
Содержание
Выход растений на сушу [Первые наземные] — процесс, причины, почему, как, последствия, вики — WikiWhat
Основные статьи: Выход растений и животных на сушу, Палеозой, Растения, Высшие растения
Содержание (план)
1. Водоросли и лишайники
2. Сосудистые растения
3. Леса-водоемы
4. Голосеменные растения
Водоросли и лишайники
Кто были первые наземные существа — точно неизвестно: сами они в палеонтологическую летопись не попали, и мы судим об их существовании по косвенным признакам. Почва образуется из горных пород лишь при участии живых организмов. С другой стороны, в воде процесс почвообразования идти не может — только на суше. Значит, если в некое время существовали почвы, то должна была существовать и наземная жизнь… Почва может захораниваться, и попадать в геологическую летопись точно так же, как животные и растения. Так вот, самые древние из этих ископаемых почв имеют возраст 2 миллиарда лет. В почвообразовании в те времена участвовали низшие растения: наземные водоросли и лишайники. Похожие водоросли и сейчас встречаются на суше в виде зеленоватого налета на влажных камнях и коре деревьев. Лишайники же — союз, образованный водорослями и грибами, — способны жить в самых суровых условиях.
В почвообразовании в те времена участвовали низшие растения: наземные водоросли и лишайники. Похожие водоросли и сейчас встречаются на суше в виде зеленоватого налета на влажных камнях и коре деревьев. Лишайники же — союз, образованный водорослями и грибами, — способны жить в самых суровых условиях.
Сосудистые растения
см. Появление сосудистых растений
В конце раннего палеозоя появились и первые по-настоящему наземные (высшие) растения. От них сохранились только споры, очень похожие на споры современных печёночников.
Печёночники — это близкие родственники мхов. В средней полосе, эти зеленые лепешки размером примерно с ладонь частенько встречаются на гарях после лесных пожаров. Ботаники полагают, что склонность этих растений селиться на голом грунте, на камнях и т.п. они унаследовали от тех времен, когда были одними из первопоселенцев на суше.
Леса-водоемы
Травы во времена позднего палеозоя не было вовсе — ей негде было бы расти. Из-за воды не было видно земли: деревья стояли «по колено», а то и «по пояс» в воде. Собственно говоря, эти леса представляли собой мелководные водоемы, заполненные упавшими стволами и валежником, сквозь слой которого и прорастали живые деревья. Вся жизнь была сосредоточена именно в этих обводненных низменностях: на возвышенных участках продолжали существовать первичные пустыни. Из-за недостатка закрепляющей почву растительности в окрестных горах размыв грунта шел очень интенсивно, и после дождей оттуда часто сходили грязевые потоки сели, хоронившие заживо все население этих лесов. Материал с сайта http://wikiwhat.ru
Из-за воды не было видно земли: деревья стояли «по колено», а то и «по пояс» в воде. Собственно говоря, эти леса представляли собой мелководные водоемы, заполненные упавшими стволами и валежником, сквозь слой которого и прорастали живые деревья. Вся жизнь была сосредоточена именно в этих обводненных низменностях: на возвышенных участках продолжали существовать первичные пустыни. Из-за недостатка закрепляющей почву растительности в окрестных горах размыв грунта шел очень интенсивно, и после дождей оттуда часто сходили грязевые потоки сели, хоронившие заживо все население этих лесов. Материал с сайта http://wikiwhat.ru
Это было нечто похожее на болото современного типа, но без торфяного слоя. То есть совершенно особенное, такого на Земле никогда больше не существовало; эти экосистемы иногда называют «лесами-водоемами». Упавшие деревья здесь не разрушались древогрызущими насекомыми, грибками и бактериями, а слеживались под водою без доступа воздуха в плотную массу и постепенно превращались в каменный уголь. Большая часть мировых запасов каменного угля образовалась именно в это время; отсюда и название этого периода.
Большая часть мировых запасов каменного угля образовалась именно в это время; отсюда и название этого периода.
Загрузка…
Голосеменные растения
«Изобретение» семени — заслуга голосеменных растений. Среди них хорошо знакомы современные хвойные — ель и сосна. Так вот, семя — это окруженный защитной оболочкой зародыш растения с запасом «продуктов» на первое время. Семени не страшны ни высыхание, ни ультрафиолет. Дождавшись благоприятных условий, зародыш прорастает сквозь оболочку. Такое «изобретение» позволило голосеменным в конце палеозоя уйти от берегов водоемов и заселить огромные пространства материков от океана до океана.
Категории:
Растения
Палеозой
Вопросы к этой статье:
Как изменялась растительность в середине палеозоя?
Где, из чего, как и почему образовались залежи каменного угля?
