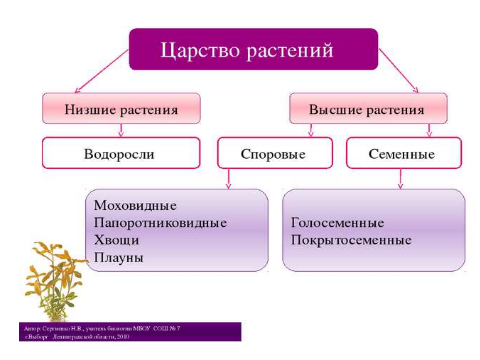
Содержание
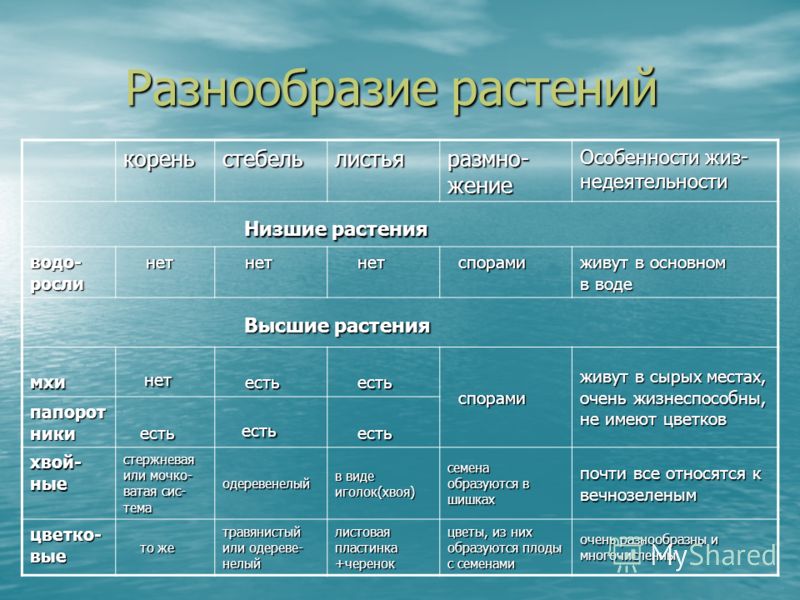
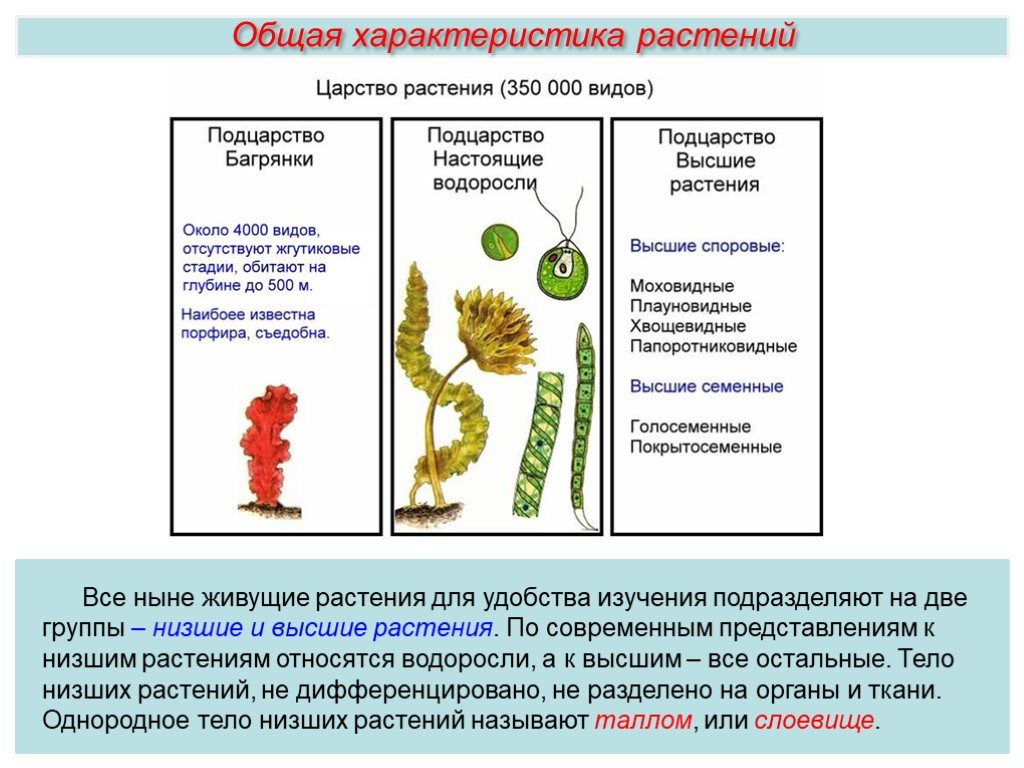
Общая характеристика высших растений
Высшие растения составляют подцарство растительного мира.
Существует предположение, что они произошли от каких-то
древних групп зеленых водорослей. Для такой гипотезы имеются
веские основания: 1) у водорослей и высших растений главный
фото-синтезирующий пигмент — хлорофилл а и каротиноиды;
2) основной запасной углевод — крахмал, который откладывается
в хлоропластах, а не в цитоплазме, как у других
фотосинтезирующих эукариот; 3) целлюлоза является важнейшим
компонентом их клеточной стенки; 4) у водорослей и некоторых
высших растений (мхи) в матриксе хлоропласта имеются особые
включения — пиреноиды; 5) при клеточном делении у растений и
некоторых водорослей образуются фрагмопласт — внутриклеточная
пластинка, зачаток клеточной стенки.
