Содержание
Оплодотворение у растений и образование зиготы. Двойное оплодотворение
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
1. План самостоятельной работы учащегося по биологии 7 класс. Раздел: Размножение Тема: Понятие об оплодотворении у растений и
образование зиготы. Двойное оплодотворение.
Биологическое значение двойного
оплодотворения.
Дата:22.04.2021г.
Цель:7.2.1.4 — описывать значение двойного
оплодотворения цветковых растений
Здравствуйте ребята сегодня мы
поговорим о размножении
растений.
Размножение – воспроизведение
себе подобных, увеличение числа
особей определенного вида,
необходимое свойство жизни,
присущее всем живым
организмам.
размножение
бесполое
Вегетативное
половое
Спорообразование
яйцеклетка
При половом размножении
развивается новый организм со
свойствами отличительными от
родительских, хотя внешне очень
похожих на родителей.
При бесполом размножении образуются новые
(дочерние) растения схожие по своим свойствам с
материнским растением.
сперматозоиды
4. Строение цветка покрытосеменных растений
Пыльца растений
5. Опыление — перенос пыльцы с тычинки на рыльце пестика
6. Опыление
самоопыление
насекомыми
животными
птицами
ветром
-Где образуются половые клетки? Как они
называются?
— Каким образом при половом размножении
происходит слияние мужских и женских
половых клеток, если они располагаются на
разных растениях?
— Как называется процесс слияния мужских и
женских половых клеток?
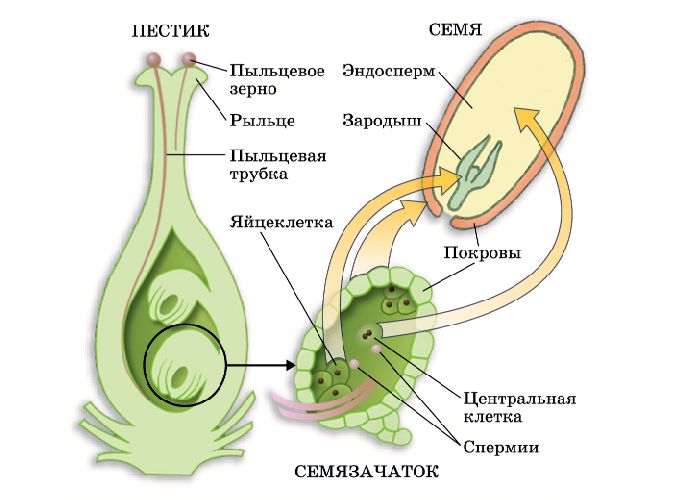
Оплодотворение – процесс
слияния женских и мужских
половых клеток.
♀ яйцеклетка х
♂сперматозоид →
зигота → зародыш.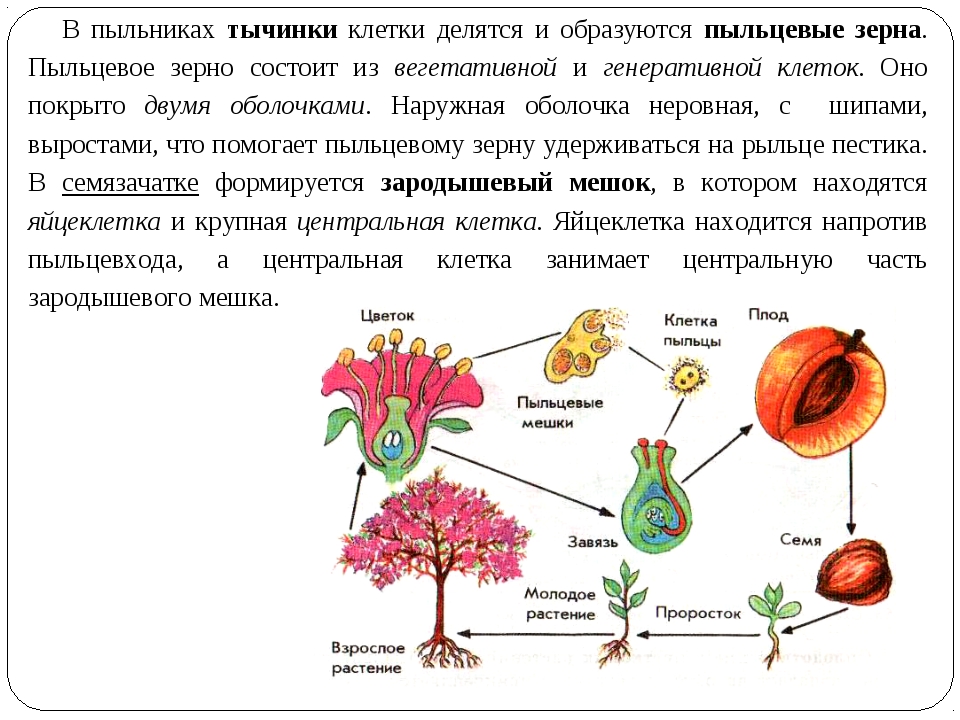
Зигота – оплодотворенная яйцеклетка.
♀ клетки — яйцеклетки
♂ клетки — спермии
образуются
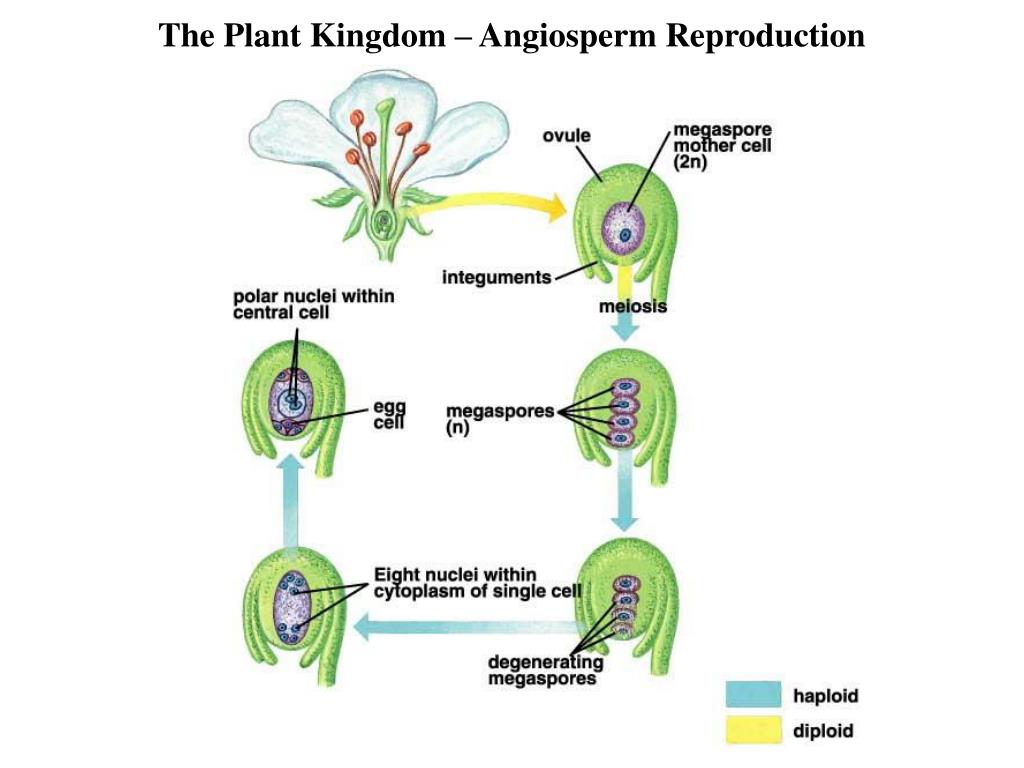
В семязачатках завязи пестика
В пыльце пыльников тычинок
Опыление
Зародышевый мешок
Яйцеклетка
Зигота
Зародыш семени
растения
Проросток
Пыльцевая трубка с двумя
спермиями
Центральная клетка
с ядром
Зигота
Эндосперм
Этот процесс
называется
двойным
оплодотворением.
Он был открыт
русским ученым
ботаником
Сергеем
Гавриловичем
Навашиным в 1898
году.
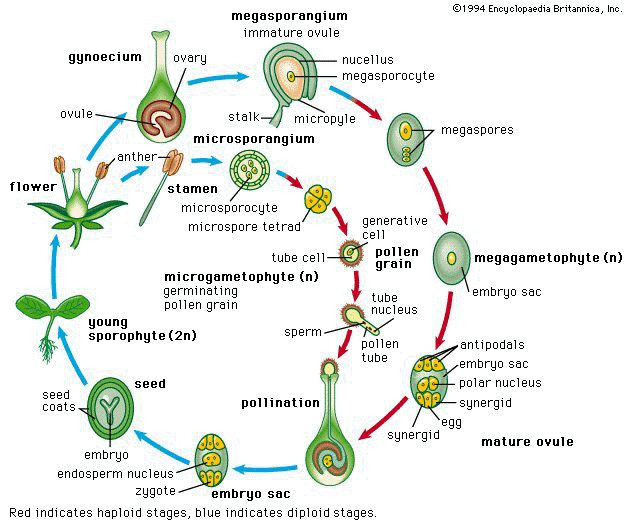
Этапы полового размножения
1.Образование половых клеток (гамет)
2. Опыление
3. Оплодотворение
4. Образование зиготы
5. Развитие дочернего организма.
