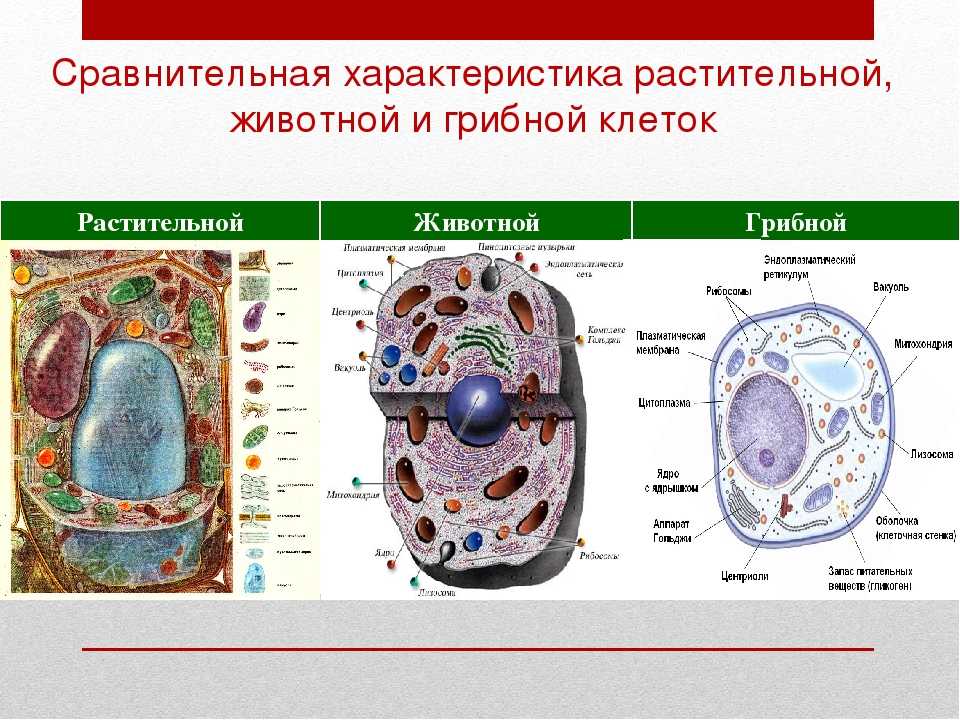
Содержание
Строение животной клетки
- Общие термины
Содержание
- Основные свойства клетки:
- Структуры живой клетки
- Сходства растительных и животных клеток
- Существенные различия между растительной и животной клеткой
Клетки животных и растений, как многоклеточных, так и одноклеточных, в принципе сходны по своему строению. Различия в деталях строения клеток связаны с их функциональной специализацией.
Основными элементами всех клеток являются ядро и цитоплазма. Ядро имеет сложное строение, изменяющееся на разных фазах клеточного деления, или цикла. Ядро неделящейся клетки занимает приблизительно 10–20% ее общего объема. Оно состоит из кариоплазмы (нуклеоплазмы), одного или нескольких ядрышек (нуклеол) и ядерной оболочки. Кариоплазма представляет собой ядерный сок, или кариолимфу, в которой находятся нити хроматина, образующие хромосомы.
Основные свойства клетки:
- обмен веществ
- чувствительность
- способность к размножению
Клетка живет во внутренней среде организма – кровь, лимфа и тканевая жидкость. Основными процессами в клетке являются окисление, гликолиз – расщепление углеводов без кислорода. Проницаемость клетки избирательна. Она определяется реакцией на высокую или низкую концентрацию солей, фаго- и пиноцитоз. Секреция – образование и выделение клетками слизеподобных веществ (муцин и мукоиды), защищающие от повреждения и участвующие в образовании межклеточного вещества.
Основными процессами в клетке являются окисление, гликолиз – расщепление углеводов без кислорода. Проницаемость клетки избирательна. Она определяется реакцией на высокую или низкую концентрацию солей, фаго- и пиноцитоз. Секреция – образование и выделение клетками слизеподобных веществ (муцин и мукоиды), защищающие от повреждения и участвующие в образовании межклеточного вещества.
Виды движений клетки:
- амебоидное (ложноножки) – лейкоциты и макрофаги.
- скользящее – фибробласты
- жгутиковый тип – сперматозоиды (реснички и жгутики)
Деление клеток:
- непрямое (митоз, кариокинез, мейоз)
- прямое (амитоз)
При митозе ядерное вещество распределяется равномерно между дочерними клетками, т.к. хроматин ядра концентрируется в хромосомах, которые расщепляются на две хроматиды, расходящиеся в дочерние клетки.
Структуры живой клетки
Хромосомы
Обязательными элементами ядра являются хромосомы, имеющие специфическую химическую и морфологическую структуру. Они принимают активное участие в обмене веществ в клетке и имеют прямое отношение к наследственной передаче свойств от одного поколения к другому. Следует, однако, иметь в виду, что, хотя наследственность и обеспечивается всей клеткой как единой системой, ядерные структуры, а именно хромосомы, занимают при этом особое место. Хромосомы, в отличие от органелл клетки, представляют собой уникальные структуры, характеризующиеся постоянством качественного и количественного состава. Они не могут взаимозаменять друг друга. Несбалансированность хромосомного набора клетки приводит в конечном счете к ее гибели.
Они принимают активное участие в обмене веществ в клетке и имеют прямое отношение к наследственной передаче свойств от одного поколения к другому. Следует, однако, иметь в виду, что, хотя наследственность и обеспечивается всей клеткой как единой системой, ядерные структуры, а именно хромосомы, занимают при этом особое место. Хромосомы, в отличие от органелл клетки, представляют собой уникальные структуры, характеризующиеся постоянством качественного и количественного состава. Они не могут взаимозаменять друг друга. Несбалансированность хромосомного набора клетки приводит в конечном счете к ее гибели.
Цитоплазма
50 фактов про кенгуру
Ушастый ёж
Паук-крестовик
Пресмыкающиеся
72 факта о растениях
Животные Сибири
Цитоплазма клетки обнаруживает весьма сложное строение. Введение методики тонких срезов и электронной микроскопии позволило увидеть тонкую структуру основной цитоплазмы.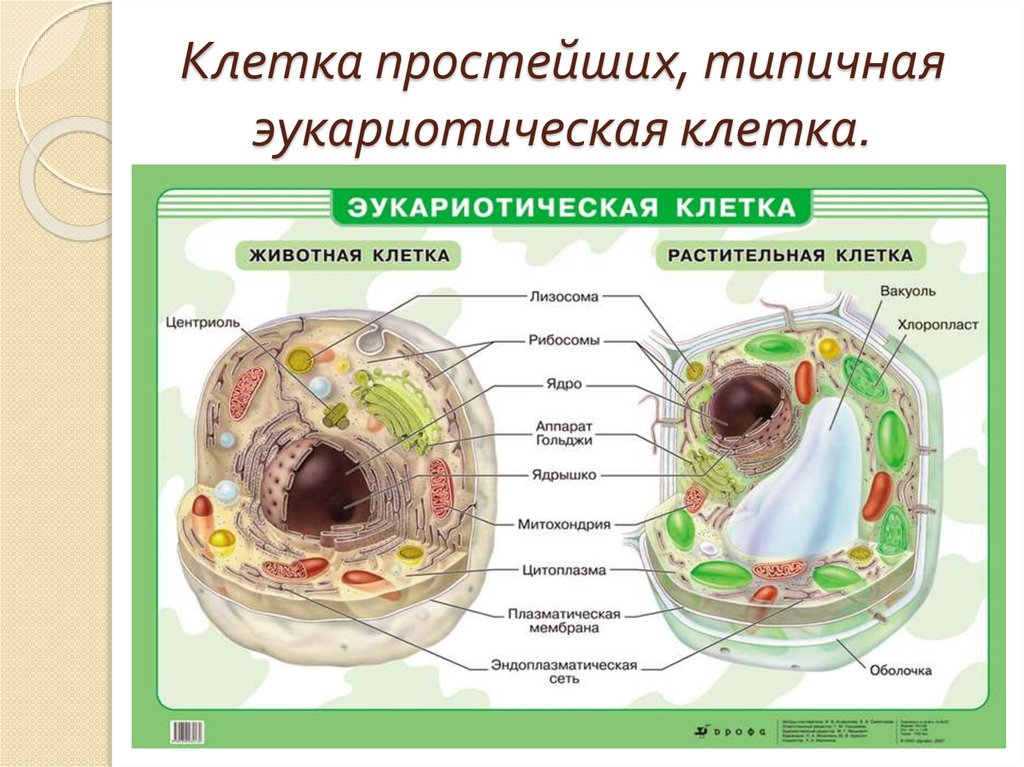 Установлено, что последняя состоит из параллельно расположенных сложных структур, имеющих вид пластинок и канальцев, на поверхности которых располагаются мельчайшие гранулы диаметром 100–120 Å. Эти образования названы эндоплазматическим комплексом. В состав этого комплекса включены различные дифференцированные органоиды: митохондрии, рибосомы, аппарат Гольджи, в клетках низших животных и растений – центросома, животных – лизосомы, у растений – пластиды. Кроме того, цитоплазме обнаруживается целый ряд включений, принимающих участие в обмене веществ клетки: крахмал, капельки жира, кристаллы мочевины и т. д.
Установлено, что последняя состоит из параллельно расположенных сложных структур, имеющих вид пластинок и канальцев, на поверхности которых располагаются мельчайшие гранулы диаметром 100–120 Å. Эти образования названы эндоплазматическим комплексом. В состав этого комплекса включены различные дифференцированные органоиды: митохондрии, рибосомы, аппарат Гольджи, в клетках низших животных и растений – центросома, животных – лизосомы, у растений – пластиды. Кроме того, цитоплазме обнаруживается целый ряд включений, принимающих участие в обмене веществ клетки: крахмал, капельки жира, кристаллы мочевины и т. д.
Мембрана
Клетка окружена плазматической мембраной (от лат. «мембрана» – кожица, пленка). Ее функции очень разнообразны, но основная – защитная: она защищает внутреннее содержимое клетки от воздействий внешней среды. Благодаря различным выростам, складкам на поверхности мембраны клетки прочно соединяются между собой. Мембрана пронизана специальными белками, через которые могут перемещаться определенные вещества, необходимые клетке или подлежащие удалению из нее. Таким образом, через мембрану осуществляется обмен веществ. Причем, что очень важно, вещества пропускаются через мембрану избирательно, за счет чего в клетке поддерживается нужный набор веществ.
Таким образом, через мембрану осуществляется обмен веществ. Причем, что очень важно, вещества пропускаются через мембрану избирательно, за счет чего в клетке поддерживается нужный набор веществ.