Высшие растения появились на суше примерно 430 млн. лет назад
лет назад
в виде небольших по размеру и примитивных по строению
риниофитов, или псилофитов. В дальнейшем их эволюция
неразрывно связана с постепенным завоеванием суши. Оказавшись
в совершенно иной наземновоздушной среде, они постепенно
адаптировались к необычной обстановке и на протяжении многих
миллионов лет дали громадное разнообразие наземных растений
различной величины и сложности строения.
Одним из ключевых событий раннего этапа выхода растений на
сушу было появление слор с прочными оболочками, позволяющими
переносить засушливые условия и распространяться ветром. В
процессе дальнейшего приспособления к наземным условиям
существования у высших растений сформировались вегетативные
органы — корень, стебель и лист, как результат
разделения функций между различными участками тела. Корни
обеспечивают закрепление растений в субстрате и
водно-минеральное питание, листья — фотосинтез, стебли —
транспорт веществ (восходящий и нисходящий токи).
Развитие эффективной проводящей системы тканей,
состоящей из ксилемы и флоэмы, решило проблему передвижения
воды и органических веществ у сосудистых растений при выходе
их на сушу. От высыхания эти растения обрели защиту в виде
покровной ткани — эпидермы или перидермы. Развитие
эпидермы привело к возникновению устьиц —
высокоспециализированных образований, регулирующих газообмен и
испарение воды растением.
Далее у высших растений произошло усиление механической
прочности путем утолщения клеточной стенки и пропитывания ее
лигнином, придающим жесткость ее целлюлозному остову.
Параллельно шла эволюция и органов полового (антеридии и
архегонии) и бесполого (спорангии) размножения. Из
одноклеточных, характерных для подавляющего большинства
водорослей, эти органы становятся многоклеточными, и стенки их
надежно защищают развивающиеся гаметы и споры от высыхания.
В жизненном цикле высших наземных растений наблюдается
закономерное чередование полового и бесполого поколений.
Гаплоидное поколение называется гаметофитом,
поскольку оно способно к половому размножению и образует
гаметы. Гаметы формируются в антеридиях и архегониях. В
результате оплодотворения образуется зигота, из которой
вырастает диплоидный спорофит. Он способен к бесполому
размножению с образованием гаплоидных спор. Последние дают
начало гаметофит-ному поколению. Одно из этих двух поколений
всегда преобладает над другим, и на его долю приходится
большая часть жизненного цикла высших растений.
В отличие от других высших растений, в жизненном цикле
моховидных преобладает гаметофит —небольшое, преимущественно
листостебельное растение, которое осуществляет функцию
фотосинтеза, водоснабжения и минерального питания. Спорофит у
Спорофит у
них развивается из оплодотворенной яйцеклетки внутри архегония
и постоянно связан с гаметофитом не только морфологически, но
и физиологически (в смысле питания), т. е. низведен до степени
органа растения, выполняющего только функцию спорообразования.
Для размножения мхов необходима вода, иначе сперматозоиды не
смогут подплыть к архегони-ям. Кроме того, только в достаточно
влажной среде у них лопаются антеридии и высвобождаются
сперматозоиды. Поэтому не случайно большинство моховидных
обитает в сырых и затененных местах.
Развитие жизненного цикла моховидных по пути возрастания
самостоятельности гаметофита и морфологического упрощения (с
потерей самостоятельности) спорофита привело к эволюционному
тупику.
В эволюции высших сосудистых растений происходит постепенная
редукция (уменьшение и упрощение) гаметофита и преобладание в
жизненном цикле спорофита. Так, у плауновидных, хвощевидных и
Так, у плауновидных, хвощевидных и
папоротниковидных гаметофит представлен маленьким (от
нескольких миллиметров до 3 см) заростком, не расчлененным на
органы, живущим несколько недель (у плаунов — несколько лет)
независимо от спорофита. На заростках в антеридиях
развиваются сперматозоиды, которые, плавая в каплях воды,
достигают архегония и сливаются с яйцеклеткой. Благодаря
крошечным размерам гаметофитов оплодотворение у хвощей,
плаунов и папоротников может происходить даже при ничтожно
малых количествах воды в виде капелек росы, тумана и Др.
У голо- и покрытосеменных растений гаметофит полностью утратил
способность к самостоятельному образу жизни, и все его
развитие протекает на спорофите внутри макроспорангия (или
семязачатка).
