
Содержание
Однодомные и двудомные растения и их основные характеристики
На основании чего классифицируются растения? Вариантов много, но сейчас мы разберемся с однодомными и двудомными растениями.
Все известные на сегодня растения можно классифицировать на 3 большие группы:
- Однодомные растения. У однодомного растения разнополые соцветия располагаются на одном растении.
- Двудомные растения. У представителей этой группы разнополые соцветия находятся на разных растениях.
- Многодомные. На одном растении можно обнаружить две разновидности соцветий. Полигамия свойственна конскому каштану, винограду, незабудке, ясеню.
В первых двух случаях цветки могут быть обоеполыми (у них есть пестики и тычинки) и раздельнополыми (у них есть или пестик, или тычинка).
Характеристика однодомных растений
Что такое однодомные растения?
Некоторые ученые пришли к выводу, что вначале были двуполые цветки, а из них уже возникли однополые. Произошло это в результате эволюционных процессов.
Произошло это в результате эволюционных процессов.
Определение 1
Однодомные растения — это растения, которые характеризуются наличием на одной особи тычиночных и пестичных соцветий.
Поскольку цветки обоих полов находятся «в одном доме», то такие растения получили название однодомные.
У цветков некоторых растений нет сформированного околоцветника — они опыляются ветром и, иногда, насекомыми (энтомофилия). Многие растения самоопыляются — в этом случае опыление происходит в чаше одного цветка. Обычно пыльца попадает в лоно с других соцветий, которые находятся на этом же цветке. Это, в частности, негативно сказывается на свойствах семян.
Пример 1
Однодомных растений очень много. Примеры однодомных растений — кукуруза, арбуз, тыква, лещина, орех, ольха, дуб, береза, грецкий орех.
В стрессовых ситуациях двудомные растения могут реорганизовываться в однодомные. Так иногда происходит с коноплей.
Грецкий орех — типичный представитель однодомных опыляемых ветром растений.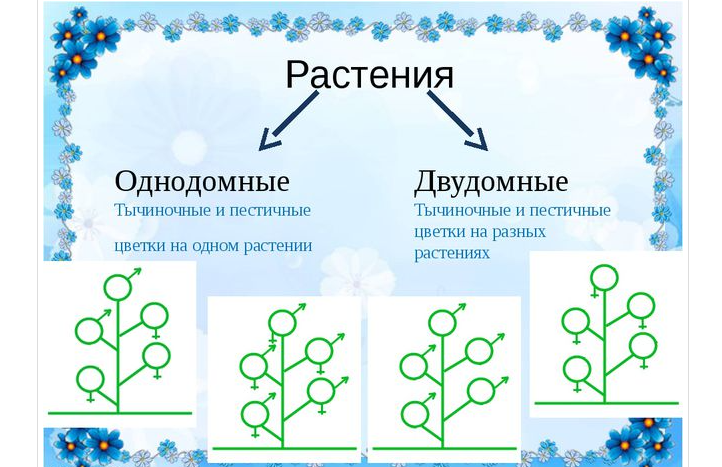 Цветки грецкого ореха могут опыляться пчелами, но только мужские. Женские цветки пчелы игнорируют. Поэтому о какой-либо значимости опыления пчелами цветков ореха говорить сложно. На одном и том же растении мужские и женские цветки распускаются с довольно большой разницей: иногда она достигает 15 дней. Как результат — происходит перекрестное опыление.
Цветки грецкого ореха могут опыляться пчелами, но только мужские. Женские цветки пчелы игнорируют. Поэтому о какой-либо значимости опыления пчелами цветков ореха говорить сложно. На одном и том же растении мужские и женские цветки распускаются с довольно большой разницей: иногда она достигает 15 дней. Как результат — происходит перекрестное опыление.
Лещина тоже относится к однодомным растениям. Мужские цветки у нее находятся в повисающих сережках, а женские — скрыты внутри почек и наружу показываются только малиновые рыльца. Опыление лещины происходит при помощи ветра. Лещина имеет односемянной орех буро-желтого цвета: он окружен колокольчатой плюской из видоизмененных прицветников.
Пример 2
Кустарник лещины — универсальное однодомное растение.
Характеристика двудомных растений
Что такое двудомные растения?
Определение 2
Двудомное растение — это растение, у которых женские и мужские цветки растут на разных растениях одного вида, что приводит к явным внешним различиям между цветками.
Процесс оплодотворения невозможен без перекрестного опыления, в ходе которого пыльца из пыльников мужских цветков переносится на рыльца женских цветков. Самую главную роль в этом переносе выполняют насекомые. Для их привлечения растения используют большие и яркие цветки. Такое опыление — более совершенное, так как позволяет укрепить вид.
К примеру, большинство фруктовых растений нуждается в представителях обоих полов. Один цветок мужского вида используется, чтобы опылить сразу несколько цветков женского вида. Только после этого происходит формирование плодов на женских цветках. По этой же причине нет необходимости обеспечивать каждое женское растение представителем противоположного пола: один мужской представитель способен опылять ряд женских особей. Сколько именно женских особей — зависит от конкретного вида.
Пример 3
Рощу финиковых пальм оплодотворяет несколько деревьев мужского пола. Один может опылить от 40 до 50 пальм.
Замечание 1
Для улучшения опыления на деревья женского пола могут привить ветку мужского дерева.
Чтобы применять все эти знания на практике, нужно не только различать двудомные растения, но и определять пол в рамках одного вида, что не всегда просто сделать.
При рассмотрении строения мужского и женского цветков можно обнаружить, что рыльце мужского цветка развито слабо или его вообще нет. При этом тычинки усеяны пыльцой.
У женских цветков тычинок практически нет, а если и есть, то такая тычинка содержит совсем мало пыльцы.
Все это должны держать в уме, в первую очередь, садоводы.
К примеру, в саду есть неплодоносящее дерево. Чаще всего такое дерево двудомное. Чтобы оно плодоносило, необходимо определить его пол, а затем высадить на участок дерево противоположного пола. Также можно привить к уже растущему дереву веточку дерева противоположного пола.
Замечание 2
Многие ландшафтные дизайнеры и просто те, кто украшают свой участок, предпочитают высаживать только растения одного пола, которые не будут плодоносить и портить опавшими плодами участок.
Мужские двудомные растения иногда производят очень много пыльцы. Так происходит, когда рядом нет женских особей. Большое количество пыльцы объясняется тем, что так повышаются ее шансы долететь до тычинок женских особей. Пыльца может летать за счет своей формы и небольшого веса.