У растений плазматическая мембрана снаружи покрыта плотной оболочкой, состоящей из целлюлозы (клетчатки). Оболочка выполняет защитную и опорную функции. Она служит внешним каркасом клетки, придавая ей определенную форму и размеры, препятствуя чрезмерному набуханию.
Ядро
Расположено в центре клетки и отделено двуслойной оболочкой. Имеет шаровидную или вытянутую форму. Оболочка – кариолемма – имеет поры, необходимые для обмена веществ между ядром и цитоплазмой. Содержимое ядра жидкое – кариоплазма, в которой содержатся плотные тельца – ядрышки. В них выделяется зернистость – рибосомы. Основная масса ядра – ядерные белки – нуклеопротеиды, в ядрышках – рибонуклеопротеиды, а в кариоплазме – дезоксирибонуклеопротеиды. Клетка покрыта клеточной оболочкой, которая состоит из белковых и липидных молекул, имеющих мозаичную структуру. Оболочка обеспечивает обмен веществ между клеткой и межклеточной жидкостью.
Оболочка обеспечивает обмен веществ между клеткой и межклеточной жидкостью.
ЭПС
Это система канальцев и полостей, на стенках которых располагаются рибосомы, обеспечивающие синтез белка. Рибосомы могут и свободно располагаться в цитоплазме. ЭПС бывают двух видов – шероховатая и гладкая: на шероховатой ЭПС (или гранулярной) располагается множество рибосом, которые осуществляют синтез белков. Рибосомы придают мембранам шероховатый вид. Мембраны гладкой ЭПС не несут рибосом на своей поверхности, в них располагаются ферменты синтеза и расщепления углеводов и липидов. Гладкая ЭПС выглядит как система тонких трубочек и цистерн.
Рибосомы
Мелкие тельца диаметром 15–20 мм. Осуществляют синтез белковых молекул, их сборку из аминокислот.
Митохондрии
Митахондрия
Это двумембранные органоиды, внутренняя мембрана которых имеет выросты – кристы. Содержимое полостей – матрикс. Митохондрии содержат большое количество липопротеидов и ферментов.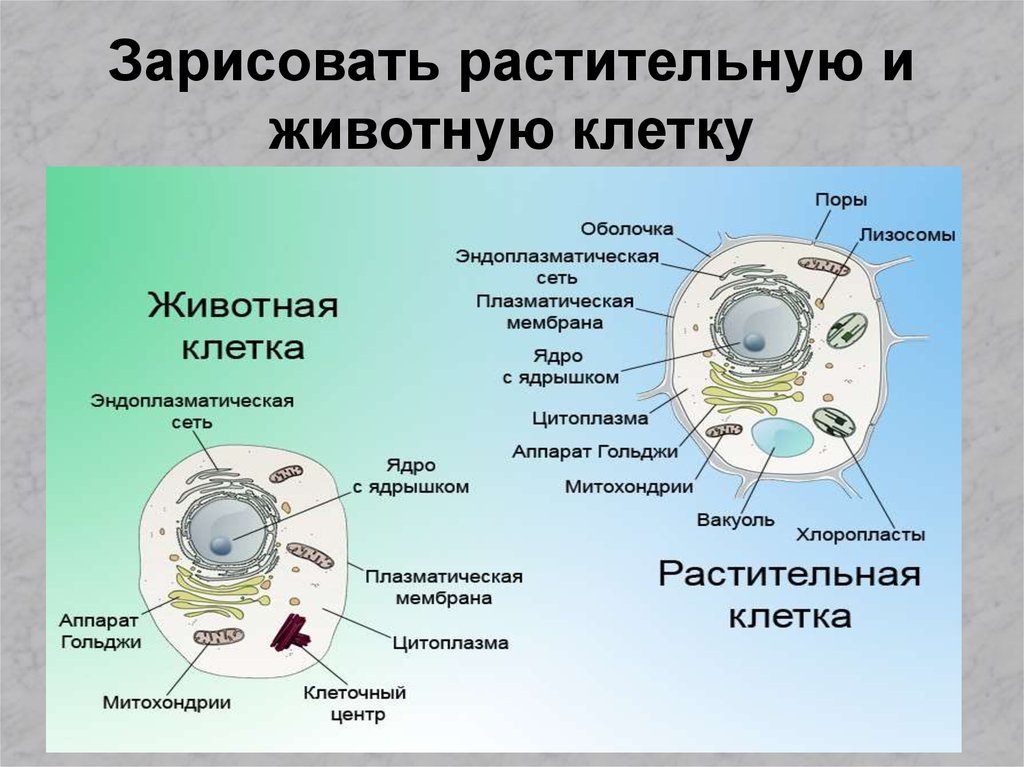 Это энергетические станции клетки.
Это энергетические станции клетки.
Пластиды (свойственны только клеткам растений!)
Их содержание в клетке – главная особенность растительного организма. Различают три основных типа пластид: лейкопласты, хромопласты и хлоропласты. Они имеют разную окраску. Бесцветные лейкопласты находятся в цитоплазме клеток неокрашенных частей растений: стеблях, корнях, клубнях. Например, их много в клубнях картофеля, в которых накапливаются зерна крахмала. Хромопласты находятся в цитоплазме цветков, плодов, стеблей, листьев. Хромопласты обеспечивают желтую, красную, оранжевую окраску растений. Зеленые хлоропласты содержатся в клетках листьев, стеблей и других частях растения, а также у разнообразных водорослей. Размеры хлоропластов 4-6 мкм, они часто имеют овальную форму. У высших растений в одной клетке содержится несколько десятков хлоропластов.
Зеленые хлоропласты способны переходить в хромопласты – поэтому осенью листья желтеют, а зеленые помидоры краснеют при созревании. Лейкопласты могут переходить в хлоропласты (позеленение клубней картофеля на свету).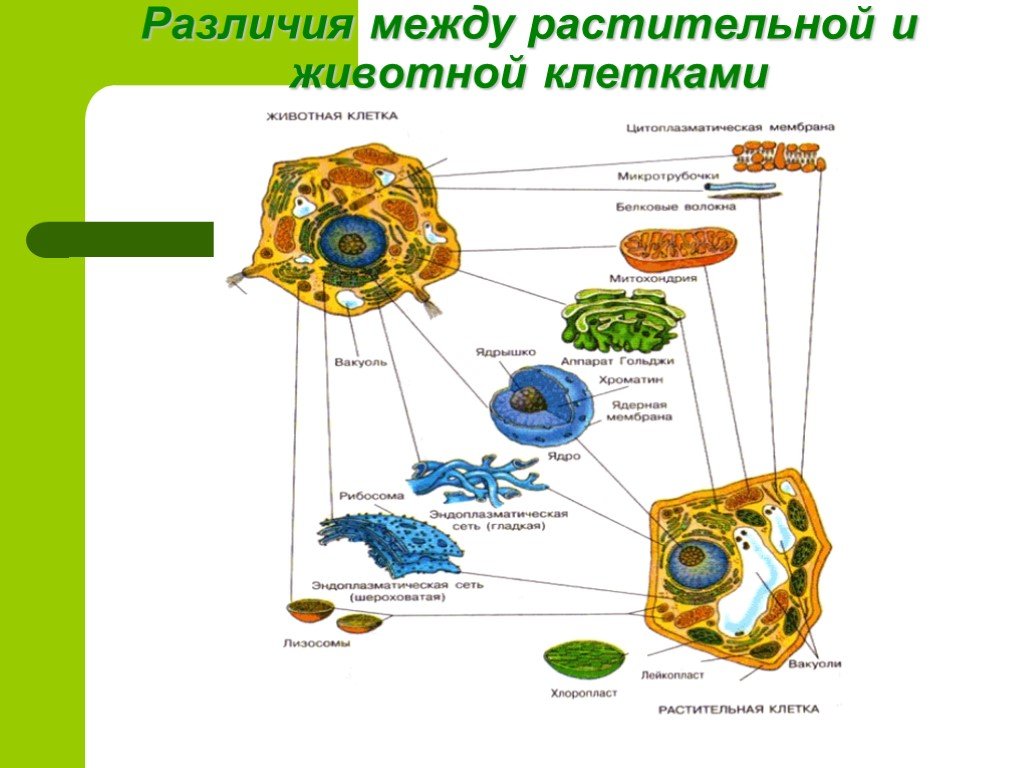 Таким образом, хлоропласты, хромопласты и лейкопласты способны к взаимному переходу.
Таким образом, хлоропласты, хромопласты и лейкопласты способны к взаимному переходу.
Основная функция хлоропластов – фотосинтез, т.е. в хлоропластах на свету осуществляется синтез органических веществ из неорганических за счет преобразования солнечной энергии в энергию молекул АТФ. Хлоропласты высших растений имеют размеры 5-10 мкм и по форме напоминают двояковыпуклую линзу. Каждый хлоропласт окружен двойной мембраной, обладающей избирательной проницаемостью. Снаружи располагается гладкая мембрана, а внутренняя имеет складчатую структуру. Основная структурная единица хлоропласта – тилакоид, плоский двумембранный мешочек, ирающий ведущую роль в процессе фотосинтеза. В мембране тилакоида расположены белки, аналогичные белкам митохондрий, которые участвуют в цепи переноса электоронов. Тилакоиды расположены стопками, напоминающие стопки монет (от 10 до 150) и называемыми гранами. Грана имеет сложное строение: в центре располагается хлорофилл, окруженный слоем белка; затем располагается слой липоидов, снова белок и хлорофилл.
Комплекс Гольджи
Это система полостей, отграниченных от цитоплазмы мембраной, может иметь разную форму. Накапливание в них белков, жиров и углеводов. Осуществление на мембранах синтеза жиров и углеводов. Образует лизосомы.
Основной структурный элемент аппарата Гольджи – мембрана, которая образует пакеты уплощенных цистерн, крупные и мелкие пузырьки. Цистерны аппарата Гольджи соединены с каналами эндоплазматической сети. Произведенные на мембранах эндоплазматической сети белки, полисахариды, жиры переносятся к аппарату Гольджи, накапливаются внутри его структур и «упаковываются» в виде вещества, готового либо к выделению, либо к использованию в самой клетке в процессе ее жизнедеятельности. В аппарате Гольджи образуются лизосомы. Кроме того, он участвует в наращивании цитоплазматической мембраны, например во время деления клетки.