Материал с сайта http://WikiWhat. ru
ru
Когда земная жизнь вышла на сушу
Первые «сухопутные» микробы могли появиться на Земле не позднее 3,2 млрд лет назад.
На древней Земле жизнь выбиралась из моря на вулканические острова. (Фото: Dan Durda / Southwest Research Institute.)
Образец осадочной породы с «сухопутным» бактериальным матом (обозначен стрелкой). (Фото: Martin Homann, European Institute for Marine Studies)
‹
›
Открыть в полном размере
Как мы все прекрасно знаем, жизнь зародилась в океане, и, проведя в нем какое-то время, эта жизнь вышла на сушу. Но в каком виде и когда именно первые живые организмы решились выбраться из воды? Обычно, когда мы говорим про освоение суши, то в голову приходят членистоногие, растения, рыбы – однако очевидно, что первыми живыми организмами на земной суше были какие-то микробы.
Хотя у микробов нет скелета, от их колоний все же остаются следы, сохраняющиеся в осадочных породах – именно такие следы ищут исследователи, занимающиеся происхождением жизни. Пока что наиболее надежными находками здесь остаются образцы возрастом 3,5 млрд лет, найденные в Западной Австралии еще в 1992 году – они считаются древнейшими следами жизни на Земле. (Стоит заметить, что время от времени появляются работы, в которых описываются еще более ранние ископаемые признаки жизни.)
Пока что наиболее надежными находками здесь остаются образцы возрастом 3,5 млрд лет, найденные в Западной Австралии еще в 1992 году – они считаются древнейшими следами жизни на Земле. (Стоит заметить, что время от времени появляются работы, в которых описываются еще более ранние ископаемые признаки жизни.)
Но следы эти в большинстве своем указывают на морскую жизнь. С земной жизнью все обстоит сложнее: во-первых, поверхность планеты в то время вообще была большей частью покрыта океанами, среди которых то тут, то там возникали вулканические острова; во-вторых, в море у следов жизни было больше шансов сохраниться в осадочных породах неповрежденными, нежели на суше; в-третьих, для «сухопутных» образцов такой древности сложнее определить возраст и сложнее описать то окружение, в котором они существовали, и потому их легко спутать с теми, которые находились в море.
До сих пор самыми древними наземными следами жизни считались образцы возрастом 2,7 млрд лет, открытые в формации Тумбиана в Австралии и в зеленокаменном поясе Барбертон в Южной Африке (зеленокаменными поясами называют особый комплекс пород возрастом 2,5–3,5 млрд лет). Однако в новой статье, опубликованной в Nature Geosciences, исследователи из Европейского института моря вместе с коллегами из других научных центров Франции, Германии и Австралии пишут, что им удалось найти еще более древние признаки «сухопутной» жизни – возрастом 3,2 млрд лет.
Однако в новой статье, опубликованной в Nature Geosciences, исследователи из Европейского института моря вместе с коллегами из других научных центров Франции, Германии и Австралии пишут, что им удалось найти еще более древние признаки «сухопутной» жизни – возрастом 3,2 млрд лет.
Авторы работы изучали древние осадочные породы из того же зеленокаменного пояса Барбертон в Южной Африке, в которых обнаружились следы бактериальных матов – сложных симбиотических сообществ бактерий и архей, в которых микробы формируют перемежающиеся слои. Судя по окружению микробных матов (точнее, следов микробных матов), они возникли именно здесь, причем в этом месте когда-то была речная дельта.
В пользу того, что микробы жили не в море, а на суше, пусть и в речной дельте, говорит и содержание изотопов углерода и азота, свойственных органическим веществам. По изотопам «сухопутные» бактериальные маты отличаются от морских, потому что те и другие сформированы микробами с разным обменом веществ. Те бактерии, которые оставили следы возрастом 3,2 млрд лет, обитали явно не в море, так что, очевидно, жизнь вышла на сушу раньше, чем все думали прежде.
Те бактерии, которые оставили следы возрастом 3,2 млрд лет, обитали явно не в море, так что, очевидно, жизнь вышла на сушу раньше, чем все думали прежде.
По материалам The Scientist
Происхождение и ранняя эволюция растений на суше
Бэнкс, Х. П. Реклассификация Psilophyta. Таксон 24 , 401–413 (1975).
Google ученый
Chaloner, WG & Sheerin, A. in The Devonian System (eds House, MR, Scrutton, CT & Bassett, MG) 145–161 (The Palaeontological Association, London, (1979)).
Google ученый
Грей, Дж. Основные эволюционные биособытия наземных растений палеозоя. Палеогеог. Палеоклимат. Палеокол. 104 , 153–169 (1993).
Google ученый
Graham, LE Origin of Land Plants (Wiley, New York, (1993)).