У голосеменных женский гаметофит — многоклеточный гаплоидный
эндосперм с двумя (у сосны) или несколькими (у других
голосеменных) архегониями; у покрытосеменных он редуцирован
обычно до семи клеток, архегониев не имеет и называется
зародышевым мешком.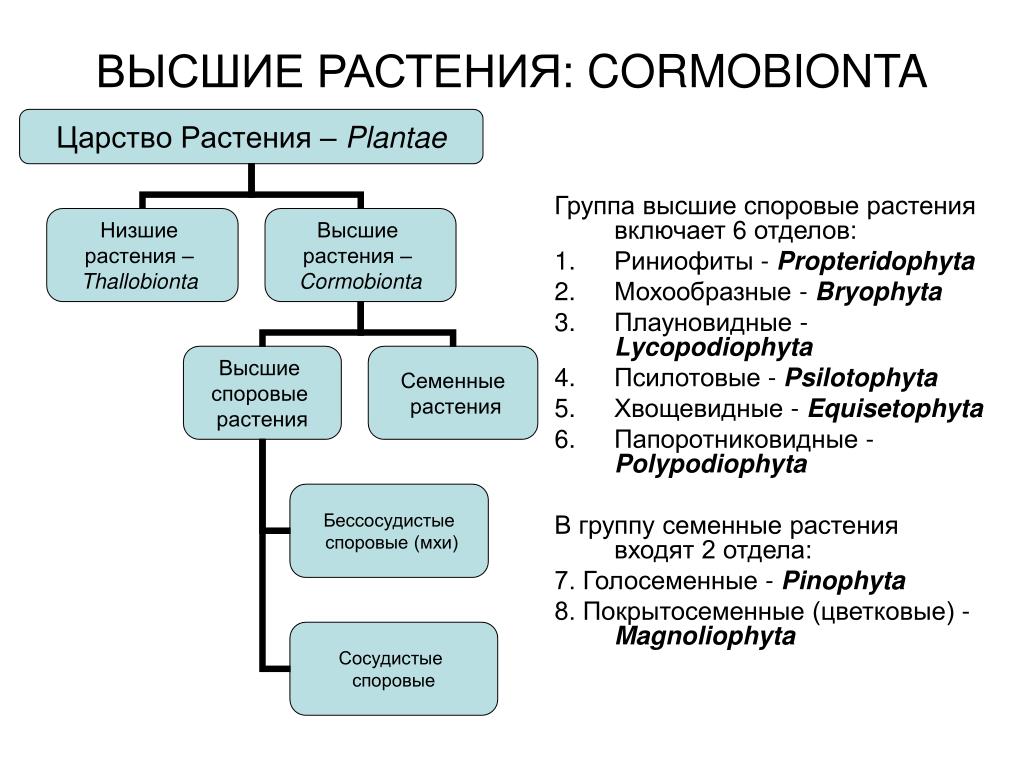 В последнем образуются яйцевой аппарат,
В последнем образуются яйцевой аппарат,
состоящий из яйцеклетки и двух клеток-синергид, вторичного
диплоидного ядра и клеток-антипод.
Мужской гаметофит семенных растений развивается из микроспоры
и представляет собой пыльцевое зерно (пыльцу), прорастающее в
пыльцевую трубку с образованием двух спермиев. При этом
впервые в эволюции растений процесс оплодотворения становится
независимым от наличия капельно-жидкой среды: спермин
доставляются к яйцеклеткам пыльцевой трубкой, что является
важнейшим приспособлением к наземному образу жизни.
Авторизация
Размер:
AAA
Цвет:
C
C
C
Изображения
Вкл.
Выкл.
Обычная версия сайта
Сведения об образовательной организации
Контакты
Старая версия сайта
Версия для слабовидящих
Версия для слабовидящих
Южно-Уральский государственный медицинский университет
- Университет
- События
- Новости
- ЮУГМУ сегодня
- Историческая справка
- Руководство
- Выборы ректора
- Лицензия, аккредитация и сертификаты
- Организационная структура
- Противодействие коррупции
- Первичная профсоюзная организация ЮУГМУ Профсоюза работников здравоохранения РФ
- Абитуриенту
- Новости для абитуриентов
- Центр довузовской подготовки
- Поступающим на специалитет
- Поступающим в ординатуру
- Поступающим в аспирантуру
- Поступающим в медицинский колледж
- Документы на право ведения образовательной деятельности
- Положения о приемной, экзаменационной и апелляционной комиссиях
- Информация об общежитиях
- Часто задаваемые вопросы
- Результаты приема студентов
- Информация для инвалидов
- Обучающемуся
- Факультеты
- Кафедры
- Медицинский колледж
- Ординатура
- Аспирантура
- Научная библиотека
- Образовательный портал
- Расписания
- Совет обучающихся ЮУГМУ
- Этический кодекс студентов медицинских вузов
- Совет студентов Минздрава России
- О допуске студентов к работе в медицинских организациях
- Иностранным обучающимся
- Медицинское обслуживание
- Информация об общежитиях
- Стипендиальное обеспечение
- Порядок перехода обучающихся с платного на бесплатное обучение
- Часто задаваемые вопросы
- Анкетирование
- Студенческие отряды
- Противодействие терроризму и экстремизму
- Специалисту
- Институт дополнительного профессионального образования
- Аккредитация специалистов
- Пациенту
- Клиника ФГБОУ ВО ЮУГМУ Минздрава России
- Профилактика новой коронавирусной инфекции, гриппа, ОРВИ, вакцинация
- Здоровый образ жизни
- Нет наркотикам!
- Научная работа
- Управление по научной и инновационной работе
- Экспериментально-биологическая клиника (виварий)
- Отдел международных связей
- НИИ иммунологии
- Центральная научно-исследовательская лаборатория
- НОЦ «Проблемы фундаментальной медицины»
- НОЦ «Клиническая фармакология»
- Конференции и другие мероприятия
- Диссертационные советы
- Журнал «Непрерывное медицинское образование и наука»
- Студенческое научное общество
- Совет молодых ученых и специалистов
Печеночник | Encyclopedia.
 com
com
Gale
Просмотры Обновлено мая 18 2018
Общие характеристики
Жизненный цикл
Распределение спор
Асексеальная репродукция
Evolution
Ресурсы
Ливер -репродукция. Мохообразная. Два других класса — это мхи и роголистники. Печеночники — небольшие зеленые наземные растения. У них нет настоящих корней, стеблей и листьев. Вместо этого они имеют надземную листовидную структуру, известную как слоевище, и подземную структуру, известную как ризоид. Большинство печеночников обитают во влажной среде и, как правило, менее устойчивы к высыханию, чем их родственники, мхи. Некоторые виды печеночников встречаются в Северной Америке с умеренным климатом, но большинство видов растет в тропиках.