Лизосомы
Тельца, отграниченные от цитоплазмы одной мембраной. Содержащиеся в них ферменты ускоряют реакцию расщепления сложных молекул до простых: белков до аминокислот, сложных углеводов до простых, липидов до глицерина и жирных кислот, а также разрушают отмершие части клетки, целые клетки. В лизосомах находится более 30 типов ферментов (вещества белковой природы, увеличивающие скорость химической реакции в десятки и сотни тысяч раз), способных расщеплять белки, нуклеиновые кислоты, полисахариды, жиры и другие вещества. Расщепление веществ с помощью ферментов называется лизисом, отсюда и происходит название органоида. Лизосомы образуются или из структур комплекса Гольджи, или из эндоплазматической сети. Одна из основных функций лизосом – участие во внутриклеточном переваривании пищевых веществ. Кроме того, лизосомы могут разрушать структуры самой клетки при ее отмирании, в ходе эмбрионального развития и в ряде других случаев.
В лизосомах находится более 30 типов ферментов (вещества белковой природы, увеличивающие скорость химической реакции в десятки и сотни тысяч раз), способных расщеплять белки, нуклеиновые кислоты, полисахариды, жиры и другие вещества. Расщепление веществ с помощью ферментов называется лизисом, отсюда и происходит название органоида. Лизосомы образуются или из структур комплекса Гольджи, или из эндоплазматической сети. Одна из основных функций лизосом – участие во внутриклеточном переваривании пищевых веществ. Кроме того, лизосомы могут разрушать структуры самой клетки при ее отмирании, в ходе эмбрионального развития и в ряде других случаев.
Вакуоли
Представляют собой полости в цитоплазме, заполненные клеточным соком, место накопления запасных питательных веществ, вредных веществ; они регулируют содержание воды в клетке.
Клеточный центр
Состоит из двух маленьких телец – центриолей и центросферы – уплотненного участка цитоплазмы. Играет важную роль при делении клеток
Органоиды движения клеток
- Жгутики и реснички, представляющие из себя выросты клетки и имеющие однотипное строение у животных и растений
- Миофибриллы – тонкие нити длиной более 1 см диаметром 1 мкм, расположенные пучками вдоль мышечного волокна
- Псевдоподии (выполняют функцию движения; за счет их происходит сокращение мышц)
Сходства растительных и животных клеток
К признакам, которыми похожи растительные и животные клетки, можно отнести следующие:
- Схожее строение системы структуры, т.
 е. наличие ядра и цитоплазмы.
е. наличие ядра и цитоплазмы. - Обменный процесс веществ и энергии близки по принципу осуществления.
- И в животной, и в растительной клетке имеется мембранное строение.
- Химический состав клеток очень похож.
- В клетках растения и животного присутствует похожий процесс клеточного деления.
- Растительная клетка и животная имеет единый принцип передачи кода наследственности.
 е. наличие ядра и цитоплазмы.
е. наличие ядра и цитоплазмы.Существенные различия между растительной и животной клеткой
Помимо общих признаков строения и жизнедеятельности растительной и животной клетки, существуют и особые отличительные черты каждой из них.
Отличия клеток заключаются в следующем:
- Наличие пластидов. В растительных клетках различают такие виды пластидов как хлоропласты, хромопласты и лейкопласты. А в животных клетках пластиды отсутствуют.
- Питание растительной клетки считается автотрофным, который, в свою очередь, разделяется на фототрофный и хемотрофный. А животная клетка питается гетеротрофным путём, который включает паразитический и сапротрофный виды.
- Процесс распада аденозинтрифосфорной кислоты в растительной клетке происходит в хлоропластах и прочих клеточных элементах, где необходима затрата энергии. В животной клетке такой процесс происходит во всех частях клетки, требующих энергетической затраты.
- Наличием клеточного центра у растений отличаются клетки низших растений. А среди животных клеток клеточный цент распространён у всех.
- Клетка растения содержит клеточную стенку из целлюлозы, а у животной клетки таковой не имеется.
- Второстепенные и необязательные компоненты растительной клетки состоят из запаса питательных веществ в качестве крахмальных зёрен, а также зёрен белка и капель масла. Также сюда входят вакуоли, содержащие клеточный сок и солевые кристаллы. А животная клетка содержит в качестве необязательных компонентов питательные вещества из зёрен и капель белков, жиров и углеводов. Также есть содержание солевых кристаллов, пигментов и конечных обменных продуктов.
- Растительные вакуоли представляют собой полости с соком. А у животной клетки имеются мелкие вакуоли, разделяющиеся на сократительные, пищеварительные и выделительные.
 А у животной клетки имеются мелкие вакуоли, разделяющиеся на сократительные, пищеварительные и выделительные.
А у животной клетки имеются мелкие вакуоли, разделяющиеся на сократительные, пищеварительные и выделительные.Таким образом, можно сказать, что растительные и животные клетки похожи между собой содержанием некоторых важных элементов и некоторыми процессами жизнедеятельности, а также имеют существенные отличия в структуре и обменных процессах.
Видео
Источники
- http://www.activestudy.info/stroenie-kletki/
http://scienceland.info/biology7/animal-cell
http://vsedz.ru/content/3-строение-растительной-и-животной-клеток
http://shkolo.ru/stroenie-kletki/
https://dist-tutor.info/mod/resource/view.php?id=12830
http://poznayka.org/s36039t1.html
http://vchemraznica.ru/chem-rastitelnaya-kletka-otlichaetsya-ot-zhivotnoj/
Различия и сходства растительной и животной клетки
Клетка — это простейший элемент строения какого-либо организма, свойственный как животному, так и растительному миру. Из чего она состоит? Сходства и различия клеток растительного и животного происхождения мы рассмотрим далее.
Из чего она состоит? Сходства и различия клеток растительного и животного происхождения мы рассмотрим далее.
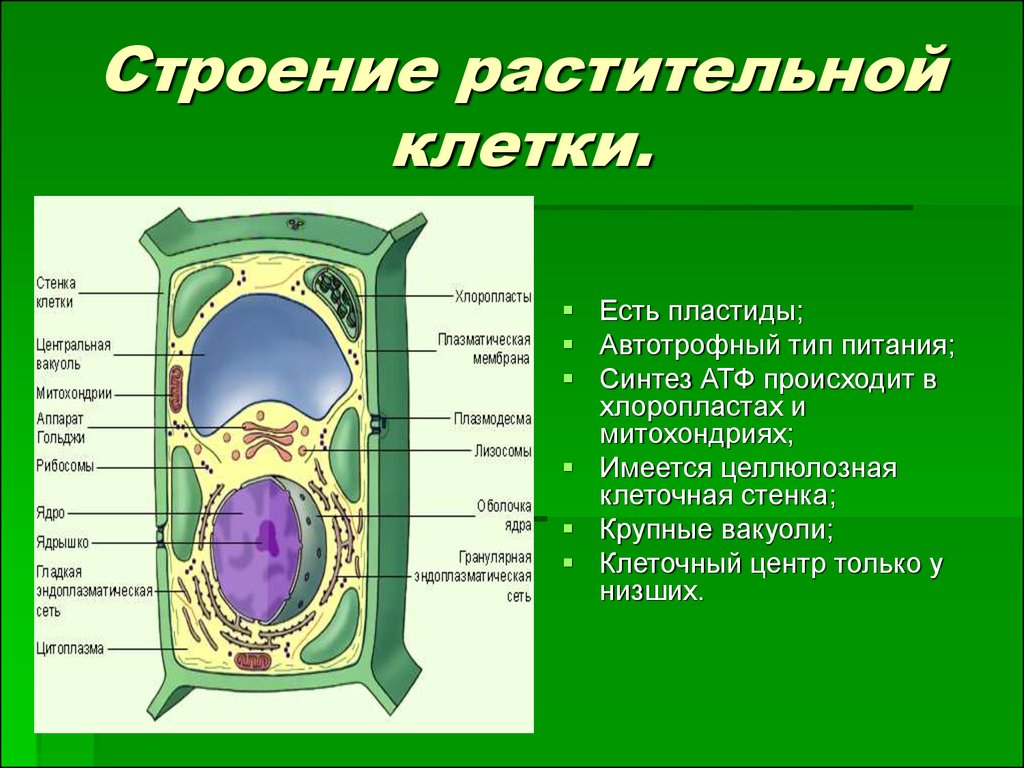
Растительная клетка
Все, чего мы раньше не видели и не знали, всегда вызывает очень сильный интерес. Как часто вы рассматривали клетки под микроскопом? Наверное, не каждый его и в глаза видел. На фото представлена растительная клетка. Основные ее части очень хорошо видны. Итак, растительная клетка состоит из оболочки, пор, мембран, цитоплазмы, вакуоли, ядерной оболочки, ядра, ядрышка и пластид.
Как видно, строение не такое и хитрое. Сразу обратим внимание на сходства растительной и животной клетки относительно строения. Здесь отметим наличие вакуоли. В растительных клетках она одна, а в животной имеется множество мелких, выполняющих функцию внутриклеточного пищеварения. Также заметим, что есть принципиальное сходство в строении: оболочка, цитоплазма, ядро. Строением мембран они тоже не отличаются.
Животная клетка
В прошлом пункте мы отметили сходства растительной и животной клетки относительно строения, но они не абсолютно идентичны, имеют различия. Например, животная клетка не имеет клеточной стенки. Также отметим наличие органоидов: митохондрии, эндоплазматическая сеть, аппарат Гольджи, лизосомы, рибосомы, клеточный центр. Обязательным элементом является ядро, которое контролирует все функции клеток, включая размножение. Это мы также отмечали, рассматривая сходства растительной и животной клетки.
Например, животная клетка не имеет клеточной стенки. Также отметим наличие органоидов: митохондрии, эндоплазматическая сеть, аппарат Гольджи, лизосомы, рибосомы, клеточный центр. Обязательным элементом является ядро, которое контролирует все функции клеток, включая размножение. Это мы также отмечали, рассматривая сходства растительной и животной клетки.
Сходства клеток
Несмотря на то что во многом клетки отличаются друг от друга, упомянем основные сходства. Сейчас нельзя точно сказать, когда и как появилась жизнь на земле. Но сейчас мирно сосуществуют многие царства живых организмов. Несмотря на то что все ведут разный образ жизни, имеют разное строение, несомненно, существует множество сходств. Это говорит о том, что все живое на земле имеет одного общего предка. Вот основные признаки сходства:
- строение клеток;
- сходство процессов обменных веществ;
- кодирование информации;
- одинаковый химический состав;
- идентичный процесс деления.