Google ученый
Мишлер Б.Д. и др. . Филогенетические взаимоотношения «зеленых водорослей» и «мохов». Энн. МО Бот. Гард. 81 , 451–483 (1994).
Google ученый
Манхарт, Дж. Р. и Палмер, Дж. Г. Получение двух интронов тРНК хлоропластов отмечает предков зеленых водорослей наземных растений. Природа 345 , 268–270 (1990).
ОБЪЯВЛЕНИЕ
КАС
пабмедGoogle ученый
Манхарт, Дж. Р. Филогенетический анализ зеленых растений rbc L последовательностей. мол. Филогенет. Эвол. 3 , 114–127 (1994).
Google ученый
Raubeson, L.A. & Jansen, R.K. Свидетельства ДНК хлоропластов о древнем эволюционном расколе сосудистых наземных растений.
Наука 255 , 1697–1699 (1992).Google ученый
Чепмен, Р. Л. и Буххейм, М. А. Последовательности генов рибосомной РНК: анализ и значение в филогении и таксономии зеленых водорослей. Крит. Преподобный завод наук. 10 , 343–368 (1991).
Google ученый
McCourt, R.M., Karol, KG, Guerlesquin, M. & Feist, M. Филогения существующих родов семейства Characeae (Charales, Charophyceae) на основе последовательностей и морфологии rbc L. утра. Дж. Бот. 83 , 125–131 (1996).
Google ученый
Прайер, К. М., Смит, А. Р. и Ског, Дж. Э. Филогенетические отношения существующих папоротников, основанные на данных морфологии и последовательностях rbc L. утра. Ферн Дж. 85 , 205–282 (1995).
Google ученый
Кранц, Х. Д. и др. . Происхождение наземных растений: филогенетические отношения между харофитами, мохообразными и сосудистыми растениями, установленные на основе полных последовательностей генов малой субъединицы рибосомной РНК. Дж. Мол. Эвол. 41 , 74–84 (1995).
Google ученый
Kranz, H.D. & Huss, V.A.R. Молекулярная эволюция папоротников и их взаимосвязь с семенными растениями: данные полных последовательностей генов 18S рРНК. Завод Сист. Эвол. 202 , 1–11 (1996).
Google ученый
Hiesel, R., von Haeseler, A. & Brennicke, A. Последовательности митохондриальных нуклеиновых кислот растений как инструмент для филогенетического анализа. Проц. Натл акад. науч. США 91 , 634–638 (1994).
Google ученый
Эдвардс, Д., Дэвис, К.Л. и Акс, Л. Бессосудистая проводящая нить в раннем наземном растении Cooksonia . Природа 357 , 683–685 (1992).
ОБЪЯВЛЕНИЕ
Google ученый
Эдвардс, Д., Дакетт, Дж. Г. и Ричардсон, Дж. Б. Печеночные признаки у самых ранних наземных растений. Природа 374 , 635–636 (1995).
ОБЪЯВЛЕНИЕ
КАСGoogle ученый
Фаннинг, У., Эдвардс, Д. и Ричардсон, Дж. Б. Разнообразное собрание ранних наземных растений из нижнего девона валлийской границы. Бот. Дж. Линн. соц. 109 , 161–188 (1992).
Google ученый
Кенрик, П. Чередование поколений у наземных растений: новые филогенетические и морфологические данные.
биол. Ред. 69 , 293–330 (1994).Google ученый
Кенрик П. и Крейн П. Р. Водопроводящие клетки ранних ископаемых наземных растений: значение для ранней эволюции трахеофитов. Бот. Газ. 152 , 335–356 (1991).
Google ученый
Реми, В. Гензель, П.Г. и Хасс, Х. Поколение гаметофитов некоторых наземных растений раннего девона. Междунар. Дж. Растениевод. 154 , 35–58 (1993).
Google ученый
Remy, W. & Hass, H. Новая информация о гаметофитах и спорофитах Aglaophyton major и выводы о возможной адаптации к окружающей среде. Преподобный Палеобот. Палинол. 90 , 175–194 (1996).
Google ученый
Реми В.
, Тейлор Т. Н., Хасс Х. и Керп Х. Везикулярная арбускулярная микориза возрастом четыреста миллионов лет. Проц. Нац. акад. науч. США 91 , 11841–11843 (1994).Google ученый
Stein, WE, Harmon, GD & Hueber, FM in International Workshop on the Biology and Evolutionary Impacts for Early Devon Plants (Westfälische Wilhelms-Universität Münster, Германия, (1994)).
Google ученый
Тейлор, Т. Н. и Осборн, Дж. М. Важность грибов в формировании палеоэкосистемы. Преподобный Палеобот. Палинол. 90 , 249–262 (1996).
Google ученый
Тейлор, В. А. Ультраструктура нижнепалеозойских диад из южного Огайо. Преподобный Палеобот. Палинол. 92 , 269–280 (1996).
