Содержание
Клеточная стенка, строение, химический состав. Видоизменения клеточной стенки
Клеточная
стенка, строение, химический
состав. Видоизменения
клеточной стенки.
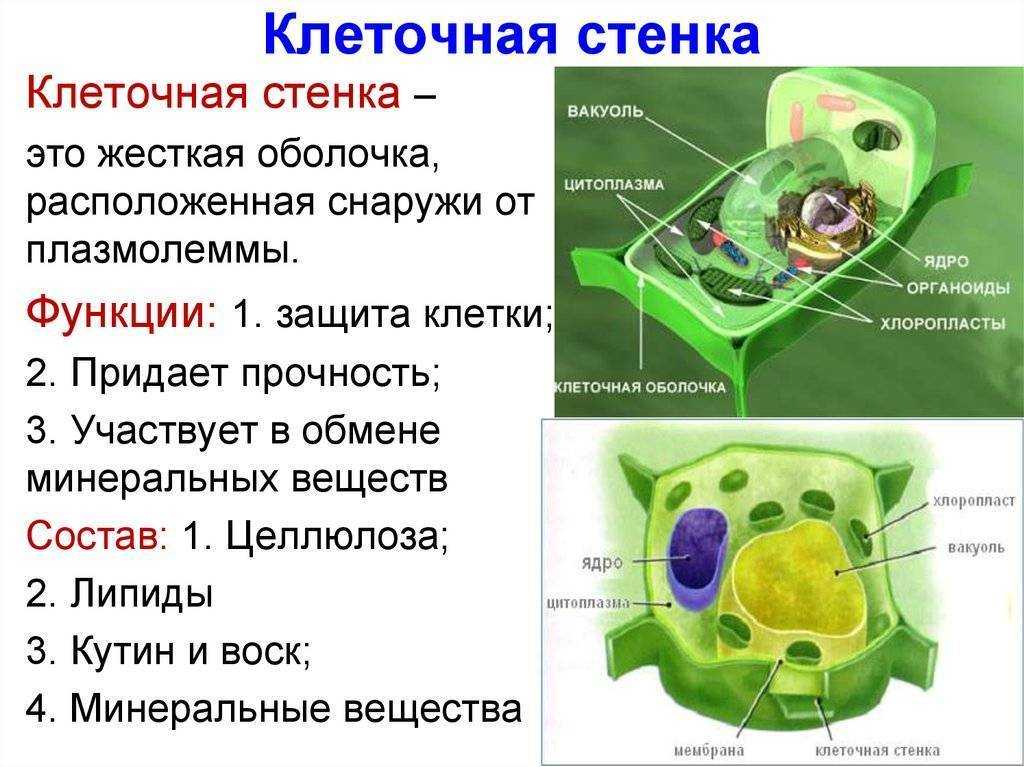
Наличие
прочной оболочки
характерная черта растительной
клетки, отличающая
ее от клетки животной.
Оболочка придает клетке
определенную форму
и прочность и защищает
живое содержимое —
протопласт и прежде
всего плазмалемму,
плотно прижатую к оболочке
изнутри. Культивируемые
на специальных питательных
средах клетки высших
растений, у которых
ферментативным путем
удаляется оболочка,
всегда принимают сферическую
форму. Без оболочки
существование растительной
клетки в обычных условиях
невозможно, так как
характерное для нее
тургорное давление,
уравновешиваемое противодавлением
оболочки, неминуемо
привело бы к разрыву
плазмалеммы и разрушению
протопласта. Совокупность
клеточных оболочек
в составе тела наземного
растения, возвышающегося
над поверхностью почвы
и не имеющего внутреннего
скелета, представляет
собой своего рода поддерживающий
остов, придающий растению
механическую прочность.
Оболочка,
как правило, бесцветна
и прозрачна, легко
пропускает солнечный
свет. По ней могут передвигаться
вода и растворенные
низкомолекулярные
вещества. У каждой клетки
есть собственная оболочка;
оболочки соседних клеток
как бы сцементированы
межклеточными веществами,
образующими так называемую
срединную пластинку.
Вследствие этого соседние
клетки оказываются
отделенными друг от
друга стенкой, образованной
двумя оболочками и
срединной пластинкой,
что дает основание
называть оболочку также
клеточной стенкой.
Оболочка
строится протопластом
клетки и поэтому
может расти, только
находясь в контакте
с ним. Очень часто оболочка
сохраняется дольше
протопласта (когда
клетка рано отмирает),
но это обычно не ведет
к прекращению существования
клетки как структурной
единицы, ибо форма клетки
благодаря прочности
оболочки не изменяется.
Поэтому растительные
клетки и после отмирания
могут выполнять важные
функции передвижения
растворов или механической
опоры. Многие типы клеток
Многие типы клеток
растения – волокна,
трахеиды, членики сосудов,
клетки пробки – во
взрослом состоянии
представляют собой
одни клеточные оболочки.
В основном из оболочек
отмерших клеток состоит
древесина.
Оболочки клеток столь
же разнообразны, как
и сами клетки. По составу
и строению оболочки
часто можно судить
о происхождении и функции
клеток различных типов.
Так, строение ископаемых
растений в основном
изучают путем исследования
оболочек их клеток.
Первоначально
к наружи от плазмалеммы
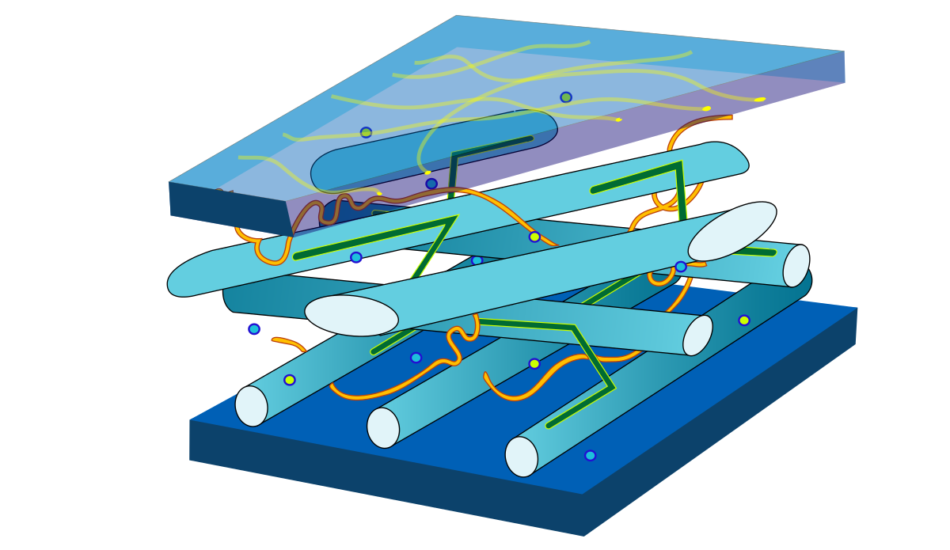
возникает первичная
клеточная стенка. Она
состоит из полисахаридов
– пектина и целлюлозы.
Первичные клеточные
стенки соседних клеток
соединены протопектиновой
срединной пластинкой.
В клеточной стенке
линейные очень длинные
молекулы целлюлозы,
состоящие из глюкозы,
собраны в пучки – мицеллы,
которые, в свою очередь,
объединяются в фибриллы
– тончайшие волоконца
неопределенной длины.
Целлюлоза образует
многомерный каркас,
который погружен в
аморфный сильно обводненный
матрикс из нецеллюлозных
углеводов: пектинов,
гемицеллюлоз и др.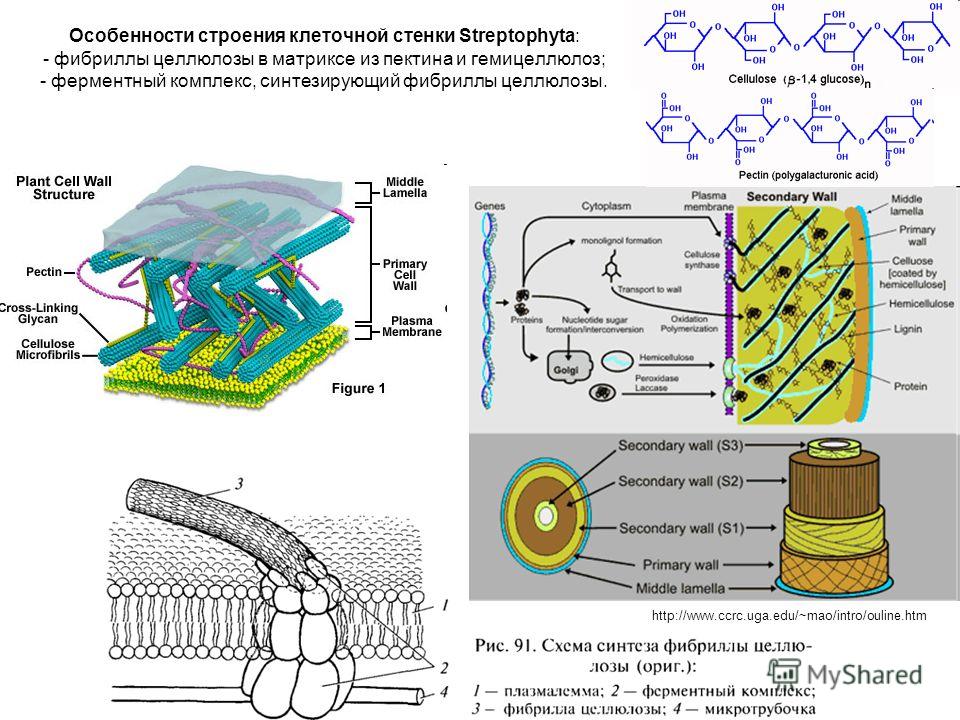 именно
именно
целлюлоза обусловливает
прочность клеточной
стенки. Микрофибриллы
эластичны и по прочности
на разрыв сходны со
сталью. Полисахариды
матрикса определяют
такие свойства стенки,
как высокая проницаемость
для воды, растворенных
мелких молекул и ионов,
сильная набухаемость.