Подобно мхам и высшим растениям, печеночники используют хлорофилл-а, хлорофилл-b и каротиноиды в качестве пигментов для фотосинтеза и хранят запасы пищи в виде крахмала. Как и у мхов и высших растений, их клеточные стенки состоят из целлюлозы.
Как и у мхов и высших растений, их клеточные стенки состоят из целлюлозы.
Подобно мхам и роголистникам, печеночники обитают только во влажной среде по двум основным причинам: во-первых, у них отсутствует сосудистая система для эффективного транспорта воды и пищи. Во-вторых, их сперматозоиды должны плыть по воде, чтобы добраться до яйцеклеток.
Талломы большинства печеночников имеют дорсивентральную морфологию. Другими словами, они имеют четкие лицевую и изнаночную стороны. В этом отношении слоевища печеночников сходны с листьями высших растений.
Название «печеночница» имеет многовековую историю и было дано этим растениям за то, что их слоевища имеют форму печени. В прежние времена люди верили в «доктрину сигнатур», согласно которой часть растения, похожая на орган тела, может использоваться для лечения заболеваний этого органа. Таким образом, печеночники использовались для лечения заболеваний печени. Западная наука давно дискредитировала учение о сигнатурах, хотя его до сих пор пропагандируют различные «нью-эйдж» и другие псевдонаучные течения.
Общие черты жизненного цикла печеночников такие же, как и у мхов. Для обоих характерно чередование поколений, в котором многоклеточный диплоидный спорофит находится в зависимости от зеленого и «лиственного» гаплоидного гаметофита. Как и у мхов, гаметофиты печеночников чаще всего встречаются в природе.
У большинства видов гаплоидная спора печеночника прорастает и дает начало одноклеточной протонеме,
небольшой нитевидной клетке. Как правило, гаплоидный гаметофит развивается из протонемы. У большинства печеночников гаметофит стелющийся, хотя у некоторых видов он прямостоячий. Как правило, гаметофит имеет подземный ризоид, специализированную одноклеточную структуру, которая прикрепляет печеночницу к субстрату и поглощает питательные вещества из почвы.
Мужские и женские репродуктивные органы, пыльники и архегонии вырастают из гаметофита. Они возникают непосредственно из слоевища или несутся на стеблях. Около 80% видов печеночников двудомные (самцы и самки на разных растениях), а остальные 20% — однодомные (самцы и самки на одном растении). Каждый архегоний производит одно яйцо; каждый антеридий производит много подвижных сперматозоидов, каждый с двумя жгутиками. Сперматозоиды должны плыть по воде, чтобы достичь архегония. Затем сперматозоиды оплодотворяют яйцеклетку, образуя диплоидную клетку. В конечном итоге он превращается в многоклеточный диплоидный спорофит.
Каждый архегоний производит одно яйцо; каждый антеридий производит много подвижных сперматозоидов, каждый с двумя жгутиками. Сперматозоиды должны плыть по воде, чтобы достичь архегония. Затем сперматозоиды оплодотворяют яйцеклетку, образуя диплоидную клетку. В конечном итоге он превращается в многоклеточный диплоидный спорофит.
Спорофит печеночников, как и спорофит мхов, имеет концевую капсулу на ножке, известную как щетинка. По мере развития спорофита внутри капсулы образуются гаплоидные споры. В целом спорофиты печеночников мельче и проще по морфологии, чем спорофиты мхов. Другое отличие состоит в том, что щетинка печеночника удлиняется после созревания капсулы, тогда как щетинка мха удлиняется до созревания капсулы.
Печеночники имеют характерный способ распространения спор. Внутри капсулы спорофита споры прикрепляются к специализированным клеткам, известным как элатеры. Элатер имеет трубчатую форму и имеет одно или несколько утолщений клеточной стенки, спирально ориентированных вдоль длинной оси клетки. Эти спиральные утолщения гигроскопичны, поскольку легко впитывают воду.
Эти спиральные утолщения гигроскопичны, поскольку легко впитывают воду.
КЛЮЧЕВЫЕ ТЕРМИНЫ
Диплоид —Ядро или клетка, содержащая две копии каждой хромосомы, образованные путем слияния двух гаплоидных ядер.
Elater — Специализированные трубчатые клетки со спирально ориентированными утолщениями клеточной стенки, к которым прикрепляются споры печеночника.
Гаметофит — гаплоидное поколение, производящее гаметы, в жизненном цикле растения.
Гемма — Многоклеточная бесполая репродуктивная структура мхов и печеночников.
Гаплоид — Ядро или клетка, содержащая по одной копии каждой хромосомы.
Мейоз — Деление клеточного ядра, при котором число хромосом уменьшается вдвое, обычно от диплоидного до гаплоидного.
Спорофит — Диплоидное спорообразующее поколение в жизненном цикле растения.
Слоевище — Одиночное тело растения без отчетливого стебля, листьев и корней.
При высыхании капсула печеночника открывается. Затем спиральные утолщения клеточных стенок элатера подсыхают, и элатер изменяет свою форму. При этом элатер выпускает связанные споры, которые затем разносятся ветром.
Как и мхи, многие виды печеночников размножаются, образуя геммы, небольшие круглые или сферические репродуктивные структуры, расположенные внутри чашечек гемм, которые формируются на вершине слоевища. Образование гемм — важная форма бесполого размножения у многих видов печеночников и мхов.