13. В результате оплодотворения образуются плоды и семена
14. Учебное задание: Откройте учебник стр.225. Выполните задание 1 и 2.
English
Русский
Правила
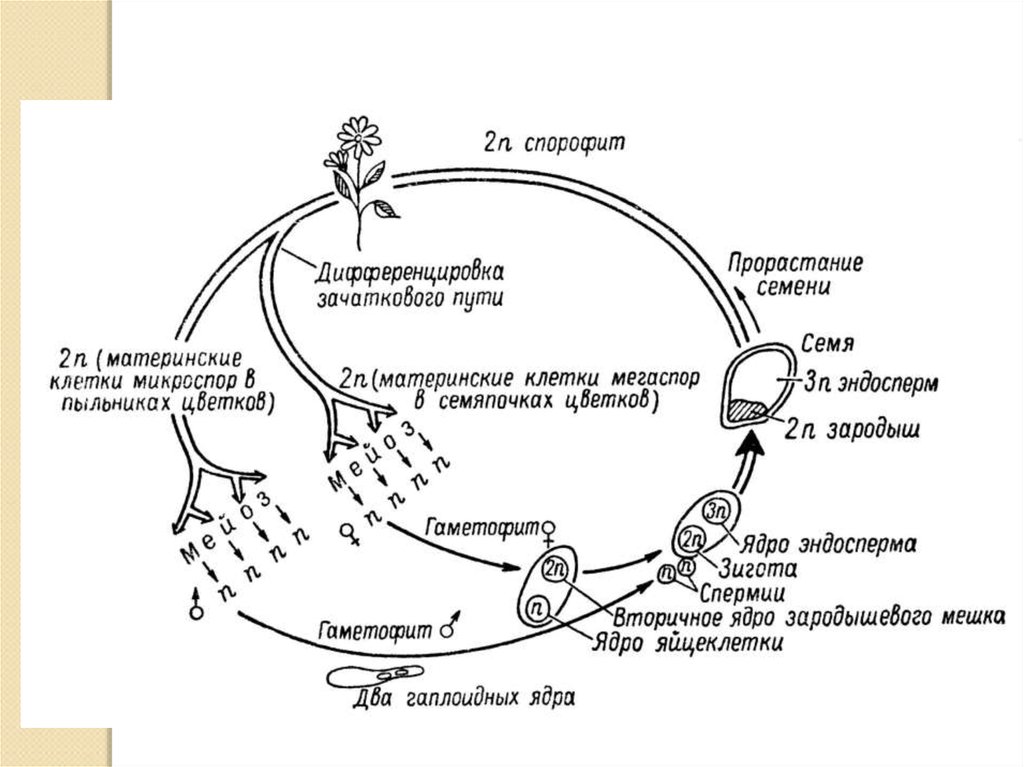
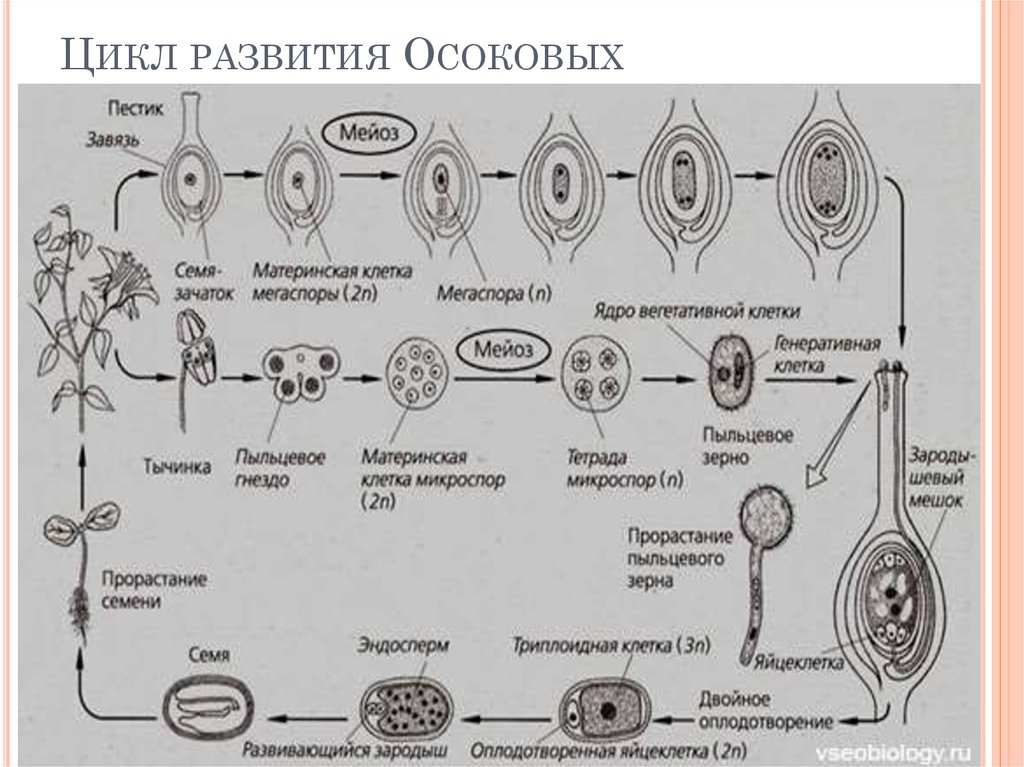
Оплодотворение двойное
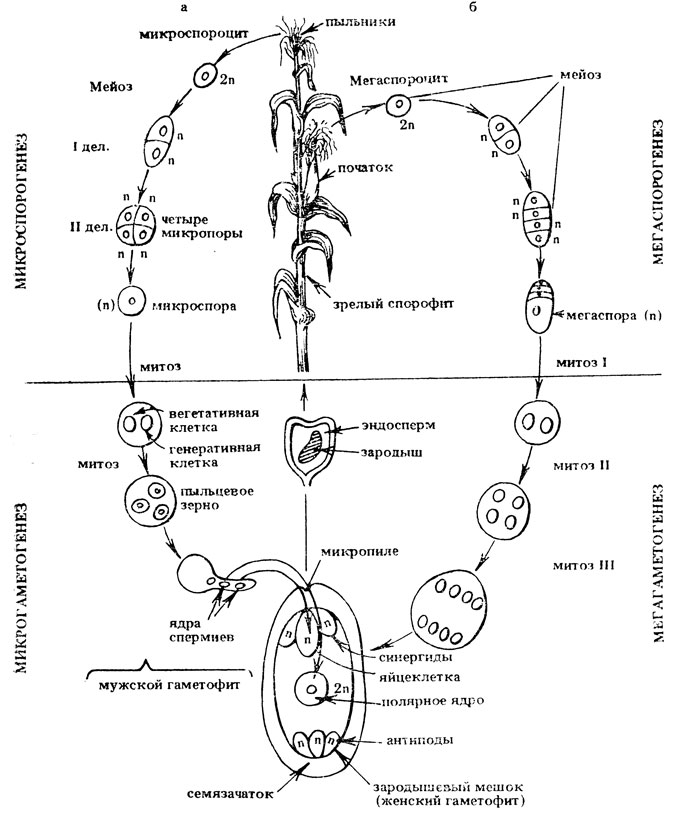
Пыльцевые зерна, попав на рыльце пестика, поглощают воду, сахара и другие питательные вещества и образуют трубку.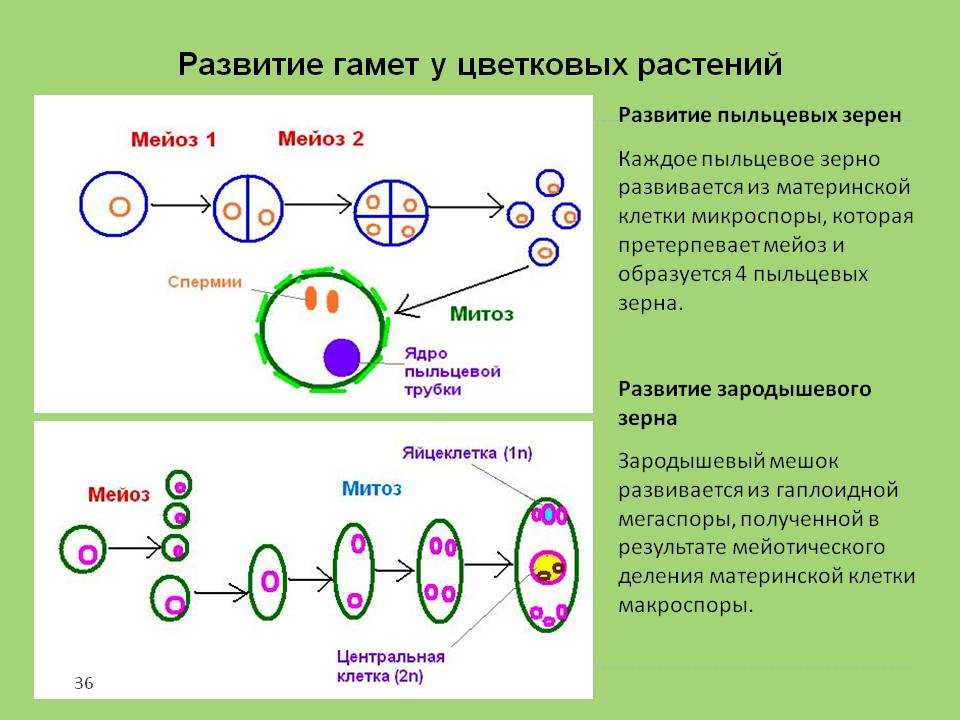 Трубка буквально прорастает в пестик до зародышевого мешка. Пыльцевая трубка проникает в зародышевый мешок, где одна мужская гамета соединяется с яйцеклеткой, образуя зиготу. После митотического деления зигота становится зародышем будущего семени. Вторая мужская гамета сливается с двумя полярными ядрами и образует эндосперм. Этот процесс известен как двойное оплодотворение.[ …]
Трубка буквально прорастает в пестик до зародышевого мешка. Пыльцевая трубка проникает в зародышевый мешок, где одна мужская гамета соединяется с яйцеклеткой, образуя зиготу. После митотического деления зигота становится зародышем будущего семени. Вторая мужская гамета сливается с двумя полярными ядрами и образует эндосперм. Этот процесс известен как двойное оплодотворение.[ …]
Двойное оплодотворение представляет собой исторически сложившееся, качественно новое явление, свойственное только покрытосеменным растениям. Биологическое значение двойного оплодотворения заключается в том, что эндосперм, обогащенный двойной наследственностью, повышает жизнеспособность и приспособленность покрытосеменных, обеспечивает их преимущество перед другими растениями в современную геологическую эпоху.[ …]
Двойное оплодотворение — тип оплодотворения, свойственный покрытосеменным растениям, при котором один из спермиев сливается с яйцеклеткой с образованием диплоидной зиготы, дающей начало зародышу семени, а второй спермпй сливается с диплоидным ядром зародышевого мешка с образованием клетки, дающей начало эндосперму семенп.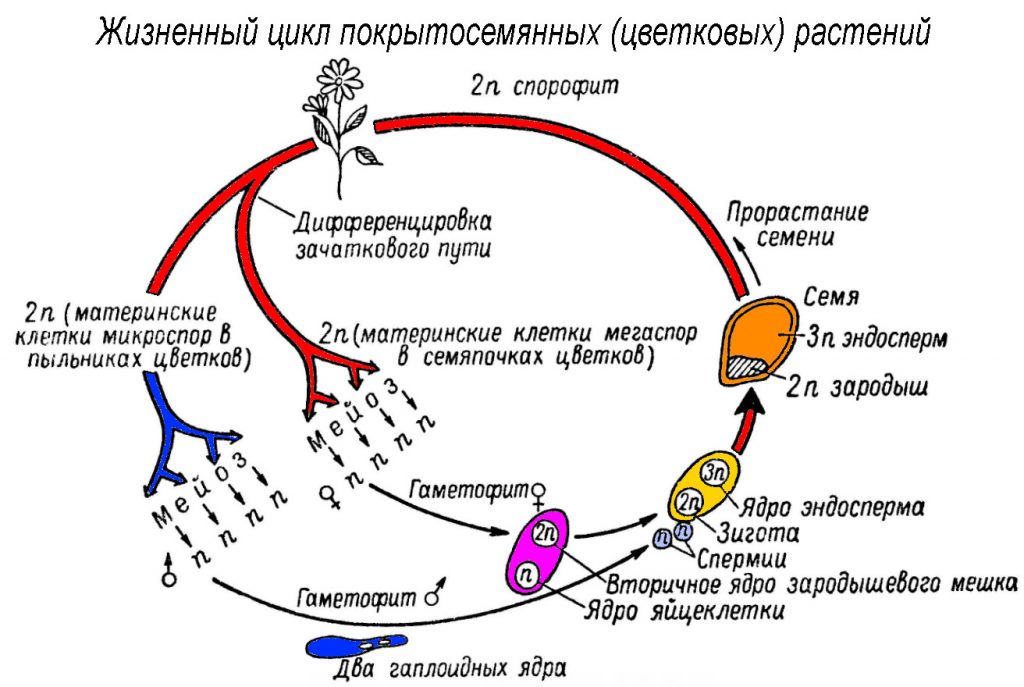 [ …]
[ …]
После двойного оплодотворения первым обычно делится первичное ядро эндосперма (рис, 124). У пшеницы его деление начинается через 3—4 ч после опыления. В это время в цитоплазме зародышевого мешка и в первичном ядре эндосперма интенсивно накапливаются нуклеиновые кислоты (РНК и ДНК), белки, полисахариды, ферменты, витамины и другие физиологически активные вещества. С момента образования зиготы и до полного формирования зародыша эндосперм служит основным источником его питания.[ …]
В момент оплодотворения мужская и женская гаметы сливаются, образуя одну клетку — зиготу, имеющую одно ядро — результат слияния ядер обеих гамет. От каждой из гамет ядро зиготы получает по гаплоидному набору хромосом, и в результате оно имеет двойной, диплоидный набор их. Из зиготы развивается организм, каждая клетка которого имеет диплоидный набор хромосом.[ …]