Благодаря матриксу
по стенкам, примыкающим
друг к другу, могут
передвигаться вода
и вещества от клетки
к клетке. Некоторые
гемицеллюлозы могут
откладываться в стенках
клеток семян в качестве
запасных веществ.
Материал
оболочек, включающий
целлюлозу, широко используется
промышленностью. Свойства
древесины, бумаги, текстильных
материалов в большой
степени зависят от
молекулярной структуры
и химического состава
оболочки, и знание их
помогает улучшать технологию
производства и качество
продукции.
Химический состав и
молекулярная организация
оболочки. Клеточная
оболочка построена
в основном из полисахаридов,
мономеры которых –
сахара — связаны
между собой гликозидной
связью (—О—) в виде
цепи.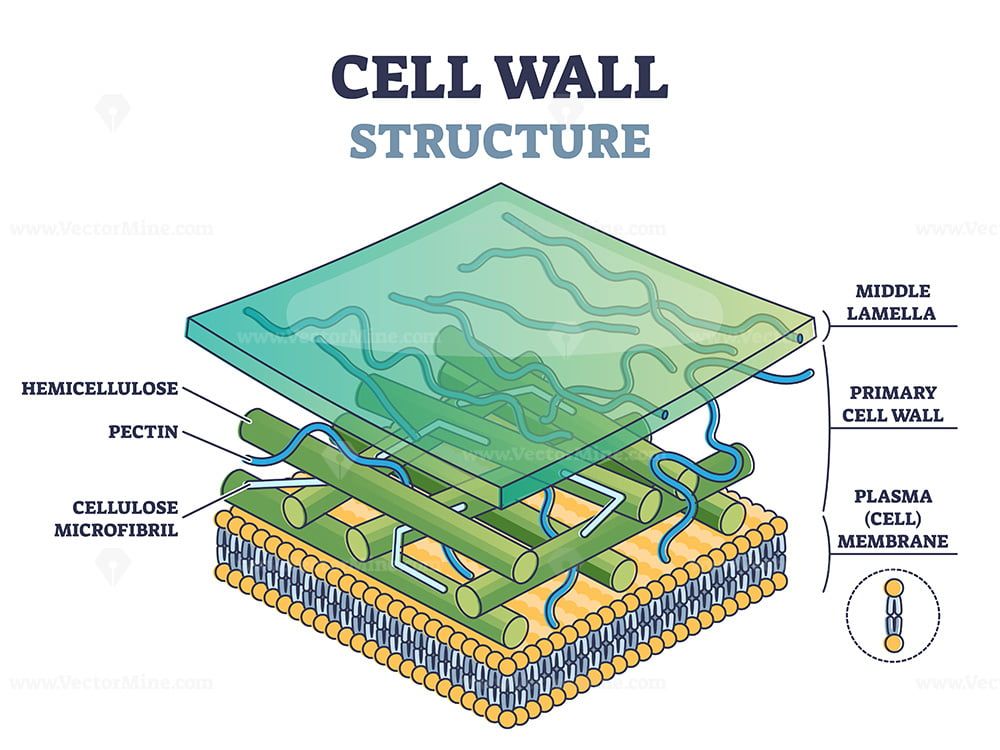 Называют полисахариды
Называют полисахариды
по составу образующих
их мономеров с прибавлением
окончания «ан».
Например, полисахарид
ксилан состоит из остатков
сахара ксилозы, маннан —
маннозы, глюкан —
глюкозы и т.д.
Кроме полисахаридов,
в состав
оболочки могут
входить белки,
минеральные соли,
лигнин, пигменты,
липиды. Обычно оболочки
пропитаны водой. Полисахариды
оболочки по своей роли
могут быть подразделены
на скелетные вещества
и вещества матрикса.
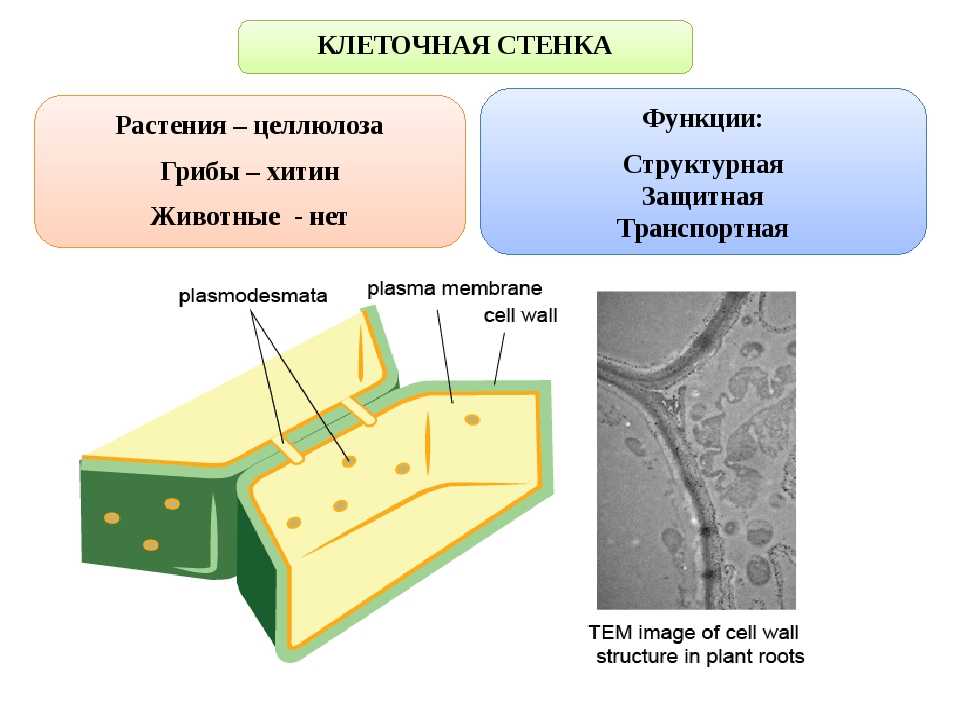
Скелетным
веществом оболочки
высших растений является
целлюлоза (клетчатка),
в химическом отношении
представляющая собой
β-1, 4-D глюкан. Число
глюкозных остатков
в молекуле и,
следовательно, длина
самих молекул могут
резко различаться в
клеточных оболочках
различных растений,
что оказывает влияние
на свойства целлюлозы.
Ее нитчатые молекулы
в оболочке располагаются
параллельно друг к
другу и группируются
по нескольку десятков
таким образом, что
возникает вытянутая
трехмерная решетка,
характерная для кристаллов.
Эти кристаллические
группировки, в образовании
которых участвуют ковалентные
и водородные связи,
составляют основу
микрофибрилл – тончайших
волокон, создающих
структурный каркас
оболочки, ее «арматуру».
Кристаллическое
состояние молекул
целлюлозы в
микрофибриллах
обусловливает
двойное лучепреломление
клеточной оболочки,
наблюдаемое
в поляризационном
микроскопе. В
микрофибриллах встречаются
паракристаллические
участки, т е. участки
с более рыхлым параллельным,
но не имеющим трехмерной
ориентации расположениемI
целлюлозных молекул.