Есть только несколько окаменелостей печеночников и мхов и нет окаменелостей роголистников. Это связано с тем, что мягкие ткани этих бриофитов плохо окаменевают. Самая старая из известных окаменелостей печеночника относится к позднему девонскому периоду, около 350 миллионов лет назад. Большинство ботаников считают, что они возникли задолго до этого.
Некоторые ботаники предположили, что в мире существует более 10 000 видов печеночников. Более реалистичная оценка составляет около 6000 человек.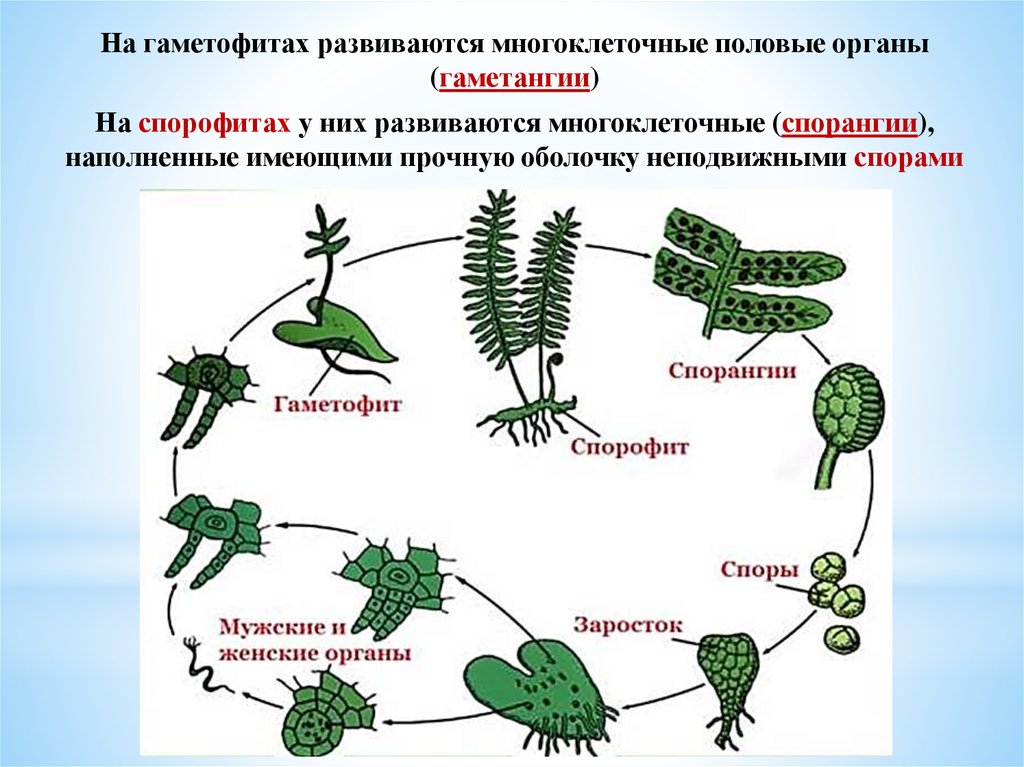 В прошлом количество видов могло быть переоценено, потому что морфология многих видов эластична, поскольку она различается в разных средах. Это делает идентификацию видов печеночников очень сложной, как правило, более сложной, чем идентификацию высших растений.
В прошлом количество видов могло быть переоценено, потому что морфология многих видов эластична, поскольку она различается в разных средах. Это делает идентификацию видов печеночников очень сложной, как правило, более сложной, чем идентификацию высших растений.
Интересно, что хотя печеночники возникли за несколько сотен миллионов лет до цветковых растений, существует несколько сотен тысяч видов цветковых растений, но только около 6000 видов печеночников. Причиной этого может быть то, что печеночники полагаются на неэффективный механизм переноса спермы водой для полового размножения. Таким образом, было высказано предположение, что большинство видов печеночников используют бесполые геммы как средство размножения. Бесполое размножение снижает генетическое разнообразие. Поскольку для развития новых видов необходимо генетическое разнообразие, печеночники и другие мохообразные, возможно, зашли в своего рода эволюционный тупик.
См. также Мохообразный.
КНИГИ
Гринуэй, Т.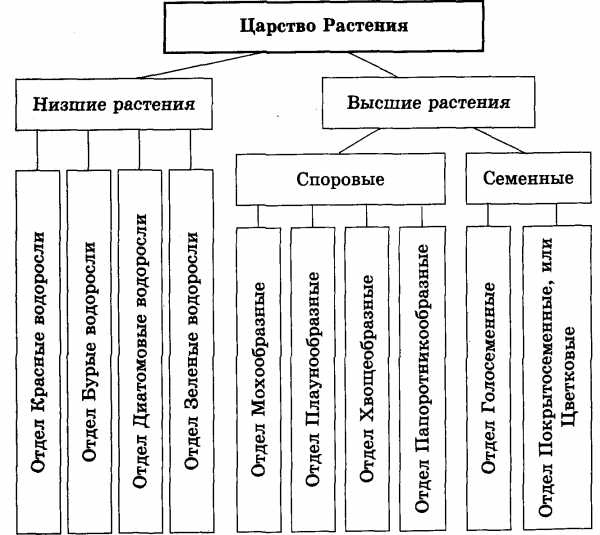 Мхи и печеночники. Остин, Техас: Steck-Vaughn, 1992.
Мхи и печеночники. Остин, Техас: Steck-Vaughn, 1992.
Маргулис, Л., и Шварц, К.В. Пять Королевств. Нью-Йорк: WH Freeman and Company, 1998.