Так происходит, когда рядом нет женских особей. Большое количество пыльцы объясняется тем, что так повышаются ее шансы долететь до тычинок женских особей. Пыльца может летать за счет своей формы и небольшого веса.
К двудомным растениям относится инжир. У инжира цветки мелкие и отличаются невзрачностью. Плодоносят только женские особи. Опыление инжира происходит только при помощи осы-бластофаги. Самцы такой осы прячутся в цветках мужских особей инжира, куда и прилетаем самка для оплодотворения. В процессе оса собирает брюхом пыльцу мужских цветков инжира, после чего разносит ее на тычинки женских цветков.
Встречаются такие двудомные растения, у которых разница между половыми хромосомами практически неразличима. Пример двудомного растения такого типа — конопля. В экстремальной ситуации она может из двудомного растения трансформироваться в однодомное. Но селекционеры выводят ее и как однодомное растение.
Замечание 3
Конопля с мужскими цветками — посконь или замашка, а с женскими — матерка.
Последняя отличается более толстыми стеблями, обилием листьев и высокорослостью. Матерка зреет позже поскони. Посконь сохнет практически сразу после цветения. Коноплю сеют из соотношения мужских и женских особей 1:1. При этом урожай все равно бывает разный. Матерка способна воспроизводить до трети общего урожая.
Замечание 3
Есть двудомные цветковые растения с промежуточными мужскими и женскими особями. Все это указывает на то, что механизм определения пола в настоящий момент довольно сложный.
Двудомные растения обладают специфическими половыми хромосомами наподобие тех, что есть у животных. Первым выявил половые хромосомы у растения печеночного мха в 1917 году Аллен. Растения мхов отличаются гаплоидностью, а вот спорангий и ножка диплоидные. Ученый обнаружил, что у мужского растения мха 7 обычных хромосом и одна небольшая Y-хромосома. У женского растения 7 Y-хромосом и одна длинная X-хромосома.
В процессе оплодотворения происходит объединение этих двух хромосомных наборов и образование спорофита с набором 14A+X-ЬY. Стадия мейоза характеризуется образованием семи пар аутосом одной пары XY. Как результат, половина спор получает набор 7A+X, а вторая — 7A+Y. Из этих спор развиваются мужские и женские особи вида.
Стадия мейоза характеризуется образованием семи пар аутосом одной пары XY. Как результат, половина спор получает набор 7A+X, а вторая — 7A+Y. Из этих спор развиваются мужские и женские особи вида.
Сегодня благодаря селекции пол растений можно смещать. Можно поменять число женских цветков у огурца и шпината: для этого нужно обработать растения перед самым цветением моноокисью углерода, этиленом или прочими восстановителями.
Соотношение мужских и женских генеративных органов (цветка) может сдвигаться в зависимости от условий минерального питания, фотопериодичности, температурного режима.
Решение задач
от 1 дня / от 150 р.
Курсовая работа
от 5 дней / от 1800 р.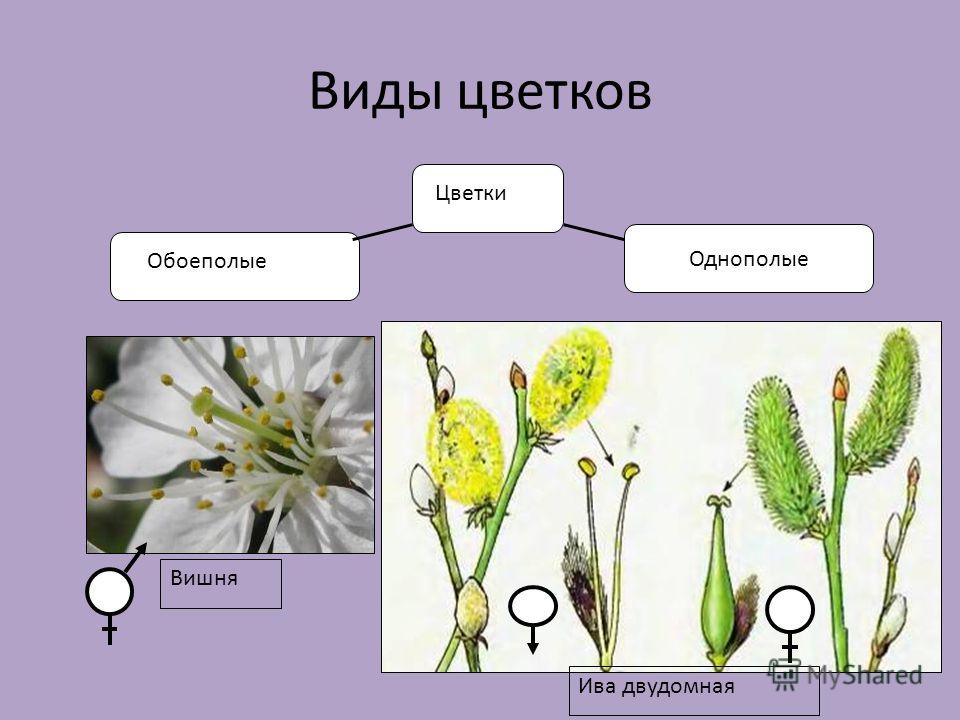
Реферат
от 1 дня / от 700 р.
Пол растения. Условия формирования. Опыление.
DzagiGrow
- /
Блог
 org/ListItem»>
org/ListItem»>/
Пол растения. Условия формирования. Опыление.
Вопрос о формировании пола у растений относится к самым сложным и наименее изученным проблемам биологии развития растений. Вместе с тем, его изучение имеет непосредственное значение для разработки приемов повышения урожайности многих сельскохозяйственных культур, особенно у тех, которые образуют раздельнополые цветки.
Определение пола у растений
Под определением пола у растений понимается формирование признаков пола у клеток, органов или особей под воздействием как генетических факторов, так и условий внешней и внутренней среды. По наличию и степени развития генеративных органов цветки делят на обоеполые (гермафродитные) и однополые (раздельнополые). Последние бывают пестичными (женскими) или тычиночными (мужскими). Если на одном растении находятся цветки разных типов, то такие растения называют однодомными. Растения, имеющие на одних экземплярах пестичные цветки, а на других — тычиночные, называются двудомными. Интересно отметить, что только 4-5% видов растений двудомны. Представителями двудомных растений являются крапивные, хмель, облепиха, тополь, ива, щавель кислый, шпинат и др.
