
Содержание
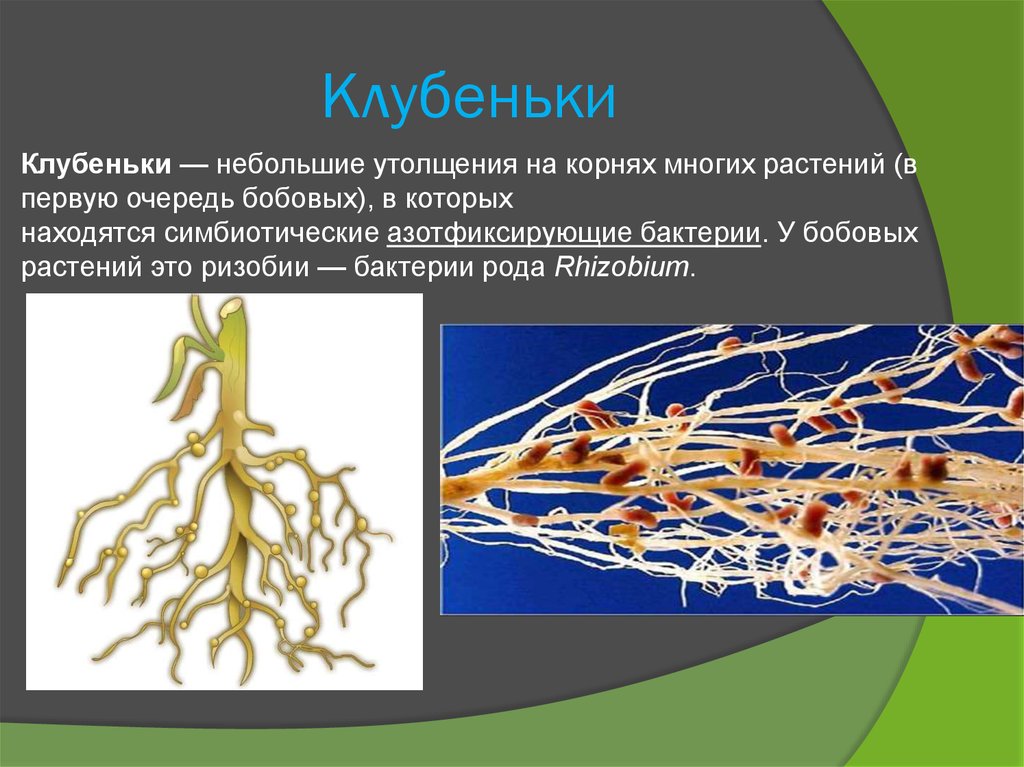
клубеньковые бактерии
Клубеньковые бактерии специфичны — отдельные виды или расы их способны образовать клубеньки на корнях лишь определенных бобовых. Так, одни из них развиваются только на корнях клевера, но не могут заражать корни гороха, люцерны, люпина и других бобовых. Группы бактерий, которые образуют клубеньки на корнях люпина и сераделлы, не заражают корни клевера и гороха и т. д. Иногда специфичность клубеньковых бактерий настолько сильно выражена, что различные разновидности од но Г? и той же культуры (или даже сорта растений) по-разному относятся к тому или другому штамму бактерии. Например, корни желтого кормового люпина не всегда хорошо заражаются клубеньковыми бактериями из корней однолетних горьких люпинов. Группы клубеньковых бактерий по их специфичности указаны на странице 382.[ …]
Клубеньковые бактерии наиболее интенсивно развиваются при реакции почвы, близкой к нейтральной. Поэтому при посевах бобовых на кислых почвах наряду с инокуляцией семян необходимо известкование почвы. Инокуляция без известкования оказывает очень слабое влияние на урожай и содержание белка. По данным К. Филлерса, при выращивании сои на кислой почве без инокуляции и без внесения извести содержание белка в семенах составляло 32,8%, при инокуляции содержание белка увеличилось на 1,2%, а при нейтрализации кислотности почвы и инокуляции содержание белка в семенах возросло на 11,2%.[ …]
Инокуляция без известкования оказывает очень слабое влияние на урожай и содержание белка. По данным К. Филлерса, при выращивании сои на кислой почве без инокуляции и без внесения извести содержание белка в семенах составляло 32,8%, при инокуляции содержание белка увеличилось на 1,2%, а при нейтрализации кислотности почвы и инокуляции содержание белка в семенах возросло на 11,2%.[ …]
Клубеньковые бактерии различных бобовых растений погибают в кислых почвах.[ …]
Клубеньковые бактерии нуждаются в достаточном притоке к ним углеводов, фосфора и кальция. В последние годы установлено, что и ряд микроэлементов (особенно молибден) играет важную роль в жизнедеятельности клубенько вых бактерий. Известкование и применение молибдена под бобовые на кислых почвах регулируют реакцию почвы; достаточная обеспеченность бобовых фосфором нужна на всех почвах.[ …]
Прокариоты (бактерии, архебактерии, цианобактерии) — одноклеточные организмы, не имеют ядра. Благодаря такому разнообразному метаболизму бактерии могут существовать в самых различных условиях среды: в воде, воздухе, почве, живых организмах. Велика роль бактерий в образовании нефти, каменного угля, торфа, природного газа, в почвообразовании, в круговоротах азота, фосфора, серы и других элементов в природе. Сапротрофные бактерии участвуют в разложении органических останков растений и животных и в их минерализации до С02, Н20, Н28, 1ЧН3 и других неорганических веществ. Вместе с грибами они являются редуцентами. Клубеньковые бактерии (азотфикси-рующие) образуют симбиоз с бобовыми растениями и участвуют в фиксации атмосферного азота в минеральные соединения, доступные растениям. Сами растения такой способностью не обладают.[ …]
Велика роль бактерий в образовании нефти, каменного угля, торфа, природного газа, в почвообразовании, в круговоротах азота, фосфора, серы и других элементов в природе. Сапротрофные бактерии участвуют в разложении органических останков растений и животных и в их минерализации до С02, Н20, Н28, 1ЧН3 и других неорганических веществ. Вместе с грибами они являются редуцентами. Клубеньковые бактерии (азотфикси-рующие) образуют симбиоз с бобовыми растениями и участвуют в фиксации атмосферного азота в минеральные соединения, доступные растениям. Сами растения такой способностью не обладают.[ …]
При отсутствии клубеньковых бактерий на корнях бобовых культур они становятся такими же потребителями азота из почвы, как и другие растения.[ …]
Важным свойством клубеньковых бактерий является их активность (эффективность), т. е. способность в симбиозе с бобовыми растениями ассимилировать молекулярный азот и удовлетворять в нем потребности растения-хозяина. В зависимости от того, в какой степени клубеньковые бактерии способствуют повышению урожайности бобовых культур (рис.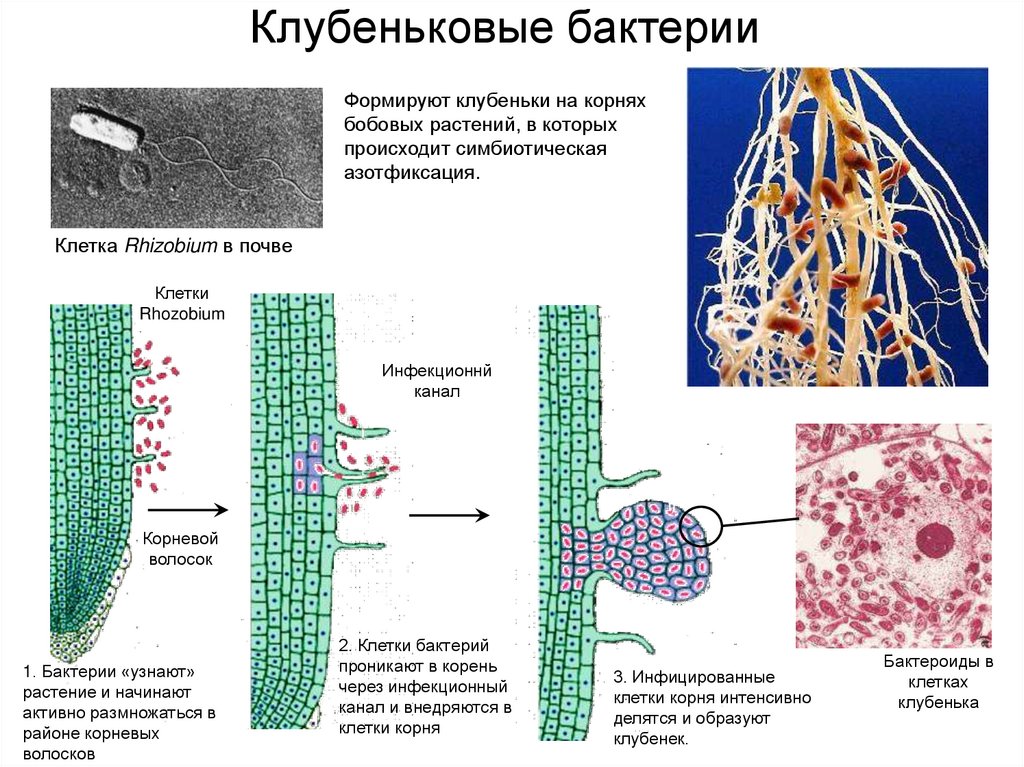 146), их принято делить на активные (эффективные), малоактивные (малоэффективные) и неактивные (неэффективные).[ …]
146), их принято делить на активные (эффективные), малоактивные (малоэффективные) и неактивные (неэффективные).[ …]
Среди азотфиксирующих бактерий выделяют свободноживу-щих в почве и клубеньковых, живущих на корнях бобовых растений (рис. 9, з). Наиболее важными представителями свобод-ноживущих азотфиксирующих бактерий являются Azotobacter и Clostridium, связывающие за год несколько десятков килограммов азота на 1 га почвы. Значительно более эффективна деятельность клубеньковых бактерий, заражающих клетки корней бобовых. В результате под бобовыми происходит микробиологическое накопление доступного для растений азота. Под площадью в 1 га, засеянной клевером, в результате действия этих бактерий может быть накоплено в 100 раз больше азота, чем свободноживущими фиксаторами этого элемента.[ …]
Поскольку размножение клубеньковых бактерий в отсутствие влаги не происходит, в случае засушливой весны инокулированные (искусственно зараженные) семена необходимо вносить глубже в почву.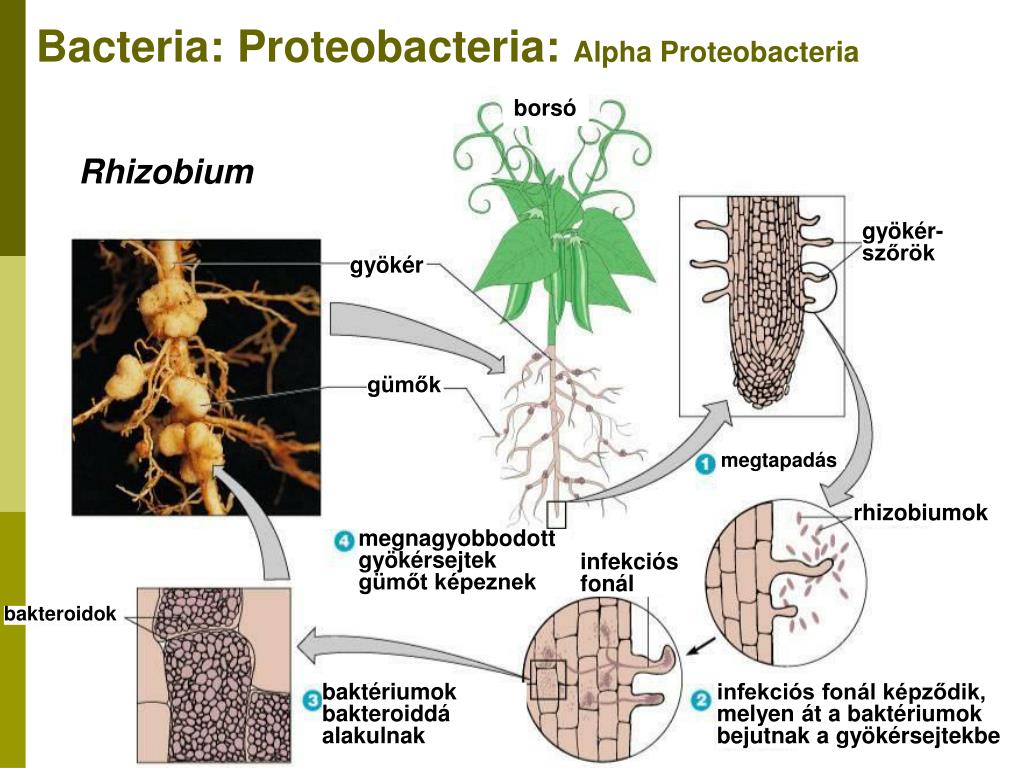 Например, в Австралии семена с нанесенными на них клубеньковыми бактериями глубоко заделывают в почву. Интересно, что клубеньковые бактерии почв засушливого климата более стойко переносят засуху, чем бактерии почв влажного климата. В этом проявляется их экологическая приспособленность.[ …]
Например, в Австралии семена с нанесенными на них клубеньковыми бактериями глубоко заделывают в почву. Интересно, что клубеньковые бактерии почв засушливого климата более стойко переносят засуху, чем бактерии почв влажного климата. В этом проявляется их экологическая приспособленность.[ …]
Кроме специфичности, расы клубеньковых бактерий различаются по вирулентности и активности. Вирулентность — способность их проникать через корневые волоски в корень бобового растения и образовывать клубеньки. Активностью клубеньковых бактерий называют способность их к усвоению азота атмосферы. Только активные штаммы этих бактерий снабжают бобовые растения азотом. При заражении корней вирулентными, но неактивными клубеньковыми бактериями образуются клубеньки, но фиксации азота не происходит. Клубеньковые бактерии, используемые для приготовления нитрагина, должны обладать большой вирулентностью и высокой активностью. Если вирулентность клубеньковых бактерий нитрагина выше вирулентности уже находящихся в почве менее активных бактерий, то это позволяет клубеньковым бактериям нитрагина проникать в корень быстрее и в большом количестве. [ …]
[ …]
Из всех этих примеров симбиоз »клубеньковых бактерий с бобовыми изучен наиболее тщательно, поскольку эти растения имеют для человека огромное значение.[ …]
Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение pH среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям pH, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация микроорганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями pH. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (pH 4,0— 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. [ …]
[ …]
Существует большое количество видов и рас клубеньковых бактерий, каждая из которых приспособлела к заражению одного или нескольких видов бобовых растений. Корневые системы бобовых растений обладают специфическими корневыми выделениями. Благодаря этому клубеньковые бактерии скопляются вокруг корневых волосков, которые при атом снручиваготся. Через корневой волосок бактерии в виде сплошного тяжа, состоящего из соединенных слизью бесчисленных бактерий, проникают в паренхиму корня. Вовможпо, бактерии выделяют гормон ауксин в именно ато является причиной разрастания тканей, образуются вздутия — клубеньки. Клетки клубеньков заполняются быстро резмножающимися бактериями, но остаются живыми и сохраняют крупные ядра. Клубеньковые бактерии заражают только полиплоидные клетки корня.[ …]
Люцерна как бобовое растение способна с помощью клубеньковых бактерий на корнях накапливать большое количество азота в почве, не уступая в этом отношении клеверу. Корневая система ее лучше развивается в 1-й год пользования, а к 3-му году накапливает азота в почве 120. ..200 кг/га. Повышая плодородие почвы, улучшая ее структуру, люцерна является хорошим предшественником в севообороте.[ …]
..200 кг/га. Повышая плодородие почвы, улучшая ее структуру, люцерна является хорошим предшественником в севообороте.[ …]