Google ученый
Kenrick, P.
& Crane, P. R. Происхождение и ранняя диверсификация наземных растений: кладистическое исследование (Smithsonian Institution Press, Вашингтон, округ Колумбия, (1997)).Google ученый
Грей, Дж. Микрофоссилии ранних наземных растений: достижения в понимании ранней земной жизни, 1970–1984 Phil. Транс. Р. Соц. Лонд. B 309 , 167–195 (1985).
Google ученый
Грей, Дж. и Буко, А. Дж. Ранние сосудистые наземные растения: доказательства и предположения. Lethaia 10 , 145–174 (1977).
Google ученый
DiMichele, WA et al .in Земные экосистемы во времени: эволюционная палеоэкология наземных растений и животных (изд. Behrensmeyer, AK) 205–325 (Univ. Chicago Press, (1992)).
Google ученый
Фаннинг У.
, Ричардсон Дж. Б. и Эдвардс Д. в Pollen and Spores (ред. Blackmore, S. & Barnes, SH) 25–47 (Clarendon, Oxford, (1991)).Google ученый
Крокен, С. Б., Грэм, Л. Э. и Кук, М. Э. Возникновение и эволюционное значение устойчивых клеточных стенок у харофитов и мохообразных. утра. Дж. Бот. 83 , 1241–1254 (1996).
Google ученый
Веллман, С. Х. и Ричардсон, Дж. Б. Комплексы спороморфов из «Нижнего старого красного песчаника» в Лорне, Шотландия. Специальные документы Палеонтолог. 55 , 41–101 (1996).
Google ученый
Эдвардс, Д. в Palaeozoic Palaeogeography and Biogeography (ред. McKerrow, WS & Scotese, CR) 233–242 (Геологическое общество, Лондон, (1990)).
Google ученый
Морел, Э.
, Эдвардс, Д. и Иньикес Родригес, М. Первая запись Cooksonia из Южной Америки в силурийских породах Боливии. Геол. Маг. 132 , 449–452 (1995).Google ученый
Tims, JD & Chambers, TC Rhyniophytina и Trimerophytina из ранней наземной флоры Виктории, Австралия. Палеонтология 27 , 265–279 (1984).
Google ученый
Кай, К.-Ю., Доу, Ю.-В. и Эдвардс, Д. Новые наблюдения за комплексом растений Придоли из северного Синьцзяна, северо-запад Китая, с комментариями о его эволюционном и палеографическом значении. Геол. Маг. 130 , 155–170 (1993).
Google ученый
Хьюбер, Ф. М. Мысли о ранних ликопсидах и зостерофиллах. Энн. МО Бот. Гард. 79 , 474–499 (1992).
Google ученый
Кай, К.
и др. . Раннесилурийское сосудистое растение. Природа 379 , 592 ((1996)).ОБЪЯВЛЕНИЕ
КАСGoogle ученый
Гэн Б.-Ю. Анатомия и морфология Pinnatiramosus , нового растения из среднего силура (венлока) Китая. Акта Бот. Грех. 28 , 664–670 (1986).
Google ученый
Raymond, A. & Metz, C. Разнообразие наземных растений Лауруссии в силуре и девоне: массовое вымирание, систематическая ошибка выборки или и то, и другое? Палеобиология 21 , 74–91 (1995).
Google ученый
Эдвардс, Д. и Дэвис, М.С. в Основные эволюционные излучения (ред. Тейлор, П.Д. и Ларвуд, Г.П.) 351–376 (Кларендон, Оксфорд, (1990)).
Google ученый
Нолл, А.
Х., Никлас, К. Дж., Генсел, П. Г. и Тиффни, Б. Х. Диверсификация признаков и закономерности эволюции ранних сосудистых растений. Палеобиология 10 , 34–47 (1984).Google ученый
Gensel, PG & Andrews, HN Растительная жизнь в девоне (Praeger, New York, (1984)).
Google ученый
Тейлор, Т. Н. и Тейлор, Э. Л. Биология и эволюция ископаемых растений (Прентис Холл, Нью-Джерси, (1993)).
Google ученый
Швейцер, Х.-Дж. Die Unterdevonflora des Rheinlandes. Palaeontographica B 189 , 1–138 (1983).
Google ученый
Gerrienne, P. Inventaire des végétaux éodévoniens de Belgique. Энн. соц. Геол. бельг. 116 , 105–117 (1993).
Google ученый
Таппан, Х. Н. Палеобиология простейших растений (Фриман, Сан-Франциско, (1980)).
Google ученый
Raven, J. Реакция растений на высокие концентрации O2: отношение к предыдущим эпизодам с высоким содержанием O2. Палеогрег. Палеоклимат. Палеокол. 97 , 19–38 (1991).