Степень кристалличности
микрофибрилл, соотношение
кристаллической и
паракристаллической
зон довольно сильно
варьируют в зависимости
от вида растения, состояния
клетки и типа оболочки,
что оказывает влияние
на механические
и физические целлюлозы
как сырья. Диаметр микрофибрилл
также колеблется у
разных клеток (обычно
в пределах 10—30 нм),
длина может достигать
нескольких микрометров.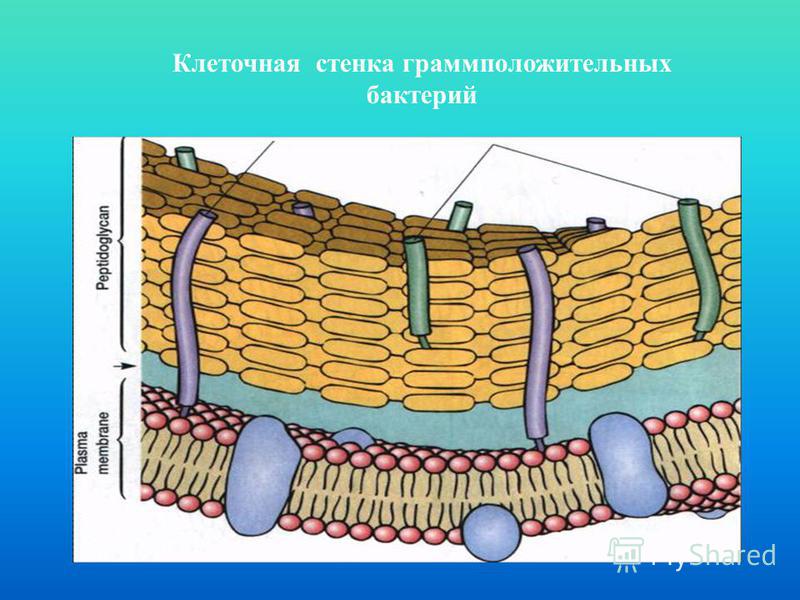
Микрофибриллы могут
объединяться в отдельные
группы или слои – макрофибриллы
толщиной 0,4-0,5 мкм, видимые
в световой микроскоп.
Рисунок
25
В
процессе жизнедеятельности
клеток, особенно в
зависимости от выполнения
какой-либо специализированной
функции, наступают
вторичные изменения
клеточной стенки.
Нередко они связаны
с радикальными изменениями
ее химического состава,
структуры и физико-химических
свойств.
Одревеснение.
Клеточная стенка
инкрустируется (пропитывается)
особым веществом
– лигнином, что
повышает твердость,
калорийность, плотность
клеточной стенки
и понижает ее пластичность
и способность
расти. Одревесневшие
клеточные стенки
не теряют способности
пропускать воду и
воздух. Протопласт
их может оставаться
живым, хотя обычно отмирает.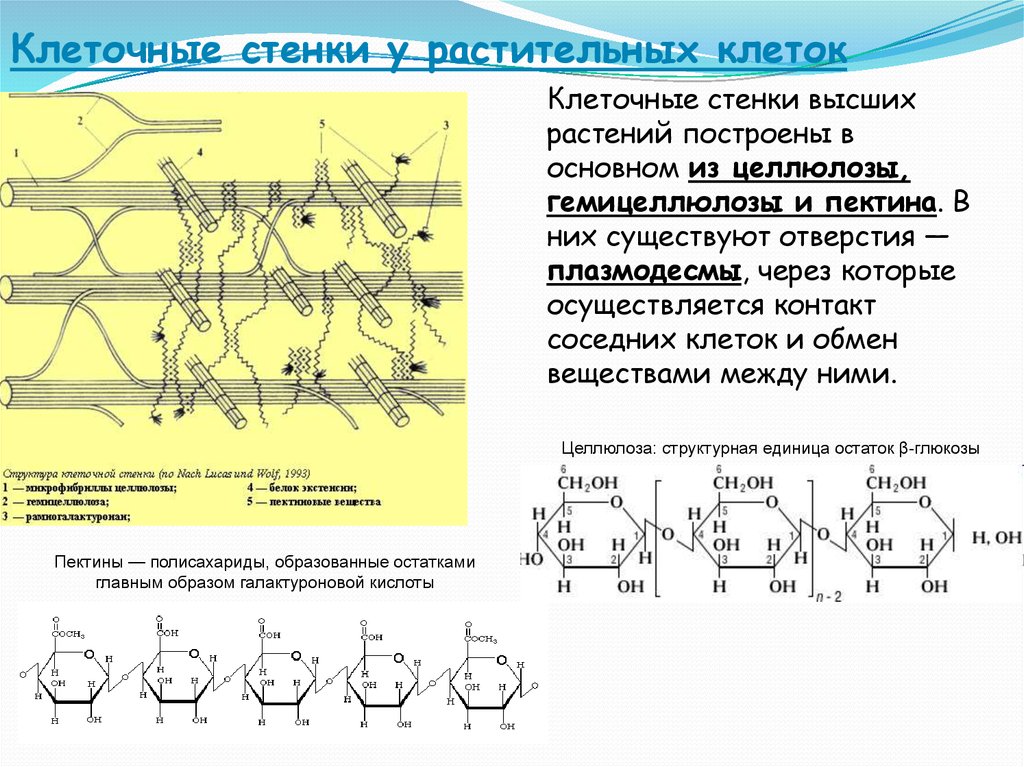
Одревеснение очень
широко распространено
в природе. Оно
обеспечивает крепость
стволов и ветвей
деревьев. Древесина
хвойных и лиственных
пород содержит целлюлозы
до 50% и лигнина 20…30%.
Одревесневают клеточные
стенки и многих трав,
особенно к концу
вегетации.
Опробковение,
или суберинизация.
В результате обильного
наслаивания в стенке
клетки химически стойкого
вещества – суберина
наступает ее опробковение.
По своей химической
природе суберин близок
к жирам. Он представляет
собой аморфное гидрофобное
соединение, которое
состоит из высокополимерных
насыщенных жирных кислот
и оксикислот сложного
состава. Суберин откладывается
преимущественно в стенках
клеток вторичной покровной
ткани – пробки, которая
является прекрасным
водо- и термоизолятором.
В малых количествах
он накапливается в
стенках клеток разнообразных
тканей. При полном опробковении
протопласт клетки отмирает,
так как ламеллы суберина
препятствуют проникновению
воды и воздуха, при
частичном – протопласт
долгое время сохраняется.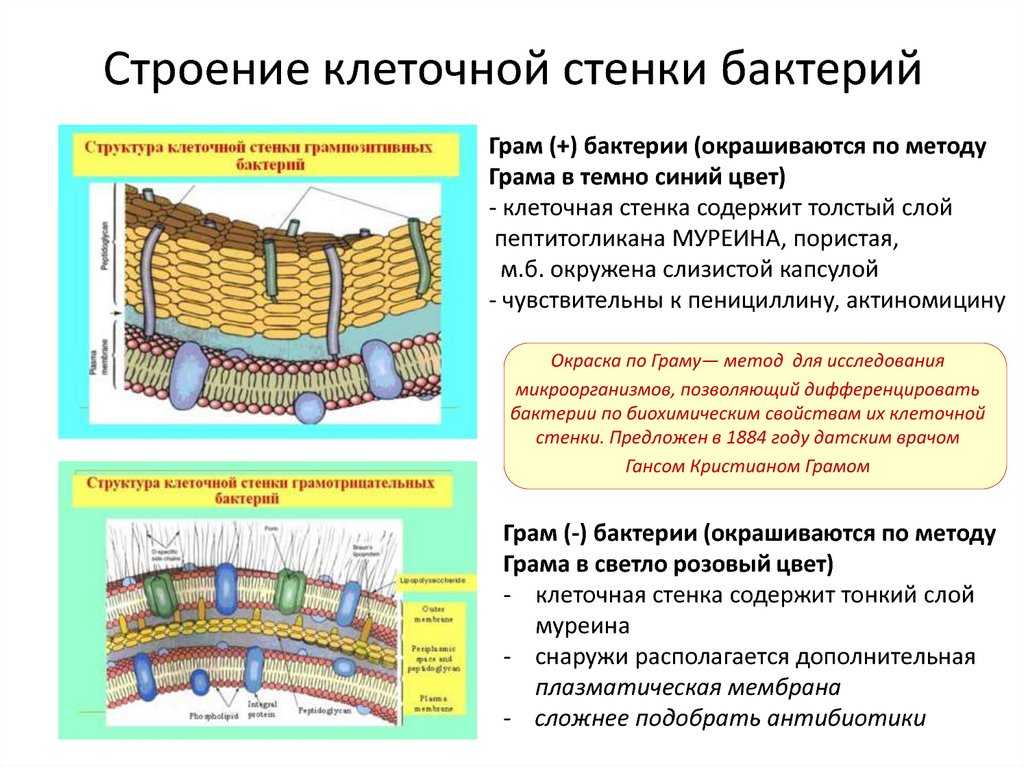
Кутинизация
– отложение кутина
– вещества, близкого
к суберину, в поверхностных
слоях наружных клеточных
стенок и на их поверхности.