При возделывании люпинов для повышения активности клубеньковых бактерий люпина применяют нитрагин, полезны также борные и молибденовые микроудобрения (семена обрабатывают раствором молибденовокислого аммония одновременно с их нитрагинизацией).[ …]
Анаэробный распад целлюлозы осуществляется только бактериями (например, бациллой Омелянского), а аэробный -многими видами бактерий, грибами, актиномицетами.[ …]
Это указывает, что фиксированный меченый азот попадает в тела бактерий из тканей высшего растения, которое является источником азотного питания для бактерий. Таким образом, фиксация атмосферного азота локализована не в теле клубеньковых бактерий, а в клубеньковой ткани высшего растения. Важная роль клубеньковых бактерий заключается в том, что они индуцируют образование этой специфической клубеньковой ткани. Дальнейшие исследования показали, что максимальное содержание меченого азота в отдельных азотистых фракциях клеточного сока клубеньков всегда приходится на амидную группу аспарагина и глутамина. Так как эта группа может рассматриваться как трансформированный аммиак, то именно •аммиак и является конечным неорганическим продуктом биологической фиксации азота.[ …]
Так как эта группа может рассматриваться как трансформированный аммиак, то именно •аммиак и является конечным неорганическим продуктом биологической фиксации азота.[ …]
| Чувствительность неинфицированных (/) и инфицированных (2) клубеньковыми бактериями растений сои и люпина к трефлану (в) и атразину (б) |
В биосфере фиксация азота осуществляется несколькими группами анаэробных бактерий и цианобактерий при нормальных температуре и давлении, благодаря высокой эффективности биокатализа. Считается, что бактерии переводят в связанную форму приблизительно 1 млрд т азота в год (мировой объем промышленной фиксации — около 100 млн т). В клубеньковых бактериях бобовых растений фиксация азота осуществляется с помощью сложного ферментного комплекса, защищенного от избытка кислорода специальным растительным гемоглобином.[ …]
Одной из причин положительного влияния молибдена на фиксацию молекулярного азота клубеньковыми бактериями является повышение под его действием активности также и дегидрогеназ, которые обеспечивают непрерывный приток активированного водорода, необходимого для восстановления атмосферного азота. [ …]
[ …]
Ценность нового препарата нитрагина состоит в длительности сохранения жизнеспособности клубеньковых бактерий и возможности заблаговременного изготовления препаратов механизированным способом. По эффективности сухой нитрагин близок к почвенному. Его можно применять в виде дуста, опудривая семена без их намачивания.[ …]
Особенно значима в круговороте азота роль симбиотических (от греч. симбиоз — сожительство) клубеньковых бактерий, локализующихся на корнях растений преимущественно семейства бобовых. Бактерии родов азотобактер или ризобиум способны путем ферментативного расщепления молекул N3 фиксировать атмосферный азот и делать его доступным корневым системам растений.[ …]
Биологическая фиксация молекулярного азота атмосферы в почве осуществляется двумя группами бактерий: свобод-ноживущими аэробными и анаэробными и клубеньковыми бактериями, живущими в симбиозе с бобовыми растениями. Важнейшим представителем первой группы из аэробов является Azotobacter, а из анаэробных — Clostridium pasteu-rianum. Благоприятная среда для активной деятельности клубеньковых бактерий — хорошо аэрируемые почвы со слабокислой и нейтральной реакцией. Деятельность бактерий азотфиксаторов имеет важное значение в общем балансе азота в земледельчески используемых почвах. Поэтому для деятельности клубеньковых бактерий важно проводить окультуривание почв. Чтобы увеличить численность клубеньковых бактерий, в почву вносят бактериальный препарат нитрагин, содержащий активные расы клубеньковых бактерий.[ …]
Благоприятная среда для активной деятельности клубеньковых бактерий — хорошо аэрируемые почвы со слабокислой и нейтральной реакцией. Деятельность бактерий азотфиксаторов имеет важное значение в общем балансе азота в земледельчески используемых почвах. Поэтому для деятельности клубеньковых бактерий важно проводить окультуривание почв. Чтобы увеличить численность клубеньковых бактерий, в почву вносят бактериальный препарат нитрагин, содержащий активные расы клубеньковых бактерий.[ …]
По данным НИИ сельскохозяйственной микробиологии, в ряде почв соответствующие той или иной бобовой культуре клубеньковые бактерии могут отсутствовать, а те, что имеются, обладают малопродуктивной системой азотфиксации. В связи с этим микробиологи провели селекционную работу. В итоге каждые три года на заводы передаются до десяти новых штаммов клубеньковых бактерий, азотфиксирующая способность которых на 10—20% превышает предыдущие эталонные штаммы. Создан и массово производится препарат ризоторфин — удобная и практичная форма поставки клубеньковых бактерий к семенам и растущим корням бобовых.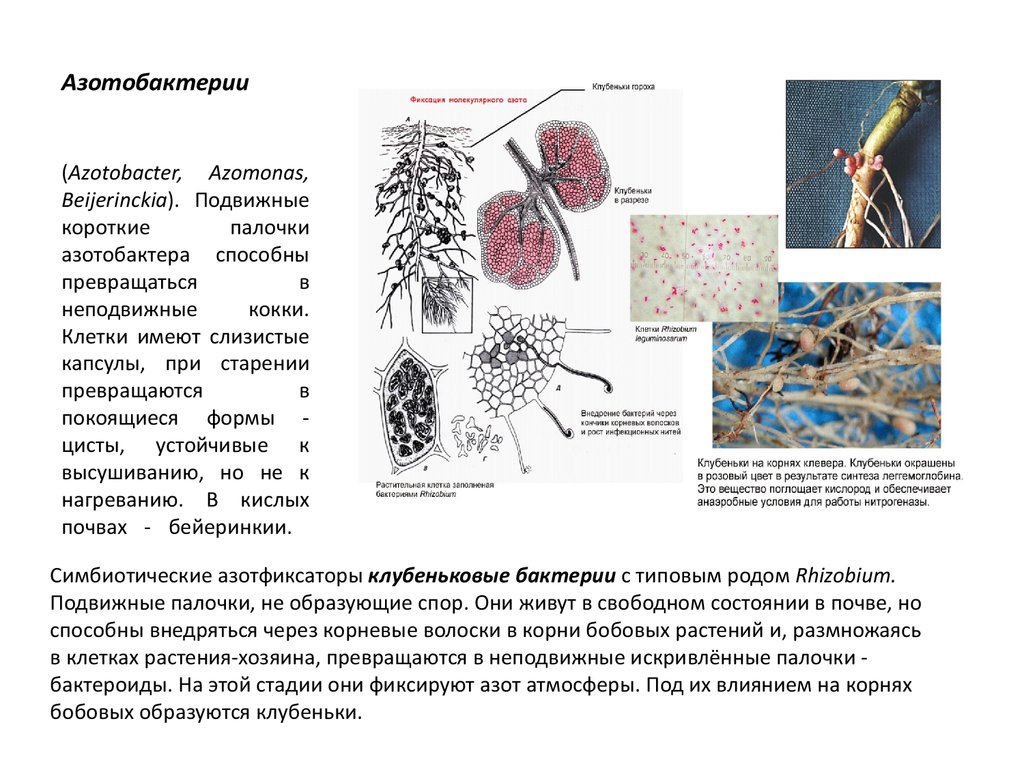 [ …]
[ …]
Органические и минеральные удобрения (фосфорнокалийные), внесенные в почву, значительно улучшают способность клубеньковых бактерий усваивать атмосферный азот. Для активизации деятельности клубеньковых бактерий кислые почвы необходимо известковать, а почвы в засушливых районах — обеспечивать влагой.[ …]
Мутуализм — это взаимодействие между двумя организмами разных видов, которое выгодно для каждого из них. Например, азотфиксирующие клубеньковые бактерии обитают на корнях бобовых растений, конвертируя атмосферный азот в форму, доступную для усвоения этими растениями. Следовательно, бактерии обеспечивают растения азотом. В свою очередь растения обеспечивают клубеньковые бактерии всеми необходимыми питательными веществами. Мутуализмом можно считать также взаимодействие между микроорганизмами, обитающими в толстом отделе кишечника человека, и самим человеком. Для микроорганизмов выгода определяется тем, что они обеспечивают свои питательные потребности за счет содержимого кишечника, а для человека выгода состоит в том, что микроорганизмы осуществляют дополнительное переваривание пищи и еще синтезируют крайне необходимый для него витамин К. В мире цветковых растений мутуализмом является опыление насекомыми растений и питание насекомых нектаром растений. Мутуализм значим и в «переработке» органических веществ. Например, переваривание целлюлозы в желудке (рубце) крупного рогатого скота обеспечивается содержащимися в нем бактериями.[ …]
В мире цветковых растений мутуализмом является опыление насекомыми растений и питание насекомых нектаром растений. Мутуализм значим и в «переработке» органических веществ. Например, переваривание целлюлозы в желудке (рубце) крупного рогатого скота обеспечивается содержащимися в нем бактериями.[ …]
Особенность питания бобовых культур заключается в том, что они не нуждаются в наличии минеральных соединений азота в почве. В симбиозе с клубеньковыми бактериями бобовые растения используют свободный азот атмосферы. Поэтому эти культуры являются источником биологического азота для сельского хозяйства.[ …]
В связи с этим в практику сельского хозяйства прочно вошел прием инокуляции — предпосевная обработка семян бобовых растений препаратом клубеньковых бактерий соответствующего вида. В разных странах технический препарат для инокуляции бобовых растений получил разные наименования. В СССР, ГДР, ФРГ и Польше он назван нитрагином. Отсюда прием инокуляции соответствующих культур в этих странах называется нитраги-низацией. Нитрагин повышает урожай бобовых растений на 10—15%, а в новых районах возделывания — на 50% и более.[ …]
Нитрагин повышает урожай бобовых растений на 10—15%, а в новых районах возделывания — на 50% и более.[ …]
Как было отмечено выше, на кислых почвах клевер растет плохо, часто изреживается, а иногда выпадает совсем и даже в 1-й год пользования. Жизнедеятельность клубеньковых бактерий на таких почвах подавляется. Кислотность почв, особенно в северо-западных областях Нечерноземной зоны (здесь более половины пашни имеет повышенную кислотность), при наличии в них подвижных форм алюминия является одной из причин снижения урожайности трав.[ …]
Фиксация атмосферного азота. Ни одно зеленое растение не может питаться непосредственно азотом атмосферы. Так как в результате деятельности денитрифицирующих бактерий непрерывно идет уменьшение в природе запасов связанного азота и перевод его в атмосферный азот, то жизни начземле грозила бы неминуемая гибель из-за азотного голода. Однако существует группа микроорганизмов, способная связывать атмосферный азот, делая его доступным для растений. Эти микроорганизмы называются азотфиксирующими бактериями, они разделяются на клубеньковые бактерии, развивающиеся на корнях бобовых растений, и на свободно живущие в почве.[ …]