ДРУГОЕ
Университет Южного Иллинойса Карбондейл. «Мохообразные: мхи, печеночники и роголистники»
Массачусетский университет, Амхерст: биологические науки. «Печеночники»
Peter A. Ensminger
The Gale Encyclopedia of Science
gale
просмотров обновлено 17 мая 2018 г.
Печеночники относятся к одному из трех классов растения . Два других класса — это мхи и роголистники. Печеночники — маленькие, зеленые, наземные растения. У них нет настоящих корней, стеблей и листьев. Вместо этого они имеют надземную листовидную структуру, известную как слоевище, и подземную структуру, известную как ризоид. Большинство печеночников обитают во влажной среде и, как правило, менее устойчивы к высыханию, чем их родственники, мхи. Много печеночников видов встречаются в умеренном климате Северной Америки , но большинство видов произрастает в тропиках.
Большинство печеночников обитают во влажной среде и, как правило, менее устойчивы к высыханию, чем их родственники, мхи. Много печеночников видов встречаются в умеренном климате Северной Америки , но большинство видов произрастает в тропиках.
Общие характеристики
Подобно мхам и высшим растениям, печеночники используют хлорофилл-а, хлорофилл-b и каротиноиды в качестве пигментов для фотосинтеза и запасают свои пищевые запасы в виде крахмала. Как и у мхов и высших растений, их стенки из клеток состоят из целлюлозы .
Подобно мхам и роголистникам, печеночники обитают только во влажной среде по двум основным причинам. Во-первых, у них отсутствует сосудистая система для эффективного транспорта вода и еда. Во-вторых, их сперматозоиды должны плыть по воде, чтобы добраться до яйцеклеток.
Талломы большинства печеночников имеют дорсивентральную морфологию. Другими словами, они имеют четкие лицевую и изнаночную стороны. В этом отношении слоевища печеночников сходны с листьями высших растений.
В этом отношении слоевища печеночников сходны с листьями высших растений.
Название «печеночница» имеет многовековую историю и было дано этим растениям, потому что их слоевища имеют форму печени. Раньше люди верили в доктрину подписей. Это диктовало, что часть растения, напоминающая тело орган можно использовать для лечения заболеваний этого органа. Таким образом, печеночники использовались для лечения заболеваний печени. Западная наука давно дискредитировала «доктрину сигнатур», хотя ее до сих пор отстаивают различные «нью-эйдж» и другие псевдонаучные течения.
Жизненный цикл
Общие черты жизненного цикла печеночников такие же, как у мхов. Оба имеют характерное чередование поколений, в котором многоклеточный диплоидный спорофит находится в зависимости от зеленого и «лиственного» гаплоидного гаметофита. Как и у мхов, гаметофиты печеночников чаще всего встречаются в природе.
У большинства видов гаплоидная спора печеночника прорастает и дает начало одноклеточной протонеме, небольшой нитевидной клетке.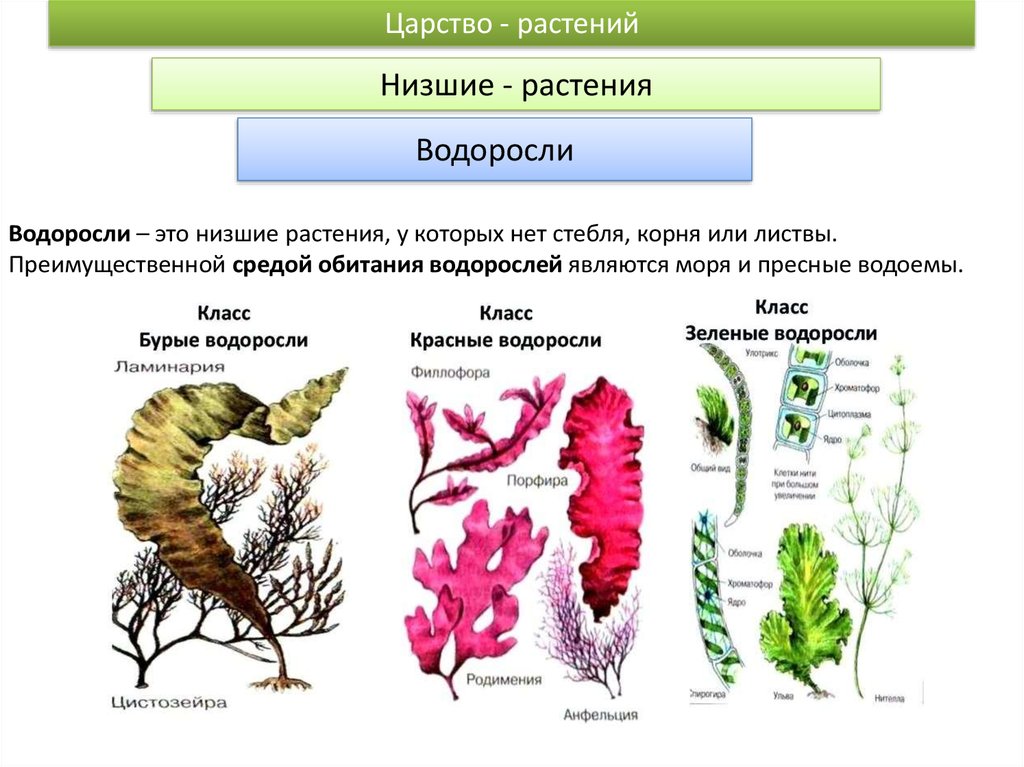 Как правило, гаплоидный гаметофит развивается из протонемы. У большинства печеночников гаметофит стелющийся, хотя у некоторых видов он прямостоячий. Как правило, гаметофит имеет подземный ризоид, специализированную одноклеточную структуру, которая прикрепляет печеночницу к субстрату и поглощает питательных веществ из почвы .