Растения, имеющие на одних экземплярах пестичные цветки, а на других — тычиночные, называются двудомными. Интересно отметить, что только 4-5% видов растений двудомны. Представителями двудомных растений являются крапивные, хмель, облепиха, тополь, ива, щавель кислый, шпинат и др.
Процесс формирования пола происходит у растений в разные периоды его развития и определяется в основном влиянием внешних условий. Дело в том, что в отличие от животных организмов процесс формирования пола у растений подвержен большим колебаниям. Под действием различных причин может происходить даже полное превращение одного пола в другой. Это связано с регуляторными процессами, обусловленными действием факторов внешней среды, и внутренними изменениями метаболического характера.
Влияние внешних факторов на определение пола у растений
Многочисленные факты свидетельствуют о влиянии на проявление признаков пола у растений таких внешних воздействий, как влажность почвы и воздуха, температура, спектральный состав света, условия минерального питания. Так, появлению женских цветков и формированию женских растений у двудомных видов способствуют низкие температуры, высокая влажность, хорошее азотное питание. Появлению мужских цветков — высокие температуры, низкая влажность, калийное питание. Определение пола зависит также от длины дня. Доказано, что короткий день приводит к появлению большего количества женских особей. Длинный световой день действует противоположным образом.
Так, появлению женских цветков и формированию женских растений у двудомных видов способствуют низкие температуры, высокая влажность, хорошее азотное питание. Появлению мужских цветков — высокие температуры, низкая влажность, калийное питание. Определение пола зависит также от длины дня. Доказано, что короткий день приводит к появлению большего количества женских особей. Длинный световой день действует противоположным образом.
Влияние фитогормонов на определение пола у растений
Исследования, проведенные на двудомных растениях (крапивные) и на однодомных с раздельнополыми цветками (кукуруза, огурцы) показали роль фитогормонов в формировании пола растения. Основной парой фитогормонов, контролирующих проявление пола у растений, являются гиббереллины и цитокинины. Гиббереллины принимают участие в формировании признаков мужского пола, а цитокинины и ауксины — женских признаков.
Уровень фитогормонов в растении определяется не только действием внешней среды, но и коррелятивными взаимосвязями между органами. Были проведены эксперименты по влиянию удаления части побегов или обрезки корней на формирование признаков пола. Удаление корней у крапивных способствовало увеличению числа мужских растений. Введение цитокинина в среду выращивания этих же растений приводило к образованию женских цветков. Предполагают, что цитокинины, образующиеся в корнях, передвигаясь в апекс (прим.: верхушка побега), включают программу, обусловливающую проявление женского пола, тогда как гиббереллины, образующиеся в листьях, включают программу, связанную с мужской сексуализацией. Однако если половая дифференцировка клеток уже началась, то воздействие фитогормонами не приведет к изменению направленности в формировании пола, предопределенного генетически. Развитие растений осуществляется по определенной генетической программе. Развертывание этой программы, в том числе проявление пола, старение и смерть организма, может в определенной мере контролироваться внешними условиями, через внутреннюю гормональную систему регуляции.
Были проведены эксперименты по влиянию удаления части побегов или обрезки корней на формирование признаков пола. Удаление корней у крапивных способствовало увеличению числа мужских растений. Введение цитокинина в среду выращивания этих же растений приводило к образованию женских цветков. Предполагают, что цитокинины, образующиеся в корнях, передвигаясь в апекс (прим.: верхушка побега), включают программу, обусловливающую проявление женского пола, тогда как гиббереллины, образующиеся в листьях, включают программу, связанную с мужской сексуализацией. Однако если половая дифференцировка клеток уже началась, то воздействие фитогормонами не приведет к изменению направленности в формировании пола, предопределенного генетически. Развитие растений осуществляется по определенной генетической программе. Развертывание этой программы, в том числе проявление пола, старение и смерть организма, может в определенной мере контролироваться внешними условиями, через внутреннюю гормональную систему регуляции. Таким образом, развитие растений происходит при взаимодействии генетических потенций и факторов внешней среды.
Таким образом, развитие растений происходит при взаимодействии генетических потенций и факторов внешней среды.
Опыление
Процесс переноса пыльцы с пыльника (мужской цветок) на рыльце пестика (женский цветок) называется опылением.
Однодомные растения имеют и мужские и женские цветки на одном растении. Например, кукуруза – мужские цветки собраны на верхушке метелкой, а женские — на стволе початками. Опыление у кукурузы происходит при помощи ветра.
Томаты, баклажаны и многие зерновые культуры относятся к самоопыляемым растениям. Например, цветки томата имеют и пестики, и тычинки. Тычинки срослись так, что в большинстве случаев пестик оплодотворяется собственной пыльцой (факультативное самоопыление).
Двудомные растения устроены так, что на мужских растениях имеются только мужские цветки с тычинками и пыльцой, а на женских – женские цветки с пестиками. Если перенос пыльцы осуществляется между цветками разных особей, такое опыление называется перекрестным. Перекрестное опыление обусловливает обмен генами, поддерживает высокий уровень генетического разнообразия популяций и создает широкое поле для естественного отбора. Для перекрестного опыления, как правило, требуется участие посредника – насекомого, животного, человека. Поэтому выращивая двудомные растения в условиях закрытого грунта, придется процесс опыления взять на себя.
Если перенос пыльцы осуществляется между цветками разных особей, такое опыление называется перекрестным. Перекрестное опыление обусловливает обмен генами, поддерживает высокий уровень генетического разнообразия популяций и создает широкое поле для естественного отбора. Для перекрестного опыления, как правило, требуется участие посредника – насекомого, животного, человека. Поэтому выращивая двудомные растения в условиях закрытого грунта, придется процесс опыления взять на себя.
Искусственное опыление растений в условиях закрытого грунта
Шаг 1. Сбор пыльцы
Если вы разводите растения для себя, в небольших масштабах, то для получения семян будет достаточно пыльцы с одной веточки мужского растения. Во избежание случайного или преждевременного опыления изолируйте мужское растение как можно скорее при появлении на нем пыльника.
Перед сбором пыльцы наденьте на ветку чистый пакет (бумажный или целлофановый). Перевяжите пакет так, чтобы он плотно прилегал к ветке, и пыльца не выпадала из него. Оставьте пакет в таком состоянии на нескольких дней. Перед снятием пакета, когда пыльцы будет собрано достаточно, легонько потрясите веточку, тогда все остатки пыльцы опадут в пакет. Аккуратно срежьте ветку и только после этого снимите пакет.