Google ученый
Штейн, А. Э., Коэн, Дж. Д., Словин, Дж. П. и Кук, Т. Дж. Метаболизм ауксинов у репрезентативных наземных растений. 900:05 утра. Дж. Бот. 82 , 1514–1521 (1995).
Google ученый
Эдвардс, Д. Новый взгляд на ранние наземные экосистемы: взгляд на мир лилипутов. Преподобный Палеобот. Палинол. 90 , 159–174 (1996).
Google ученый
Эдвардс, Д.
, Фаннинг, У. и Ричардсон, Дж. Б. Устьица и стеромы ранних наземных растений. Природа 323 , 438–440 (1986).ОБЪЯВЛЕНИЕ
Google ученый
Рэйвен, Дж. А. Сравнительная физиология адаптации растений и членистоногих к земле. Фил. Транс. Р. Соц. Лонд. B 309 , 273–288 (1985).
Google ученый
Рэйвен, Дж. А. Эволюция сосудистых растений в связи с количественным функционированием мертвых водопроводящих клеток и устьиц. биол. 68 , 337–363 (1993).
Google ученый
Никлас, К. Дж. Аллометрия растений: масштабирование формы и процесса. (Университет Чикаго Пресс, (1994)).
Google ученый
Бирбауэр, Р. в Геологические факторы и эволюция растений (изд.
Тиффни, Б.Х.) 47–92 (Издательство Йельского университета, Нью-Хейвен, Коннектикут, (1985)).Google ученый
Бернер, Р. А. GEOCARB II: пересмотренная модель атмосферного CO2 за фанерозойское время. утра. J. Sci. 294 , 56–91 (1994).
Google ученый
Мора, К.И., Дризе, С.Г. и Коларуссо, Л.А. Уровни CO2 в атмосфере от среднего до позднего палеозоя по почвенным карбонатам и органическому веществу. Наука 271 , 1105–1107 (1996).
Google ученый
Алгео, Т. Дж., Бернер, Р., Мейнард, Дж. Б. и Шеклер, С. Э. Позднедевонские океанические бескислородные события и биотические кризисы: «укорененные» в эволюции сосудистых наземных растений? GSA Today 5 , 45, 64–66 (1995).
Google ученый
Retallack, GJ in Paleosols: its Recognition and Interpretation (ed.
Wright, VP) (Blackwell, Oxford, (19)86)).Google ученый
Нолл, А. Х. Ранняя эволюция эукариот: геологическая перспектива. Наука 256 , 622–627 (1992).
Google ученый
Бенгтсон С. (редактор) Ранняя жизнь на Земле. (Columbia Univ. Press, Нью-Йорк, (1994)).
Google ученый
Тейлор, Т. Н., Хасс, Х., Реми, В. и Керп, Х. Самый старый ископаемый лишайник. Природа 378 , 244 (1995).
ОБЪЯВЛЕНИЕ
КАСGoogle ученый
Хемсли, А. Р. в Ультраструктура ископаемых спор и пыльцы (ред. Курманн, М. Х. и Дойл, Дж. А.) 1–21 (Королевские ботанические сады, Кью, (1994)).
Google ученый
Хьюбер, Ф.
М. в Международный семинар по биологии и эволюционным последствиям растений раннего девона (Westfälische Wilhelms-Universität, Münster, (1994)).Google ученый
Симон, Л., Буске, Дж., Левек, К. и Лалонд, М. Происхождение и диверсификация эндомикоризных грибов с сосудистыми растениями. Природа 363 , 67–69 (1993).
ОБЪЯВЛЕНИЕ
Google ученый
Селден, П.А. и Эдвардс, Д. в Evolution and the Fossil Record (редакторы Allen, KC & Briggs, DEG) 122–152 (Белхейвен, Лондон, (1989)).
Google ученый
Грей, Дж. и Шир, В. Ранняя жизнь на суше. утра. науч. 80 , 444–456 (1992)).
Google ученый
Грей, Дж. и Буко, А. Дж. Останки неморских животных раннего силура и природа ранней континентальной экосистемы.
Акта Палеонтол. пол. 38 , 303–328 (1994).Google ученый
Retallack, G.J. & Feakes, C.R. Следы окаменелостей позднеордовикских животных на суше. Наука 235 , 61–63 (1987).
Google ученый
Скотт А.С., Стивенсон Дж. и Чалонер В.Г. Взаимодействие и совместная эволюция растений и членистоногих в палеозое и мезозое. Фил. Транс. Р. Соц. Лонд. B 336 , 129–165 (1992).
Google ученый
Бэнкс, Х. П. и Колтхарт, Б. Дж. Взаимодействие растений, животных и грибов у тримерофитов раннего девона из Гаспе, Канада. утра. Дж. Бот. 80 , 992–1001 (1993).