Как видно из приведенного перечня, сходства растительной и животной клетки многочисленны, несмотря на такое разнообразие форм жизни.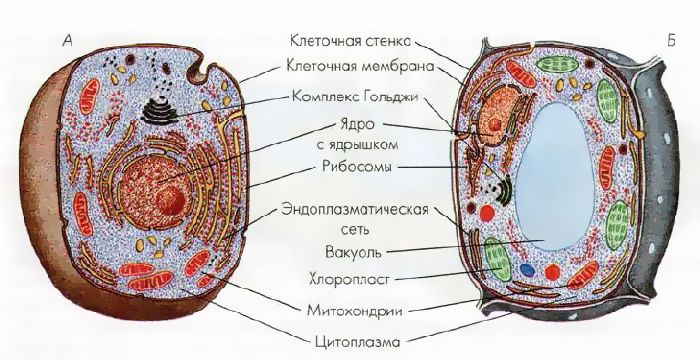
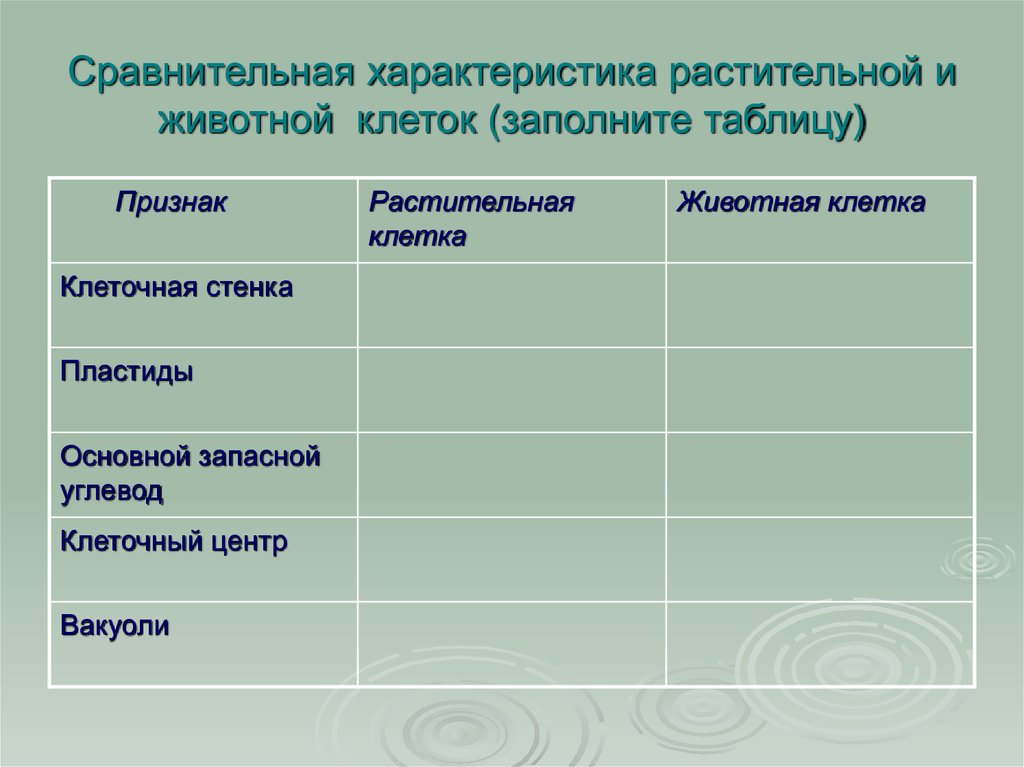
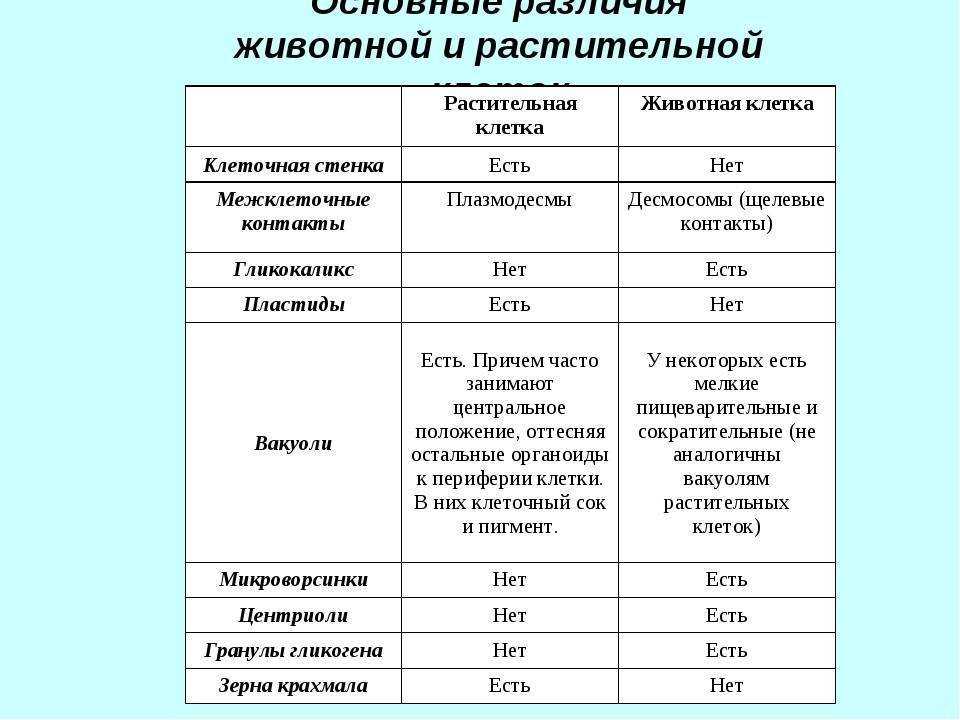
Различия клеток. Таблица
Несмотря на большое количество сходных черт, клетки животного и растительного происхождения имеют много различий. Для наглядности приводим таблицу:
| Признаки | Растительная клетка | Животная клетка |
| Целлюлозная клеточная стенка | + | — |
| Пластиды | + | — |
| Основной запас углеводов | крахмал | гликоген |
| Клеточный центр | — | + |
| Вакуоль | Одна | Многочисленные |
| Синтез АТФ | Хлоропластах, митохондриях | Митохондриях |
| Способ питания | Автотрофный | Гетеротрофный |
Основное отличие заключается в способе питания. Как видно из таблицы, растительная клетка имеет автотрофный способ питания, а животная — гетеротрофный. Это связано с тем, что растительная клетка содержит хлоропласты, то есть растения сами синтезируют все необходимые для выживания вещества, используя энергию света и фотосинтез. Под гетеротрофным способом питания понимается попадание в организм необходимых веществ с пищей. Эти же вещества являются и источником энергии для существа.
Под гетеротрофным способом питания понимается попадание в организм необходимых веществ с пищей. Эти же вещества являются и источником энергии для существа.
Отметим, что есть и исключения, например, зеленые жгутиконосцы, которые способны получать необходимые вещества двумя способами. Так как для процесса фотосинтеза необходима солнечная энергия, то автотрофный способ питания они применяют в светлое время суток. Ночью же они вынуждены употреблять уже готовые органические вещества, то есть питаются гетеротрофным способом.
Сходство и различие между животной и растительной клеткой – LORECENTRAL
по БИОЛОГИИ, Научная разница
По мере того, как в клеточной биологии разрабатывается больше методов, мы узнаем больше о великих тайнах и секретах, которые скрывают клетки. Знание растительных и животных клеток очень важно для понимания функционирования живых существ, понимания болезней, от которых они страдают, разработки терапевтических методов против этих болезней или получения продуктов, представляющих коммерческий интерес.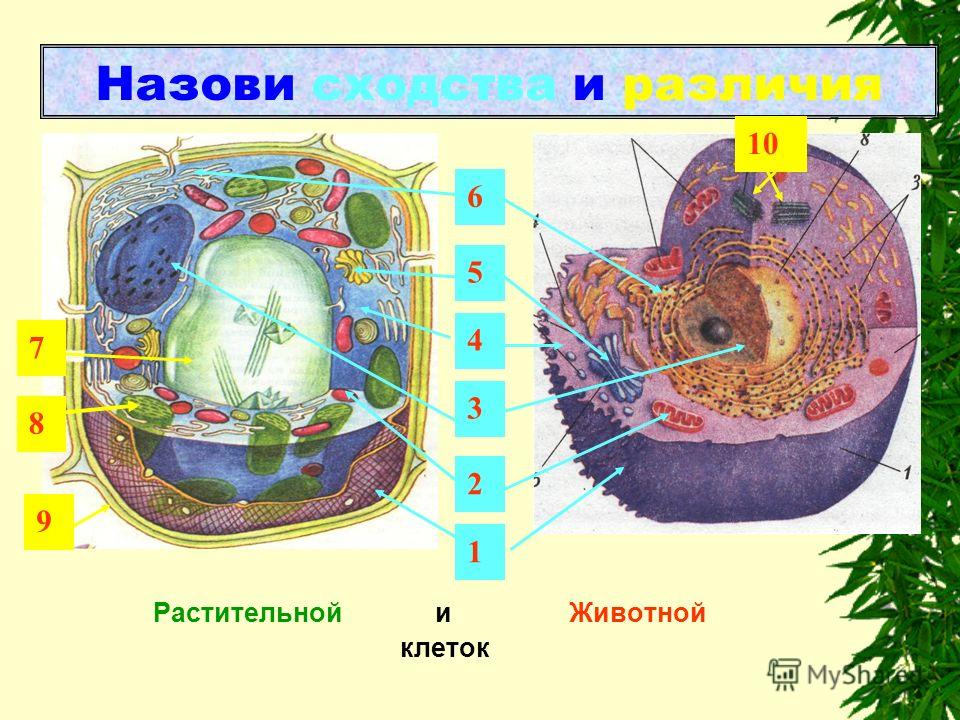
Также важно понимать сходство и различие между животными и растительными клетками . В этой статье lorecentral.org мы расскажем вам.
Содержание
Что такое клетка
Клетка является морфологической и функциональной единицей всех живых существ , способной воспроизводить, расти, метаболизировать, взаимодействовать с другими клетками и захватывать сигналы от внеклеточную среду или запрограммировать собственную смерть. Живое существо может иметь одну клетку и быть одноклеточным, как в случае бактерий, или иметь несколько клеток и быть многоклеточным, как в случае животные и растения .