В спермии двойная спираль ДНК стабильная. Порядок азотистых оснований в мостиках и их относительная численность на известном участке ДНК и дают тот или другой геи, поскольку расположением оснований определяется чередование аминокислот в белке, образующемся в развивающемся животном под влиянием данного участка ДНК.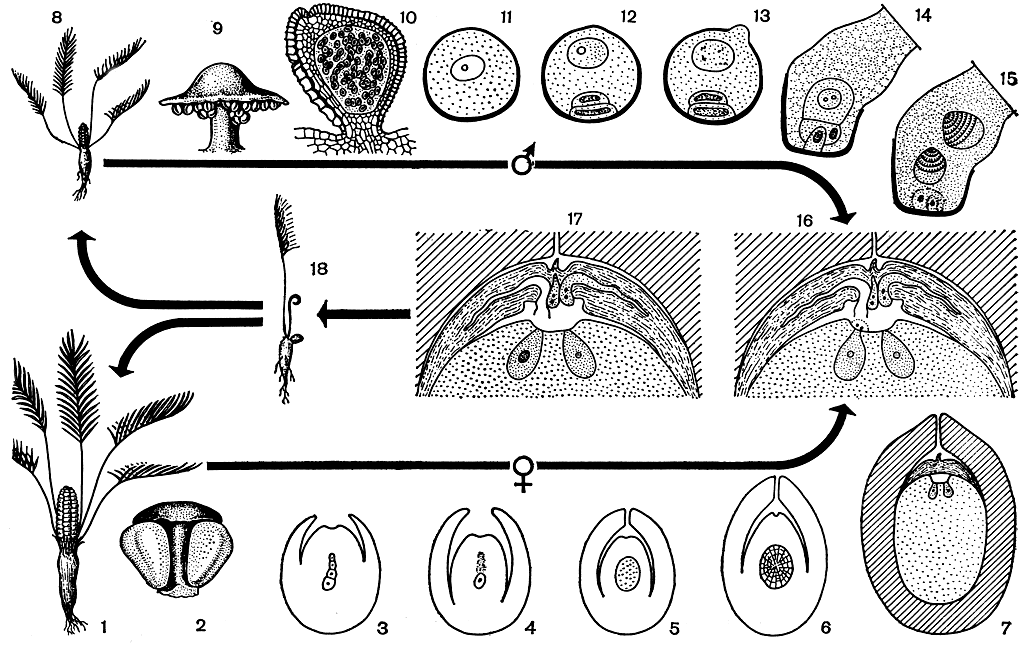 Но белок ДНК образует только после оплодотворения, когда она становится способной к превращению в РНК (рибонуклеиновую кислоту): ДНК->РНК->белок.[ …]
Но белок ДНК образует только после оплодотворения, когда она становится способной к превращению в РНК (рибонуклеиновую кислоту): ДНК->РНК->белок.[ …]
Впоследствии из оплодотворенной яйцеклетки (зиготы) образуется зародыш, а из оплодотворенного ядра центральной клетки — эндосперм. Двойное оплодотворение присуще всем систематическим группам покрытосеменных растений.[ …]
Постмитотический тип двойного оплодотворения детально исследован Е. Н. Герасимовой-Навашиной у рябчика горного (Fritillaria tenella) (рис. Она же изучала промежуточный тип двойного оплодотворения у гальтонии. Премитотический процесс оплодотворения у пшеницы показан на рисунках 111, Л и 113.[ …]
Как было отмечено выше, оплодотворение у цветковых (покрытосеменных) имеет существенную отличительную особенность в виде двойного оплодотворения (С. Г. Навашин, 1896), которое сводится к тому, что в зародышевом мешке гаплоидная яйцеклетка и диплоидная центральная клетка оплодотворяются спермиями, в результате чего образуется диплоидный зародыш и триплоидная клетка, развивающаяся в клетки эндосперма (см.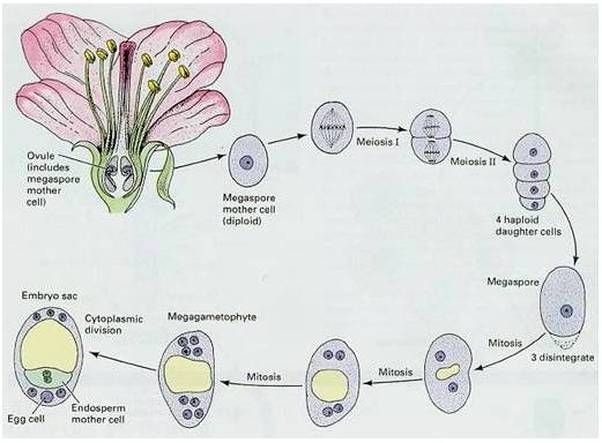 гл. II).[ …]
гл. II).[ …]
Герасимова-Навашнна Е. Н. Двойное оплодотворение покрытосеменных и некоторые теоретические аспекты//Проблемы эмбриологии.— Киев: Наукова думка, 1971.[ …]
Правильным представлением об оплодотворении у покрытосеменных мы обязаны выдающемуся русскому цитологу-эмбриологу С. Г. Навашину, открывшему в 1898 г. двойное оплодотворение у лилейных и астровых. Он впервые показал, что у цветковых растений при оплодотворении один из спермиев сливается с ядром яйцеклетки, а другой — с ядром центральной клетки или полярными ядрами (рис. 109).[ …]
У плодовых растений после завершения двойного оплодотворения из с.емяпочки формируется семя, а наружные покровы семяпочки (или интегументы) превращаются в кожуру семени. Клетки нуцеллуса используются растущим зародышем или реже превращаются в питательную ткань — перисперм. У болв-шинства плодовых и ягодных растений эндосперм и клетки нуцеллуса идут на формирование зародыша, а запасные пластические вещества локализуются в семядолях зародыша (рис.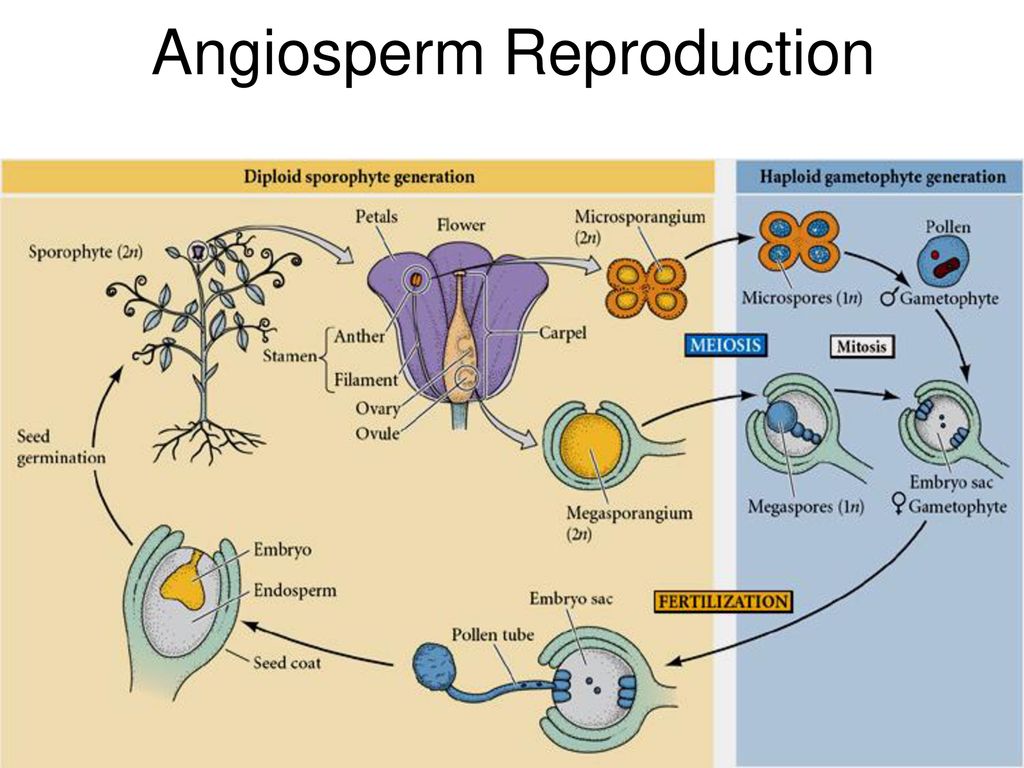 28).[ …]
28).[ …]
При изучении структуры спермиев в процессе двойного оплодотворения С. Г. Навашин обнаружил, что их ядра находятся в состоянии поздней телофазы. Механизм движения спермиев в пыльцевой трубке и зародышевом мешке он рассматривал как активный процесс самостоятельного передвижения.[ …]