Google ученый
Эдвардс, Д., Селдон, П. А., Ричардсон, Дж. Б. и Акс, Л.
Копролиты как свидетельство взаимодействия растений и животных в силуро-девонских наземных экосистемах. Природа 377 , 329–331 (1995).ОБЪЯВЛЕНИЕ
КАСGoogle ученый
Аллен, Дж. Р. Л. Морская вода к пресной: седиментология прерванного экологического перехода (лудлоу-зигенский период) в англо-валлийском регионе. Фил. Транс. Р. Соц. Лонд. B 309 , 85–104 (1985).
Google ученый
Мелконян М. и Сурек Б. Филогения Chlorophyta: соответствие между ультраструктурными и молекулярными данными. Бык. соц. Зоол. о. 120 , 191–208 (1995).
Google ученый
Бремер, К., Хамфрис, С.Дж., Мишлер, Б.Д. и Черчилль, С.П. О кладистических отношениях в зеленых растениях. Таксон 36 , 339–349 (1987).
Google ученый
Гарбари, Д. Дж., Ренцалья, К. С. и Дакетт, Дж. Г. Филогения наземных растений: кладистический анализ, основанный на мужском гаметогенезе. Завод Сист. Эвол. 188 , 237–269 (1993).
Google ученый
Capesius, I. Амолекулярная филогения мохообразных на основе кодируемых в ядре генов 18S рРНК. J. Физиол растений. 146 , 59–63 (1995).
Google ученый
Тейлор Т. Н. Происхождение наземных растений: ответы, вопросы. Таксон 37 , 805–833 (1988).
Google ученый
Ротвелл, Г. В. в Птеридиология в перспективе (ред. Камю, Дж. М., Гибби, М. и Джонс, Р. Дж.) (Королевские ботанические сады, Кью) (в печати).
Альберт, В.
А. и др. . Функциональные ограничения и доказательства филогении наземных растений. Энн. МО Бот. Гард. 81 , 534–567 (1994).Google ученый
Эдвардс, Д., Фаннинг, У. и Ричардсон, Дж. Б. Угольные спорангии нижнего девона из Шропшира: Salopella Edwards & Richardson и > Tortilicaulis Edwards. Бот. Дж. Линн. соц. 116 , 89–110 (1994).
Google ученый
Бейтман, Р. М., ДиМишель, В. А. и Уиллард, Д. А. Экспериментальный кладистический анализ анатомически сохранившихся плауновидных из каменноугольного периода Еврамерики: эссе по палеоботанической филогенетике. Энн. МО Бот. Гард. 79 , 500–559 (1992).
Google ученый
Feist, M. & Grambast-Fesssard, N. в Calcareous Algae and Stromatolites (изд.
Riding, R.) 189–203 (Springer, Berlin, (1991)).Google ученый
Hébant, C. in Bryophyte Systematics (ред. Clarke, GCS & Duckett, JG) 365–383 (Академический, Лондон, (19)79)).
Google ученый

 Наука 255 , 1697–1699 (1992).
Наука 255 , 1697–1699 (1992).

 биол. Ред. 69 , 293–330 (1994).
биол. Ред. 69 , 293–330 (1994). , Тейлор Т. Н., Хасс Х. и Керп Х. Везикулярная арбускулярная микориза возрастом четыреста миллионов лет. Проц. Нац. акад. науч. США 91 , 11841–11843 (1994).
, Тейлор Т. Н., Хасс Х. и Керп Х. Везикулярная арбускулярная микориза возрастом четыреста миллионов лет. Проц. Нац. акад. науч. США 91 , 11841–11843 (1994). & Crane, P. R. Происхождение и ранняя диверсификация наземных растений: кладистическое исследование (Smithsonian Institution Press, Вашингтон, округ Колумбия, (1997)).
& Crane, P. R. Происхождение и ранняя диверсификация наземных растений: кладистическое исследование (Smithsonian Institution Press, Вашингтон, округ Колумбия, (1997)). , Ричардсон Дж. Б. и Эдвардс Д. в Pollen and Spores (ред. Blackmore, S. & Barnes, SH) 25–47 (Clarendon, Oxford, (1991)).
, Ричардсон Дж. Б. и Эдвардс Д. в Pollen and Spores (ред. Blackmore, S. & Barnes, SH) 25–47 (Clarendon, Oxford, (1991)). , Эдвардс, Д. и Иньикес Родригес, М. Первая запись Cooksonia из Южной Америки в силурийских породах Боливии. Геол. Маг. 132 , 449–452 (1995).