В многоклеточных организмах клетки не являются отдельными компартментами, но способны общаться друг с другом посредством сигналов, чтобы координировать и реагировать на сигналы, поступающие из внешней среды, так что организм связан с внешней средой. Клетки также способны регулировать свой рост до поступления сигналов и программировать собственную гибель клеток за счет накопления мутаций или сбоев в их функционировании.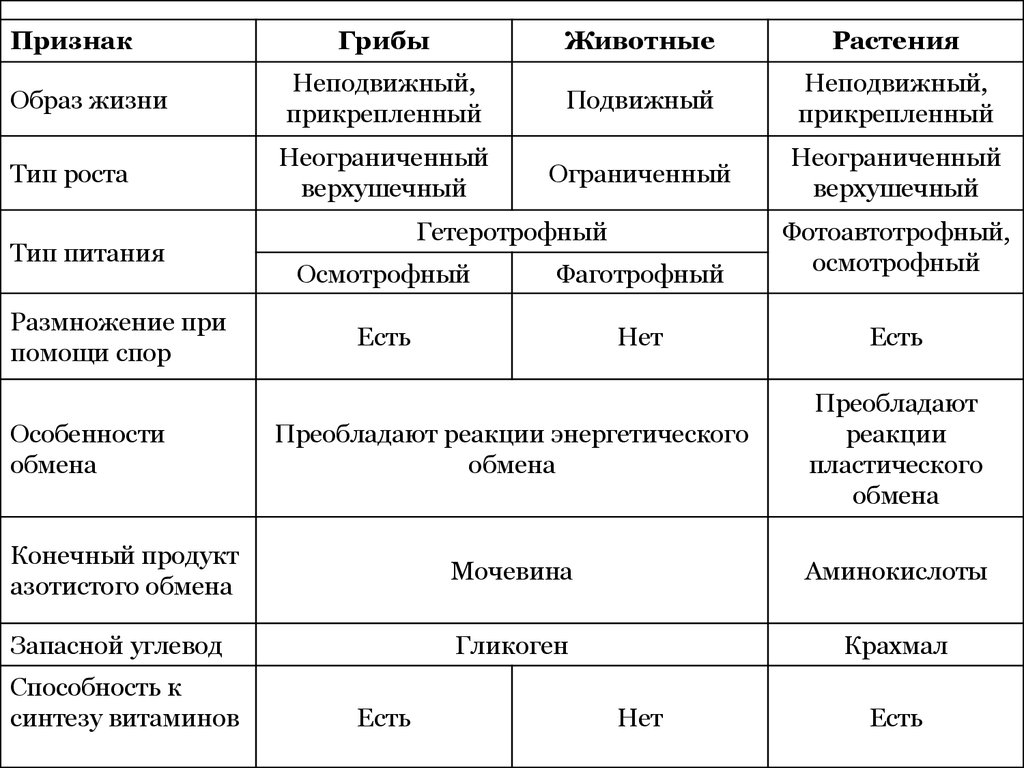 Примером важности этих реакций является рак, когда клетка начинает выходить из-под контроля из-за своей неспособности координировать свои действия с окружающей средой и планировать свою смерть.
Примером важности этих реакций является рак, когда клетка начинает выходить из-под контроля из-за своей неспособности координировать свои действия с окружающей средой и планировать свою смерть.
Но никакое изучение клетки было бы невозможно без технического прогресса и инноваций в области оптики и электроники, которые позволили разработать микроскопы для наблюдения структур размером всего в нанометр.
Сходства между клетками животных и растений
Таким образом, зная, что такое клетка, мы объясняем, что сходств между клетками растений и животных являются:
- Они являются основными морфологическими и функциональными единицами.
- И растительные, и животные клетки являются эукариотическими клетками. Эукариотические клетки, в отличие от прокариот (бактерии и археи), имеют, среди прочего, организованное клеточное ядро с защитным покрытием, клеточные органеллы, цитоскелет (клеточный скелет) и организованный и упакованный в хромосомы геном.
- Они окружены полупроницаемой плазматической мембраной, ограничивающей цитоплазму.
- Размер от 10 до 100 мкм. Клетки животных могут достигать 30 мкм, в то время как у овощей 100 мкм (микрон составляет одну тысячную миллиметра).
- Учитывая их небольшой размер, их нельзя наблюдать невооруженным глазом и требуется помощь микроскопов.
Сходство и различие между животной и растительной клеткой
Различия между животной и растительной клеткой
Теперь, когда мы упомянули о сходствах, мы уточним, что различий, которые мы можем найти между растительной и животной клеткой , таковы:
- Растительные клетки имеют клеточной стенки вне плазматической мембраны. Эта стенка придает им большую жесткость и состоит из целлюлозы, лигнина и других компонентов. Клетки животных не имеют этой клеточной стенки. Некоторые компоненты клеточной стенки растений имеют множество коммерческих и промышленных применений.
- В растительной клетке есть хлоропласты. Эти хлоропласты содержат пигменты, такие как хлорофилл или каротин, и обеспечивают процесс фотосинтеза. Многие из этих пигментов имеют коммерческое или промышленное применение. Клетки животных не имеют хлоропластов.
- Клетки растений обладают способностью производить себе пищу из неорганических компонентов посредством явления фотосинтеза, которое называется автотрофным питанием.
- Клетки животных не обладают способностью производить собственное питание из неорганических компонентов, что называется гетеротрофным питанием.
- В процессе фотосинтеза растительная клетка способна преобразовывать солнечную или световую энергию в химическую энергию.
- В клетках животных энергия обеспечивается митохондриями.
- Клетки растений имеют цитоплазму, занятую крупными вакуолями на 90% своего пространства, иногда даже в виде одной крупной вакуоли. Эти вакуоли служат для хранения продуктов метаболизма и удаления продуктов жизнедеятельности. В то время как животные клетки имеют вакуоли, но маленькие и не занимают так много места.
- В клетках животных есть органелла, называемая центросомой (отвечающая за разделение хромосом для деления между дочерними клетками), а в клетках растений — нет.
- Клетки растений обычно имеют призматическую форму, тогда как клетки животных могут иметь различную форму.
 Эти хлоропласты содержат пигменты, такие как хлорофилл или каротин, и обеспечивают процесс фотосинтеза. Многие из этих пигментов имеют коммерческое или промышленное применение. Клетки животных не имеют хлоропластов.
Эти хлоропласты содержат пигменты, такие как хлорофилл или каротин, и обеспечивают процесс фотосинтеза. Многие из этих пигментов имеют коммерческое или промышленное применение. Клетки животных не имеют хлоропластов.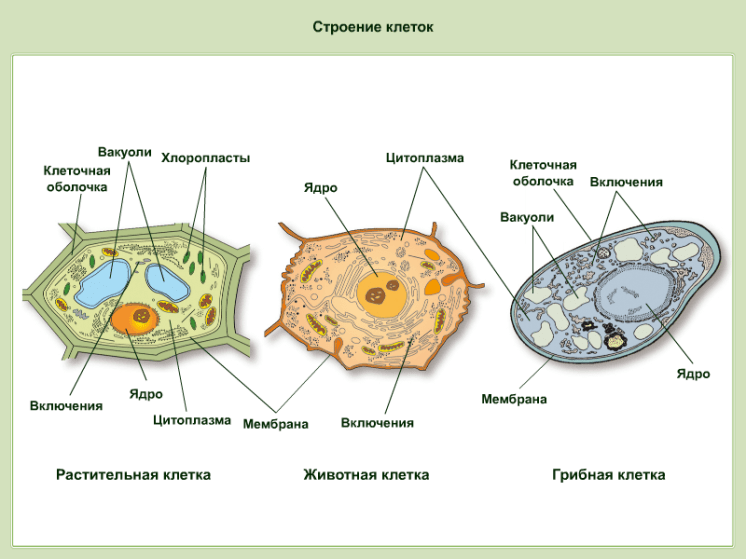
7 частей легких (и их функции)
по
ГЭВИН ТОМАС
Какова функция глиальных клеток?
по
ГЭВИН ТОМАС
В чем разница между сгибанием и разгибанием?
по
ГЭВИН ТОМАС
Примеры ферментов в организме и их субстратов
по
ГЭВИН ТОМАС
Примеры ферментов (и их функции)
по
ГЭВИН ТОМАС
Примеры рестриктаз
по
ГЭВИН ТОМАС
Чем отличаются африканские слоны от азиатских
Сходства и различия между растениями и животными
Происходит ли сходное метаболическое перепрограммирование в Fe-дефицитных растительных клетках и опухолевых клетках животных?
Клетки растений и животных сильно отличаются друг от друга по многим функциям. Однако сравнение их поведения в различных стрессовых условиях обнаруживает интригующее сходство. Интересно, что одним из таких случаев является сравнение метаболического перепрограммирования в клетках растений при дефиците Fe и развитии рака в клетках животных.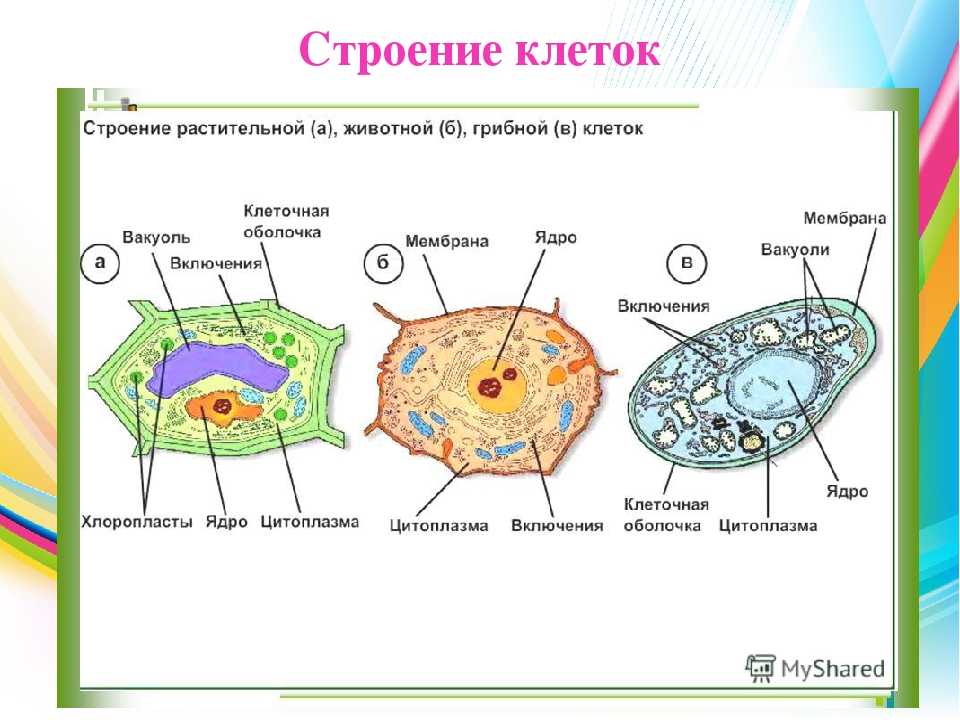
В недавних обзорах изучалась идея о том, что дефицит Fe вызывает перепрограммирование метаболизма у двудольных растений (Vigani, 2012) для поглощения Fe из почвы. Вкратце, в условиях ограниченного роста Fe клетки корней перепрограммируют свой метаболизм следующим образом:
1 Увеличение гликолиза и нарушение дыхательной цепи. Fe является важным кофактором для нескольких митохондриальных ферментов, и при дефиците Fe функции митохондрий сильно нарушаются, не теряя значения в регуляции метаболических ответов на этот стресс, и, что удивительно, в Fe-дефицитных корнях количество митохондрий увеличивается, даже если они менее эффективны. В этом контексте Fe-дефицитные клетки активируют гликолиз, чтобы обеспечить энергией механизм приобретения Fe (Vigani, 2012, и ссылки в нем).