Эти микроорганизмы называются азотфиксирующими бактериями, они разделяются на клубеньковые бактерии, развивающиеся на корнях бобовых растений, и на свободно живущие в почве.[ …]
В результате этих исследований было найдено, что фиксированный бобовыми меченый газообразный азот атмосферы первоначально в больших количествах содержится только в клеточном соке клубеньковой ткани, представляющей гипертрофированную корневую ткань бобовых, откуда он затем постепенно переходит в другие органы растений. В клубеньковых бактериях меченый азот при экспозиции растений от 6 до 48 часов совершенно отсутствует или содержится в крайне незначительных количествах, обычно не выходящих за пределы возможной ошибки эксперимента (табл. 4).[ …]
Кроме того, П. С. Коссовичу принадлежат не потерявшие и до настоящего времени значения исследования круговорота в природе и хозяйстве серы и хлора, а также доказательство положения, что клубеньковые бактерии связывают азот атмосферы, поступивший через корни, а не через листья бобовых растений. Он с успехом изучал и корневые выделения культур, особенно выделение углекислого газа, связывая его с усвояющей способностью корней.[ …]
Он с успехом изучал и корневые выделения культур, особенно выделение углекислого газа, связывая его с усвояющей способностью корней.[ …]
Сообщество организмов, основанное на обоюдной пользе, когда два вида создают друг другу благоприятную среду для развития, называется симбиозом. Примером могут служить взаимоотношения между клубеньковыми бактериями и бобовыми растениями. Клубеньковые бактерии получают от бобового растения безазотис-тые органические вещества и минеральные соли, а взамен предоставляют ему азотистые вещества, синтезированные ими из атмосферного азота.[ …]
К аминоавтотрофам относятся микроорганизмы, использующие азот аммиачных солей, азотнокислых солей и мочевину. Аминоавтотрофы при использовании азота минеральных соединений предварительно переводят его в аммиачный азот, а затем потребляют для построения аминокислот, из которых синтезируют белки. Предварительный перевод азота в аммиак объясняется тем, что в составе микробной клетки азот находится в восстановленном состоянии в форме амино- (Nh3) и иминогрупп (NH).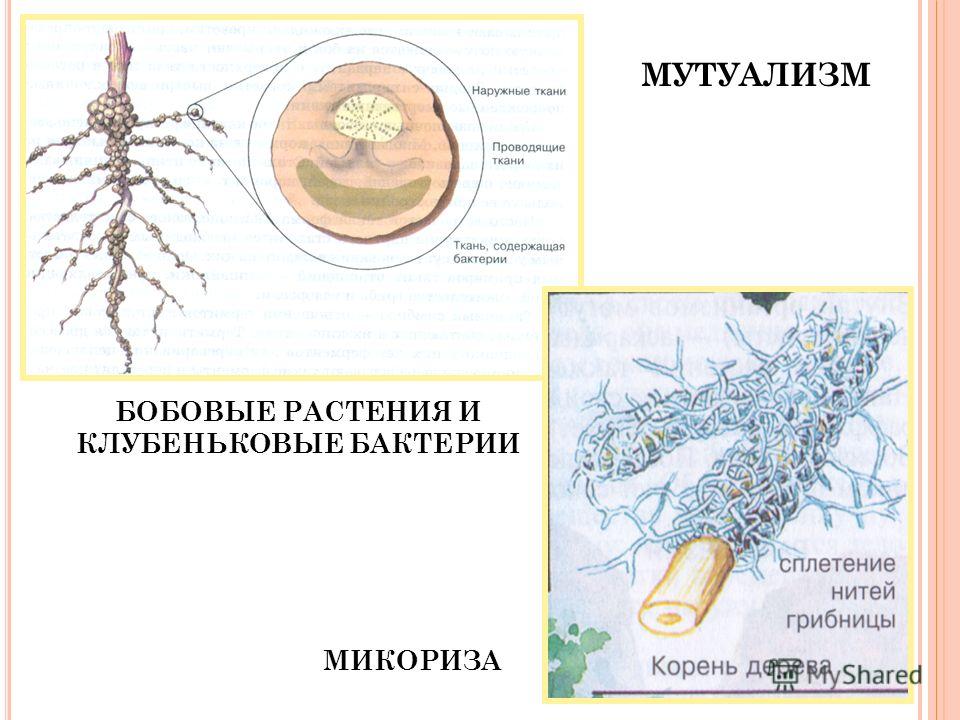 Роль азота в белковых веществах протоплазмы бактерий состоит в том, что он придает белкам реактивность. Азот трудно входит в состав соединений из-за инертности, но легко из них выходит.[ …]
Роль азота в белковых веществах протоплазмы бактерий состоит в том, что он придает белкам реактивность. Азот трудно входит в состав соединений из-за инертности, но легко из них выходит.[ …]
Влияние предшественника сказывается прежде всего на обеспеченности удобряемой культуры азотом. Растения, высеваемые после бобовых, оставляющих некоторое количество азота, ассимилированного клубеньковыми бактериями из атмосферы, лучше отзываются на фосфор, чем следующие по другим предшественникам. С. П. Кулжинский (1935) иллюстрировал это положение данными украинских опытных станций (табл. 72).[ …]
За последнее время накапливается все больше данных о том, что многие лизогенные культуры содержат 2, 3, 4 и более умеренных фагов, т. е. являются полилизогенными. Например, многие актиномицеты, проактиномицеты, клубеньковые бактерии и некоторые спороносные бактерии содержат 4 и более фагов. Содержащиеся в полилизогенных культурах фаги часто резко различаются между собой по форме частиц, антигенным свойствам и спектру лити-ческого действия. Полилизогенные культуры можно экспериментально получить с помощью воздействия на них одновременно или последовательно различными умеренными фагами. Полученные таким способом культуры не отличаются от выделенных из природных источников.[ …]
Полилизогенные культуры можно экспериментально получить с помощью воздействия на них одновременно или последовательно различными умеренными фагами. Полученные таким способом культуры не отличаются от выделенных из природных источников.[ …]
Эти изменения могут быть как положительными для плодородия, так и отрицательными. Примером положительных изменений является устранение избыточной кислотности в результате известкования, накопление азота за счет деятельности клубеньковых бактерий при посевах бобовых, удаление вредных солей в орошаемых почвах после их промывок, улучшение водно-воздушного режима за счет рыхления подпахотного слоя и т. д.[ …]
Зеленое удобрение прежде всего обогащает почву органическим веществом и азотом. Нередко, й зависимости от условий его применения, на гектаре пашни запахивают 35—45 т органической массы, содержащей 150— 200 кг азота, фиксированного из воздуха клубеньковыми бактериями (при посеве бобовых сидератов).[ …]
Мутуализм (симбиоз): каждый из видов может жить, расти и размножаться только в присутствии другого. Симбионтами могут быть только растения, или растения и животные, или только животные. Характерным примером пищеобусловленных симбионтов являются клубеньковые бактерии и бобовые, микориза некоторых грибов и корни деревьев, лишайники и термиты.[ …]
Симбионтами могут быть только растения, или растения и животные, или только животные. Характерным примером пищеобусловленных симбионтов являются клубеньковые бактерии и бобовые, микориза некоторых грибов и корни деревьев, лишайники и термиты.[ …]
Однако в природе и хозяйстве имеются существенные различия в круговороте азота и фосфора. Как известно, воздух почти на 4Д состоит из молекулярного азота. И хотя он недоступен высшим растениям, но усваивается некоторыми микроорганизмами, в частности клубеньковыми бактериями, которые живут на корнях бобовых культур. Эти бактерии снабжают азотистой пищей не только бобовые. При запахивании в почву пожнивных остатков и разложении корней остается достаточное количество азота для культуры, высеваемой после бобовых, особенно после клевера и люцерны.[ …]
Горох только в том случае обогащает почву азотом, когда на его корнях развиваются клубеньки, при этом чем больше и мощнее, тем лучше почва обогащается азотом. В этих целях семена гороха необходимо в день посева обработать нитрагином под навесом, предохраняя азотфиксирующие бактерии от губительного воздействия солнечных лучей.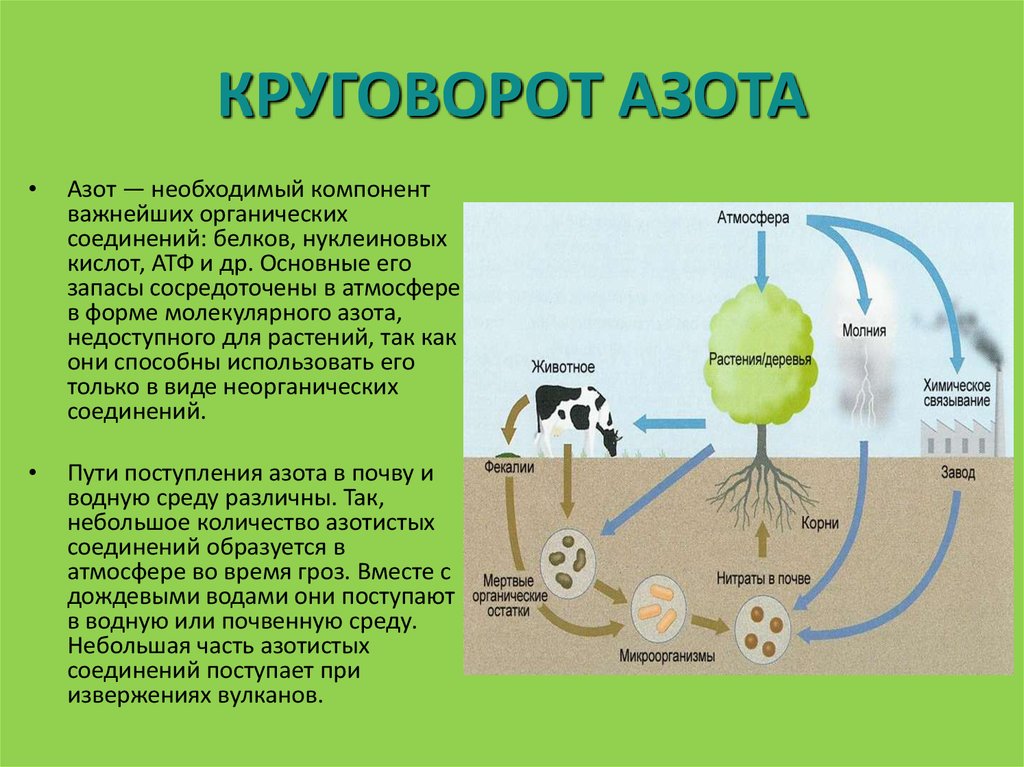 Содержимым одной бутылки нитрагина, растворенного в 2 л воды, смачивают семена, перелопачивая их. Подсохшие семена без промедления высевают. Нитрагиниза-ция вызывает раннее образование клубеньков и содействует лучшему развитию растений. Применение нитрагина эффективно при раннем посеве гороха во влажную почву. На неизвесткованных кислых почвах клубеньковые бактерии развиваются плохо и действие нитрагина резко снижается.[ …]
Содержимым одной бутылки нитрагина, растворенного в 2 л воды, смачивают семена, перелопачивая их. Подсохшие семена без промедления высевают. Нитрагиниза-ция вызывает раннее образование клубеньков и содействует лучшему развитию растений. Применение нитрагина эффективно при раннем посеве гороха во влажную почву. На неизвесткованных кислых почвах клубеньковые бактерии развиваются плохо и действие нитрагина резко снижается.[ …]
СИМБИОЗ (С.) — устойчивое совместное существование двух или нескольких видов организмов, при котором оба партнера (симбионта) или один из них получают преимущества в отношениях с внешней средой. С. — широкое понятие, которое включает как отношения взаимопомощи (мутуализм., комменсализм), так и паразитизм, поскольку хозяин и паразит сосуществуют длительное время. СИМБИОТРОФЫ (С.)— микроорганизмы (грибы, бактерии, одноклеточные простейшие животные), которые связаны отношениями мутуализма с растениями или животными (грибы микоризы, клубеньковые бактерии бобовых, бактерии и простейшие (амебы) пищеварительного тракта млекопитающих, включая человека). С. являются важнейшим компонентом экосистемы и условно относятся к консумен-там, так как их пищей служат прижизненные выделения органического вещества растений и еще не разложившиеся ткани животных.[ …]
С. являются важнейшим компонентом экосистемы и условно относятся к консумен-там, так как их пищей служат прижизненные выделения органического вещества растений и еще не разложившиеся ткани животных.[ …]
Типичным примером симбиоза может служить тесное сожительство между грибами и водорослями, приводящее к образованию более сложного и более приспособленного к природным условиям растительного организма — лишайника. Другим ярким примером симбиотического сожительства в почве является симбиоз грибов с высшими растениями, когда грибы образуют на корнях растений м и-к о р и з у. Явно выраженный симбиоз наблюдается между клубеньковыми бактериями и бобовыми растениями.[ …]