, Эдвардс, Д. и Иньикес Родригес, М. Первая запись Cooksonia из Южной Америки в силурийских породах Боливии. Геол. Маг. 132 , 449–452 (1995). и др. . Раннесилурийское сосудистое растение. Природа 379 , 592 ((1996)).
и др. . Раннесилурийское сосудистое растение. Природа 379 , 592 ((1996)). Х., Никлас, К. Дж., Генсел, П. Г. и Тиффни, Б. Х. Диверсификация признаков и закономерности эволюции ранних сосудистых растений. Палеобиология 10 , 34–47 (1984).
Х., Никлас, К. Дж., Генсел, П. Г. и Тиффни, Б. Х. Диверсификация признаков и закономерности эволюции ранних сосудистых растений. Палеобиология 10 , 34–47 (1984).
 , Фаннинг, У. и Ричардсон, Дж. Б. Устьица и стеромы ранних наземных растений. Природа 323 , 438–440 (1986).
, Фаннинг, У. и Ричардсон, Дж. Б. Устьица и стеромы ранних наземных растений. Природа 323 , 438–440 (1986). Тиффни, Б.Х.) 47–92 (Издательство Йельского университета, Нью-Хейвен, Коннектикут, (1985)).
Тиффни, Б.Х.) 47–92 (Издательство Йельского университета, Нью-Хейвен, Коннектикут, (1985)). Wright, VP) (Blackwell, Oxford, (19)86)).
Wright, VP) (Blackwell, Oxford, (19)86)). М. в Международный семинар по биологии и эволюционным последствиям растений раннего девона (Westfälische Wilhelms-Universität, Münster, (1994)).
М. в Международный семинар по биологии и эволюционным последствиям растений раннего девона (Westfälische Wilhelms-Universität, Münster, (1994)).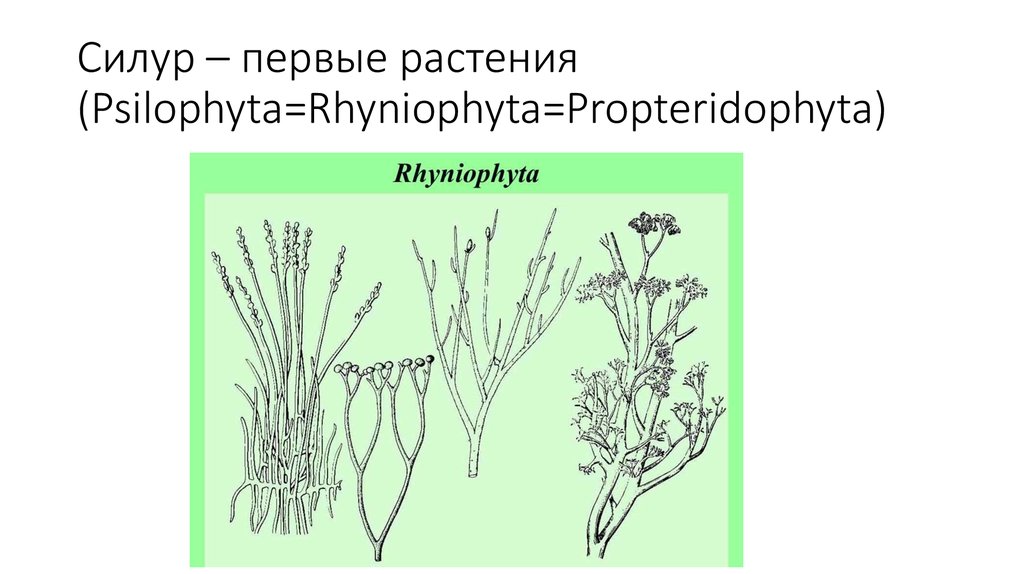 Акта Палеонтол. пол. 38 , 303–328 (1994).
Акта Палеонтол. пол. 38 , 303–328 (1994). Копролиты как свидетельство взаимодействия растений и животных в силуро-девонских наземных экосистемах. Природа 377 , 329–331 (1995).
Копролиты как свидетельство взаимодействия растений и животных в силуро-девонских наземных экосистемах. Природа 377 , 329–331 (1995).
 А. и др. . Функциональные ограничения и доказательства филогении наземных растений. Энн. МО Бот. Гард. 81 , 534–567 (1994).
А. и др. . Функциональные ограничения и доказательства филогении наземных растений. Энн. МО Бот. Гард. 81 , 534–567 (1994). Riding, R.) 189–203 (Springer, Berlin, (1991)).