При слиянии сперматозоида с яйцеклеткой во время оплодотворения восстанавливается двойной набор хромосом, образуется зигота, из которой развивается организм человека.[ …]
Весь ;>тот процесс, как известно, принято намывать двойным оплодотворением. Ведь согласно общепринятому в литературе определению, оплодотворение (еипгамин) — это процесс слияния, мужской п женской половых клеток (гамет) с образованием зиготы, из которой и дальнейшем разнимается новый организм. Такое определение оплодотворения можно найти в любом учебнике биологии и в любом :нщнклопедичоском словаре (в том число в советских шщпклопедиях, в Большой и Малой). И даже в известном «Словаре русского языка» С. И. Ожегова (1973) мы читаем: «Оплодотворить. 1. Создать зародыш в ком-чем-н. слиянием мужской и женской половых клеток. 2. Послужить источником развития, совершенствования». Слияние одного из сиормпев с яйцеклеткой является, несомненно, оплодотворением, но тройное слияние по является, строго говоря, оплодотворением, так как 1) центральная клетка — не гамета и 2) в результате ¡»того слияния но образуется зигота, из которой и дальнейшем развивался бы новый организм. Очевидно, тройное слияние является оплодотворением лишь в указанном Ожеговым, втором, переносном смысле. Другими словами, в выражении «двойное оплодотворение» термин «оплодотворение» применяется в двух разных, смыслах — прямом и переносном. Тем не менее выражение «двойное онлодотиороние» настолько широко вошло в литературу, что было бы нецелесообразно его заменить (а попытки такого рода делались, в том числе известным, немецким ботаником. Достаточно, если мы будем помнить, что речь идет здесь о двух разных биологических процессах, условно объединяемых общим .названием.[ .
1. Создать зародыш в ком-чем-н. слиянием мужской и женской половых клеток. 2. Послужить источником развития, совершенствования». Слияние одного из сиормпев с яйцеклеткой является, несомненно, оплодотворением, но тройное слияние по является, строго говоря, оплодотворением, так как 1) центральная клетка — не гамета и 2) в результате ¡»того слияния но образуется зигота, из которой и дальнейшем развивался бы новый организм. Очевидно, тройное слияние является оплодотворением лишь в указанном Ожеговым, втором, переносном смысле. Другими словами, в выражении «двойное оплодотворение» термин «оплодотворение» применяется в двух разных, смыслах — прямом и переносном. Тем не менее выражение «двойное онлодотиороние» настолько широко вошло в литературу, что было бы нецелесообразно его заменить (а попытки такого рода делались, в том числе известным, немецким ботаником. Достаточно, если мы будем помнить, что речь идет здесь о двух разных биологических процессах, условно объединяемых общим .названием.[ .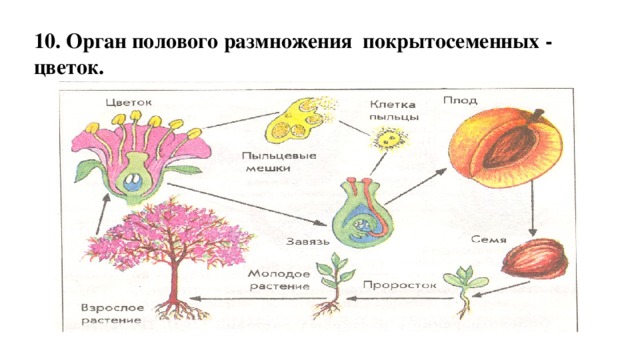 ..]
..]