2 Увеличение анаэробных реакций. Лопес-Миллан и др. (2000, 2009) сообщили, что дефицит железа повышает активность ферментов брожения, таких как лактатдегидрогеназа (ЛДГ) и пируватдекарбоксилаза (ПДК), которые могут способствовать окислению пула пиридиновых нуклеотидов в корнях томата и сахарной свеклы. Эти данные также были подтверждены микрочиповым анализом Fe-дефицитного Arabidopsis (Thimm et al., 2001) и протеомными данными, полученными из корней сахарной свеклы и огурца, в которых предполагаемые белки алкогольдегидрогеназы и малатдегидрогеназы накапливались при дефиците Fe ( Donnini et al., 2010; Rellán-álvarez et al., 2010).
Эти данные также были подтверждены микрочиповым анализом Fe-дефицитного Arabidopsis (Thimm et al., 2001) и протеомными данными, полученными из корней сахарной свеклы и огурца, в которых предполагаемые белки алкогольдегидрогеназы и малатдегидрогеназы накапливались при дефиците Fe ( Donnini et al., 2010; Rellán-álvarez et al., 2010).
3 Увеличение окислительного пентозофосфатного пути (OPP). В частности, некоторые цитозольные NAD(P) + -зависимые дегидрогеназы, такие как глюкозо-6-фосфатдегидрогеназа (окислительный пентозофосфатный путь; Vigani, 2012), проявляли повышенную активность для производства восстанавливающих эквивалентов, необходимых для поддержания поглощения Fe. .
Удивительно, но аналогичное метаболическое перепрограммирование происходит в опухолевых клетках животных: весь их метаболизм реорганизуется для поддержания роста и пролиферации клеток. Вкратце, некоторые раковые клетки характеризуются следующими метаболическими адаптациями:
1 Альтерация, связанная с опухолью, представляет собой «феномен Варбурга», при котором глюкоза преимущественно превращается в лактат путем усиления гликолиза и ферментативных реакций, а не полностью окисляется путем окислительного фосфорилирования (OXOPHOS; Warburg et al. , 1924; Warburg, 1956; Брахими-Хорн и др., 2007). Поэтому это метаболическое изменение часто называют «аэробным гликолизом» (Vander Heiden et al., 2009; рисунок 1), поскольку это ферментативный путь, фактически происходящий в аэробных условиях. Кроме того, как наблюдается в клетках растений с дефицитом Fe, некоторые опухолевые клетки содержат аномально большое количество митохондрий (Carew and Huang, 2002).
, 1924; Warburg, 1956; Брахими-Хорн и др., 2007). Поэтому это метаболическое изменение часто называют «аэробным гликолизом» (Vander Heiden et al., 2009; рисунок 1), поскольку это ферментативный путь, фактически происходящий в аэробных условиях. Кроме того, как наблюдается в клетках растений с дефицитом Fe, некоторые опухолевые клетки содержат аномально большое количество митохондрий (Carew and Huang, 2002).
2 Раковые клетки продуцируют угольную и молочную кислоты, при этом лактат является основным конечным продуктом аэробного гликолиза. Такие кислоты способствуют инвазии опухоли (Swietach et al., 2007) и подавляют противораковые иммунные эффекторы (Fischer et al., 2007). Лактат, продуцируемый опухолевыми клетками, может поглощаться стромальными клетками для регенерации пирувата, который затем может быть экструдирован для дозаправки раковых клеток или использован для дыхательной цепи (Koukourakis et al., 2006).
3 Опухоли могут метаболизировать глюкозу через OPP с образованием NADPH, который обеспечивает функцию антиоксидантной защиты клетки от враждебного микроокружения, что приводит к толерантности к химиотерапевтическим агентам (Gatenby and Gillies, 2004).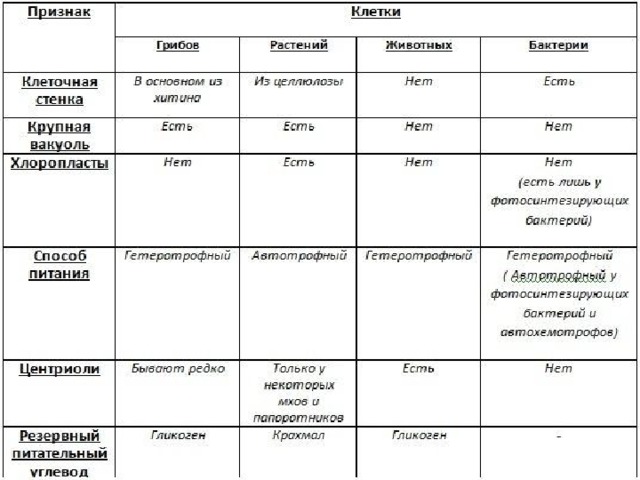
Один из механизмов активации аэробного гликолиза включает фактор, индуцируемый гипоксией (HIF), фактор транскрипции, который активируется не только гипоксическим стрессом, но также онкогенным, воспалительным, метаболическим и окислительным стрессом (Harris, 2002; Semenza, 2007). ; Тейлор и Пуиссегюр, 2007). HIF-1 состоит из конститутивно экспрессируемой субъединицы HIF-1β и активатора транскрипции, регулируемого кислородом, HIF-1α. Стабильность и активность α-субъединицы HIF регулируются посттрансляционными модификациями (например, гидроксилированием). При нормоксии гидроксилирование двух остатков пролина и ацетилирование остатка лизина в домене кислородзависимой деградации (ODDD) HIF-1α запускает его ассоциацию с лигазным комплексом pVHL E3, что приводит к деградации HIF-1α через убиквитин-протеасомные пути. Механизм гидроксилирования опосредован пролилгидроксилазой (PHD), которая принадлежит к семейству 2-оксоглутарат-зависимых диоксигеназ и катализирует образование 4-гидроксипролина, для чего требуются 2-оксоглутарат и O 2 в качестве косубстратов, Fe 2+ в качестве кофактора и аскорбат для оптимальной активности. Накопление HIF-1 было обнаружено при различных видах рака. Когда по какой-либо причине механизм деградации, зависящий от гидроксилирования, не активируется, HIF-1 накапливается и стимулирует превращение глюкозы в пируват и лактат, активируя переносчик глюкозы изоформы 1 (GLUT1), гексокиназу (НК1 и НК2) и лактатдегидрогеназу А. LDHA), а также лактат-экструдирующий фермент монокарбоксилатный переносчик 4 (MCT4; Semenza, 2007). По-видимому, эффект Варбурга, опосредованный индукцией HIF-1, по-видимому, строго связан с гипоксией. Тем не менее, в опухолевых клетках было постулировано, что гипоксия представляет собой важный аспект биологии рака, но только как поздно возникающее событие. Таким образом, гипоксию нельзя рассматривать как основную причину переключения раковых клеток на аэробный гликолиз (Vander Heiden et al., 2009).). В опухолевых клетках происходит метаболическое переключение с более энергичного (окислительное фосфорилирование) на менее (аэробный гликолиз) эффективного метаболизма, потому что, будучи пролиферирующими клетками, они имеют важные метаболические потребности, выходящие за рамки простого высокого производства АТФ.
Накопление HIF-1 было обнаружено при различных видах рака. Когда по какой-либо причине механизм деградации, зависящий от гидроксилирования, не активируется, HIF-1 накапливается и стимулирует превращение глюкозы в пируват и лактат, активируя переносчик глюкозы изоформы 1 (GLUT1), гексокиназу (НК1 и НК2) и лактатдегидрогеназу А. LDHA), а также лактат-экструдирующий фермент монокарбоксилатный переносчик 4 (MCT4; Semenza, 2007). По-видимому, эффект Варбурга, опосредованный индукцией HIF-1, по-видимому, строго связан с гипоксией. Тем не менее, в опухолевых клетках было постулировано, что гипоксия представляет собой важный аспект биологии рака, но только как поздно возникающее событие. Таким образом, гипоксию нельзя рассматривать как основную причину переключения раковых клеток на аэробный гликолиз (Vander Heiden et al., 2009).). В опухолевых клетках происходит метаболическое переключение с более энергичного (окислительное фосфорилирование) на менее (аэробный гликолиз) эффективного метаболизма, потому что, будучи пролиферирующими клетками, они имеют важные метаболические потребности, выходящие за рамки простого высокого производства АТФ. Действительно, во время пролиферации клетки должны воспроизвести все свое клеточное содержимое: глюкоза должна быть направлена на синтез нескольких клеточных молекул, таких как ацетил-КоА, гликолитических промежуточных продуктов и рибозы для биосинтеза жирных кислот, заменимых аминокислот и аминокислот. нуклеотидов соответственно (Vander Heiden et al., 2009; Фигура 1). OXOPHOS не нарушается, но замедляется, чтобы в достаточной степени удовлетворить энергетический запрос на жизнь клетки и сбалансировать соотношение АТФ/АДФ (Vander Heiden et al., 2009).
Действительно, во время пролиферации клетки должны воспроизвести все свое клеточное содержимое: глюкоза должна быть направлена на синтез нескольких клеточных молекул, таких как ацетил-КоА, гликолитических промежуточных продуктов и рибозы для биосинтеза жирных кислот, заменимых аминокислот и аминокислот. нуклеотидов соответственно (Vander Heiden et al., 2009; Фигура 1). OXOPHOS не нарушается, но замедляется, чтобы в достаточной степени удовлетворить энергетический запрос на жизнь клетки и сбалансировать соотношение АТФ/АДФ (Vander Heiden et al., 2009).