Д. Н. Прянишников доказывал, что для нашей страны более перспективно не травополье, а интенсивные плодосменные севообороты. Именно они пришли на смену трехполью зернового типа, господствовавшему на протяжении тысячи лет в Западной Европе. При трехполье треть земли пустовала (поздний пар), а две трети засевались зерновыми культурами. Бобовые не возделывали, что исключало возможность мобилизации азота воздуха с помощью клубеньковых бактерий и отрицательно сказывалось на азотном питании растений и круговороте азота в земледелии. В этом севообороте почти отсутствовали пропашные, в том числе картофель и корнеплоды, что приводило к засоренности полей и постоянному недостатку кормов. На протяжении столетий крестьянское хозяйство (за исключением кулацкой верхушки) не могло вырваться из порочного круга, отмеченного известным русским агрономом XVIII в. А. Т. Болотовым, который писал: «…без навоза земля не дает урожая, а навоза мало, так как мало скота, а скота мало, так как мало кормов, а кормов мало, так как без навоза земля не дает урожая» (1779).[ …]
Бобовые не возделывали, что исключало возможность мобилизации азота воздуха с помощью клубеньковых бактерий и отрицательно сказывалось на азотном питании растений и круговороте азота в земледелии. В этом севообороте почти отсутствовали пропашные, в том числе картофель и корнеплоды, что приводило к засоренности полей и постоянному недостатку кормов. На протяжении столетий крестьянское хозяйство (за исключением кулацкой верхушки) не могло вырваться из порочного круга, отмеченного известным русским агрономом XVIII в. А. Т. Болотовым, который писал: «…без навоза земля не дает урожая, а навоза мало, так как мало скота, а скота мало, так как мало кормов, а кормов мало, так как без навоза земля не дает урожая» (1779).[ …]
Трудности хранения, транспортировки и применения агаровых и бульонных препаратов нитрагина, а также небольшой срок их годности являются серьезными причинами, способствующими вытеснению этого препарата из производства. Сыпучие порошковидные препараты нитрагина обладают несомненными преимуществами по сравнению с агаровыми и бульонными. Технология их изготовления проще и экономичнее. Торфяные культуры дольше хранятся и легче транспортируются. Они защищают клетки клубеньковых бактерий от непосредственного контакта с удобрениями и сохраняют их в жизнеспособном состоянии на семенах, особенно при гранулировании семян с известью.[ …]
Технология их изготовления проще и экономичнее. Торфяные культуры дольше хранятся и легче транспортируются. Они защищают клетки клубеньковых бактерий от непосредственного контакта с удобрениями и сохраняют их в жизнеспособном состоянии на семенах, особенно при гранулировании семян с известью.[ …]
Высшее растение по схеме является источником углеродсодержащих соединений. Их трансформация обеспечивает получение энергетического материала для процессов активации и восстановления N2. Активированный азот — конечный акцептор электронов. Продукты неполного окисления углеродсодержащих соединений служат акцепторами 1ЧНз и в клубеньках образуют аминокислоты, которые становятся доступными высшему растению. Растения выполняют роль накопителя углеродсодержащих соединений (продуктов фотосинтеза) и поставщика энергии. Клубеньковые бактерии в стадии бактероидов проявляют способность с помощью нитрогеназы переносить активированный водород к азоту. Путь от N2 до Т Шз рассматривается как восстановительный процесс. [ …]
[ …]
Различные почвенные микроорганизмы неодинаково относятся к кислотности почвы. Плесневые грибы лучше развиваются при pH 3—6 и могут расти даже при более высокой кислотности. Среди грибов встречается много паразитов и возбудителей различных болезней культурных растений, например паразит свеклы РЬота Ье1ае, возбудитель килы у крестоцветных Р1азтос1шр110га Ьгайвшае. Развитие их в кислых почвах усиливается. В то же время многие полезные почвенные микроорганизмы лучше развиваются при нейтральной и слабощелочной реакции.[ …]
Растения гороха обладают способностью усваивать азот из воздуха, обогащая им почву, в связи с чем основным направлением в системе удобрения должно быть применение фосфорных и калийных удобрений. Хорошо развитая корневая система гороха отличается высокой усвояющей способностью питательных веществ, однако для значительного увеличения урожая нельзя рассчитывать на использование последействия удобрений в севообороте, в связи с чем необходимо непосредственное внесение минеральных удобрений под горох. Он отзывчив на применение фосфорных и калийных удобрений. Фосфор способствует ускорению созревания растений. При недостатке его в почве клубеньковые бактерии слабо развиваются на корнях, и урожай снижается. При недостатке калия листья желтеют, и бобы плохо развиваются, особенно на легких по механическому составу почвах. На кислых почвах горох резко снижает урожай, ослабляется деятельность клубеньковых бактерий. Поэтому известкование кислых почв является обязательным условием повышения урожая гороха и более высокой эффективности внесенных минеральных удобрений. Доза извести зависит от кислотности почвы (3…6 т/га), известь лучше вносить под предшествующие культуры.[ …]
Он отзывчив на применение фосфорных и калийных удобрений. Фосфор способствует ускорению созревания растений. При недостатке его в почве клубеньковые бактерии слабо развиваются на корнях, и урожай снижается. При недостатке калия листья желтеют, и бобы плохо развиваются, особенно на легких по механическому составу почвах. На кислых почвах горох резко снижает урожай, ослабляется деятельность клубеньковых бактерий. Поэтому известкование кислых почв является обязательным условием повышения урожая гороха и более высокой эффективности внесенных минеральных удобрений. Доза извести зависит от кислотности почвы (3…6 т/га), известь лучше вносить под предшествующие культуры.[ …]
Одним из наиболее важных процессов взаимодействия микроорганизмов с высшим растением является симбиотическая фиксация атмосферного азота — основного элемента, определяющего величину и качество урожая. Общее количество молекулярного азота, вовлекаемого в биологический круговорот симбиотической бобово-ризоби-альной системой, только в СССР составляет ежегодно 3 млн т. Многолетний отечественный и зарубежный опыт показывает, что эффективный бобово-ризобиальный симбиоз — не только залог получения высокого и качественного урожая бобовых культур, а следовательно, и возможности решения проблемы пищевого белка, но и наиболее экономичный источник пополнения запасов азота в почве. Для использования дешевого ’’биологического азота” в сельскохозяйственном производстве многих стран увеличивают посевные площади под бобовыми культурами, а также широко применяют предпосевную обработку семян препаратами клубеньковых бактерий, получаемых на основе активных штаммов Rhizobium. Характер и эффективность симбиотических взаимоотношений бобового растения с клубеньковыми бактериями зависят от физиолого-биохимического состояния обоих партнеров, в связи с чем влияние каких-либо факторов на одного из них непременно отражается на продуктивности системы в целом.[ …]
Многолетний отечественный и зарубежный опыт показывает, что эффективный бобово-ризобиальный симбиоз — не только залог получения высокого и качественного урожая бобовых культур, а следовательно, и возможности решения проблемы пищевого белка, но и наиболее экономичный источник пополнения запасов азота в почве. Для использования дешевого ’’биологического азота” в сельскохозяйственном производстве многих стран увеличивают посевные площади под бобовыми культурами, а также широко применяют предпосевную обработку семян препаратами клубеньковых бактерий, получаемых на основе активных штаммов Rhizobium. Характер и эффективность симбиотических взаимоотношений бобового растения с клубеньковыми бактериями зависят от физиолого-биохимического состояния обоих партнеров, в связи с чем влияние каких-либо факторов на одного из них непременно отражается на продуктивности системы в целом.[ …]
Концентрация в почве различных элементов напрямую зависит от количества в
Купить минеральные удобрения, mineral fertilizers в Санкт-Петербурге
В каталоге товаров/продукции представлены минеральные удобрения —
хелат удобрения, хелатные удобрения, простые водорастворимые удобрения, комплексные минеральные удобрения, микроудобрения, категории: chelate fertilizer, chelated fertilizer, simple water-soluble fertilizers, complex fertilizers, microfertilizers, ; хелат удобрения, хелаты, удобрения в форме хелатов, хелатные удобрения, комплексные хелатные удобрения, хелатные микроудобрения, комплексные хелатные микроудобрения, удобрения простые водорастворимые, нитрат кальция (кальциевая селитра), нитрат калия (калиевая селитра), монокалий фосфат, сульфат калия, нитрат магния, сульфат магния, , ,
хелат удобрения, хелатные удобрения
хелат удобрения, хелатные удобрения
Подробнее. ..