Дозревание икринок. Вполне развитая в яичнике икринка, прежде чем быть оплодотворенной, должна еще дозреть, в ней происходит процесс изменения зародышевого пузырька или ядра. Всякая клетка, животная и растительная, состоит из протоплазмы и ядра, с ядрышком внутри и особого тельца, состоящего из тончайших радиально расположенных нитей ахроматического (неокрашивающегося) белкового вещества, называемого центрозомой. Эта центрозома лежит то рядом с ядром, то посредине между поверхностью желтка и центрально-лежащим ядром. Самое ядро состоит из нитей хроматина (окрашивающегося вещества, лежащего в массе нуклеина (белковое вещество), и ядрышка в середине. Нити хроматина, или хромозомы, находятся всегда в определенном числе (от одной до нескольких сот у разных видов). Перед делением клетки центрозома делится на две звездочки, отходящие к противоположным полюсам ядра. Ядро же своими нитями ахроматина образует двойной конус центрозомы. Хроматозомы ядра внутри этого двойного конуса или веретена образуют петли, которые передвигаются в середине веретена попарно и располагаются в два ряда, разделяющих веретено как бы пластинкой на две половины. Тогда происходит разделение веретена и всего ядра на две половины, влекущее за собою деление всей клетки протоплазмы, которая окружает каждую из вновь образовавшихся ядер. Этот процесс носит название кариокинезиса, или митозы.[ …]
Тогда происходит разделение веретена и всего ядра на две половины, влекущее за собою деление всей клетки протоплазмы, которая окружает каждую из вновь образовавшихся ядер. Этот процесс носит название кариокинезиса, или митозы.[ …]
Открытие в 1896 г. выдающимся русским ученым Сергеем Гавриловичем Навашииым двойного оплодотворения у покрытоременных растений положило начало новой .эре исследований в биологической науке. С. Г. Навашин является также основателем науки о ядре — кариологии.[ …]
Детально изучая поведение спермиев в зародышевом мешке, Е. Н. Герасимова-Навашина предложила различать два основных типа двойного оплодотворения: премитотический, когда объединение половых ядер происходит перед первым митозом зиготы, и постмитотический, когда объединение половых ядер наступает в начале первого митоза зиготы. Ею установлена также промежуточная форма двойного оплодотворения, характеризующаяся образованием собственной оболочки вокруг ядра спермия при медленном его погружении в ядро яйцеклетки.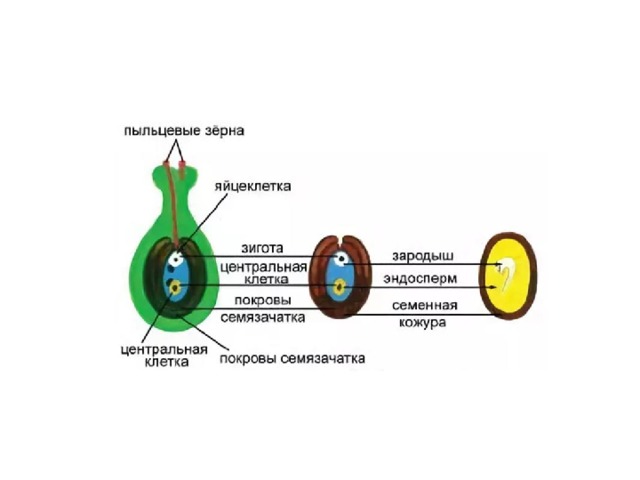 При этом объединение половых ядер происходит во время первого митоза зиготы (рис. 110).[ …]
При этом объединение половых ядер происходит во время первого митоза зиготы (рис. 110).[ …]
Семена у покрытосеменных растений образуются из семяпочки в результате двойного оплодотворения. В пыльце сосредоточено большое количество физиологически активных веществ — фитогормоиов (ауксинов), ферментов, много аминокислоты пролина. Все это приводит к тому, что уже сразу после опыления интенсивность обмена веществ в системе пыльца — пестик резко усиливается. Пыльцевая трубка несет два спермия. В результате слияния первого спермия с яйцеклеткой образуется зигота, дающая при последующем развитии зародыш семени. Анатомически ткани зародыша целиком состоят из первичной меристемы. При слиянии второго спермия со вторичным ядром зародышевого мешка образуется триплоидное ядро. Оно начинает делиться и вместе с цитоплазмой образует множество «леток составляющих эндосперм. После опыления и особенно оплодотворения завязь начинает разрастаться. Вместе с пыльцой в зародышевый мешок привносятся как сами фнтогормоны (ауксины), так и ферменты, катализирующие синтез гормонов.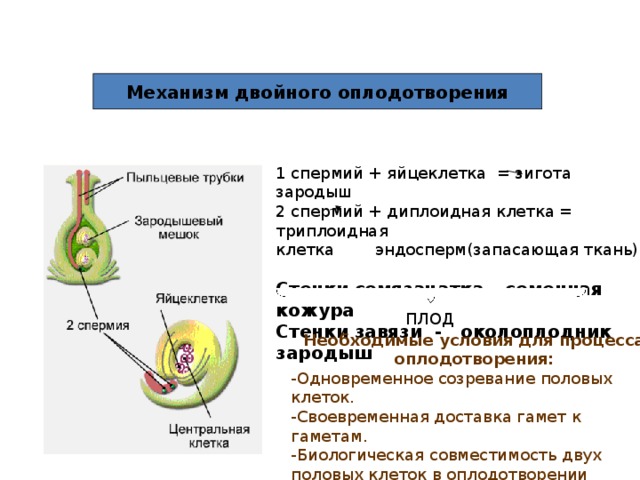 Дыхание развивающегося после оплодотворения семени идет очень иптенсивно. Семя становится центром притяжения питательных веществ. В процессе развития п роста зародыша ткань эндосперма потребляется и к моменту созревания семян может частично или полностью исчезнуть. Некоторые семена лишены эндосперма, питательные вещества сосредоточены у них в тканях самого зародыша (в семядолях).[ …]
Дыхание развивающегося после оплодотворения семени идет очень иптенсивно. Семя становится центром притяжения питательных веществ. В процессе развития п роста зародыша ткань эндосперма потребляется и к моменту созревания семян может частично или полностью исчезнуть. Некоторые семена лишены эндосперма, питательные вещества сосредоточены у них в тканях самого зародыша (в семядолях).[ …]
У одного конца семенного рубчика находится трудно различимый еемявходный след, пли микропиле, — место проникновения пыльцевой трубки в семяпочку при ее оплодотворении. У другого конца рубчика располагается небольшой, чаще двойной, бугорок — халаза, являющийся основанием семяпочки.[ …]