Перевяжите пакет так, чтобы он плотно прилегал к ветке, и пыльца не выпадала из него. Оставьте пакет в таком состоянии на нескольких дней. Перед снятием пакета, когда пыльцы будет собрано достаточно, легонько потрясите веточку, тогда все остатки пыльцы опадут в пакет. Аккуратно срежьте ветку и только после этого снимите пакет.
Шаг 2. Хранение пыльцы
Если опыление женских растений будет производиться не сразу, то пыльцу надо умело сохранить, т. к. в естественных условиях она хранится недолго. Ее легко уничтожить высокой температурой и влажностью. Но в морозилке пыльца может храниться несколько месяцев. Поэтому собранную пыльцу аккуратно достаем из пакета и высыпаем на бумагу. Для удаления растительных остатков можно просеять пыльцу через сито (под сито также кладется бумага). Затем предварительно стерилизованным скребком собираем пыльцу с бумаги, помещаем ее в стерильную емкость и убираем в морозилку.
Шаг 3. Процесс опыления
Процесс опыления
Если процесс опыления производится сразу после сбора пыльцы, то можно просто надеть пакет с пыльцой на женское растение и слегка потрясти. Для надежности результата можно оставить пакет на женском растении на сутки. Убедитесь, что при опылении нет сквозняков, и пыльца не рассыпалась. Если в оранжерее есть растения, не предназначенные для опыления – изолируйте их, либо проводите опыление в отдельном месте.
ВАЖНО: перед каждой новой процедурой опыления делайте уборку там, где опыляете. Перед тем, как относить женские растения после оплодотворения обратно в оранжерею, обрызгайте их водичкой, чтобы смыть и уничтожить остатки пыльцы. Это позволит предотвратить опыление остальных растений.
Если процесс опыление производится спустя какое-то время и пыльца хранилась в морозилке, то опылять придется вручную кисточкой. Для этого окунаем кисть в контейнер с пыльцой и аккуратно и легко на¬носим кистью пыльцу на пестики. Здесь следует быть внимательным: пыльца не должна случайно стряхнуться с кисточки. Данный способ всем хорош, однако семян в итоге получается не так много, как в первом случае.
Здесь следует быть внимательным: пыльца не должна случайно стряхнуться с кисточки. Данный способ всем хорош, однако семян в итоге получается не так много, как в первом случае.
Несомненно, закрытый грунт имеет свои особенности и некоторые сложности воспроизводства условий окружающей среды для успешного развития растений. Но
современный уровень науки и знаний позволяют получать отличные урожаи и даже вывести потомство в виде семян.
Предыдущая статья
17 Мая 2016
LED — диодное освещение для растений: преимущества и недостатки.
Следующая статья
22 Июня 2016
Торф. Использование в качестве субстрата.
Комментарии
Чтобы оставлять комментарии вам необходимо войти под своим аккаунтом. Если вы еще
не
зарегистрированы, то можете пройти регистрацию, которая займет
всего пару минут.
Стань первым, кто оставил комментарий к этой статье
Знания
Стань продвинутым гровером.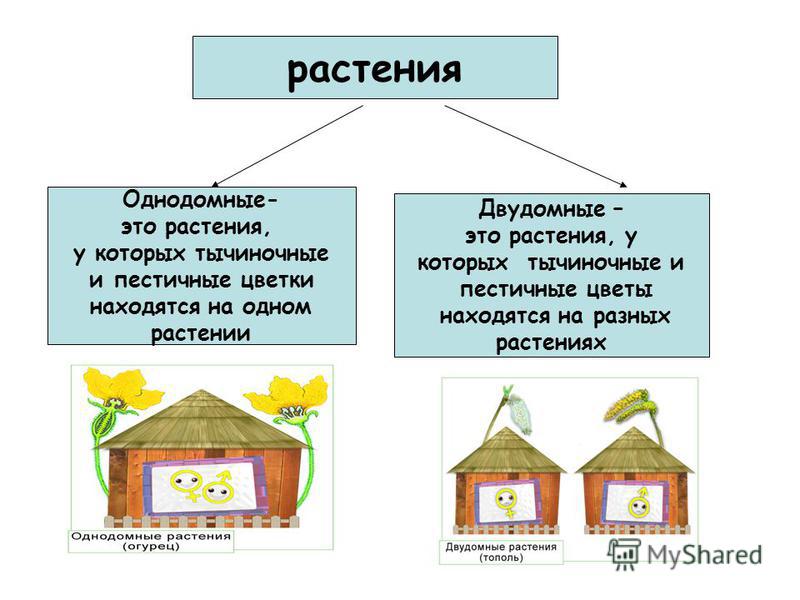 Получай полезные статьи раз в две недели!
Получай полезные статьи раз в две недели!
Я согласен на обработку персональных данных, а также с условиями подписки
Знания
Стань продвинутым гровером. Получай полезные статьи раз в две недели!
Я согласен на обработку персональных данных, а также с условиями подписки
© 2013 — 2022 ИП Ежов А.А.
Все права защищены.
Цветение кукурузы
Рис. 1. Растения кукурузы с недавно появившимися кистями и колосом на поле в округе Кодингтон, Южная Дакота.
У кукурузы цветение определяется по развитию метелки (самец) и колоса (самка). В отличие от большинства других полевых культур, кукуруза является однодомным растением, то есть двумя отдельными цветками на одном растении. Роль кисточки состоит в том, чтобы производить достаточное количество пыльцы для оплодотворения семяпочек в женских цветках или колосе. Полностью выросшая здоровая кисточка может производить от двух до пяти миллионов пыльцевых зерен. Например, если есть 1000 шелковых колосков, это будет от 2000 до 5000 пыльцевых зерен, произведенных для оплодотворения каждого колоска. Обычно кончик кисточки можно увидеть примерно в то же время, когда виден кончик формирующегося уха. Выпадение пыльцы (с кисточки) начинается примерно за один или два дня до появления первого шелка. Выпадение пыльцы продолжалось в течение нескольких дней — около недели с пиком производства примерно на третий день. Опыление кукурузы — это энергичный процесс, и здоровое растение гарантирует наличие пыльцы на каждом появившемся шелке. Успешное опыление приводит к успешному оплодотворению и оптимальному развитию ядра.
Полностью выросшая здоровая кисточка может производить от двух до пяти миллионов пыльцевых зерен. Например, если есть 1000 шелковых колосков, это будет от 2000 до 5000 пыльцевых зерен, произведенных для оплодотворения каждого колоска. Обычно кончик кисточки можно увидеть примерно в то же время, когда виден кончик формирующегося уха. Выпадение пыльцы (с кисточки) начинается примерно за один или два дня до появления первого шелка. Выпадение пыльцы продолжалось в течение нескольких дней — около недели с пиком производства примерно на третий день. Опыление кукурузы — это энергичный процесс, и здоровое растение гарантирует наличие пыльцы на каждом появившемся шелке. Успешное опыление приводит к успешному оплодотворению и оптимальному развитию ядра.