..
Купить хелат удобрения — эффективные хелатные удобрения для растений — chelate fertilizer в ХИМСНАБ-СПБ, контактный телефон +7-812-337-18-93 (plugin.block chelating_agent_trace_element) представлены в каталоге ХИМСНАБ-СПБ. Области применения микроудобрений, дозы применения препарата для различных культур, растворимость и содержание хилатирующего агента а также показатель активности водородны…
простые водорастворимые удобрения
простые водорастворимые удобрения
Подробнее…
комплексные минеральные удобрения
комплексные минеральные удобрения
Подробнее…
микроудобрения
микроудобрения
Подробнее. ..
..
Купить органо минеральные удобрения, organo mineral fertilizers в Санкт-Петербурге
В каталоге товаров/продукции представлены органо минеральные удобрения —
ОМУ удобрения, лигногуматы, категории: WMD fertilizer, Lignohumate, ; , ,
ОМУ удобрения
ОМУ удобрения
Подробнее…
лигногуматы
лигногуматы
Подробнее…
Инновационные гуминовые препараты и удобрения со склада в Санкт-Петербурге в компании ООО «ХИМСНАБ-СПБ» В каталоге ХИМСНАБ-СПБ можно купить развличные виды органо минеральных удобрений для растений и культур: лигногуматы и ОМУ удобрения. В зависимости от типа удобрения — варианты упаковки, фасовки: мешок или канистра. марка А — порошкообразный продукт; марка АМ — порошкообра…
В зависимости от типа удобрения — варианты упаковки, фасовки: мешок или канистра. марка А — порошкообразный продукт; марка АМ — порошкообра…
Оставьте заявку ON-LINE или позвоните. Менеджер компании ответит на ваши вопросы.
Оставить заявку ON-LINE
Или звонок в рабочее время
Широкий ассортимент
В каталоге компании более 4000 наименований продукции в 200 товарных категориях: химические реактивы, лаб. оборудование и посуда, аксессуары и принадлежности для лабораторий, различные виды удобрений, химическое сырьеи многе другое. Можно подобрать продукцию воспользовавшись фильтром характеристик.
Опт и розница
Осуществляем продажу оптом и в розницу. В каталоге Химснаб-СПБ можно заказать широкий спектр веществ различных квалификаций: «Технический» («тех. »); «Чистый» («ч.»); «Чистый для анализа» («ч.д.а.»); «Химически чистый» («х.ч.»); «Особо чистый» («ос.ч.»); имп.: неорганические реактивы, органические реактивы, особо чистые вещества, растворы (буферные растворы, растворы для очистки и хранения электродов, растворы для кондуктометров), химическое сырье и компоненты. Продукции для лабораторных исследований.
»); «Чистый» («ч.»); «Чистый для анализа» («ч.д.а.»); «Химически чистый» («х.ч.»); «Особо чистый» («ос.ч.»); имп.: неорганические реактивы, органические реактивы, особо чистые вещества, растворы (буферные растворы, растворы для очистки и хранения электродов, растворы для кондуктометров), химическое сырье и компоненты. Продукции для лабораторных исследований.
Проверенные поставщики
Компания реализует товары и продукцию только от проверенных поставщиков гарантирующих качестно продукции.
Консультация по продукции
Менеджеры компании проконсультируют вас по ассортименту реализуемой продукции, звоните в рабочее время
Доставка
География потребителей выходит за пределы России, компания «Химснаб-СПБ» осуществляет доставку приобретаемых товаров и продукции по Санкт-Петербургу, Ленинрадской обл, России и странам СНГ.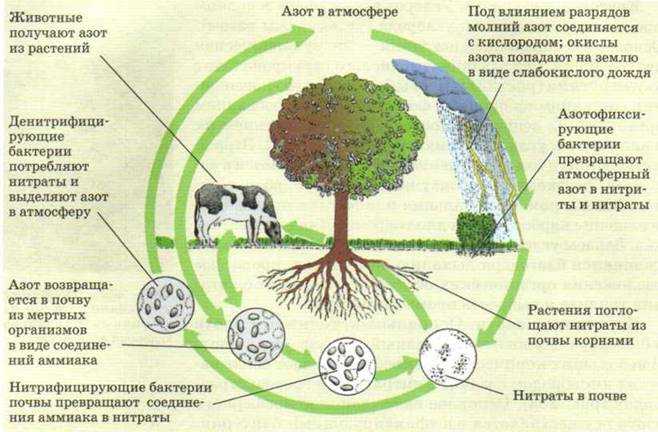
Индивидуальный подход
Строим свое сотрудничество с клиентом с учетом всех пожеланий клиента.
Гибкий и индивидуальный подход к каждому клиенту, ориентированность на долгосрочные партнерские отношения, строгое соблюдение оговоренных сроков и предоставления документов заказчику являются неоспоримыми преимуществами компании «Химснаб-СПБ». Мы заботимся о том, чтобы каждый наш клиент остался доволен приобретаемой продукцией и полученным результатом, который является нашим общим успехом!
Малотоннажная химия
Реализация продукции малотоннажной химии: продукция химической и нефтехимической промышленности. Малотоннажная химия дает возможность на скромном оборудовании и в небольших объемах производить дорогостоящие модификаторы, пластификаторы, ингибиторы и другие микродобавки, способные наделять конечный продукт новыми свойствами
Комплексное снабжение, оснащение
Компания Химснаб-СПБ имеет многолетний опыт работы на рынке химической продукции и лабораторного оборудования. Компания тесно сотрудничает со многими промышленными и производственными организациями и имеет возможность осуществлять комплексное снабжение и оснащение предприятии различных отраслений промышленности необходимым оборудованием и расходными материалами.
Компания тесно сотрудничает со многими промышленными и производственными организациями и имеет возможность осуществлять комплексное снабжение и оснащение предприятии различных отраслений промышленности необходимым оборудованием и расходными материалами.
Предствленная информация на страницах данного интернет-сайта и в каталоге продукции носит исключительно информационный характер и ни при каких условиях не является публичной офертой, определяемой положениями Статьи 437 (2) Гражданского кодекса РФ. Для получения подробной информации о наличии и стоимости указанных товаров и (или) услуг,обращайтесь к менеджерам отдела продаж: форма обратной связи, e-mail, телефон.
Реализация продукции для сельского хозяйства, химической, строительной, нефтегазовой, металлургической, текстильной, кожевенной, и других отраслей промышленности.
7 812-337-18-93
Возникли вопросы, звоните: пн-пт с 9:00 до 17:00 или оставьте Ваш телефон и мы Вам перезвоним.
Возможности «ХИМСНАБ-СПБ»:
Предлагаем широкие возможности для комплектации химической продукцией производства и исследовательских лабораторий в различных отраслях промышленности.
Приемущества «ХИМСНАБ-СПБ»:
Комплексное снабжение:
«ХИМСНАБ-СПБ» — Ваш надежный поставщик
Поставка химической продукции и лабораторного оборудования является ключевым направлением деятельности компании с 1996 года.
Компания «ХИМСНАБ-СПБ» успешно осуществляет поставку широкого спектра лабороторного оборудования, приборов и другой химической продукции на рынке Северо-Запада Российской Федерации.