У Crépis capillaris ядра спермиев при слиянии с женскими половыми ядрами находятся в периоде G прерванного митотического цикла (стадия телофазы), который они заканчивают в зиготе (рис. 112, 118). У астровых и злаковых в момент оплодотворения спермии находятся в стадии поздней телофазы митоза. На рисунке 114 показан конец двойного оплодотворения у кукурузы в момент слияния мужских ядрышек с женскими в ядрах зиготы и первичного ядра эндосперма; при этом второй сперматозоид находится в полярных ядрах; в яйцеклетке процесс слияния уже закончен, присутствует ядрышко спермия; слева виден контур второй пыльцевой трубки со спермиями.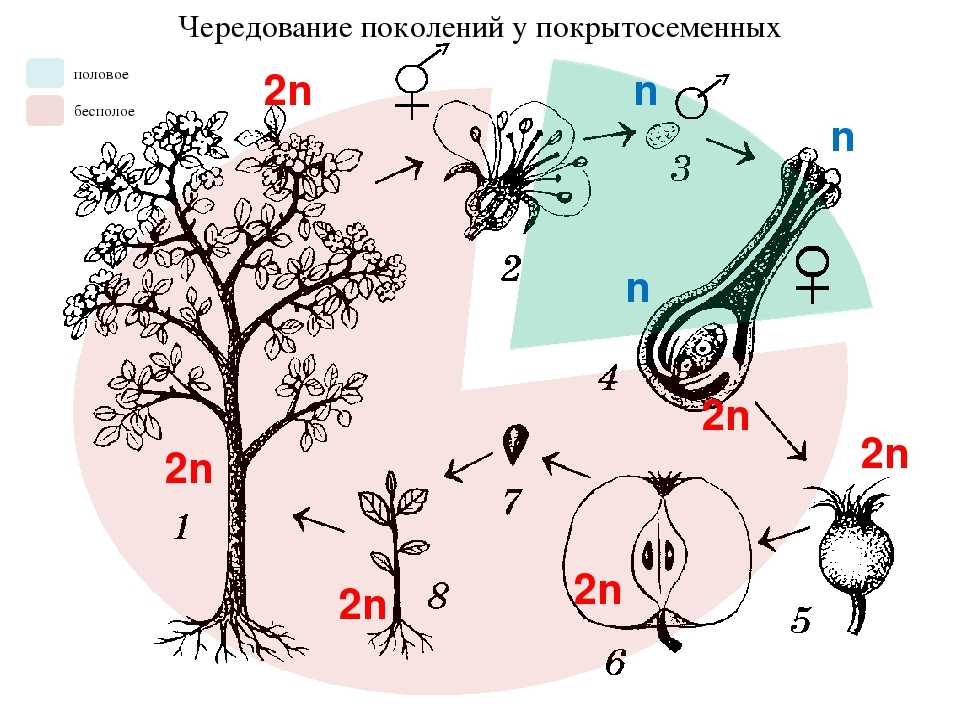 [ …]
[ …]
Заканчивая изложение экспериментов Г. Менделя, свидетельствующих о независимом характере перераспределения генов, остановимся кратко на перераспределении генов при других поли-гибридных скрещиваниях. Количество генных комбинаций в полигибридных скрещиваниях (например, тетрагибридных) возрастает с вовлечением в наблюдения каждой новой пары генов, ибо каждая новая пара приводит к двойному увеличению типов гамет, продуцируемых гибридами Г, к тройному увеличению количества разных генотипов среди гибридов Г2 и к увеличению в 4 раза количества возможных комбинаций при оплодотворении гамет, продуцируемых гибридами Е„.[ …]
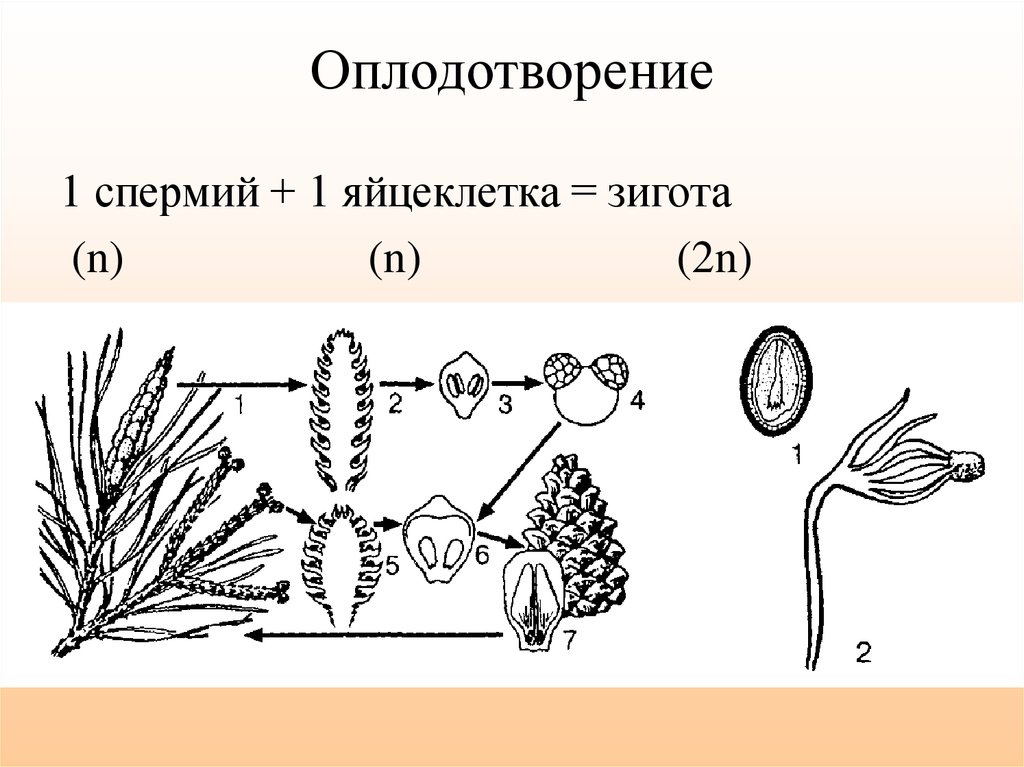
Половое размножение — это образование нового организма при участии двух родительских особей. При половом размножении происходит слияние половых клеток — гамет мужского и женского организма. Новый организм несет наследственную информацию от обоих родителей. Половые клетки формируются в результате особого типа деления. В этом случае в отличие от клеток взрослого организма, которые несут диплоидный (двойной) набор хромосом, образующиеся гаметы имеют гаплоидный (одинарный) набор.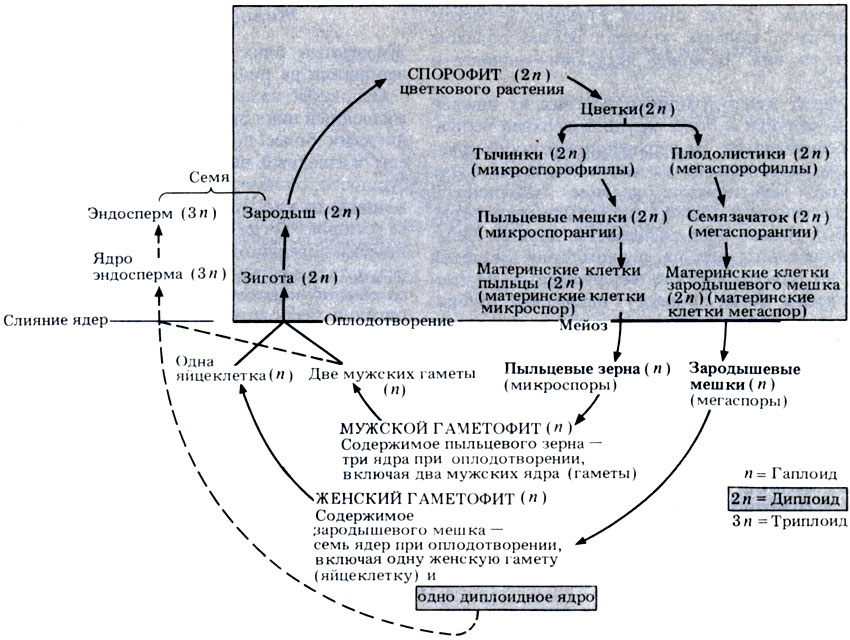 В результате оплодотворения парный, диплоидный набор хромосом восстанавливается. Одна хромосома из пары является отцовской, а другая — материнской. Гаметы образуются в половых железах или в специализированных клетках в процессе мейоза.[ …]
В результате оплодотворения парный, диплоидный набор хромосом восстанавливается. Одна хромосома из пары является отцовской, а другая — материнской. Гаметы образуются в половых железах или в специализированных клетках в процессе мейоза.[ …]
Птицы — животные, приспособленные к полетам в земной атмосфере. Распространены по всему земному шару и насчитывают около 9 тыс. видов. Тело птиц покрыто перьями, а передние конечности превратились в крылья. В строении тела птиц имеются особенности, например, кости скелета у них полые, хорошо развита грудина-киль. Птицы —теплокровные животные (до 42° С). Легкие у них ячеистые и имеются воздушные мешки для активной вентиляции (это так называемое двойное дыхание). Сердце четырехкамерное; артериальная и венозная кровеносная системы разделены; пищеварительная, выделительная и половая системы у птиц и рептилий очень похожи. Нервная система птиц развита весьма хорошо, особенно передний мозг-мозжечок. Поведение птиц весьма сложное и у них выработано много условных рефлексов.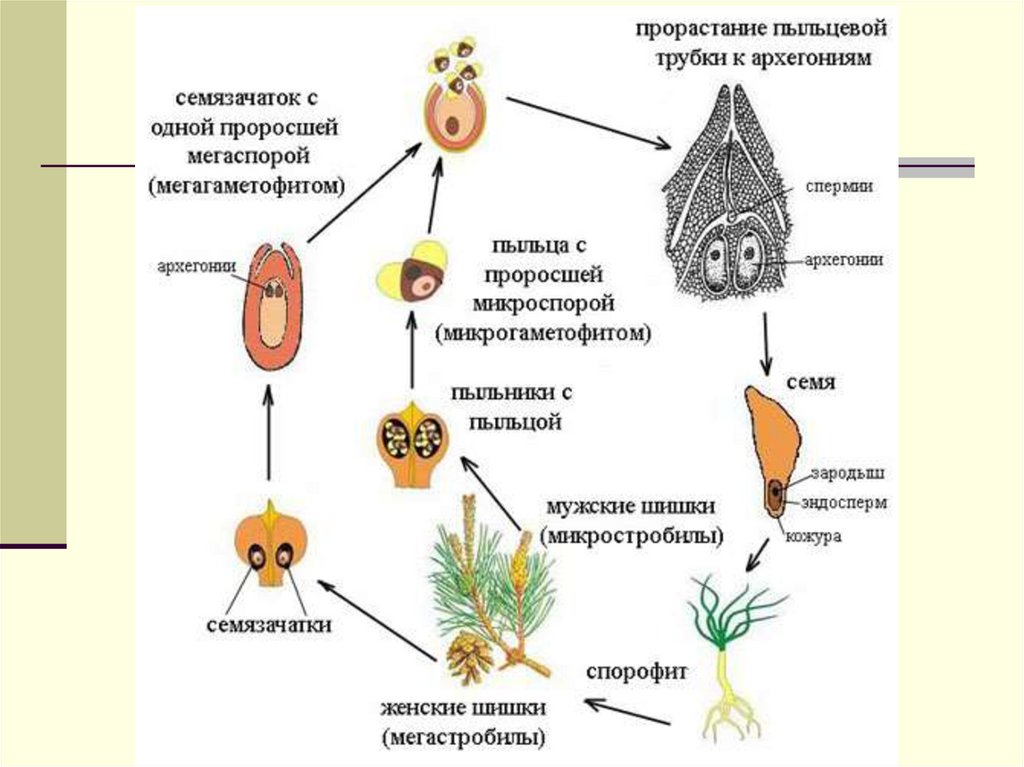 Оплодотворение внутреннее; яйца откладываются, как правило, в гнезда; для птиц, как и рептилий, характерна забота о потомстве.[ …]
Оплодотворение внутреннее; яйца откладываются, как правило, в гнезда; для птиц, как и рептилий, характерна забота о потомстве.[ …]
Жизненный цикл высших растений состоит из двух ритмически чередующихся фаз, или «поколений» (рис. 1),— полового (г а м е т о-ф и т а) и бесполого (спорофита). Половые органы развиваются на гаметофите. Гаметофит может быть обоеполым. В таком случае на нем развиваются как антеридии, так и архегонии. Но у подавляющего большинства высших растений (в том числе у всех голосеменных и цветковых растений) он однополый и несет или только антеридии (мужской гаметофит), или только архегонии (женский гаметофит). В результате оплодотворения, т. е. слияния мужской гаметы с женской, образуется новая клетка с двойным набором хромосом (отцовским и материнским), называемая зиготой.[ …]
Покрытосеменные растения | Определение, воспроизведение, примеры, характеристики, жизненный цикл, таксономия и факты
цветок змеиной тыквы
Посмотреть все СМИ
- Ключевые люди:
- Адольф-Теодор Броньяр
Роберт Браун
Чарльз Э. Бесси
Бесси
Чарльз Джозеф Чемберлен
Джон Бартрам
 Бесси
Бесси- Похожие темы:
- клада магнолид
паразитическое растение
цветок
австробайлейлес
коммелинид
Просмотреть весь связанный контент →
Популярные вопросы
Что такое покрытосеменные растения?
Покрытосеменные – это растения, дающие цветы и содержащие семена в плодах. Это самая большая и разнообразная группа в царстве Plantae, насчитывающая около 300 000 видов. Покрытосеменные растения составляют примерно 80 процентов всех известных живых зеленых растений. Примеры варьируются от обыкновенного одуванчика и трав до древних магнолий и высокоразвитых орхидей. Покрытосеменные также составляют подавляющее большинство всей растительной пищи, которую мы едим, включая зерновые, бобовые, фрукты, овощи и большинство орехов.
цветок
Узнайте больше о цветах, одной из определяющих характеристик покрытосеменных растений.