Опыление кукурузы и погода
Хотя может показаться, что процесс опыления зависит от погодных условий, в большинстве случаев он очень хорошо справляется с неблагоприятными ситуациями. Пыльца не так легко смывается с кисточки, потому что при чрезмерно высокой влажности она почти не высыпается. Пыльца, попавшая на шелк, также имеет тенденцию довольно легко противостоять погоде, потому что поверхность шелка липкая, и прорастание происходит сразу после попадания пыльцы.
Пыльца, попавшая на шелк, также имеет тенденцию довольно легко противостоять погоде, потому что поверхность шелка липкая, и прорастание происходит сразу после попадания пыльцы.
В растущей среде тепловой и водный стресс обычно могут идти рука об руку. Стресс из-за дефицита воды обычно оказывает большее влияние на опыление кукурузы, чем тепловой стресс. Тепловой стресс может не проявляться до тех пор, пока температура не превысит 86°F при сухой почве или 92°F при достаточной влажности почвы и высокой влажности.
В условиях высокой температуры растениям кукурузы требуется больше энергии для поддержания нормальных физиологических процессов. Стресс от высокой температуры и/или низкой влажности может привести к высыханию открытых шелковых тканей и неправильному приему пыльцы. Это может уменьшить количество потенциальных ядер в строке. Когда присутствует только высокая температура, растение может получить доступ к воде из более глубокого профиля почвы, и в этом случае прямое воздействие тепла, вероятно, будет низким.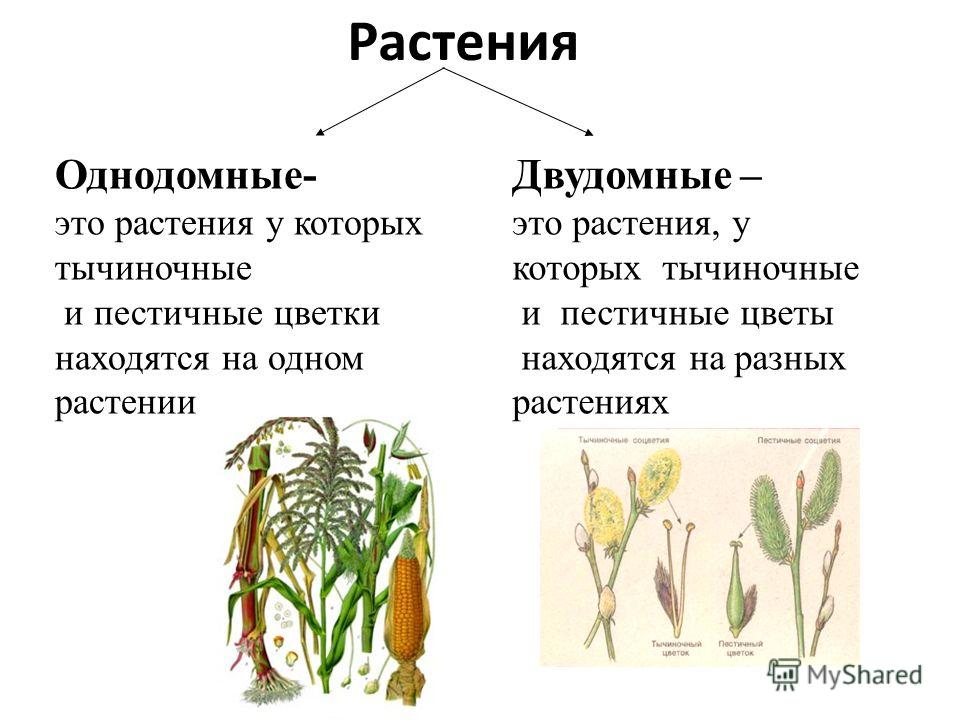
Если присутствует только водный стресс, появление шелка и удлинение колоса происходит медленнее, чем скорость сбрасывания пыльцы, что влияет на количество потенциальных зерен в ряду. Это может привести к ошибке Earl с отсутствующими ядрами. Продолжительные засушливые условия также могут повлиять на развитие семян после опыления.
Рекомендуемое чтение:
- Хоефт, Р.Г., Э.Д. Нафзигер, Р.Р. Джонсон и С.Р. Олдрич. Как растет кукуруза. В Современное производство кукурузы и сои. 2000. Публикация MCSP, 1520 Yorkshire Dr., Шампейн, Иллинойс.
- Coulter, J. 2018. Сухие условия во время опыления кукурузы.
МикроРНК, детерминация пола и меристемы цветков кукурузы | Биология генома
- Миниобзор
- Опубликовано:
- Джо Энн Бэнкс 1
Биология генома
том 9 , Номер статьи: 204 (2008)
Процитировать эту статью
8318 Доступ
22 цитаты
Сведения о показателях
Аннотация
Определение пола в цветках кукурузы включает прерывание развития тычинок или пестика. Недавняя работа по изучению генов, контролирующих этот процесс, показала, что микроРНК участвует как в определении пола мужского соцветия, так и в модели его роста.
Недавняя работа по изучению генов, контролирующих этот процесс, показала, что микроРНК участвует как в определении пола мужского соцветия, так и в модели его роста.
Решение развиваться как мужчина или женщина является фундаментальным для большинства форм жизни. У животных генетические и экологические сигналы, влияющие на это решение, разнообразны и являются предметом интенсивного изучения, однако у растений этому вопросу уделялось относительно мало внимания. Почему разница? В то время как однополость распространена у животных, она не так распространена у цветковых растений. Модельный завод Arabidopsis thaliana , например, имеет совершенные (обеполые) цветки с тычинками, производящими пыльцу, и пестиками, образующими семяпочки, завернутыми в мутовки лепестков и чашелистиков. Хотя у нас есть четкое представление о том, как у Arabidopsis контролируется идентичность цветочных органов, это растение имеет ограниченную ценность для понимания того, как виды растений достигли однополости. По большей части цветковые растения с обоеполыми цветками коллективно развили множество дополнительных и интересных механизмов, позволяющих избежать самооплодотворения, чего большинство видов животных избегает, следя за тем, чтобы каждая особь была однополой, а популяции имели равные пропорции мужских и женских особей.