Кутинизация предохраняет
органы растения от
избыточного испарения,
вымывания продуктов
метаболизма осадками
и защищает органы от
проникновения паразитов,
а также от механических
повреждений. Кроме
того, кутин поглощает
ультрафиолетовые лучи
и служит как бы радиационным
экраном.
Минерализация.
В процессе жизнедеятельности
стенки поверхностных
клеток побегов могут
накапливать минеральные
вещества, особенно
кремнезем и углекислый
кальций.некоторые группы
растений, напрмер многие
виды осок, злаков, хвощей,
всегда имеют сильно
инкрустированные солями
клетки покровной ткани
– эпидермы. Причем
инкрустации может подвергаться
не только клеточная
стенка, но и разнообразные
выросты эпидермы –
трихомы. Кроме того,
минеральные вещества
откладываются и на
поверхности клеток
покровной ткани – эпидермы.
В результате минерализации
осевые органы приобретают
большую механическую
прочность.
Ослизнение.
Превращение целлюлозы
и пектина в слизи м
близкие к ним камеди,
представляющие собой
полимерные углеводы,
которые отличаются
способностью к сильному
набуханию при соприкосновении
с водой. Ослизнение
наблюдается в клеточных
сиенках кожуры семян,
например у льна. Образование
слизей имеет большое
приспособительное
значение. При прорастании
семян слизь закрепляет
их на определенном
месте, легко поглощает
и удерживает влагу,
защищает семена от
высыхания, улучшает
водный режим всходов.
Ослизнение клеточных
стенок корневых влосков
обеспечивает прочное
склеивание их с частицами
почвы.
Мацерация
– растворение
межклеточного вещества,
приводящее к разъединению
клеток. Естественная
мацерация происходит
в зрелых плодах.
Искусственно ее проводят,
например, при мочке
льна для освобождения
прядильного сырья
– групп клеток
лубяных волокон.
D-рамнан и пируватсодержащая тейхуроновая кислота клеточной стенки Rathayibacter sp. ВКМ Ac-2759 – выпуск 4, том 86, 2021 – Биохимия
БИОХИМИЯ, 2021, том 86, вып. 4, с. 595–606
УДК 577.124
© 2021 А.С. Шашков 1, Е.М. Тульская 2*[email protected], Н.В. Потехина 2, А.С. Дмитренок 1, С.Н. Сенченкова 1, В.А. Зайчиков 2, Л.В. Дорофеева 3, Л.И. Евтушенко 3
1 Институт органической химии им. Н.Д. Зелинского РАН, 119991 Москва, Россия
2 Московский государственный университет имени М.В. Ломоносова, биологический факультет, 119234 Москва, Россия
3 Всероссийская коллекция микроорганизмов (ВКМ), Институт биохимии и физиологии микроорганизмов им. Г.К. Скрябина, ФИЦ Пущинский научный центр биологических исследований РАН, 142290 Пущино, Московская обл., Россия
Поступила в редакцию 18.01.2021
После доработки 16.02.2021
Принята к публикации 16.02.2021
DOI: 10.31857/S0320972521040114
КЛЮЧЕВЫЕ СЛОВА: Rathayibacter, клеточная стенка, гликополимеры, D-рамнан, тейхуроновая кислота, пируват.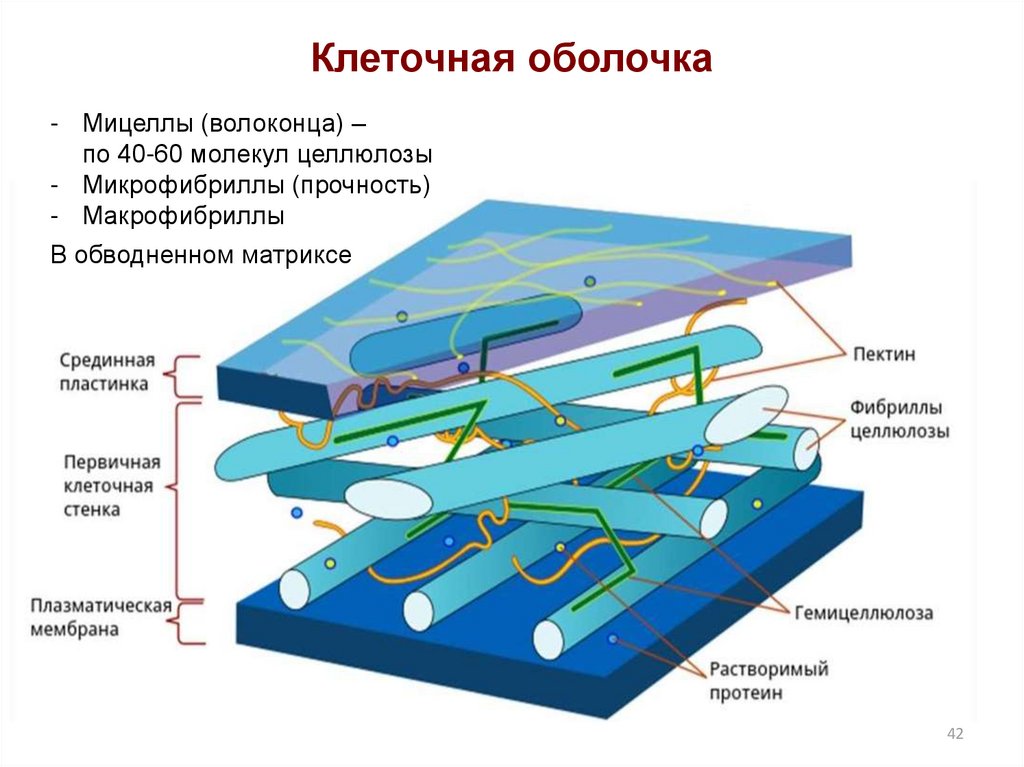
Аннотация
Rathayibacter sp. ВКМ Ac-2759 (семейство Microbacteriaceae, класс Actinobacteria) содержит в клеточной стенке два гликополимера. Основная цепь рамнана, гликополимера 1, построена из повторяющихся тетрасахаридных звеньев, несущих на нередуцирующем конце терминальные остатки арабинофуранозы: →3)-α-[α-D-Araf-(1→2)]-D-Rhap-(1→2)-α-D-Rhap-(1→3)-α-D-Rhap-(1→2)-α-D-Rhap-(1→. Подобно другим изученным видам рода Rathayibacter, в нейтральном гликополимере штамма ВКМ Ac-2759 рамноза присутствует в D-конфигурации. Ацеталированная пировиноградной кислотой тейхуроновая кислота, гликополимер 2, состоит из линейных тетрасахаридных звеньев: →4)-β-D-GlcpA-(1→4)-β-D-Galp-(1→4)-β-D-Glcp-(1→3)-β-[4,6-S-Pyr]-D-Manp-(1→. Структуры гликополимеров установлены химическими и ЯМР-спектроскопическими методами и описаны впервые для прокариотных микроорганизмов. Полученные данные могут быть востребованы в таксономических исследованиях и работах по выяснению механизмов колонизации и инфекции растений бактериями рода Rathayibacter.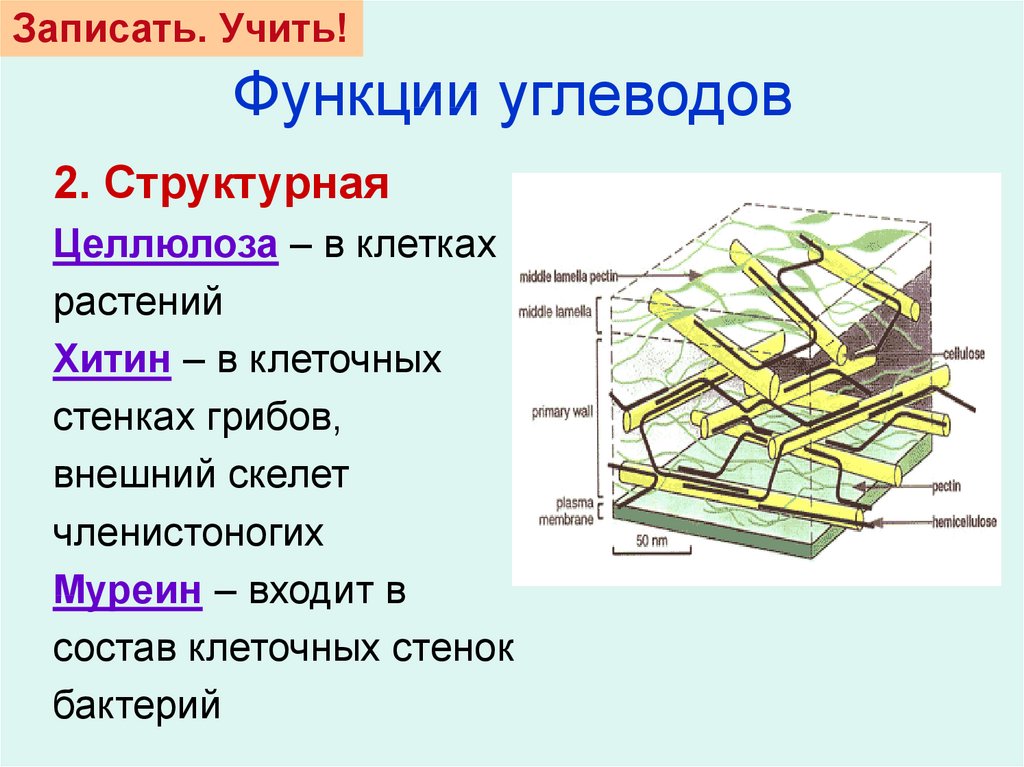
Сноски
* Адресат для корреспонденции.