Как правило, гаплоидный гаметофит развивается из протонемы. У большинства печеночников гаметофит стелющийся, хотя у некоторых видов он прямостоячий. Как правило, гаметофит имеет подземный ризоид, специализированную одноклеточную структуру, которая прикрепляет печеночницу к субстрату и поглощает питательных веществ из почвы .
Мужские и женские репродуктивные органы, антеридии и архегонии, вырастают из гаметофита. Они возникают непосредственно из слоевища или несутся на стеблях. Около 80% видов печеночников двудомные (самцы и самки на разных растениях), а остальные 20% — однодомные (самцы и самки на одном растении). Каждый архегоний производит одно яйцо; каждый антеридий производит много подвижных сперматозоидов, каждый с двумя жгутиками . Сперматозоиды должны плыть по воде, чтобы достичь архегония. Затем сперматозоиды оплодотворяют яйцеклетку, образуя диплоидную клетку. В конечном итоге он превращается в многоклеточный диплоидный спорофит.
Спорофит печеночников, как и спорофит мхов, имеет концевую капсулу на ножке, известную как щетинка.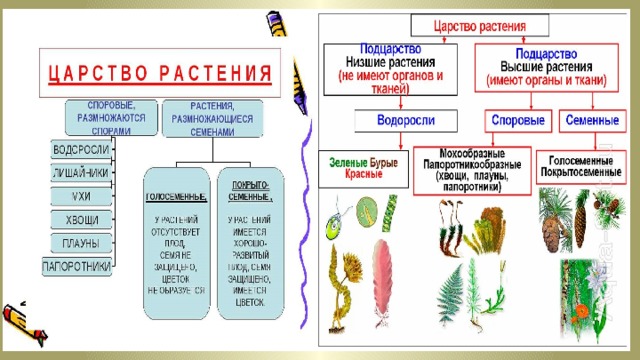 По мере развития спорофита внутри капсулы образуются гаплоидные споры. В целом спорофиты печеночников мельче и проще по морфологии, чем спорофиты мхов. Другое отличие состоит в том, что щетинка печеночника удлиняется после созревания капсулы, тогда как щетинка мха удлиняется до созревания капсулы.
По мере развития спорофита внутри капсулы образуются гаплоидные споры. В целом спорофиты печеночников мельче и проще по морфологии, чем спорофиты мхов. Другое отличие состоит в том, что щетинка печеночника удлиняется после созревания капсулы, тогда как щетинка мха удлиняется до созревания капсулы.
Распространение спор
Печеночники имеют характерный способ распространения спор. Внутри капсулы спорофита споры прикрепляются к специализированным клеткам, известным как элатеры. Элатер имеет трубчатую форму и имеет одно или несколько утолщений клеточной стенки, спирально ориентированных вдоль длинной оси клетки. Эти спиральные утолщения гигроскопичны, поскольку легко впитывают воду.
При высыхании капсула печеночника раскрывается. Затем спиральные утолщения клеточных стенок элатера подсыхают, и элатер изменяет свою форму. Когда это происходит, элатер выпускает связанные споры, которые затем рассеиваются ветром .
Бесполое размножение
Как и мхи, многие виды печеночников размножаются, образуя почки. Геммы представляют собой небольшие круглые или сферические репродуктивные структуры, которые находятся внутри чашечек гемм. Чашечки почки формируются на вершине слоевища. Образование гемм является важной формой бесполое размножение у многих видов печеночников и мхов.
Геммы представляют собой небольшие круглые или сферические репродуктивные структуры, которые находятся внутри чашечек гемм. Чашечки почки формируются на вершине слоевища. Образование гемм является важной формой бесполое размножение у многих видов печеночников и мхов.
Эволюция
Есть лишь несколько окаменелостей печеночников и мхов и нет окаменелостей роголистников. Это связано с тем, что мягкая ткань этих мохообразных плохо окаменевает. Самая старая из известных окаменелостей печеночника относится к позднему девонскому периоду, около 350 миллионов лет назад. Большинство ботаников считают, что они возникли задолго до этого.
Некоторые ботаники предположили, что в мире существует более 10 000 видов печеночников. Более реалистичная оценка составляет около 6000 видов. В прошлом количество видов могло быть переоценено, потому что морфология многих видов пластична, поскольку она различается в разных средах. Это делает идентификацию
видов печеночников очень сложно, как правило, сложнее, чем у высших растений.
Интересно, что хотя печеночники возникли за несколько сотен миллионов лет до цветковых растений, существует несколько сотен тысяч видов цветковых растений и только около 6000 видов печеночников. Причина этого может заключаться в том, что печеночники полагаются на неэффективный механизм переноса спермы водой для полового размножения . Таким образом, было высказано предположение, что большинство видов печеночников используют бесполые геммы как средство размножения. Бесполое размножение снижает генетическое разнообразие. Поскольку для развития новых видов необходимо генетическое разнообразие, печеночники и другие мохообразные, возможно, зашли в своего рода эволюционный тупик.
См. также Мохообразные.
Ресурсы
книги
Гринуэй, Т. Мхи и печеночники. Орландо: Рейнтри Стек-Вон, 1992.
Маргулис, Л. и К.В. Шварц. Пять Королевств. Вт.ч. Free-man and Company, 1998.
Питер А. Энсмингер
КЛЮЧЕВЫЕ ТЕРМИНЫ
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
- Диплоидный
— Ядро или клетка, содержащие две копии каждой хромосомы, образованные путем слияния двух гаплоидных ядер.