По большей части цветковые растения с обоеполыми цветками коллективно развили множество дополнительных и интересных механизмов, позволяющих избежать самооплодотворения, чего большинство видов животных избегает, следя за тем, чтобы каждая особь была однополой, а популяции имели равные пропорции мужских и женских особей.
Из тех видов цветковых растений, которые дают однополые цветки, некоторые являются раздельнополыми, имеют исключительно мужские или женские цветки и однополые особи, тогда как другие являются однодомными и дают как однополые мужские, так и женские цветки, но на одном и том же растении. Кукуруза ( Zea mays ) — однодомное растение, которое дает несовершенные (однополые) мужские цветки или соцветия в метелке и несовершенные женские соцветия в колосе. У кукурузы несовершенные соцветия возникают из-за остановки развития тычинок в женских цветках и аборта зачатков пестика в мужских, что позволяет предположить, что детерминация пола не является проблемой идентичности цветочных органов, а скорее вопросом того, что регулирует остановку развития цветков. развитие тычинок или пестиков в колосе и кистевидных соцветиях соответственно. У кукурузы применение растительного гормона гиббереллина (GA) к метелкам делает метелку женственной, тогда как истощение эндогенного GA делает початки маскулинизированными [1], указывая на то, что развитие тычинок и пестика находится под гормональным контролем. Учитывая богатую историю генетики кукурузы и агрономическое значение ее семян, кукуруза является отличной и важной системой для понимания того, как однополость может развиваться у цветковых растений.
развитие тычинок или пестиков в колосе и кистевидных соцветиях соответственно. У кукурузы применение растительного гормона гиббереллина (GA) к метелкам делает метелку женственной, тогда как истощение эндогенного GA делает початки маскулинизированными [1], указывая на то, что развитие тычинок и пестика находится под гормональным контролем. Учитывая богатую историю генетики кукурузы и агрономическое значение ее семян, кукуруза является отличной и важной системой для понимания того, как однополость может развиваться у цветковых растений.
Мутации, делающие соцветия метелки женскими, можно легко идентифицировать по наличию семян и пушистых шелков (женских рыльцов) в метелке мужского растения (рис. 1а). Шесть таких мутантов tasselseed были идентифицированы более 50 лет назад [2], и два из них, ts4 (рецессивная мутация) и Ts6 (доминантная мутация), были недавно клонированы и охарактеризованы Chuck et al . [3]. Что особенно важно в этом недавнем исследовании, так это то, что оно показывает, как регулируется детерминация пола (в данном случае выкидыш пестика) и как этот процесс связан с механизмом, который регулирует, когда меристема переключается с индетерминантной на детерминантную, или конечную. государство. Понимание этой связи в первую очередь требует базового понимания структуры и развития кисточки.
государство. Понимание этой связи в первую очередь требует базового понимания структуры и развития кисточки.
Рисунок 1
Организация мужских соцветий и цветков кукурузы. (а) Соцветия (метелки) растений дикого типа (слева) и ts4 (справа). Нормальная мужская кисточка состоит из ветвей (шипов), каждая из которых состоит из колосков, содержащих по два цветка. Нити в мутанте ts4 — это женские стигмы. (b) Схематическое изображение двух мужских колосков, каждый из которых состоит из двух цветков и окружен двумя прицветниками, называемыми колосками. Каждый цветочек состоит из трех тычинок, окруженных двумя прицветниками, цветковой чешуей и цветком (лодикулы здесь не показаны). Фотографии и иллюстрации предоставлены Джорджем Чаком.
Изображение в полный размер
Архитектура кукурузной соцветия довольно сложна (рис. 1а), даже несмотря на то, что сотни отдельных соцветий соцветий чрезвычайно уменьшены в размерах и просты по структуре, как показано на рис. 1б. Каждый мужской цветочек состоит из двух прицветников (цветковая чешуя и цветковая чешуя), двух мелких чешуек, называемых лодикулами, и трех тычинок. Соцветия организованы в единицы, называемые колосками, при этом каждый колосок окружен двумя прицветниками, называемыми колосками. У кукурузы в каждом мужском колоске по два цветка, а в колосе — по одному. Колоски коллективно организованы в единицы, называемые шипами, при этом множество шипов вместе составляют кисточку. Организация колоса, колосков и цветков определяется апикальной меристемой соцветия, которая содержит популяцию стволовых клеток, доставляющих новые дочерние клетки растущему растению. По мере развития колосовидного соцветия меристема соцветия непрерывно образует новые производные ветвей, которые инициируют парные меристемы колосков; после ветвления каждая меристема пары колосков претерпевает переход с образованием двух меристем колосков; затем каждая меристема колоска претерпевает переход с образованием двух меристем цветка, которые инициируют органы цветка.
1б. Каждый мужской цветочек состоит из двух прицветников (цветковая чешуя и цветковая чешуя), двух мелких чешуек, называемых лодикулами, и трех тычинок. Соцветия организованы в единицы, называемые колосками, при этом каждый колосок окружен двумя прицветниками, называемыми колосками. У кукурузы в каждом мужском колоске по два цветка, а в колосе — по одному. Колоски коллективно организованы в единицы, называемые шипами, при этом множество шипов вместе составляют кисточку. Организация колоса, колосков и цветков определяется апикальной меристемой соцветия, которая содержит популяцию стволовых клеток, доставляющих новые дочерние клетки растущему растению. По мере развития колосовидного соцветия меристема соцветия непрерывно образует новые производные ветвей, которые инициируют парные меристемы колосков; после ветвления каждая меристема пары колосков претерпевает переход с образованием двух меристем колосков; затем каждая меристема колоска претерпевает переход с образованием двух меристем цветка, которые инициируют органы цветка. В конце концов все меристемы заканчиваются, цветки созревают, раскрываются и сбрасывают пыльцу.
В конце концов все меристемы заканчиваются, цветки созревают, раскрываются и сбрасывают пыльцу.
Кисти мутантных растений ts4 отличаются от растений дикого типа по нескольким параметрам (см. рис. 1а): они имеют феминизированные соцветия, демонстрируют усиленное ветвление колосков и обычно дают более двух соцветий на колосок. Увеличение ветвления и числа цветков отражает неспособность меристем переходить из индетерминантного состояния в детерминантное по мере развития метелки. Таким образом, в дополнение к стимуляции развития самцов ген дикого типа ts4 функционирует, придавая детерминированность меристемам колосков и пар колосков.