- Широкий ассортимент продукции
- Опт и розница
- Консультация по продукции
- Доставка транспортными компаниями
- Индивидуальный подход
- Проверенные поставщики
- Малотоннажная химия
- Комплексное снабжение, оснащение
О компании Химснаб-СПБ
-
Нитрагин
бактериальные удобрения
Похожие статьи
Марки удобрений Акварин комплексные водорастворимые минеральные удобрения
-
30 ноября 2008
дефицит элементов питания растений, недостаток минеральных удобрений
-
01 апреля 2015
дефицит кобальта у растений, недостаток элемента кобальт
-
02 апреля 2015
важные микроэлементы для развития и роста растений
-
01 мая 2015
Хелатное удобрение способствует активизации ферментов и процессы фотосинтеза в структуре растений
-
01 июня 2015
Хелатное удобрение способствует повышению морозоустойчивости и засухоустойчивости
-
02 июня 2015
Хелатное удобрение способствует ускорению роста и развития стеблей и листьев растения
-
06 июня 2015
Хелатное удобрение способствует повышению урожайности культур
-
07 июня 2015
Хелатное удобрение способствует улучшению качества выращенного урожая
-
08 июня 2015
Концентрированный растворимый комплекс Akvarin
-
01 января 2016
Удобрения для листовых подкормок растений Akvarin
-
02 января 2016
Особенности внесение водорастворимых комплексных минеральных удобрений
-
03 января 2016
Углеродно-азотный баланс конкреции и его регуляция при повышенной концентрации углекислого газа
1. Phillips DA. Эффективность симбиотической азотфиксации у бобовых. Ежегодный обзор физиологии растений . 1980; 31: 29–49. [Google Scholar]
Phillips DA. Эффективность симбиотической азотфиксации у бобовых. Ежегодный обзор физиологии растений . 1980; 31: 29–49. [Google Scholar]
2. Стейси Г., Либо М., Брехенмахер Л., Ван Дж., Мэй Г.Д. Генетика и функциональная геномика клубеньков бобовых. Текущее мнение по биологии растений . 2006;9(2):110–121. [PubMed] [Академия Google]
3. Олдройд GED, Дауни Дж.А. Координация морфогенеза клубеньков с ризобиальной инфекцией бобовых. Ежегодный обзор биологии растений . 2008; 59: 519–546. [PubMed] [Google Scholar]
4. Мюррей Д.Д. Инвазия по приглашению: ризобиальная инфекция бобовых. Молекулярные взаимодействия растений и микробов . 2011;24(6):631–639. [PubMed] [Google Scholar]
5. Oldroyd GED, Murray JD, Poole PS, Downie JA. Правила участия в бобово-ризобиальном симбиозе. Ежегодный обзор генетики . 2011;45:119–144. [PubMed] [Google Scholar]
6. Олдройд GED. Говори, дружи и вступай: сигнальные системы, которые способствуют полезным симбиотическим ассоциациям у растений. Nature Reviews Микробиология . 2013;11(4):252–263. [PubMed] [Google Scholar]
Nature Reviews Микробиология . 2013;11(4):252–263. [PubMed] [Google Scholar]
7. Venkateshwaran M, Volkening JD, Sussman MR, Ané J-M. Симбиоз и социальная сеть высших растений. Текущее мнение по биологии растений . 2013;16(1):118–127. [PubMed] [Google Scholar]
8. Kosslak RM, Bohlool BB. Подавление образования клубеньков с одной стороны расщепленной корневой системы сои, вызванное предварительной инокуляцией другой стороны. Физиология растений . 1984; 75: 125–130. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Делвс А.С., Мэтьюз А., Дей Д.А., Картер А.С., Кэрролл Б.Дж., Грессхофф П.М. Регуляция соево-ризобиального симбиоза клубеньков побеговыми и корневыми факторами. Физиология растений . 1986; 82: 588–590. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Caetano-Anollés G, Gresshoff PM. Генетический контроль клубеньков растений. Ежегодный обзор микробиологии . 1991; 45: 345–382. [PubMed] [Академия Google]
11. Ferguson BJ, Indrasumunar A, Hayashi S, et al. Молекулярный анализ развития клубеньков бобовых и ауторегуляции. Журнал интегративной биологии растений . 2010;52(1):61–76. [PubMed] [Google Scholar]
Ferguson BJ, Indrasumunar A, Hayashi S, et al. Молекулярный анализ развития клубеньков бобовых и ауторегуляции. Журнал интегративной биологии растений . 2010;52(1):61–76. [PubMed] [Google Scholar]
12. Nishimura R, Hayashit M, Wu G-J, et al.
HAR1 опосредует системную регуляцию развития симбиотических органов. Природа . 2002;420(6914):426–429. [PubMed] [Google Scholar]
13. Лохар Д.П., ВанденБош К.А. Прививка между модельными бобовыми демонстрирует роль корней и побегов в определении типа клубенька и специфичности хозяина/ризобий. Журнал экспериментальной ботаники . 2005;56(416):1643–1650. [PubMed] [Google Scholar]
14. Schnabel E, Journet E-P, de Carvalho-Niebel F, Duc G, Frugoli J. Ген Medicago truncatula SUNN кодирует CLV1-подобную киназу рецептора повторов, богатую лейцином, которая регулирует клубеньки. число и длина корня. Молекулярная биология растений . 2005;58(6):809–822. [PubMed] [Google Scholar]
15. Magori S, Oka-Kira E, Shibata S, et al. Слишком много любви, корневой регулятор, связанный с дистанционным контролем нодуляции в Лотос японский
Magori S, Oka-Kira E, Shibata S, et al. Слишком много любви, корневой регулятор, связанный с дистанционным контролем нодуляции в Лотос японский
. Молекулярные взаимодействия растений и микробов . 2009;22(3):259–268. [PubMed] [Google Scholar]
16. Ёсида С., Фунаяма-Ногучи С., Кавагути М. Пленти, новый мутант с гипернодуляцией в Lotus japonicus
. Физиология растений и клеток . 2010;51(9):1425–1435. [PubMed] [Google Scholar]
17. Bourion V, Laguerre G, Depret G, Voisin A-S, Salon C, Duc G. Генетическая изменчивость клубеньков и роста корней влияет на фиксацию и накопление азота в горохе. Анналы ботаники . 2007;100(3):589–598. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Cabeza RA, Lingner A, Liese R, et al. Активность клубеньков суперклубенькового мутанта Mt sunn не лимитируется фотосинтезом в оптимальных условиях роста. Международный журнал молекулярных наук . 2014;15:6031–6045. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Burdon JJ, Gibson AH, Searle SD, Woods MJ, Brockwell J. Изменение эффективности симбиотических ассоциаций между аборигенными ризобиями и умеренным климатом Австралийская акация Внутривидовые взаимодействия. Журнал прикладной экологии . 1999;36(3):398–408. [Google Scholar]
20. Denton MD, Coventry DR, Bellotti WD, Howieson JG. Распространение, численность и симбиотическая эффективность Rhizobium leguminosarum bv. trifolii из щелочных пастбищных почв Южной Австралии. Австралийский журнал экспериментального сельского хозяйства . 2000;40(1):25–35. [Google Scholar]
21. Garau G, Reeve WG, Brau L, et al. Симбиотические потребности различных видов Medicago spp. предложить эволюцию Sinorhizobium meliloti и S. medicae с растениями-хозяевами, по-разному адаптированными к рН почвы. Растения и почва . 2005;276(1-2):263–277. [Google Scholar]
22. Терполелли Дж. Дж., О’Хара Г. В., Тивари Р. П., Дилворт М. Дж., Хоуисон Дж. Г. Модель бобовых Medicago truncatula A17 плохо сочетается по фиксации N 2 с секвенированным микросимбионтом Sinorhizobium meliloti 1021. Новый фитолог . 2008;179(1):62–66. [PubMed] [Академия Google]
Терполелли Дж. Дж., О’Хара Г. В., Тивари Р. П., Дилворт М. Дж., Хоуисон Дж. Г. Модель бобовых Medicago truncatula A17 плохо сочетается по фиксации N 2 с секвенированным микросимбионтом Sinorhizobium meliloti 1021. Новый фитолог . 2008;179(1):62–66. [PubMed] [Академия Google]
23. Амаргер Н. Селекция штаммов Rhizobium на их конкурентоспособность в отношении образования клубеньков. Биология и биохимия почв . 1981;13(6):481–486. [Google Scholar]
24. Синглтон П.В., Стокингер К.Р. Компенсация неэффективного образования клубеньков на сое. Растениеводство . 1983; 23: 69–72. [Google Scholar]
25. Хан М., Штудер Д. Конкурентоспособность мутанта anif- Bradyrhizobium japonicum по сравнению со штаммом дикого типа. Письма по микробиологии FEMS . 1986;33(1):143–148. [Google Scholar]
26. Kiers ET, Rousseau RA, West SA, Denlson RF. Санкции против хозяина и мутуализм бобовых- ризобий . Природа . 2003; 425 (6953): 78–81. [PubMed] [Google Scholar]
Природа . 2003; 425 (6953): 78–81. [PubMed] [Google Scholar]
27. Гейдж DJ. Анализ развития инфекционных нитей с использованием Gfp- и DsRed-экспрессирующих штаммов Sinorhizobium meliloti
. Журнал бактериологии . 2002;184(24):7042–7046. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Denison RF, Kiers ET. Истории жизни симбиотических ризобий и микоризных грибов. Актуальная биология . 2011;21(18):R775–R785. [PubMed] [Google Scholar]
29. Oono R, Anderson CG, Denison RF. Неспособность фиксировать азот нерепродуктивными симбиотическими ризобиями вызывает санкции хозяина, которые снижают приспособленность их репродуктивных клонов. Труды Королевского общества B: биологические науки . 2011;278(1718):2698–2703. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. West SA, Kiers ET, Simms EL, Denison RF. Санкции и стабильность мутуализма: почему ризобии фиксируют азот? Труды Королевского общества B: биологические науки . 2002; 269 (1492): 685–694. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2002; 269 (1492): 685–694. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Denison RF, Kiers ET. Альтернативы образу жизни для ризобий: мутуализм, паразитизм и отказ от симбиоза. Письма по микробиологии FEMS . 2004;237(2):187–193. [PubMed] [Google Scholar]