- Elater
— Специализированные трубчатые клетки со спирально ориентированными утолщениями клеточной стенки, к которым прикрепляются споры печеночника.
- Гаметофит
— Гаплоидное поколение, образующее гаметы, в жизненном цикле растения.
- Гемма
— Многоклеточная бесполая репродуктивная структура мхов и печеночников.
- Гаплоид
— Ядро или клетка, содержащая по одной копии каждой хромосомы.
- Мейоз
— Деление клеточного ядра, при котором число хромосом уменьшается вдвое, обычно от диплоидного до гаплоидного.
- Спорофит
— Диплоидное спорообразующее поколение в жизненном цикле растения.

- Слоевище
—Одиночное тело растения без отчетливых стебля, листьев и корней.
Энциклопедия науки Гейла
oxford
просмотра обновлено 18 мая 2018
liv·er·wort / ˈlivərˌwərt; -ˌwôrt/ •
н. небольшое зеленое растение без цветов (класс Hepaticae) с листовидными стеблями или лопастными листьями, встречающееся во влажных местообитаниях. Печеночники не имеют настоящих корней и размножаются спорами, высвобождающимися из коробочек.
Оксфордский карманный словарь современного английского языка
oxford
просмотров обновлено 14 мая 2018 г.0007 Любой из c. 9000 видов крошечных нецветущих зеленых растений, у которых, как и у родственных им мхов, отсутствуют специализированные ткани для транспортировки воды, пищи и минералов в теле растения. Печеночники относятся к типу растений Bryophyta. См. также мох
См. также мох
Всемирная энциклопедия
oxford
просмотров обновлено 11 мая 2018 г.
Словарь наук о растениях МАЙКЛ АЛЛАБИ
Роль арабидопсиса в исследованиях растений
Роль арабидопсиса в исследованиях растений
Растения жизненно важны для нашего существования. Они обеспечивают кислород, которым мы дышим, пищу, которую мы едим, волокна для нашей одежды, материалы для строительства наших домов и сырье для нашей промышленности. Четверть наших лекарственных препаратов производится из видов растений.
бумага, на которой напечатан этот отчет, является растительным продуктом.
Тем не менее, несмотря на важный вклад растений в наш уровень жизни, о них известно гораздо меньше, чем о мышах, мухах или бактериях, населяющих наш кишечник. Нам нужно больше узнать о том, как растут и развиваются растения; как они производят
полезные химические вещества; как они защищаются от вредителей; и как они чувствуют, реагируют и даже изменяют нашу среду. Один из способов узнать об этом — изучить гены растения. Информация, которую растения используют для роста и развития, а также для
Один из способов узнать об этом — изучить гены растения. Информация, которую растения используют для роста и развития, а также для
взаимодействуют с окружающей их средой, закодировано в их геномах. Чтобы полностью понять растения, нам нужно прочитать и интерпретировать их геномную информацию.
В 1980-х годах росло осознание того, что значительные инвестиции в исследования многих различных растений, таких как кукуруза, масличный рапс и соя, ослабляют усилия, направленные на полное понимание основных свойств всех растений. Ученые начали
понимать, что цель полного понимания физиологии и развития растений настолько амбициозна, что ее лучше всего достичь, обратившись к модельным видам растений, которые затем изучают многие ученые. К счастью, потому что все цветковые растения
близкородственными, полное секвенирование всех генов одного репрезентативного вида растений даст много знаний обо всех высших растениях. Точно так же открытие функций белков, продуцируемых модельными видами, может многое предложить.
сведения о роли белков во всех высших растениях.
За последние 8–10 лет растение Arabidopsis thaliana стало общепризнанным модельным растением для таких исследований. Несмотря на то, что это некоммерческий член семейства горчичных, ученые предпочитают его, потому что он развивается,
размножается и реагирует на стресс и болезни почти так же, как и многие сельскохозяйственные растения. Более того, Arabidopsis легко и недорого выращивать, и он дает много семян; это позволяет проводить обширные генетические эксперименты, часто с участием десятков
тысячи растений. Кроме того, Arabidopsis имеет сравнительно небольшой геном, что упрощает и облегчает генетический анализ. По сравнению с другими растениями у него отсутствуют повторяющиеся, менее информативные последовательности ДНК, которые усложняют анализ генома.
Первоначально было много споров о том, поможет ли лучшее понимание Arabidopsis в селекции коммерческих культур, и много споров по поводу решений о выделении ограниченных ресурсов на этот некоммерческий вид. Однако,
Однако,
многие достижения, о которых сообщалось за последние несколько лет, ясно свидетельствуют о том, что это растение является не только очень важным модельным видом для фундаментальных исследований, но и чрезвычайно ценным для прикладных ученых и селекционеров растений. Публикации по
Arabidopsis в высококачественных журналах растет в геометрической прогрессии после значительного увеличения инвестиций многих правительств. В Соединенных Штатах, например, Министерство сельского хозяйства США, Министерство энергетики, Национальный
Институты здравоохранения и Национальный научный фонд совместно предоставили 7,5 миллионов долларов США в 1990 году на исследования Arabidopsis и 22 миллиона долларов США в 1993 году. Европейское сообщество инвестировало значительную часть своей биотехнологии.
исследовательские ресурсы до Исследование генома арабидопсиса за последние 5 лет. На самом деле, многие из ведущих мировых лабораторий в области растениеводства инициировали программы с использованием Arabidopsis , и многие молодые ученые-растения решили начать
свою карьеру с использованием этого вида.