Чтобы понять, как эти гены кистевидного семени контролируют определение пола в мужских цветках, Чак и его коллеги [3] клонировали ген ts4 и обнаружили, что он является членом семейства микроРНК (миРНК) miR172, которые, как известно, регулируют APETALA2 ( AP2 ) семейство транскрипционных факторов [4, 5].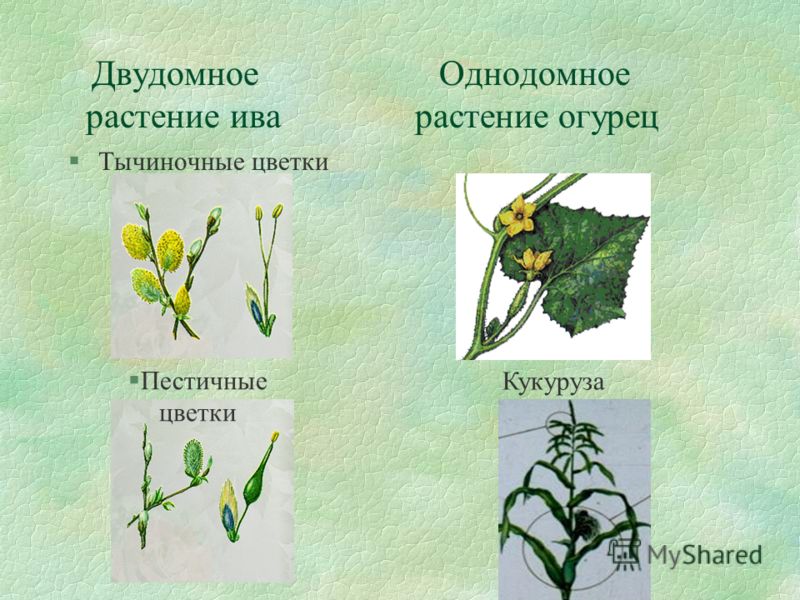 Как оказалось, те же авторы ранее показали, что ген индетерминантного колоска1 ( ids1 ) кукурузы кодирует AP2 -подобный ген [6]. Как и кисточки ts4 , кисточки ids1 производят дополнительные соцветия; однако цветков ids1 в кисточке не феминизированы, как цветочки ts4 .
Как оказалось, те же авторы ранее показали, что ген индетерминантного колоска1 ( ids1 ) кукурузы кодирует AP2 -подобный ген [6]. Как и кисточки ts4 , кисточки ids1 производят дополнительные соцветия; однако цветков ids1 в кисточке не феминизированы, как цветочки ts4 .
Чтобы узнать, является ли ids1 целью ts4 , Chuck и др. . [3] исследовали фенотипы двойных мутантов ts4 ids1 и обнаружили, что ids1 подавляли дефект определения пола у ts4 , подтверждая предположение, что ids1 является целью ts4 . В другой серии интересных экспериментов авторы также обнаружили, что Ts6 и ids1 являются вариантами одного и того же гена, хотя эти два мутанта имеют разные фенотипы (кисточковые соцветия растений Ts6 часто феминизированы [7], тогда как ids1 цветков нет). Сравнение последовательностей Ts6 и ids1 показало, что две независимые мутации Ts6 изменяют 5′-конец сайта связывания miR172 в этом AP2 -подобный ген, приводящий к доминантному фенотипу, тогда как два независимых мутанта ids1 вызываются вставками транспозона Mutator в разные интроны одного и того же гена, что приводит к фенотипу с потерей функции.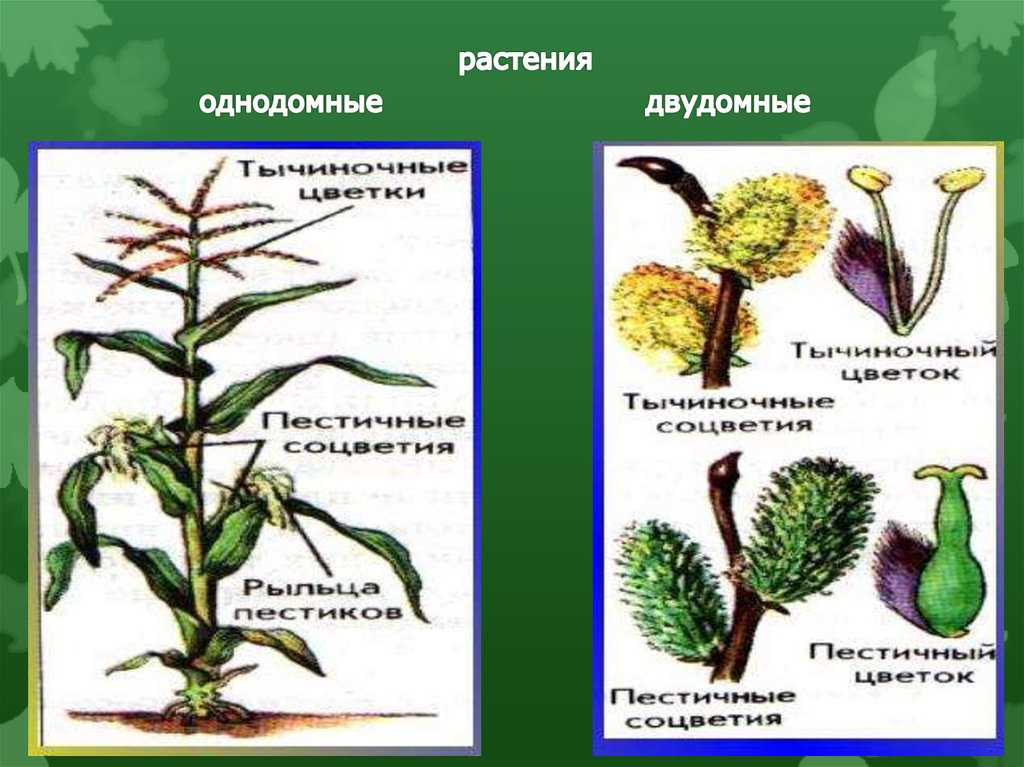 Поскольку транскриптов ids1 присутствуют как в растениях дикого типа, так и в мутантных растениях ts4 , ts4 , вероятно, регулируют синтез белка IDS1. Трансляционная репрессия белка AP2 с помощью miR172 также наблюдалась у Arabidopsis [5]. Иммунолокализация дикого типа и мутантных метелок ts4 показала, что IDS1 эктопически экспрессируется в пестиках мутантных ts4 соцветий и повышается во время инициации цветочной меристемы. Эти наблюдения привели к созданию простой модели регуляции определения пола с помощью IDS1 в кистевидных соцветиях кукурузы, показанной на рисунке 2.