32. Агилар О.М., Рива О., Пельтцер Э. Анализ Rhizobium etli и его симбиоза с диким Phaseolus vulgaris поддерживает коэволюцию в центрах диверсификации хозяев. Труды Национальной академии наук Соединенных Штатов Америки . 2004;101(37):13548–13553. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Zanetti ME, Blanco FA, Beker MP, Battaglia M, Aguilar OM. Субъединица C ядерного фактора растений NF-Y, необходимая для ризобиальной инфекции и развития клубеньков, влияет на выбор партнера в симбиозе обыкновенной фасоли- Rhizobium etli . Растительная клетка . 2010;22(12):4142–4157. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Петрони К., Кумимото Р.В., Гнесутта Н. и др. Беспорядочная жизнь факторов транскрипции растений NUCLEAR FACTOR Y. Растительная клетка . 2012;24(12):4777–4792. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Петрони К., Кумимото Р.В., Гнесутта Н. и др. Беспорядочная жизнь факторов транскрипции растений NUCLEAR FACTOR Y. Растительная клетка . 2012;24(12):4777–4792. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Benedito VA, Torres-Jerez I, Murray JD, et al. Атлас экспрессии генов модельного бобового растения Medicago truncatula
. Журнал завода . 2008;55(3):504–513. [PubMed] [Google Scholar]
36. He J, Benedito VA, Wang M, et al. Медикаго укороченный 9Веб-сервер атласа экспрессии генов 0004. Биоинформатика BMC . 2009;10, статья 441 [бесплатная статья PMC] [PubMed] [Google Scholar]
37. Libault M, Farmer A, Joshi T, et al. Интегрированный атлас транскриптома сельскохозяйственной модели Glycine max и его использование в сравнительном анализе растений. Журнал завода . 2010;63(1):86–99. [PubMed] [Google Scholar]
38. Северин А.Дж., Вуди Дж.Л., Болон Ю.Т. и соавт. RNA-Seq Atlas Glycine max : справочник по транскриптому сои. BMC Биология растений . 2010;10, статья 160 [бесплатная статья PMC] [PubMed] [Google Scholar]
BMC Биология растений . 2010;10, статья 160 [бесплатная статья PMC] [PubMed] [Google Scholar]
39. Joshi T, Patil K, Fitzpatrick MR, et al. База знаний о сое (SoyKB): веб-ресурс по трансляционной геномике сои. BMC Genomics . 2012;13(приложение 1, статья S15) [бесплатная статья PMC] [PubMed] [Google Scholar]
40. Hackenberg D, Keetman U, Grimm B. Гомологическая субъединица NF-YC2 арабидопсиса и табака активируется фотоокислительным стрессом и вызывает цветение. Международный журнал молекулярных наук . 2012;13(3):3458–3477. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Roth LE, Stacey G. Высвобождение бактерий в клетки-хозяева азотфиксирующих клубеньков сои: симбиосомная мембрана происходит из трех источников. Европейский журнал клеточной биологии . 1989;49(1):13–23. [PubMed] [Google Scholar]
42. Lodwig EM, Leonard M, Marroqui S, et al. Роль полигидроксибутирата и гликогена в бактероидах гороха и фасоли в качестве соединений для хранения углерода. Молекулярные взаимодействия растений и микробов . 2005;18(1):67–74. [PubMed] [Google Scholar]
Молекулярные взаимодействия растений и микробов . 2005;18(1):67–74. [PubMed] [Google Scholar]
43. Oono R, Schmitt I, Sprent JI, Denison RF. Множественное эволюционное происхождение признаков бобовых, ведущее к крайней ризобиальной дифференциации. Новый фитолог . 2010;187(2):508–520. [PubMed] [Google Scholar]
44. Van de Velde W, Zehirov G, Szatmari A, et al. Растительные пептиды регулируют конечную дифференциацию бактерий в симбиозе. Наука . 2010;327(5969):1122–1126. [PubMed] [Академия Google]
45. Saalbach G, Erik P, Wienkoop S. Характеристика протеомики перибактериоидного пространства и препаратов перибактериоидной мембраны из симбиосом гороха ( Pisum sativum ). Протеомика . 2002; 2: 325–337. [PubMed] [Google Scholar]
46. Hohnjec N, Lenz F, Fehlberg V, et al. Сигнальный пептид модульного нодулина MtNOD25 Medicago truncatula действует как адресная метка для специфического нацеливания белков на азотфиксирующие симбиосомы. Молекулярные взаимодействия растений и микробов . 2009;22(1):63–72. [PubMed] [Google Scholar]
Молекулярные взаимодействия растений и микробов . 2009;22(1):63–72. [PubMed] [Google Scholar]
47. Meckfessel MH, Blancaflor EB, Plunkett M, Dong Q, Dickstein R. Множественные домены в белке mtenod8, включая сигнальный пептид, нацеливают его на симбиосому. Физиология растений . 2012;159(1):299–310. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Mergaert P, Nikovics K, Kelemen Z, et al. Новое семейство Medicago truncatula , состоящее из более чем 300 специфичных для клубеньков генов, кодирующих небольшие секретируемые полипептиды с консервативными цистеиновыми мотивами. Физиология растений . 2003;132(1):161–173. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Mergaert P, Uchiumi T, Alunni B, et al. Эукариотический контроль бактериального клеточного цикла и дифференцировки в симбиозе Rhizobium и бобовых. Труды Национальной академии наук Соединенных Штатов Америки . 2006;103(13):5230–5235. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Alunni B, Kevei Z, Redondo-Nieto M, Kondorosi A, Mergaert P, Kondorosi E. Геномная организация и эволюционные взгляды на Гены GRP и NCR , два больших семейства генов, специфичных для узелков, в Medicago truncatula
. Молекулярные взаимодействия растений и микробов . 2007;20(9):1138–1148. [PubMed] [Google Scholar]
51. Керест А., Мергерт П., Кондороси Э. Развитие бактероидов в клубеньках бобовых: эволюция взаимной выгоды или жертв? Молекулярные взаимодействия растений и микробов . 2011;24(11):1300–1309. [PubMed] [Google Scholar]
52. Wang D, Griffitts J, Starker C, et al. Путь секреции белка, специфичный для узелков, необходим для азотфиксирующего симбиоза. Наука . 2010;327(5969):1126–1129. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Cabeza R, Koester B, Liese R, et al. Анализ транскриптома секвенирования РНК раскрывает новое понимание молекулярных аспектов воздействия нитратов на активность клубеньков Medicago truncatula .
. Физиология растений . 2014; 164:400–411. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Haag AF, Baloban M, Sani M, et al. Защита Sinorhizobium от богатых цистеином антимикробных пептидов хозяина имеет решающее значение для симбиоза. ПЛОС Биология . 2011;9(10)e1001169 [бесплатная статья PMC] [PubMed] [Google Scholar]
55. Marlow VL, Haag AF, Kobayashi H, et al. Существенная роль белка BacA в поглощении укороченного эукариотического пептида у Sinorhizobium meliloti
. Журнал бактериологии . 2009;191(5):1519–1527. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Karunakaran R, Haag AF, East AK, et al.
BacA необходим для развития бактероидов в клубеньках галегоидных, но не фазеолоидных бобовых культур. Журнал бактериологии . 2010;192(11):2920–2928. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Удварди М.К., Дэй Д.А. Транспорт метаболитов через симбиотические мембраны клубеньков бобовых.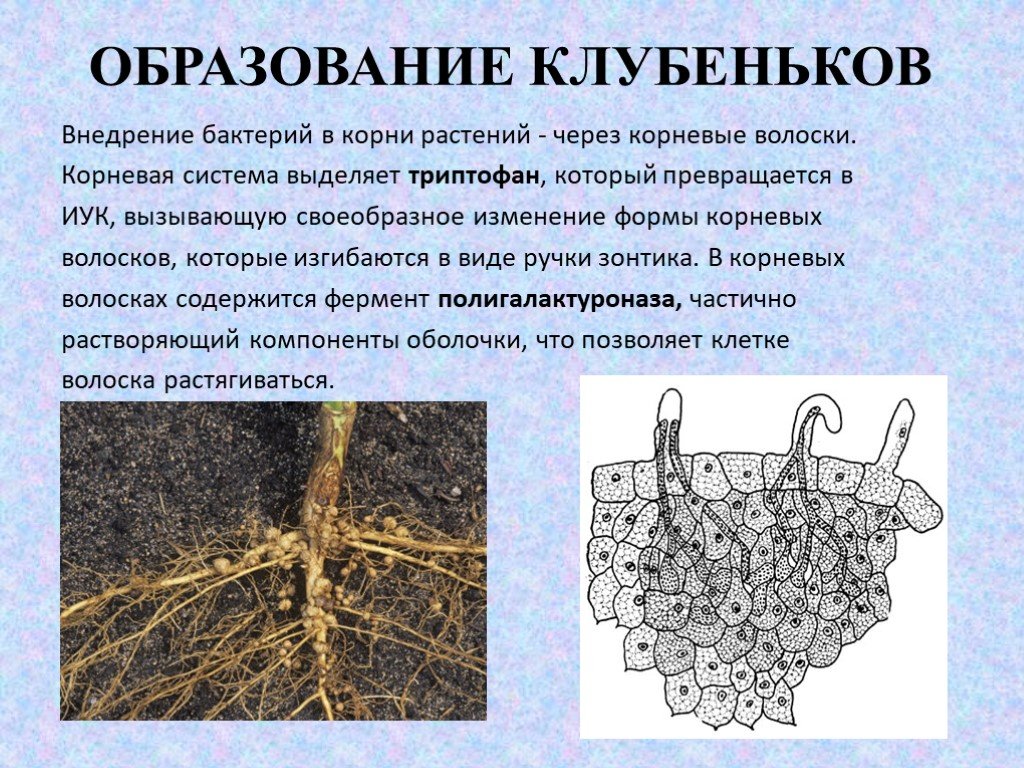 Ежегодный обзор биологии растений . 1997; 48: 493–523. [PubMed] [Google Scholar]
Ежегодный обзор биологии растений . 1997; 48: 493–523. [PubMed] [Google Scholar]
58. Benedito VA, Li H, Dai X, et al. Геномная инвентаризация и анализ транскрипции транспортеров Medicago truncatula . Физиология растений . 2010;152(3):1716–1730. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Масалкар П., Уоллес И.С., Хван Дж.Х., Робертс Д.М. Взаимодействие цитозольной глутаминсинтетазы корневых клубеньков сои с С-концевым доменом нодулина 26 мембраны симбиосомы акваглицеропорина. Журнал биологической химии . 2010;285(31):23880–23888. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Fotelli MN, Tsikou D, Kolliopoulou A, et al. Клубеньки усиливают фиксацию и рециркуляцию темнового СО 2 в модели бобовых Lotus japonicus
. Журнал экспериментальной ботаники . 2011;62(8):2959–2971. [PubMed] [Google Scholar]
61. Удварди М., Пул П.С. Транспорт и метаболизм в бобово-ризобиальных симбиозах. Ежегодный обзор биологии растений . 2013; 64: 781–805. [PubMed] [Google Scholar]
Ежегодный обзор биологии растений . 2013; 64: 781–805. [PubMed] [Google Scholar]
62. Тренер MA, Charles TC. Роль метаболизма ПОБ в симбиозе ризобий с бобовыми. Прикладная микробиология и биотехнология . 2006;71(4):377–386. [PubMed] [Академия Google]
63. Libault M, Farmer A, Brechenmacher L, et al. Полный транскриптом клетки корневого волоска сои, одноклеточная модель, и ее изменение в ответ на инфекцию Bradyrhizobium japonicum . Физиология растений . 2010;152(2):541–552. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Libault M, Govindarajulu M, Berg RH, et al. Соевый белок с двойной направленностью участвует в заражении Bradyrhizobium japonicum корневых волосков сои и клеток коры. Молекулярные взаимодействия растений и микробов . 2011;24(9):1051–1060. [PubMed] [Google Scholar]
65. Newcomb EH, Tandon SR, Kowal RR. Ультраструктурная специализация по продукции уреидов в неинфицированных клетках корневых клубеньков сои.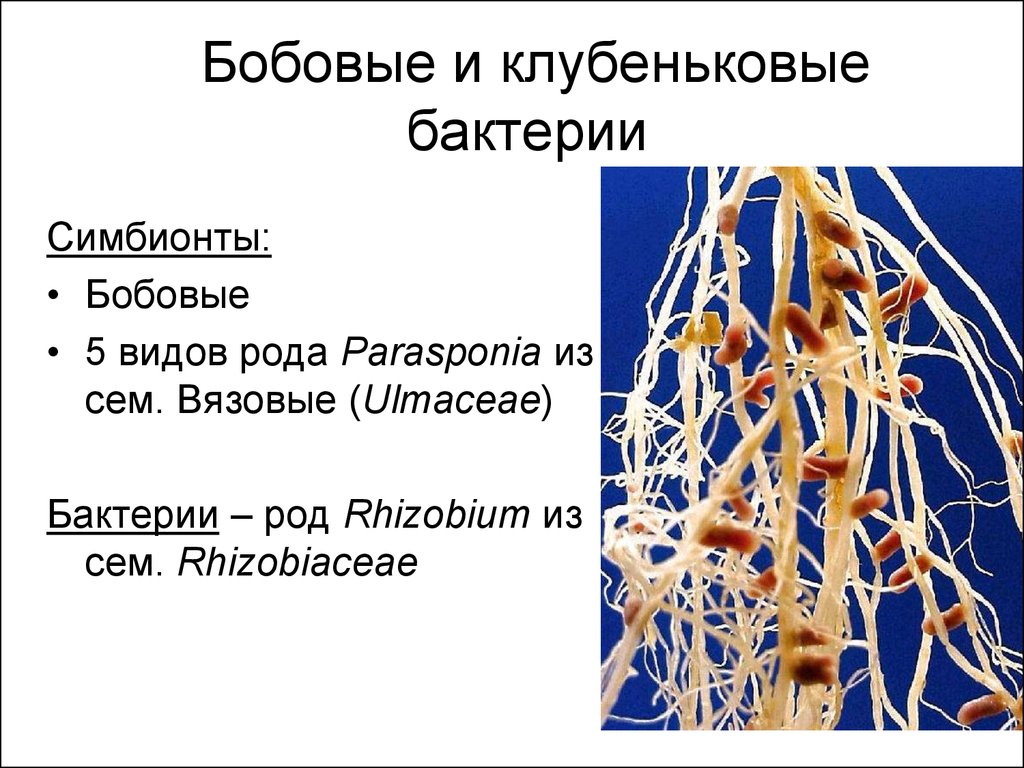 Протоплазма . 1985;125(1-2):1–12. [Google Scholar]
Протоплазма . 1985;125(1-2):1–12. [Google Scholar]
66. Бергерсен Ф.Дж., Пиплс М.Б., Тернер Г.Л. Роль поли- β -гидроксибутирата в бактероидах корневых клубеньков сои. Труды Королевского общества B: биологические науки . 1991;245(1312):59–64. [Google Scholar]
67. Чермола М., Федорова Э., Тате Р., Риччио А., Фавр Р., Патриарка Э.Дж. Инвазия клубеньков и дифференцировка симбиосом во время симбиоза Rhizobium etli-Phaseolus vulgaris . Молекулярные взаимодействия растений и микробов . 2000;13(7):733–741. [PubMed] [Google Scholar]
68. Wang C, Saldanha M, Sheng X, et al. Роль поли-3-гидроксибутирата (ПГБ) и гликогена в симбиозе Sinorhizobium meliloti с Медикаго сп
. Микробиология . 2007;153(2):388–398. [PubMed] [Google Scholar]
69. Valls M, Genin S, Boucher C. Комплексная регуляция системы секреции типа III и других детерминант вирулентности в Ralstonia solanacearum
. Патогены PLoS . 2006;2(8, статья e82) [бесплатная статья PMC] [PubMed] [Google Scholar]
Патогены PLoS . 2006;2(8, статья e82) [бесплатная статья PMC] [PubMed] [Google Scholar]