Чем покрытосеменные отличаются от голосеменных?
Основное различие между покрытосеменными и голосеменными заключается в том, как развиваются их семена. Семена покрытосеменных растений развиваются в завязях цветков и окружены защитным плодом. Семена голосеменных обычно образуются в однополых шишках, известных как стробилы, и у растений отсутствуют плоды и цветы. Кроме того, все покрытосеменные, кроме самых древних, содержат проводящие ткани, известные как сосуды, в то время как голосеменные (за исключением Gnetum ) нет. Покрытосеменные имеют большее разнообразие в своих привычках роста и экологических ролях, чем голосеменные.
Семена покрытосеменных растений развиваются в завязях цветков и окружены защитным плодом. Семена голосеменных обычно образуются в однополых шишках, известных как стробилы, и у растений отсутствуют плоды и цветы. Кроме того, все покрытосеменные, кроме самых древних, содержат проводящие ткани, известные как сосуды, в то время как голосеменные (за исключением Gnetum ) нет. Покрытосеменные имеют большее разнообразие в своих привычках роста и экологических ролях, чем голосеменные.
Подробнее ниже:
Структура и функция: соцветия
В чем разница между покрытосеменными и голосеменными растениями?
Узнайте больше о различиях между покрытосеменными и голосеменными растениями.
Чем похожи покрытосеменные и голосеменные растения?
Обе группы сосудистых растений содержат ксилему и флоэму. За исключением очень немногих видов покрытосеменных (например, облигатных паразитов и микогетеротрофов), обе группы полагаются на фотосинтез для получения энергии.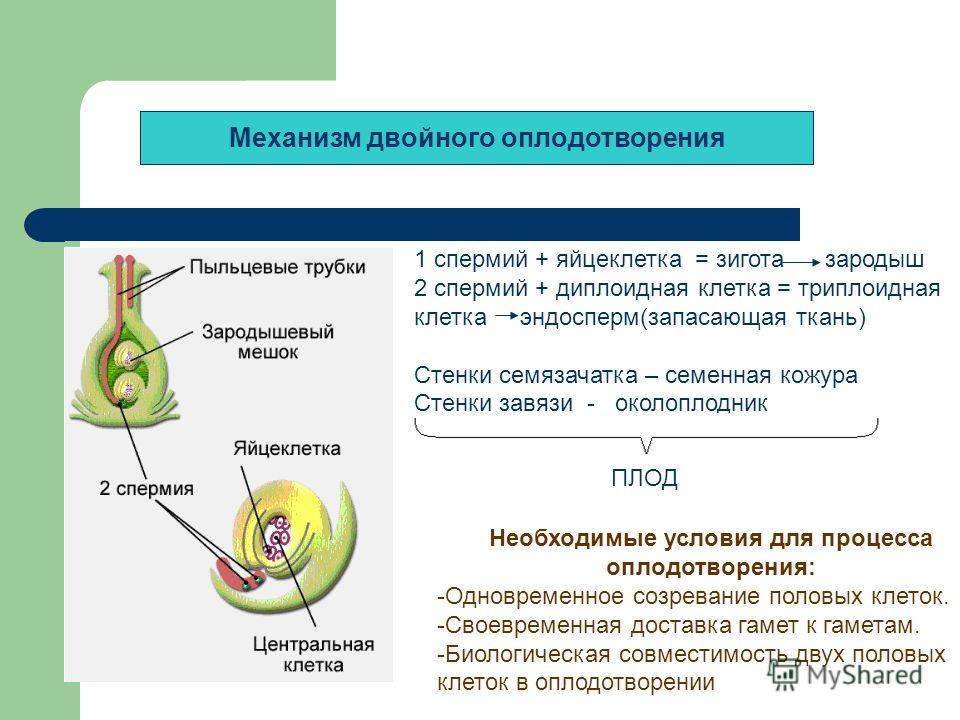 И покрытосеменные, и голосеменные используют семена в качестве основного средства размножения, и оба используют пыльцу для облегчения оплодотворения. Голосеменные и покрытосеменные имеют жизненный цикл, включающий смену поколений, и оба имеют редуцированную стадию гаметофита.
И покрытосеменные, и голосеменные используют семена в качестве основного средства размножения, и оба используют пыльцу для облегчения оплодотворения. Голосеменные и покрытосеменные имеют жизненный цикл, включающий смену поколений, и оба имеют редуцированную стадию гаметофита.
голосеменные
Прочтите о голосеменных растениях.
покрытосеменное , также называемое цветковое растение , любой из примерно 300 000 видов цветковых растений, самой большой и разнообразной группы в царстве Plantae. Покрытосеменные растения составляют примерно 80 процентов всех известных ныне живущих зеленых растений. Покрытосеменные — это сосудистые семенные растения, у которых семязачаток (яйцо) оплодотворяется и развивается в семя в закрытой полой завязи. Сама завязь обычно заключена в цветке, той части покрытосеменного растения, которая содержит мужские или женские репродуктивные органы или и то, и другое. Плоды образуются из созревающих цветочных органов покрытосеменных растений и поэтому характерны для покрытосеменных растений. Напротив, у голосеменных растений (например, хвойных и саговниковых), другой большой группы сосудистых семенных растений, семена развиваются не в завязи, а обычно остаются открытыми на поверхности репродуктивных структур, таких как шишки.
Напротив, у голосеменных растений (например, хвойных и саговниковых), другой большой группы сосудистых семенных растений, семена развиваются не в завязи, а обычно остаются открытыми на поверхности репродуктивных структур, таких как шишки.
Узнайте, как покрытосеменные и голосеменные растения хранят свои семена
Просмотреть все видео к этой статье
В отличие от таких несосудистых растений, как мохообразные, у которых все клетки в теле растения участвуют во всех функциях, необходимых для поддержки, питания и расширения тела (например, питание, фотосинтез и деление клеток), покрытосеменные развили специализированные клетки и ткани, которые выполняют эти функции, а затем развили специализированные сосудистые ткани (ксилема и флоэма), которые переносят воду и питательные вещества во все области тела растения. . Специализация растительного тела, развившаяся как приспособление к преимущественно наземной среде обитания, включает обширные корневые системы, закрепляющие растение и поглощающие воду и минеральные вещества из почвы; стебель, поддерживающий растущее тело растения; и листья, которые являются основными местами фотосинтеза для большинства покрытосеменных растений. Еще одним значительным эволюционным достижением по сравнению с несосудистыми и более примитивными сосудистыми растениями является наличие локализованных областей для роста растений, называемых меристемами и камбиями, которые увеличивают длину и ширину тела растения соответственно. За исключением определенных условий, эти области являются единственными областями, в которых происходит митотическое деление клеток в теле растения, хотя дифференцировка клеток продолжается в течение всей жизни растения.
Еще одним значительным эволюционным достижением по сравнению с несосудистыми и более примитивными сосудистыми растениями является наличие локализованных областей для роста растений, называемых меристемами и камбиями, которые увеличивают длину и ширину тела растения соответственно. За исключением определенных условий, эти области являются единственными областями, в которых происходит митотическое деление клеток в теле растения, хотя дифференцировка клеток продолжается в течение всей жизни растения.
Покрытосеменные доминируют на поверхности Земли и растительности в большем количестве сред, особенно в наземных местообитаниях, чем любая другая группа растений. В результате покрытосеменные растения являются наиболее важным конечным источником пищи для птиц и млекопитающих, включая человека. Кроме того, цветковые растения являются наиболее экономически важной группой зеленых растений, служащих источником фармацевтических препаратов, продуктов из волокна, древесины, декоративных растений и других коммерческих продуктов.
Хотя таксономия покрытосеменных до сих пор известна не полностью, последняя система классификации включает в себя большой объем сравнительных данных, полученных в результате изучения последовательностей ДНК. Он известен как система ботанической классификации покрытосеменных Phylogeny Group IV (APG IV). Покрытосеменные стали считаться группой на уровне отдела (сопоставимом с уровнем типа в системах классификации животных), называемой Anthophyta, хотя система APG признает только неформальные группы выше уровня порядка.
Викторина «Британника»
Назови этот цветок!
Можете ли вы отличить сирень от гиацинта?
В этой статье порядки или семейства даны, как правило, в скобках, после народного или научного названия растения. В соответствии с таксономическими соглашениями роды и виды выделены курсивом. Высшие таксоны легко идентифицируются по их суффиксам: семейства оканчиваются на -aceae , а порядки — на -ales .