Поскольку транскриптов ids1 присутствуют как в растениях дикого типа, так и в мутантных растениях ts4 , ts4 , вероятно, регулируют синтез белка IDS1. Трансляционная репрессия белка AP2 с помощью miR172 также наблюдалась у Arabidopsis [5]. Иммунолокализация дикого типа и мутантных метелок ts4 показала, что IDS1 эктопически экспрессируется в пестиках мутантных ts4 соцветий и повышается во время инициации цветочной меристемы. Эти наблюдения привели к созданию простой модели регуляции определения пола с помощью IDS1 в кистевидных соцветиях кукурузы, показанной на рисунке 2.
Рисунок 2
0067 IDS1 стенограмма. В нормальных мужских цветках мРНК IDS1 не транслируется из-за присутствия миРНК ts4 (миРНК172). Если белок IDS1 продуцируется либо в результате мутации с потерей функции в гене ts4 , либо в результате мутации сайта связывания микроРНК в ids1 (которая произошла у мутанта Ts6 ), цветочек женский.
Полноразмерное изображение
Неясно, как ids1 влияет на ветвление и переходы меристемы, хотя некоторые подсказки дает характеристика мутации ap2 в Arabidopsis . У Arabidopsis эктопическая экспрессия мутантной формы ap2 с измененным сайтом связывания miRNA172 приводит к нескольким дефектам цветков, включая потерю детерминантности цветков [5]. Этот фенотип имитирует агамный фенотип ( ag ), что согласуется с функцией AG , необходимой для инициации плодолистика [8]. Чак и др. . [3] предполагают, что аналогичная связь между ids1 и AG -подобный ген могут встречаться в кукурузе. Снижая экспрессию AG в цветке, пролонгированная экспрессия ids1 будет предотвращать дифференцировку стволовых клеток в меристеме, тем самым позволяя меристеме колоска образовывать дополнительные меристемы цветка. Эту и другие гипотезы еще предстоит проверить.
С точки зрения эволюции было бы интересно посмотреть, работает ли механизм определения пола у кукурузы, определяемый ts4 и ids1 является общим для других однодомных или раздельнополых видов растений, включая односпоровые папоротники, половые фенотипы которых определяются в гаметофитной фазе роста и у которых развитие меристемы и детерминация пола идут рука об руку [9] .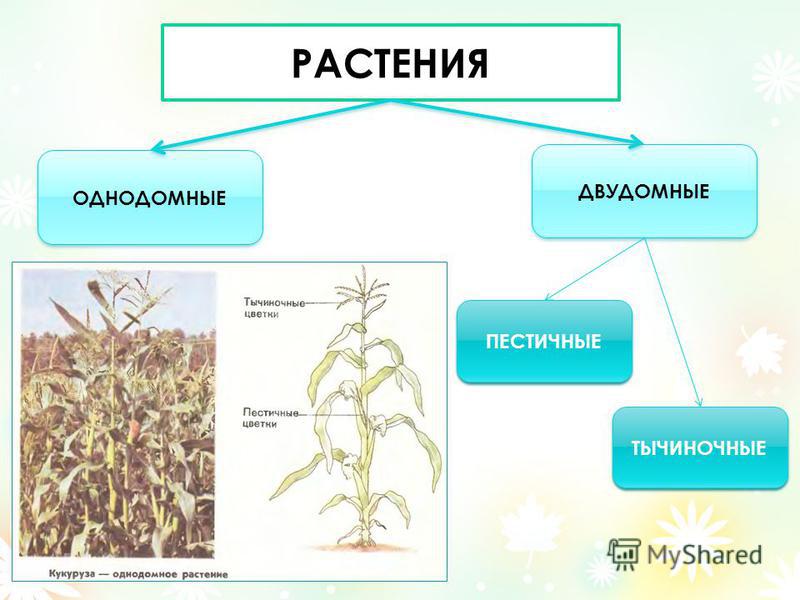 Учитывая, что однодомность эволюционировала несколько раз в разных линиях покрытосеменных растений, а за ней часто следует двудомность [10], исследование Chuck et al . [3] предоставляет новые инструменты для понимания того, насколько широко распространен этот механизм определения пола.
Учитывая, что однодомность эволюционировала несколько раз в разных линиях покрытосеменных растений, а за ней часто следует двудомность [10], исследование Chuck et al . [3] предоставляет новые инструменты для понимания того, насколько широко распространен этот механизм определения пола.
Ссылки
Nickerson NH: Продолжительное лечение гиббереллиновой кислотой пяти различных видов кукурузы. Энн Мо Бот Гард. 1959, 46: 19-37. 10.2307/2394566.
Артикул
КАСGoogle ученый
Вейт Б., Шмидт Р.Дж., Хейк С., Янофски М.Ф.: Развитие цветков кукурузы: новые гены и старые мутанты. Растительная клетка. 1993, 5: 1205-1215. 10.1105/тпк.5.10.1205.
Артикул
пабмед
ПабМед ЦентральныйGoogle ученый
Chuck G, Meeley R, Irish E, Sakai H, Hake S: МикроРНК tasselseed4 кукурузы контролирует детерминацию пола и судьбу клеток меристемы, воздействуя на Tasselseed6/неопределенный колосок1.
 Нат Жене. 2007, 39: 1517-1521. 10.1038/нг.2007.20.
Нат Жене. 2007, 39: 1517-1521. 10.1038/нг.2007.20.Артикул
пабмед
КАСGoogle ученый
Aukerman MJ, Sakai H: Регуляция времени цветения и идентичности цветочных органов с помощью микроРНК и ее APETALA2-подобных генов-мишеней. Растительная клетка. 2003, 15: 2730-2741. 10.1105/тпк.016238.
Артикул
пабмед
КАС
ПабМед ЦентральныйGoogle ученый
Chen X: МикроРНК в качестве репрессора трансляции APETALA2 в развитии цветка Arabidopsis . Наука. 2004, 303: 2022-2025. 10.1126/научн.1088060.
Артикул
пабмед
КАСGoogle ученый
Чак Г., Мили Р.Б., Хейк С.: Контроль судьбы меристемы колосков кукурузы с помощью APETALA2-подобного гена неопределенного колоска1. Гены Дев. 1998, 12: 1145-1154.
Артикул
пабмед
КАС
ПабМед ЦентральныйGoogle ученый
 Нат Жене. 2007, 39: 1517-1521. 10.1038/нг.2007.20.
Нат Жене. 2007, 39: 1517-1521. 10.1038/нг.2007.20.