Финансирование
Работа выполнялась в рамках программы исследований № ЦИТИС: АААА-А16-116021660068-1, запланированных в МГУ имени М.В. Ломоносова, кафедра микробиологии 2016–2022 гг.
Благодарности
Авторы благодарны Галине Матвеевне Стрешинской за помощь в исследованиях и обсуждении статьи.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Соблюдение этических норм
Настоящая статья не содержит описания выполненных авторами исследований с участием людей или использованием животных в качестве объектов.
Список литературы
1. Zgurskaya, H. I., Evtushenko, L. I., Akimov, V. N., and Kalakoutskii, L. V. (1993) Rathayibacter gen, nov., including the species Rathayibacter rathayi comb. nov., Rathayibacter tritici comb. nov., Rathayibacter iranicus comb., nov., and six strains from annual grasses, Int. J. Sys. Bacteriol. , 43, 143-149, doi: 10.1099/00207713-43-1-143.
, 43, 143-149, doi: 10.1099/00207713-43-1-143.
2. Evtushenko, L. I., and Dorofeeva, L. V. (2012) Genus XXII. Rathayibacter Zgurskaya, Evtushenko, Akimov and Kalakoutskii 1993, 147VP in Bergey’s manual of systematic bacteriology (Goodfellow, M., Kämpfer, P., Busse, H.-J., Trujillo, M. E., Suzuki, K.-I., Ludwig, W., Whitman, W. B., eds.) 2nd Edn., Vol. 5, Springer, New York, Dordrecht, Heidelberg, London, pp. 953-964.
3. Murray, T. D., Schroeder, B. K., Schneider, W. L., Luster, D. G., Sechler, A., et al. (2017) Rathayibacter toxicus, other Rathayibacter species inducing bacterial head blight of grasses, and the potential for livestock poisonings, Phytopathology, 107, 804-815, doi: 10.1094/PHYTO-02-17-0047-RVW.
4. Murray, T. D., Barrantes-Infante, B., and Schroeder, B. K. (2020) First report of bacterial head blight of Pseudoroegneria spicata subsp. spicata caused by Rathayibacter agropyri in Idaho, Plant Disease, 104, 1534, doi: 10.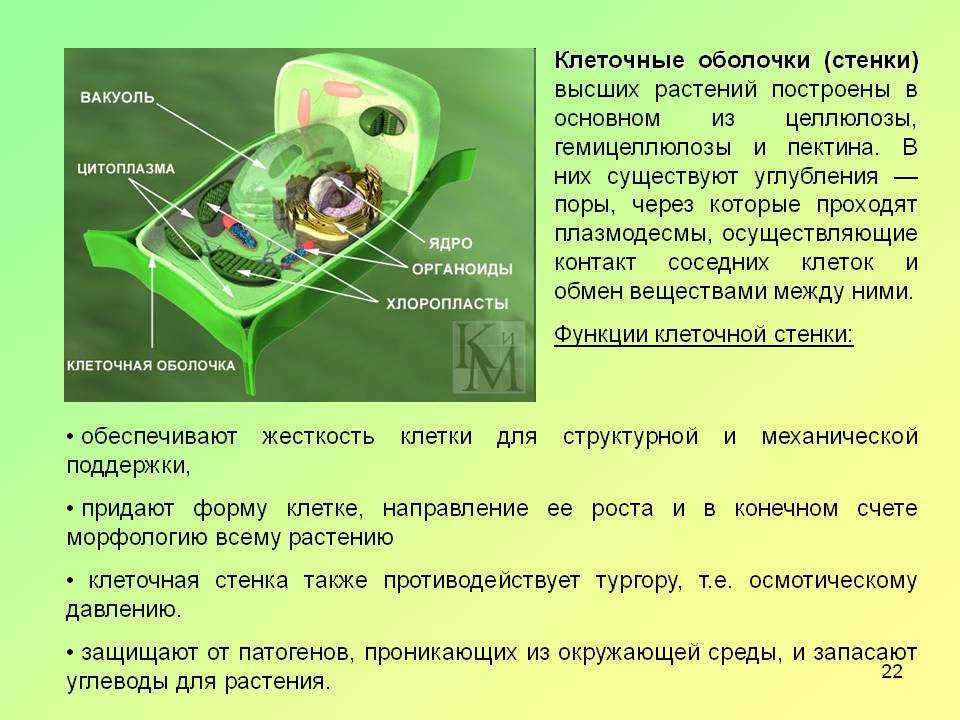 1094/PDIS-06-19-1233-PDN.
1094/PDIS-06-19-1233-PDN.
5. Starodumova, I. P. (2018) Development of the classification system for actinobacteria of the genus Rathayibacter, candidate dissertation (in Russian), Moscow.
6. Shashkov, A. S., Tul’skaya, E. M., Dmitrenok, A. S., Streshinskaya, G. M., Potekhina, N. V., et al. (2018) Rhamnose-containing cell wall glycopolymers from Rathayibacter toxicus VKM Ac-1600 and “Rathayibacter tanaceti” VKM Ac-2596, Biochemistry (Moscow), 83, 717-726, doi: 10.1134/S0006297918060093.
7. Dmitrenok, A. S., Shashkov, A. S., Streshinskaya, G. M., Tul’skaya, E. M., Potekhina, N. V., et al. (2019) New rhamnose-contaning glycopolymers from Rathayibacter iranicus VKM Ac-1602T cell wall, Carbohydr. Res., 482, 107728, doi: 10.1016/j.carres.2019.06.007.
8. Shashkov, A. S., Tul’skaya, E. M., Streshinskaya, G. M., Dmitrenok, A. S., Potekhina, N. V., et al. (2020) Rhamnomannans and teichuronic acid from the cell wall of Rathayibacter tritici VKM Ac-1603T, Biochemistry (Moscow), 85, 369-377, doi: 10.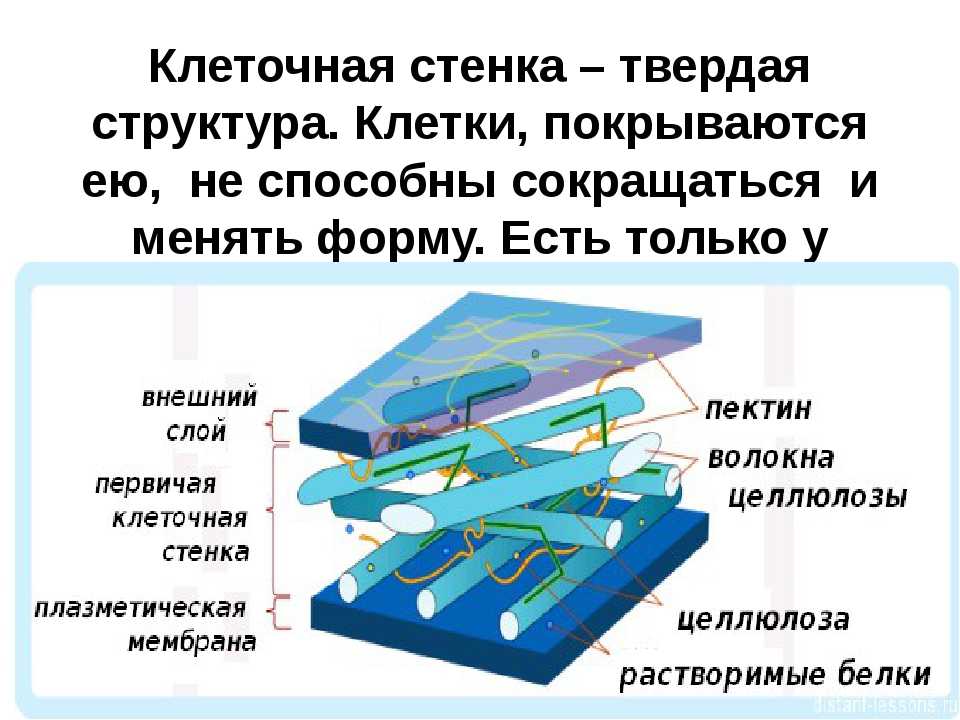 1134/S0006297920030128.