70. Marchetti M, Capela D, Glew M, et al. Экспериментальная эволюция патогена растений в симбионт бобовых. PLoS Биология . 2010;8(1)e1000280 [бесплатная статья PMC] [PubMed] [Google Scholar]
71. IPCC. Изменение климата 2014: смягчение последствий изменения климата . 2014. [Google Scholar]
72. Диас С., Грайм Дж. П., Харрис Дж., Макферсон Э. Доказательства механизма обратной связи, ограничивающего реакцию растений на повышенное содержание углекислого газа. Природа . 1993;364(6438):616–617. [Google Scholar]
73. Rice CW, Garcia FO, Hampton CO, Owensby CE. Микробная реакция почвы в высокотравных прериях на повышенное содержание CO 2
. Растения и почва . 1994;165(1):67–74. [Google Scholar]
74. Soussana JF, Hartwig UA. Влияние повышенного содержания CO 2 на симбиотическую фиксацию N 2 : связь между циклами углерода и азота в пастбищных экосистемах. Растения и почва . 1996;187(2):321–332. [Google Scholar]
Растения и почва . 1996;187(2):321–332. [Google Scholar]
75. Zanetti S, Hartwig UA, Lüscher A, et al. Стимуляция фиксации симбиотика N 2 в Trifolium repens L. при повышенном рСО в атмосфере 2 в пастбищной экосистеме. Физиология растений . 1996;112(2):575–583. [Бесплатная статья PMC] [PubMed] [Google Scholar]
76. Hungate BA, Dukes JS, Shaw MR, Luo Y, Field CB. Атмосферная наука. Азот и изменение климата. Наука . 2003;302(5650):1512–1513. [PubMed] [Google Scholar]
77. Роджерс А., Эйнсворт Э.А., Лики А.Д.Б. Усилит ли повышенная концентрация углекислого газа преимущества фиксации азота в бобовых? Физиология растений . 2009;151(3):1009–1016. [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Luo Y, Su B, Currie WS, et al. Прогрессирующее ограничение реакции экосистем азотом на повышение концентрации CO 2 в атмосфере. БиоНаука . 2004;54(8):731–739. [Google Scholar]
Растения, которые поглощают азот из воздуха, процветают в засушливых условиях – Florida Museum Science
разнообразны в засушливых районах страны. Этот вывод противоречит преобладающему предположению, что азотфиксаторы должны быть сравнительно наиболее разнообразны в средах, где азот в почве ограничен.
Этот вывод противоречит преобладающему предположению, что азотфиксаторы должны быть сравнительно наиболее разнообразны в средах, где азот в почве ограничен.
Результаты были особенно неожиданными, учитывая, что у азотфиксаторов отсутствуют признаки, часто присущие сухим почвам, такие как толстые запасающие воду стебли кактусов. «На первый взгляд, азотфиксаторы не обязательно приспособлены к засушливым экосистемам», — сказал ведущий автор Джошуа Доби, докторант кафедры биологии Университета Флориды.
Причина такой неожиданной закономерности не сразу была ясна, но Доби подозревает, что это связано с тем, как азотфиксаторы и нефиксаторы используют этот элемент.
Растения проявляют творческий подход к освоению азота
Растения включают азот почти во все структуры и реакции, происходящие в их клетках. Без него они не смогли бы производить белки, создавать ферменты или даже фотосинтезировать. С таким высоким спросом на азот он часто является одним из самых больших ограничений для роста растений — его просто не хватает.
Чтобы компенсировать этот недостаток, растения неоднократно разрабатывали инновационные способы выжимания из окружающей среды как можно большего количества азота. Венериные мухоловки и липкая росянка развили структуры, позволяющие красть азот у насекомых, навозные мхи растут исключительно на помете животных, а видоизмененные листья некоторых растений-кувшинов служат домом для ночлега летучих мышей в обмен на богатые азотом фекалии, которые они оставляют после себя.
В то время как азот может быть труднодоступным в почве, его почти неиссякаемый запас находится вне досягаемости. Газообразный азот составляет около 78% земной атмосферы, но, подобно тому, как люди умирают от жажды, потерявшись в море, растения совершенно не способны его поглощать.
Бактерии, с другой стороны, неоднократно осваивали трюк фиксации атмосферного азота. Случайный союз между такой бактерией и растением привел к одному из крупнейших ботанических излучений на Земле, давшему начало семейству бобовых, которое сегодня насчитывает более 18 000 видов. На корнях многих растений фасоли образуются полые узелки, способствующие росту бактерий. Растения наполняют эти предкамеры сахаром, чтобы кормить своих бактериальных партнеров и извлекать азот, который они выделяют в виде отходов.
На корнях многих растений фасоли образуются полые узелки, способствующие росту бактерий. Растения наполняют эти предкамеры сахаром, чтобы кормить своих бактериальных партнеров и извлекать азот, который они выделяют в виде отходов.
Опосредованная бактериями фиксация азота в растениях была открыта голландским ботаником Мартинусом Бейеринком, который в то время изучал растительные галлы и не понимал, почему такие галлы могут образовываться на корнях без явной помощи паразитических насекомых.
Симбиотические отношения, которые позволяют растениям получать доступ к атмосферному азоту, многократно развивались в отдаленно родственных группах, включая цветковые растения, саговники и водные папоротники.
Ботаники десятилетиями предполагали, что растения, содержащие азотфиксирующие бактерии, должны быть более разнообразными в таких экосистемах, как саванны и луга. Лесные пожары регулярно охватывают эти места, превращая деревья и кустарники, наполненные питательными веществами, в дым и пепел, которые уносятся ветром. Любые питательные вещества, оставшиеся в почве после пожара, могут ускользнуть от корней во время дождя, прежде чем прорастающие растения успеют их поглотить.
Любые питательные вещества, оставшиеся в почве после пожара, могут ускользнуть от корней во время дождя, прежде чем прорастающие растения успеют их поглотить.
Фиксация азота подготовила растения к более прохладной и сухой планете
Исследователи из Музея естественной истории Флориды, Университета штата Луизиана и Университета штата Миссисипи решили определить, какие факторы окружающей среды сыграли наибольшую роль в формировании сообществ азотфиксирующих растений. В Соединенных Штатах. Используя данные Национальной сети экологических обсерваторий (NEON), они проанализировали записи как местных, так и инвазивных видов из более чем 40 мест в США, включая Пуэрто-Рико.
Согласно их результатам, количество азотфиксаторов увеличивалось в среде с низким содержанием азота и уменьшалось в более сухих местообитаниях, как и ожидалось. В засушливой среде обычно меньше видов растений, чем в тех, где выпадает больше осадков, и азотфиксаторы не являются исключением из этого правила.
Неожиданность возникла, когда исследователи специально изучили разнообразие природных фиксаторов азота. Хотя видов было меньше, разнообразие аборигенных азотфиксаторов резко возросло в засушливых регионах, независимо от количества азота, доступного в почве.
Разница между количеством видов, называемая видовым богатством, и разнообразием аналогична выбору цветовой палитры. Палитра с 16 оттенками синего содержит больше оттенков, чем палитра с восемью дополнительными цветами; однако вторая палитра содержит большее разнообразие цветов и охватывает более широкий спектр видимого света.
Хотя первое цветовое колесо имеет в два раза больше оттенков, чем второе, колесо с восемью дополнительными цветами охватывает больший спектр видимого света.
Флоридский музей. Изображение Джеральда Пинсона.
Существуют тысячи видов цветковых растений, в которых обитают азотфиксирующие бактерии, и часто в пустынях и кустарниковых равнинах бок о бок растут особи, имеющие лишь отдаленное родство друг с другом. К ним относятся знакомые примеры, такие как мескитовые деревья и ольха.
К ним относятся знакомые примеры, такие как мескитовые деревья и ольха.
В то время как растения, любящие засуху, такие как кактусы, развили целый ряд черт, которые позволяют им процветать в пустынях, Доби считает, что растения с богатым запасом азота имеют встроенное преимущество. «Это не обязательно потому, что засушливость способствует разнообразию», — сказал он. «У растений с большим содержанием азота более толстая кутикула, что делает их устойчивыми к потере воды».
Большинство линий растений, фиксирующих азот, зародились в меловом периоде, когда динозавры еще существовали, а температура была выше, чем сегодня. За последние 50 миллионов лет климат Земли постепенно охлаждался и высыхал, что привело к образованию обширных лугов и обширных пустынь. Растения, которые не могли расти в этих новых условиях, постепенно выпалывали, объясняет Доби, в то время как многие азотфиксаторы, которые хорошо подходили для этого нового мира, диверсифицировались в освободившихся ландшафтах.
«Это исследование дает нам действительно хорошее представление о том, почему растительные сообщества такие, какие они есть сегодня», — сказал Доби, добавив, что он беспокоится о том, что условия, поддерживающие разнообразную флору в засушливых регионах, могут не продлиться долго. «Поскольку из-за изменения климата становится все влажнее и теплее, черты, которые сделали эти растения хорошо адаптированными и разнообразными, больше не будут очень полезными. Многие из уникальных растительных сообществ, которые существуют сегодня, в долгосрочной перспективе окажутся под угрозой».
Авторы опубликовали свое исследование в журнале Global Ecology and Biogeography.
Роберт Гуральник из Музея естественной истории Флориды, Дайджанг Ли из Университета штата Луизиана, а также Райан Фолк и Каролина Синискалчи из Университета штата Миссисипи также являются соавторами исследования.
Финансирование исследования было частично предоставлено Институтом биоразнообразия Университета Флориды и Национальным научным фондом (грант № 1916632).