Рисунок 1. Схематическое представление различий между окислительным фосфорилированием, анаэробным гликолизом и аэробным гликолизом (эффект Варбурга) у животных и метаболическими реакциями, вызванными дефицитом железа в растительных клетках . В присутствии кислорода нормальные клетки (как животных, так и растений) сначала метаболизируют глюкозу в пируват посредством гликолиза, а затем полностью окисляют большую часть этого пирувата в митохондриях в ходе окислительного фосфорилирования (ОКСОФОС).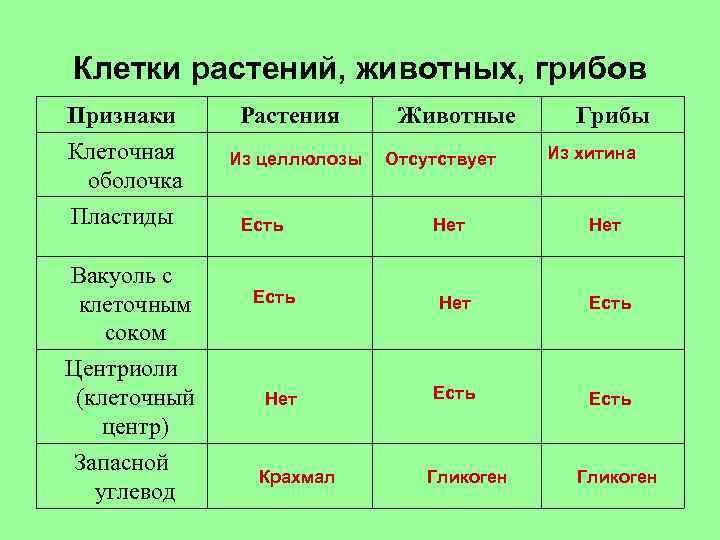 Когда кислород ограничен, клетки могут перенаправлять пируват, образующийся в результате гликолиза, от митохондриального OXOPHOS, индуцируя ферментативный путь (анаэробный гликолиз). Эффект Варбурга, наблюдаемый в раковых клетках, имеет тенденцию превращать большую часть глюкозы в лактат независимо от того, присутствует кислород или нет (аэробный гликолиз). Сходный эффект Варбурга можно наблюдать и в Fe-дефицитной растительной клетке, в которой замедление OXOPHOS из-за недостатка Fe определяет активацию гликолизно-ферментативных реакций. Поскольку это метаболическое переключение произошло в присутствии кислорода, мы можем рассматривать его как индукцию аэробного гликолиза. (Рисунок представляет собой адаптацию/реинтерпретацию рисунка 2, приведенного в Vander Heiden et al., 2009.).
Когда кислород ограничен, клетки могут перенаправлять пируват, образующийся в результате гликолиза, от митохондриального OXOPHOS, индуцируя ферментативный путь (анаэробный гликолиз). Эффект Варбурга, наблюдаемый в раковых клетках, имеет тенденцию превращать большую часть глюкозы в лактат независимо от того, присутствует кислород или нет (аэробный гликолиз). Сходный эффект Варбурга можно наблюдать и в Fe-дефицитной растительной клетке, в которой замедление OXOPHOS из-за недостатка Fe определяет активацию гликолизно-ферментативных реакций. Поскольку это метаболическое переключение произошло в присутствии кислорода, мы можем рассматривать его как индукцию аэробного гликолиза. (Рисунок представляет собой адаптацию/реинтерпретацию рисунка 2, приведенного в Vander Heiden et al., 2009.).
Перемещая акцент на растения, кажется, что метаболические изменения, вызванные дефицитом железа, имитируют метаболические изменения, наблюдаемые в раковых клетках, что выдвигает интригующую гипотезу.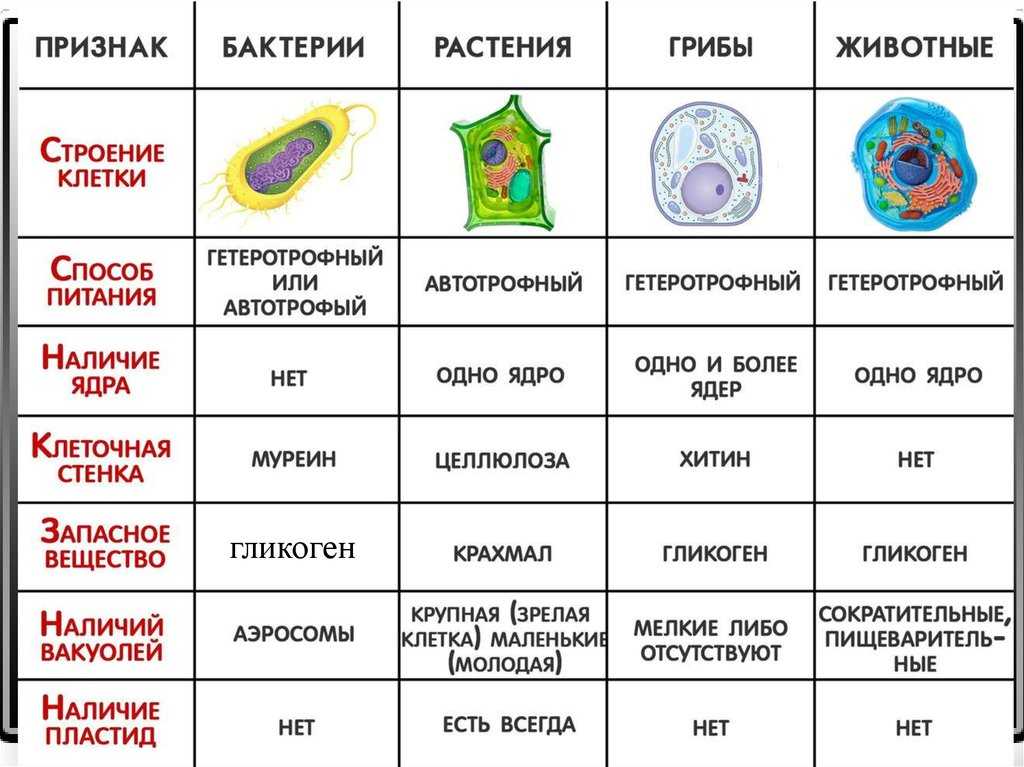 Низкое содержание Fe вызывает замедление OXOPHOS, а также индукцию гликолиза и ферментативных реакций даже в аэробных условиях. Поскольку, как и в опухолевых клетках, так и в Fe-дефицитных растительных клетках, метаболизм глюкозы в основном (не полностью, митохондрии все еще работают в обоих случаях) направлен на гликолиз плюс ферментативные реакции, можем ли мы постулировать варбургоподобный эффект и в Fe -дефицитные растения? В опухолевых клетках индукция гликолиза в сторону ферментативной реакции называется аэробным гликолизом, а не анаэробным гликолизом, поскольку он происходит независимо от присутствия кислорода. Поскольку в растительных клетках с дефицитом Fe наблюдаемый метаболический переключатель, аналогичный таковому в опухолевых клетках животных, первоначально является следствием недостатка Fe, а не недостатка кислорода, можно ли его рассматривать как аэробный гликолиз? (Фигура 1). Кроме того, в растительных клетках с дефицитом Fe глюкоза, направленная на процесс аэробного гликолиза, позволяет ей удовлетворить как (i) потребность в энергии для выживания клеток, так и (ii) потребность в гликолитических промежуточных продуктах для синтеза нескольких полезных метаболических соединений.
Низкое содержание Fe вызывает замедление OXOPHOS, а также индукцию гликолиза и ферментативных реакций даже в аэробных условиях. Поскольку, как и в опухолевых клетках, так и в Fe-дефицитных растительных клетках, метаболизм глюкозы в основном (не полностью, митохондрии все еще работают в обоих случаях) направлен на гликолиз плюс ферментативные реакции, можем ли мы постулировать варбургоподобный эффект и в Fe -дефицитные растения? В опухолевых клетках индукция гликолиза в сторону ферментативной реакции называется аэробным гликолизом, а не анаэробным гликолизом, поскольку он происходит независимо от присутствия кислорода. Поскольку в растительных клетках с дефицитом Fe наблюдаемый метаболический переключатель, аналогичный таковому в опухолевых клетках животных, первоначально является следствием недостатка Fe, а не недостатка кислорода, можно ли его рассматривать как аэробный гликолиз? (Фигура 1). Кроме того, в растительных клетках с дефицитом Fe глюкоза, направленная на процесс аэробного гликолиза, позволяет ей удовлетворить как (i) потребность в энергии для выживания клеток, так и (ii) потребность в гликолитических промежуточных продуктах для синтеза нескольких полезных метаболических соединений.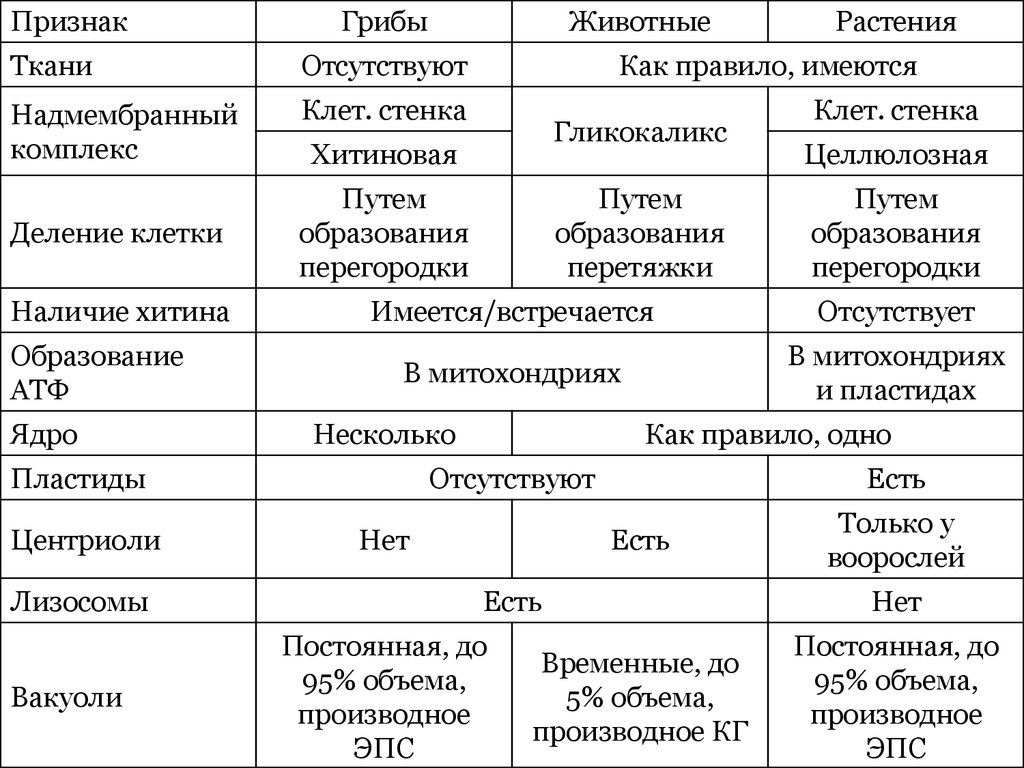 в клетку для преодоления дефицита Fe, например органические кислоты.
в клетку для преодоления дефицита Fe, например органические кислоты.