1134/S0006297920030128.
9. Shashkov, A. S., Potekhina, N. V., Dmitrenok, A. S., Tul’skaya, E. M., Senchenkova, S. N., et al. (2021) D-Rhamnan and teichuronic acid from the cell wall of Rathayibacter caricis VKM Ac-1799T, Carbohydr. Res., 499, 108233.
10. Tarlachkov, S. V., Starodumova, I. P., Dorofeeva, L. V., Prisyazhnaya, N. V., Leyn, S. A., et al. (2020) Complete and draft genome sequences of 12 plant-associated Rathayibacter strains of known and putative new species, Microbiol. Resour. Announc., 9, e00316-20, doi: 10.1128/MRA.00316-20.
11. Potekhina, N. V., Streshinskaya, G. M., Tul’skaya, E. M., and Shashkov, A. S. (2011) Cell wall teichoic acids in the taxonomy and characterization of Gram-positive bacteria, in Methods in Microbiology (Rainey, F. A., and Oren, A., eds.) Vol. 38, Chap. 6, Academic Press, Elsevier, pp. 132-164, doi: 10.1016/B978-0-12-387730-7.00020-6.
12. Leontein, K., Lindberg, B., and Lönngren, J. (1978) Assignment of absolute configuration of sugars by glc of their acetylated glycosides formed from chiral alcohols, Carbohyd. Res., 62, 359-362, doi: 10.1016/S0008-6215(00)80882-4.
Leontein, K., Lindberg, B., and Lönngren, J. (1978) Assignment of absolute configuration of sugars by glc of their acetylated glycosides formed from chiral alcohols, Carbohyd. Res., 62, 359-362, doi: 10.1016/S0008-6215(00)80882-4.
13. Gerwig, G. J., Kamerling, I. P., and Vliegenthart, J. F. G. (1979) Determination of the absolute configuration of monosaccharides in complex carbohydrates by capillary GLC, Carbohydr. Res., 77, 1-7, doi: 10.1016/s0008-6215(00)83788-x.
14. Shashkov, A. S., Lipkind, G. M., Knirel, Y. A., and Kochetkov, N. K. (1988) Stereometrical factors determining the effects of glycosylation on the 13C chemical shifts in carbohydrates, Magn. Reson. Chem., 26, 735-747, doi: 10.1002/mrc.1260260904.
15. Garegg, P. J., Jansson, P.-E., Lindberg, B., Lindh, F., Lönngren, J. (1980) Configuration of the acetal carbon atom of pyruvic acid acetals in some bacterial polysaccharides, Carbohydr. Res., 79, 127-132, doi: 10.1016/S0008-6215(00)83666-6.
Res., 79, 127-132, doi: 10.1016/S0008-6215(00)83666-6.
16. Mistou, M.-Y., Sutcliffe, I. C., and van Sorge, N. M. (2016) Bacterial glycobiology: rhamnose-containing cell wall polysaccharides in Gram-positive bacteria, FEMS Microbiol. Rev., 40, 464-479, doi: 10.1093/femsre/fuw006.
17. Knirel, Y. A. (2010) O-Specific polysaccharides of Gram-negative bacteria, in Microbial Glycobiology (Holst, O., Brennan, P. J., von Itzstein, M., eds.) Vol. 4, Academic Press, pp. 57-73, doi: 10.1016/B978-0-12-374546-0.X0001-6.
18. Schaffer, C., Muller, N., Christian, R., Graninger, M., Wugeditsch, T., et al. (1999) Complete glycan structure of the S-layer glycoprotein of Aneurinibacillus thermoaerophilus GS4-97, Glycobiology, 9, 407-414, doi: 10.1093/glycob/9.4.407.
19. Kaczynski, Z., Karapetyan, G., Evidente, A., Iacobellis, N. S., and Holst, O. (2006) The structure of a putative exopolysaccharide of Burkholderia gladioli pv.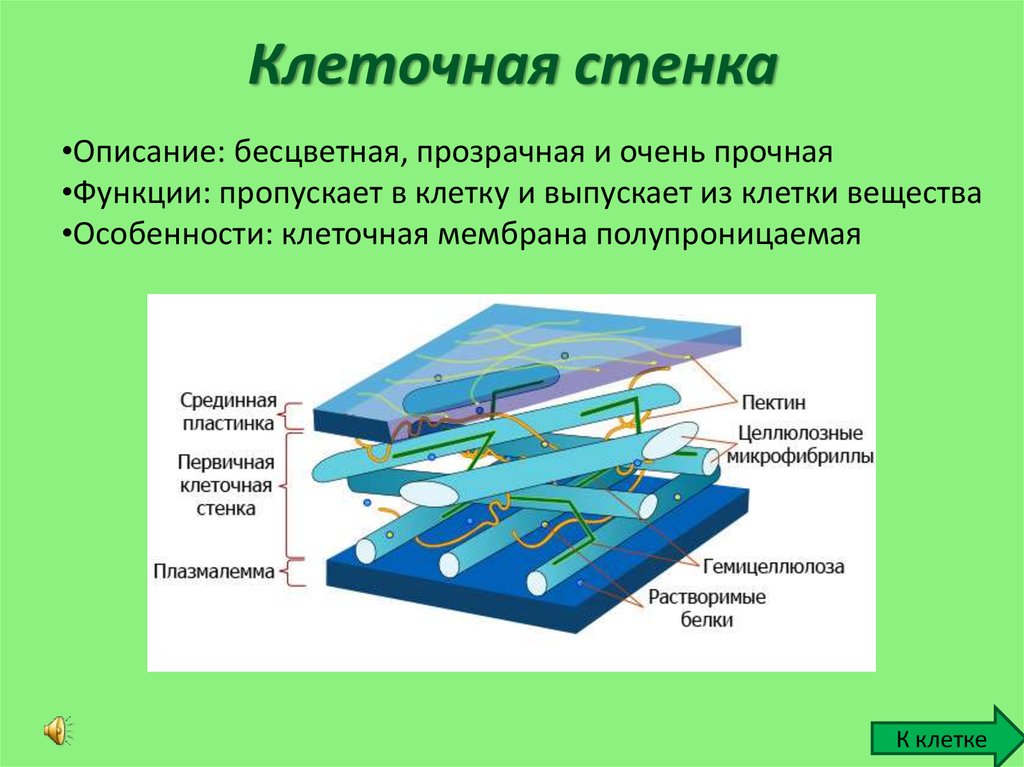 agaricicola, Carbohydr. Res., 341, 285-288, doi: 10.1016/j.carres.2005.10.020.
agaricicola, Carbohydr. Res., 341, 285-288, doi: 10.1016/j.carres.2005.10.020.
20. Cimmino, A., Marchi, G., Hanuszkiewicz, A., Surico, G., Evidente, A., and Holst, O. (2008) The structure of the O-specific polysaccharide of the lipopolysaccharide from Pantoea agglomerans strain FL1, Carbohydr. Res., 343, 392-396, doi: 10.1016/j.carres.2007.10.023.
