, с. 97, нет. 13: 7037-7042.
- KENNEDY, B.K., 2004. Определен генетический барьер для самоопыления. ЮрекОсторожно! 19 мая. [2]
- КОХЛИ, М.М. & FRANCIS, M., 2000. Применение биотехнологий в селекции пшеницы. Материалы конференции в Ла Эстансуэла, Уругвай, 19-20 ноября 1998 г. Монтевидео, Уругвай: CIMMYT. [3]
- KWV, Южная Африка. (2005). Установление новых мировых стандартов улучшения виноградников. Вититек ©
- МАРЕ, Г.Ф. 2005. Seleksiemetodes vir kruisbestuiwende gewasse. Кафедра генетики. Стелленбосский университет. Конспект лекций.
- TAKEBAYASHI, N. & MORELL, P.L., 2001. Является ли самооплодотворение эволюционным тупиком? Пересмотр старой гипотезы с генетическими теориями и макроэволюционным подходом. Американский журнал ботаники. 88:1143-1150.
- ВИВЬЕ, Массачусетс (2004 г.
 Стелленбосский университет. Конспект лекций.
Стелленбосский университет. Конспект лекций.