Riding, R.) 189–203 (Springer, Berlin, (1991)).Ссылки для скачивания
6: Ранние наземные растения — Биология LibreTexts
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 27713
- Мелисса Ха, Мария Морроу и Камми Алжирс
- Колледж Юба, Колледж Редвудс и Колледж Вентура через Инициативу открытых образовательных ресурсов ASCCC
Существует четыре основные эволюционные группы наземных растений: мохообразные, бессемянные сосудистые растения (СВП), голосеменные и покрытосеменные. Эти группы представляют собой основные изменения в структуре растений и характеристиках жизненного цикла с течением времени, которые совпадают с крупными изменениями в эволюции Земли в целом. Ранняя Земля выглядела бы совсем иначе, чем планета, которую вы знаете. Когда растения впервые вышли из воды, почвы не было. Земной ландшафт был бы скалистым, потенциально скользким от микробной слизи. Отсутствие окружающей воды будет означать, что ткани могут высохнуть, а также увеличить воздействие кислорода и разрушительных длин волн солнечного света. Им также потребуется больше структурной поддержки, без воды, чтобы плавать, а изменения температуры будут гораздо более резкими и быстрыми, чем в их прежней водной среде обитания. Тем не менее, у них было бы много CO 2 и повышенный доступ солнечного света для фотосинтеза.
Эти группы представляют собой основные изменения в структуре растений и характеристиках жизненного цикла с течением времени, которые совпадают с крупными изменениями в эволюции Земли в целом. Ранняя Земля выглядела бы совсем иначе, чем планета, которую вы знаете. Когда растения впервые вышли из воды, почвы не было. Земной ландшафт был бы скалистым, потенциально скользким от микробной слизи. Отсутствие окружающей воды будет означать, что ткани могут высохнуть, а также увеличить воздействие кислорода и разрушительных длин волн солнечного света. Им также потребуется больше структурной поддержки, без воды, чтобы плавать, а изменения температуры будут гораздо более резкими и быстрыми, чем в их прежней водной среде обитания. Тем не менее, у них было бы много CO 2 и повышенный доступ солнечного света для фотосинтеза.
- 6.1: Знакомство с ранними наземными растениями
- Мохообразные были первой группой растений, эволюционировавших на суше, за ними последовали бессемянные сосудистые растения. Эти ранние растения в сопровождении своих грибковых мутуалистов и других микробов превратили каменистый земной ландшафт в экосистему со слоистыми почвами и сложными биотическими сообществами. Синапоморфии мохообразных связаны с проблемами жизни на суше, тогда как синапоморфии бессемянных сосудистых растений связаны с увеличением высоты и возможностями для мейоза (т. Е. Конкуренцией).
- 6.2: Мохообразные
- Существует около 23 000 видов мохообразных, относящихся к трем различным линиям: Anthocerotophyta, Marchantiophyta и Bryophyta. Эти ранние растения, лишенные сосудистой ткани, обычно имеют распростертую форму и растут вплотную к субстрату. У них нет настоящих корней, но есть якорные клетки, называемые ризоидами, которые отходят от гаметофита. Мохообразные имеют жизненный цикл с преобладанием гаметофитов, а спорофиты растут из мегагаметофитов.
- 6.2.1: Anthocerotophyta
- 6.2.2: Marchantiophyta
- 6. 2.3: Bryophyta
- 6.3.3: 3.SINSIRLESRELESRELE 9083.SINLERSELESRELENLER. воду по клеткам древесной ксилемы вверх от настоящих корней, по стеблям, вплоть до их листьев. Фотосинтезирующие ткани могут распределять сахара через живые клетки флоэмы по всему растению. SVP являются доминирующими спорофитами с редуцированными талоидными гаметофитами. Спорофиты разветвлены со множеством участков для образования спор. Их можно разделить на две линии: Lycopodiopsida и Polypodiopsida.
- 6.3.1: Lycopodiopsida
- 6.3.2: Polypodiopsida
- 6.4: Краткое изложение глав
 Эти ранние растения в сопровождении своих грибковых мутуалистов и других микробов превратили каменистый земной ландшафт в экосистему со слоистыми почвами и сложными биотическими сообществами. Синапоморфии мохообразных связаны с проблемами жизни на суше, тогда как синапоморфии бессемянных сосудистых растений связаны с увеличением высоты и возможностями для мейоза (т. Е. Конкуренцией).
Эти ранние растения в сопровождении своих грибковых мутуалистов и других микробов превратили каменистый земной ландшафт в экосистему со слоистыми почвами и сложными биотическими сообществами. Синапоморфии мохообразных связаны с проблемами жизни на суше, тогда как синапоморфии бессемянных сосудистых растений связаны с увеличением высоты и возможностями для мейоза (т. Е. Конкуренцией).Мария Морроу (CC-BY-NC)
Эта страница под названием 6: Ранние наземные растения распространяется под лицензией CC BY-NC 4.0, авторами, ремиксами и/или кураторами являются Мелисса Ха, Мария Морроу и Камми Алжирс (Инициатива открытых образовательных ресурсов ASCCC).