| |||||||||||||||||||||||
Ключевое слово SearchAdvanced Search Поисковые книги для: Советы по поиску
96696. Show all books public access books [?] | |||||||||||||||||||||||
 7: 7006677777776 гг. 200520042003200220012000199919981997199619951994199319921991199019891988198719861985198419831982198119801979197819771976197519741973197219711970196919681967196619651964196319621961196019591958195719561955195419531952195119501949194819471946194519441943194219411940 to 200520042003200220012000199919981997199619951994199319921991199019891988198719861985198419831982198119801979197819771976197519741973197219711970196919681967196619651964196319621961196019591958195719561955195419531952195119501949194819471946194519441943194219411940
7: 7006677777776 гг. 200520042003200220012000199919981997199619951994199319921991199019891988198719861985198419831982198119801979197819771976197519741973197219711970196919681967196619651964196319621961196019591958195719561955195419531952195119501949194819471946194519441943194219411940 to 200520042003200220012000199919981997199619951994199319921991199019891988198719861985198419831982198119801979197819771976197519741973197219711970196919681967196619651964196319621961196019591958195719561955195419531952195119501949194819471946194519441943194219411940Клеточная мембрана — клеточная стенка
Снаружи клетки многих видов клеток покрыты жесткой структурой, называемой клеточной стенкой, независимо от того, являетесь ли вы растением, грибком, многими бактериями, а также протестной группой. называются водорослями. Теперь клеточная стенка состоит из материала, называемого целлюлозой, хитином или пептидогликаном.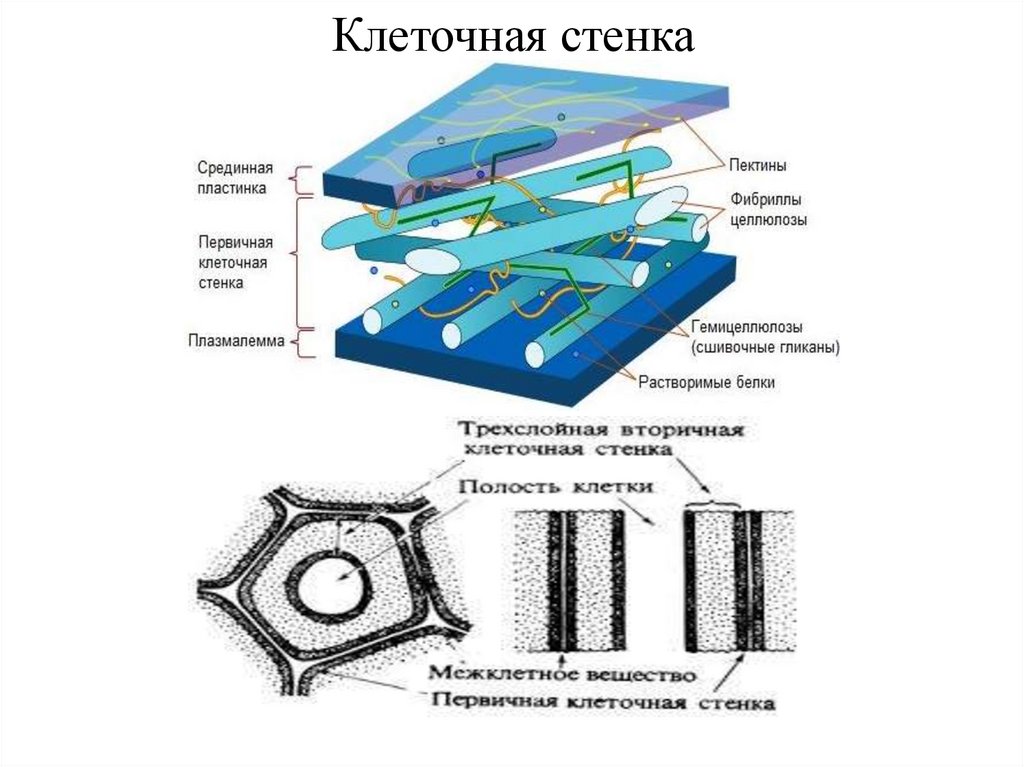 Там есть другие. Некоторые водоросли даже используют стекло и диоксид кремния в качестве своих клеточных стенок, но в целом все они обеспечивают структуру и поддержку клетки, находящейся внутри этой клеточной стенки.
Там есть другие. Некоторые водоросли даже используют стекло и диоксид кремния в качестве своих клеточных стенок, но в целом все они обеспечивают структуру и поддержку клетки, находящейся внутри этой клеточной стенки.
Давайте быстро взглянем на эту растительную клетку, и вы увидите толстую внешнюю клеточную стенку, которая помогает удерживать клетку внутри, защищая ее, а также, поскольку у них нет скелетов, как у вас и у меня, если бы они их не имели. Имейте эту жесткую конструкцию снаружи, они просто рухнут, и вместо высоких деревьев у вас будут просто маленькие капли зеленой слизи. Теперь, если мы вернемся сюда, клеточная мембрана, которую часто называют плазматической мембраной, — это то, что вы найдете непосредственно под этой клеточной стенкой, и она состоит из чего-то, называемого фосфолипидным бислоем, о котором я расскажу позже. Теперь фосфолипидный бислой является одним из ключевых элементов, позволяющих клеточной или плазматической мембране контролировать то, что может входить в клетку или выходить из нее.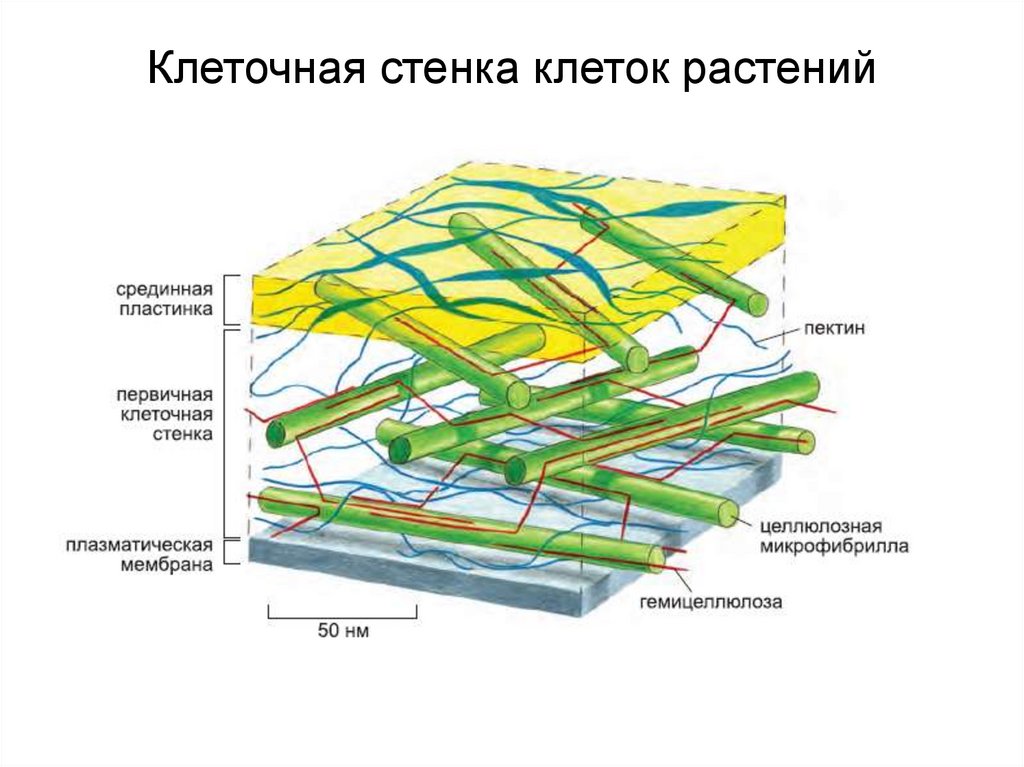 На самом деле, из-за некоторых других вещей, встроенных в фосфолипидный бислой, он также способен изменять то, что он может пропустить, от момента к моменту, что придает ему не только полупроницаемую природу фосфолипидного двойного слоя, но и фактически подмножество называется избирательно проницаемым барьером.
На самом деле, из-за некоторых других вещей, встроенных в фосфолипидный бислой, он также способен изменять то, что он может пропустить, от момента к моменту, что придает ему не только полупроницаемую природу фосфолипидного двойного слоя, но и фактически подмножество называется избирательно проницаемым барьером.
Еще одна вещь, которая вызвана не фосфолипидами, а некоторыми другими молекулами, входящими в состав мембраны, заключается в том, что она участвует в межклеточном взаимодействии. Итак, еще раз, если мы посмотрим на нашу растительную клетку, вот клеточная стенка. Сразу под ним находится тонкий бислой фосфолипидов. Теперь это называется бислоем фосфолипидов, потому что эта структура здесь и эта структура здесь, каждая из них является фосфолипидом, и вы видите один, два слоя, ну да.