Для сравнения покрытосеменных растений с другими основными группами растений см.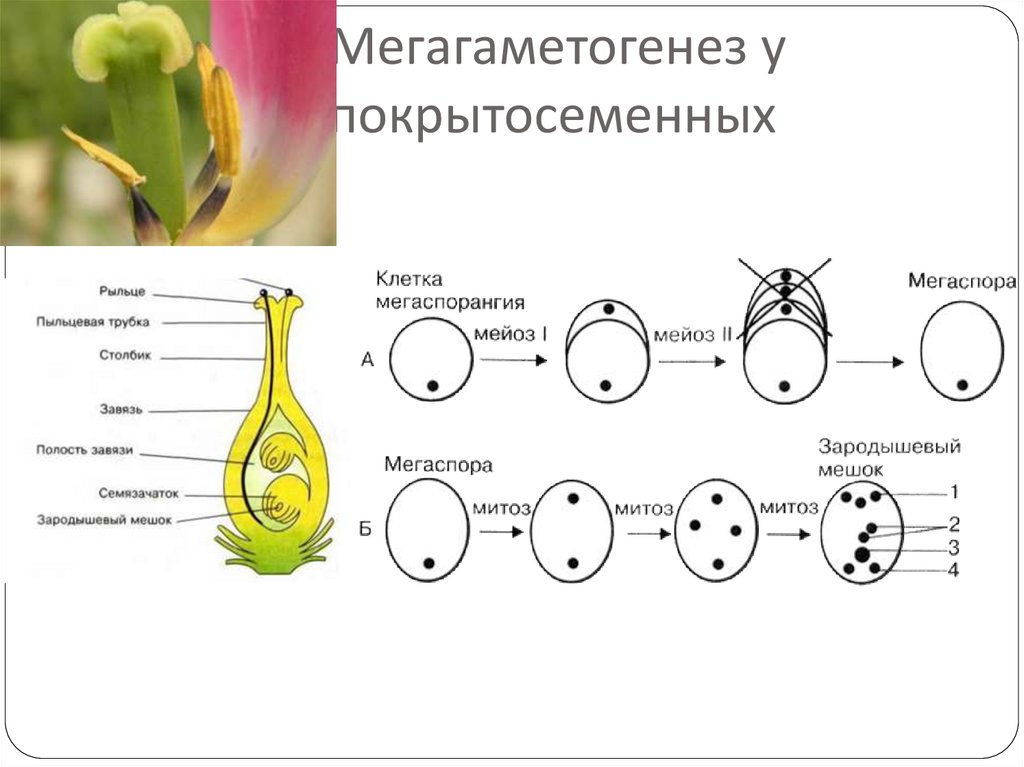 растение, мохообразное, папоротник, низшее сосудистое растение и голосеменное растение.
растение, мохообразное, папоротник, низшее сосудистое растение и голосеменное растение.
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Подпишитесь сейчас
Развитие зигот растений: последние идеи и применение к клональным семенам
Обзор
. 2021 фев; 59:101993.
doi: 10.1016/j.pbi.2020.101993.
Epub 2021 7 января.
Имтияз Хандай
1
, Венкатесан Сундаресан
2
Принадлежности
- 1 Факультет биологии растений, Калифорнийский университет, Дэвис, Калифорния, США; Инновационный институт геномики, Калифорнийский университет, Беркли, Калифорния, США.
- 2 Кафедра биологии растений, Калифорнийский университет, Дэвис, Калифорния, США; Институт инновационной геномики, Калифорнийский университет, Беркли, Калифорния, США; Департамент наук о растениях, Калифорнийский университет, Дэвис, Калифорния, США. Электронный адрес: [email protected].
 Электронный адрес:
Электронный адрес: PMID:
33422964
DOI:
10.1016/j.pbi.2020.101993
Обзор
Имтияз Хандай и др.
Curr Opin Plant Biol.
2021 фев.
. 2021 фев; 59:101993.
дои: 10.1016/j.pbi.2020.101993.
Epub 2021 7 января.
Авторы
Имтияз Хандай
1
, Венкатесан Сундаресан
2
Принадлежности
- 1 Факультет биологии растений, Калифорнийский университет, Дэвис, Калифорния, США; Инновационный институт геномики, Калифорнийский университет, Беркли, Калифорния, США.
- 2 Кафедра биологии растений, Калифорнийский университет, Дэвис, Калифорния, США; Институт инновационной геномики, Калифорнийский университет, Беркли, Калифорния, США; Департамент наук о растениях, Калифорнийский университет, Дэвис, Калифорния, США. Электронный адрес: [email protected].
PMID:
33422964
DOI:
10.1016/j.pbi.2020.101993
Абстрактный
У цветковых растений гаплоидные гаметы — яйцеклетка и сперматозоид сливаются, образуя первую диплоидную клетку — зиготу. Зигота является исходной стволовой клеткой, дающей начало всем эмбриональным и постэмбриональным тканям и органам. В отличие от животных, продукты как материнских, так и отцовских генов участвуют в начальном развитии зигот у растений.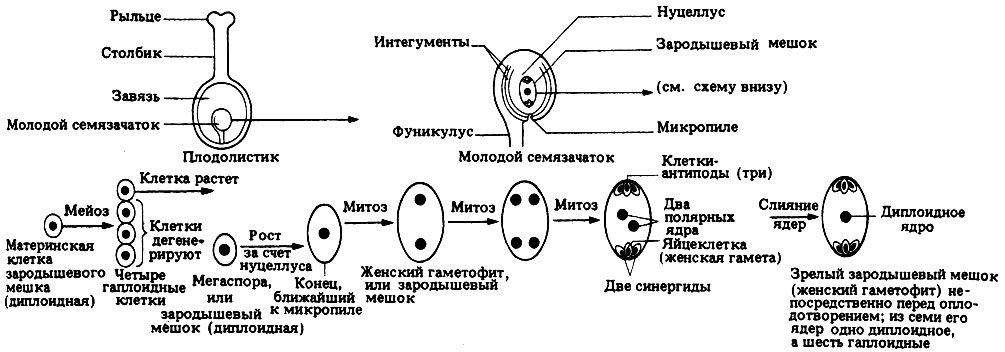 Здесь мы обсуждаем последние достижения в понимании зиготического перехода и инициации зародыша у покрытосеменных, включая роль родительского вклада в экспрессию генов в зиготе. Далее мы обсудим использование этих знаний в сельскохозяйственной биотехнологии посредством синтетического апомиксиса. Партеногенез, полученный путем манипулирования эмбриогенными факторами в сочетании с мутациями, минующими мейоз, делает возможным клональное размножение гибридных культур через семена.
Здесь мы обсуждаем последние достижения в понимании зиготического перехода и инициации зародыша у покрытосеменных, включая роль родительского вклада в экспрессию генов в зиготе. Далее мы обсудим использование этих знаний в сельскохозяйственной биотехнологии посредством синтетического апомиксиса. Партеногенез, полученный путем манипулирования эмбриогенными факторами в сочетании с мутациями, минующими мейоз, делает возможным клональное размножение гибридных культур через семена.
Copyright © 2021 Elsevier Ltd. Все права защищены.
Похожие статьи
Успехи в изучении активации зигот у высших растений.
Li DX, Chen SJ, штаб-квартира Tian.
Ли Д.С. и др.
Зигота. 2021 фев; 29(1):12-19. дои: 10.1017/S0967199420000568. Epub 2020 15 октября.
Зигота. 2021.PMID: 33054867
Обзор.
Эмбриогенный триггер риса, экспрессируемый самцами, перенаправлен на бесполое размножение через семена.
Хандай И., Скиннер Д., Ян Б., Мерсье Р., Сундаресан В.
Хандай I и др.
Природа. 2019 Январь; 565 (7737): 91-95. doi: 10.1038/s41586-018-0785-8. Epub 2018 12 декабря.
Природа. 2019.PMID: 30542157
Мейоз, нередуцированные гаметы и партеногенез: значение для инженерии образования клональных семян у сельскохозяйственных культур.
Ронсерет А., Вьель-Кальсада Дж.П.
Ронсере А. и др.
Завод Репрод. 2015 июнь; 28 (2): 91-102. doi: 10.1007/s00497-015-0262-6. Epub 2015 22 марта.
Завод Репрод. 2015.PMID: 25796397
Обзор.
Можем ли мы использовать редактирование генов, чтобы вызвать апомиксис у половых растений?
Шебен А., Хойсгаард Д.
Шебен А. и соавт.
Гены (Базель). 2020 12 июля; 11 (7): 781. doi: 10.3390/genes11070781.
Гены (Базель). 2020.PMID: 32664641
Бесплатная статья ЧВК.Инженерный апомиксис: клональные семена приближаются к полям.
Андервуд СиДжей, Мерсье Р.
Андервуд CJ и др.
Annu Rev Plant Biol. 2022 20 мая; 73: 201-225. doi: 10.1146/annurev-arplant-102720-013958. Epub 2022 9 февраля.
Annu Rev Plant Biol. 2022.PMID: 35138881
Обзор.
 doi: 10.3390/genes11070781.
doi: 10.3390/genes11070781.Посмотреть все похожие статьи
Цитируется
Способы доставки биомакромолекул растений в 21 веке.
Рустги С., Навид С., Виндхэм Дж., Чжан Х., Демирер Г.С.
Рустги С. и др.
Передний геном Эд. 2022, 14 октября; 4:1011934. doi: 10.3389/fgeed.2022.1011934. Электронная коллекция 2022.
Передний геном Эд. 2022.PMID: 36311974
Бесплатная статья ЧВК.Обзор.
Контролируемая индукция партеногенеза в трансгенном рисе через Посттрансляционная активация PsASGR-BBML .
Сидху Г.С., Коннер Дж.А., Озиас-Акинс П.
Сидху Г.С. и др.
Фронт завод науч. 2022, 8 июля; 13:925467. doi: 10.3389/fpls.2022.925467. Электронная коллекция 2022.
Фронт завод науч. 2022.PMID: 35873991
Бесплатная статья ЧВК.Миграция проверетена перед первым асимметричным делением в прорастающей споре Marchantia polymorpha .
Сакаи Ю., Хигаки Т., Исидзаки К., Нишихама Р., Кочи Т., Хасэдзава С.
Сакаи Ю. и др.
Завод Биотехнолог (Токио). 2022 25 марта; 39 (1): 5-12. doi: 10.5511/plantbiotechnology.