Фосфолипид — это особый тип липида или молекулы жира, один конец которого имеет фосфатную головку. Фосфат — это ион, который имеет сильный отрицательный заряд. Эти маленькие волнистые кусочки на конце — это хвосты из двух жирных кислот, их иногда называют.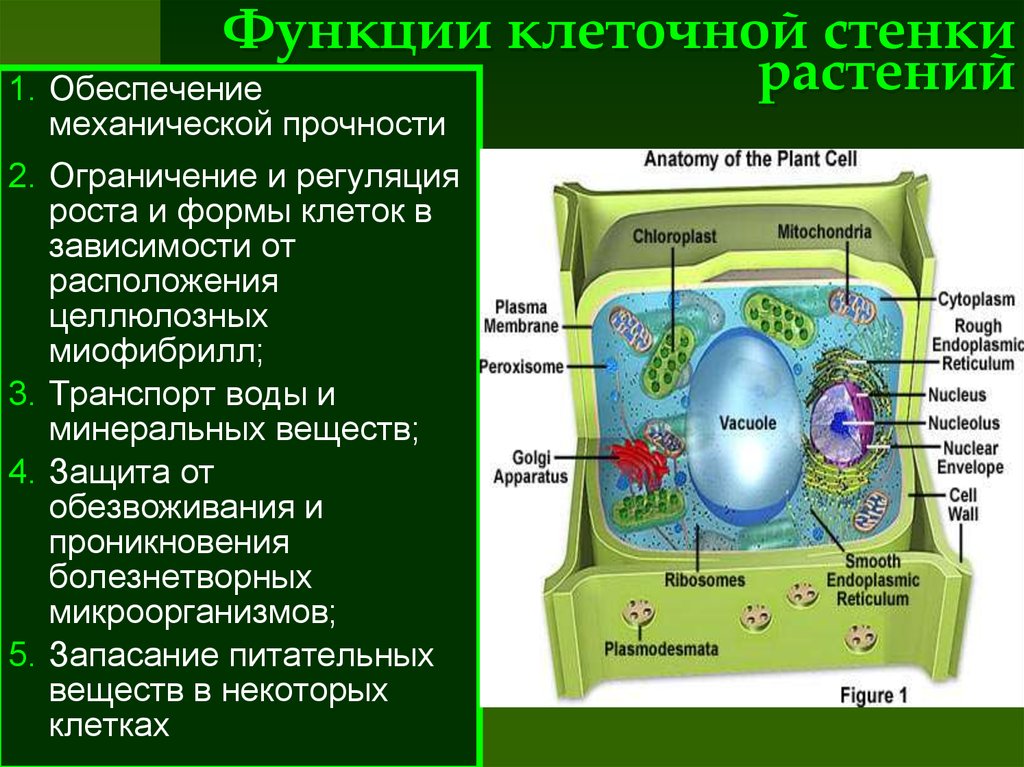 Я часто думал, что они выглядят как маленькие ножки какого-то парня в плавательном жилете, который просто плавает в воде. Эти ноги — жирная часть, а жиры, как вы могли видеть, если вам когда-либо приходилось мыть посуду, жиры не смешиваются с водой. Вот они прячутся от воды внутри и снаружи камеры. Я часто думал о фосфолипидах, фосфатах, которые могут взаимодействовать с водой, и хвостах жирных кислот, которые не могут, это как вы и двое ваших младших братьев и сестер. Ты классный, ты классный. Все хотят взаимодействовать с вами, все являются водой. Вы можете установить с ними водородную связь, когда взаимодействуете с ними на вечеринке.
Я часто думал, что они выглядят как маленькие ножки какого-то парня в плавательном жилете, который просто плавает в воде. Эти ноги — жирная часть, а жиры, как вы могли видеть, если вам когда-либо приходилось мыть посуду, жиры не смешиваются с водой. Вот они прячутся от воды внутри и снаружи камеры. Я часто думал о фосфолипидах, фосфатах, которые могут взаимодействовать с водой, и хвостах жирных кислот, которые не могут, это как вы и двое ваших младших братьев и сестер. Ты классный, ты классный. Все хотят взаимодействовать с вами, все являются водой. Вы можете установить с ними водородную связь, когда взаимодействуете с ними на вечеринке.
Твои младшие братья и сестры, никто не хочет с ними общаться, и им на самом деле наплевать на общение с кем-либо еще. Они просто хотят сидеть и смотреть на свои карты покемонов или на то, чем они увлекаются. Так что эта фосфатная голова взаимодействует с водой, а покемоны здесь нет, и что вы делаете, так это то, что если вы идете на вечеринку, и ваши родители приковывают к вам своих младших братьев и сестер, вы прячете их за собой.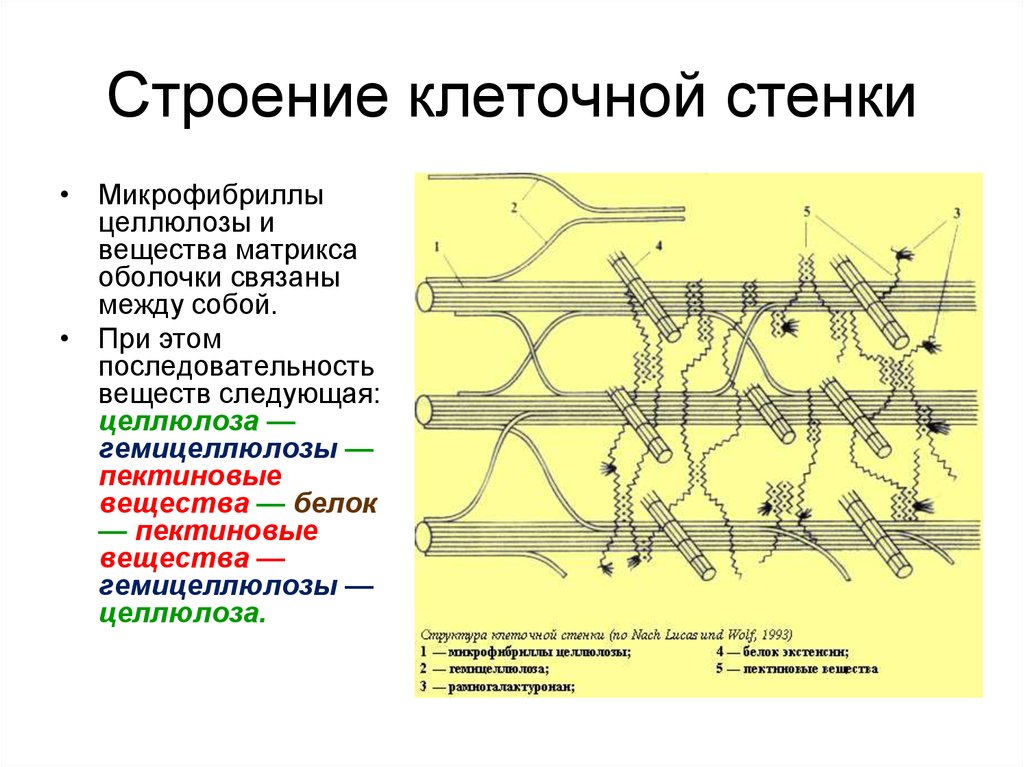 И вы можете сформировать стену из старших братьев и сестер, а младшие братья и сестры прячутся за ними, и фактически вы можете сформировать здесь этот двойной слой. Все, что пытается пройти, должно иметь возможность взаимодействовать с хвостами жирных кислот, а не слишком много химических веществ могут. Вы должны быть незаряжены и, как правило, должны быть маленькими. И большинство молекул не могут этого сделать. Именно поэтому он непроницаем для большинства молекул, то есть они не могут проникнуть сквозь него. Некоторые молекулы могут, например газообразный кислород и газообразный углекислый газ. Они могут проникать прямо через него, потому что иногда некоторые вещи могут проникать, а другие нет, это называется полупроницаемостью.
И вы можете сформировать стену из старших братьев и сестер, а младшие братья и сестры прячутся за ними, и фактически вы можете сформировать здесь этот двойной слой. Все, что пытается пройти, должно иметь возможность взаимодействовать с хвостами жирных кислот, а не слишком много химических веществ могут. Вы должны быть незаряжены и, как правило, должны быть маленькими. И большинство молекул не могут этого сделать. Именно поэтому он непроницаем для большинства молекул, то есть они не могут проникнуть сквозь него. Некоторые молекулы могут, например газообразный кислород и газообразный углекислый газ. Они могут проникать прямо через него, потому что иногда некоторые вещи могут проникать, а другие нет, это называется полупроницаемостью.
Итак, я упоминал ранее, что клеточная мембрана на самом деле избирательно проницаема, потому что она может изменить то, что разрешено и что не может войти. Вот как на самом деле будет выглядеть ваша клеточная мембрана. Здесь эти фиолетовые шарики представляют белок, встроенный в мембрану. Здесь мы видим наш бислой фосфолипидов. Эти маленькие оранжевые штучки представляют собой молекулы стероидов или холестерина, которые помогают, придают некоторую жесткость мембране. И они текут вокруг. Это иногда называют моделью жидкой мозаики, потому что она текучая и состоит из маленьких кусочков, очень похожих на мозаику.
Здесь мы видим наш бислой фосфолипидов. Эти маленькие оранжевые штучки представляют собой молекулы стероидов или холестерина, которые помогают, придают некоторую жесткость мембране. И они текут вокруг. Это иногда называют моделью жидкой мозаики, потому что она текучая и состоит из маленьких кусочков, очень похожих на мозаику.
Эти белки, некоторые из них являются насосами или каналами. Насосы, как следует из названия, могут качать вещи в клетку или из нее, если она тратит некоторую энергию. У каналов есть маленькие дверцы, которые они могут открывать или закрывать, открывать, когда они хотят, чтобы что-то вошло, или закрывать, когда они этого не хотят.
Теперь некоторые из этих других вещей, которые прикреплены, являются сахарным тростником, полисахаридами, которые, особенно когда они присоединены к белкам, они называются гликопротеинами, когда они прикреплены к некоторым из фосфатных головок здесь, они называются гликолипидами, потому что фосфолипид — это разновидность липидов.