Несмотря на то, что метаболическое перепрограммирование Fe-дефицитных клеток растений имеет сходство с таковым, происходящим в опухолевых клетках животных, важно принимать во внимание некоторые метаболические аспекты, которыми эти две системы могут отличаться друг от друга. Хотя эффект Варбурга считается отличительной чертой исследований рака, он возникает в некоторых специфических опухолевых клетках. Действительно, некоторые опухоли содержат две субпопуляции раковых клеток, которые различаются по своим путям выработки энергии, но, по-видимому, они функционируют симбиотически. Одна субпопуляция состоит из глюкозозависимых клеток (клетки, отображающие эффект Варбурга), которые секретируют лактат, тогда как клетки, принадлежащие ко второй субпопуляции, преимущественно импортируют и используют лактат, продуцируемый их соседями (Hanahan and Weinberg, 2011 и ссылки в этой работе). У Fe-дефицитных растений наличие столь же специализированных клеточных субпопуляций пока не наблюдается.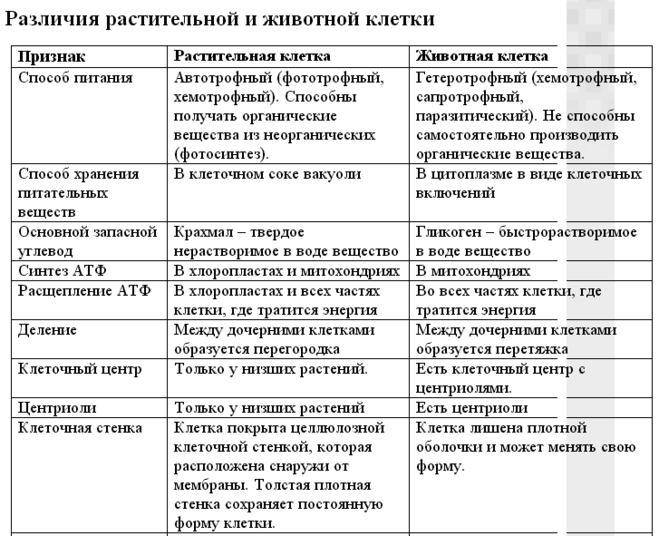 Однако в корнях растений действительно присутствуют разные ткани и, следовательно, разные «клеточные популяции». Было замечено, что при дефиците железа внешние клеточные слои корня преимущественно вовлечены в активацию типичных ферментов (т. е. железо-хелатредуктазы, H + -АТФаза), участвующие в механизме поглощения Fe из почвы (Santi, Schmidt, 2008). Кроме того, накопление белка H + -ATPase было особенно высоким в ризодермальных и энтодермальных клетках по сравнению с другими слоями Fe-дефицитных корней огурца (Dell’Orto et al., 2002). Таким образом, можно ли рассматривать ризодермальные клетки у растений с дефицитом Fe как глюкозозависимые клетки, тогда как корковые слои можно рассматривать как вторую субпопуляцию клеток, наблюдаемую в опухолях? Этот вопрос до сих пор остается открытым, так как нет данных о различном метаболическом перепрограммировании между разными слоями корня: действительно, все исследования, проведенные по метаболическому перепрограммированию у растений с дефицитом железа, были проведены на всей корневой системе без дискриминации.
Однако в корнях растений действительно присутствуют разные ткани и, следовательно, разные «клеточные популяции». Было замечено, что при дефиците железа внешние клеточные слои корня преимущественно вовлечены в активацию типичных ферментов (т. е. железо-хелатредуктазы, H + -АТФаза), участвующие в механизме поглощения Fe из почвы (Santi, Schmidt, 2008). Кроме того, накопление белка H + -ATPase было особенно высоким в ризодермальных и энтодермальных клетках по сравнению с другими слоями Fe-дефицитных корней огурца (Dell’Orto et al., 2002). Таким образом, можно ли рассматривать ризодермальные клетки у растений с дефицитом Fe как глюкозозависимые клетки, тогда как корковые слои можно рассматривать как вторую субпопуляцию клеток, наблюдаемую в опухолях? Этот вопрос до сих пор остается открытым, так как нет данных о различном метаболическом перепрограммировании между разными слоями корня: действительно, все исследования, проведенные по метаболическому перепрограммированию у растений с дефицитом железа, были проведены на всей корневой системе без дискриминации.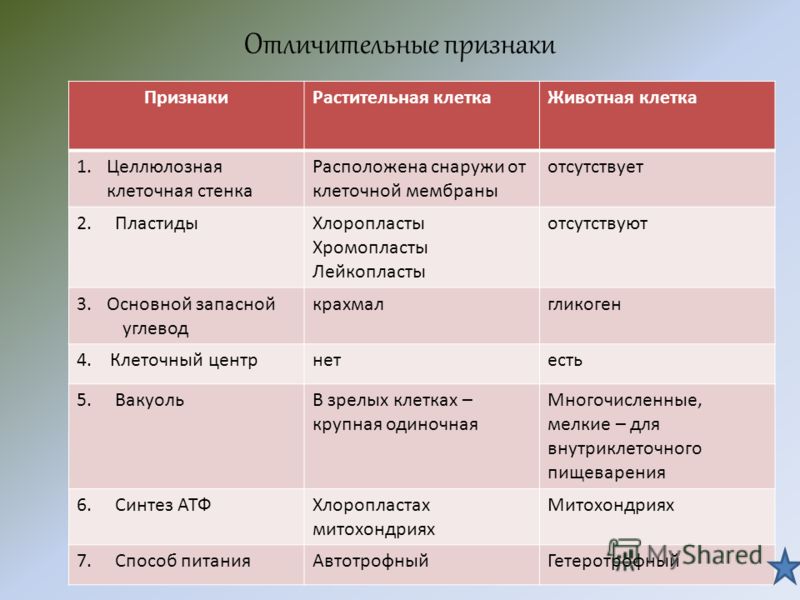 между различными тканями и слоями корня.
между различными тканями и слоями корня.
Еще одно важное различие между клетками растений с дефицитом Fe и опухолевыми клетками животных заключается в различном использовании глюкозы. Действительно, в опухолевых клетках животных эффект Варбурга поддерживается повышенным поглощением глюкозы пролиферирующими клетками (см. выше). В Fe-дефицитных растительных клетках нарушение фотосинтеза приводит к снижению доступности глюкозы во всем растении, что, по-видимому, противоречит тому, что наблюдается в опухолевых клетках. С другой стороны, у растений с дефицитом Fe, пока доступна глюкоза, она, по-видимому, направлена на поддержание перепрограммированного метаболизма, а не на производство биомассы, что приводит к снижению роста растений, что является общим следствием дефицита Fe у растений.
Одним из важных аспектов биологии опухолей в клетках животных является гипоксия. Как было сказано ранее, гипоксия важна для поздних событий в развивающихся раковых клетках и не является непосредственной причиной эффекта Варбурга.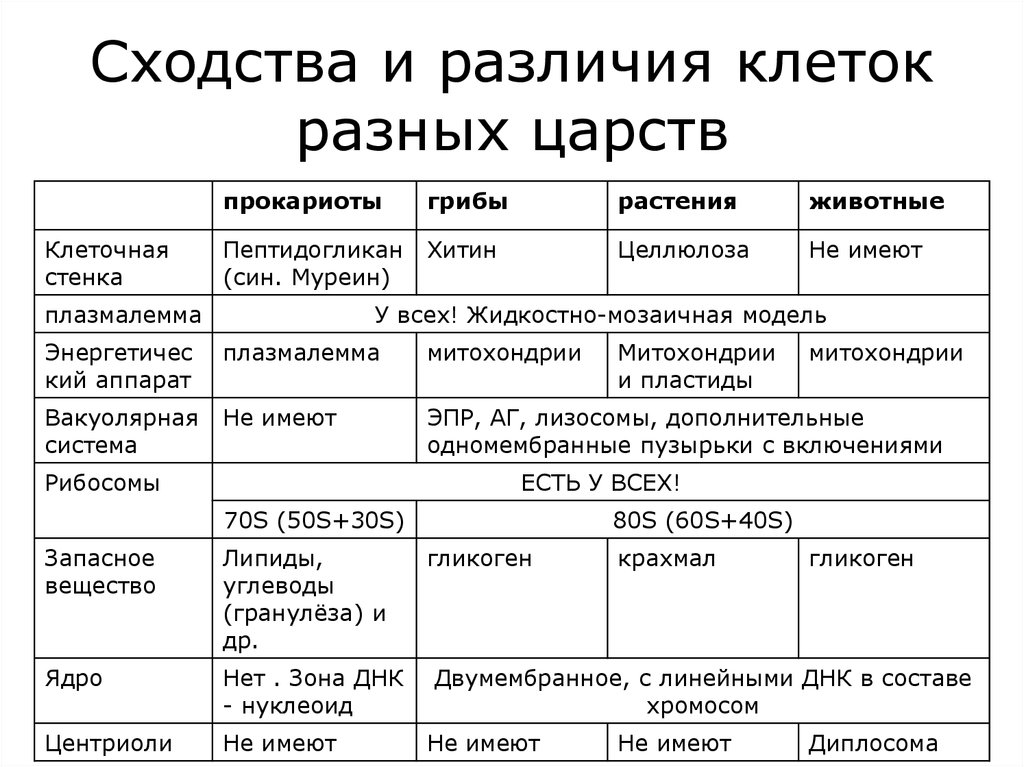 Таким образом, может быть важно рассмотреть вопрос о том, сравним ли процесс метаболической адаптации, вызывающий гипоксию, в растительных и животных клетках. Подобно тому, что наблюдается у животных, растительные клетки, подвергнутые гипоксии, демонстрируют аналогичные метаболические эффекты (Licausi, 2011). Действительно, наиболее очевидным следствием нехватки кислорода является дефицит энергии. Однако у растений энергетическая депривация является обычным следствием почти всех абиотических стрессов, отрицательно влияющих на фотосинтез и/или дыхание. В этих условиях клетки все еще способны производить АТФ посредством окислительного фосфорилирования, но скорость потребления кислорода снижается, и клетки оптимизируют использование энергии путем реорганизации метаболических потоков. С этой точки зрения метаболические изменения, происходящие при дефиците Fe, аналогичны изменениям, вызываемым гипоксией. Тем не менее, метаболические изменения, индуцируемые гипоксией, у растений опосредуются арсеналом транскрипционных факторов, отличных от системы HIF-P4DH, действующей в раковых клетках (Licausi et al.
Таким образом, может быть важно рассмотреть вопрос о том, сравним ли процесс метаболической адаптации, вызывающий гипоксию, в растительных и животных клетках. Подобно тому, что наблюдается у животных, растительные клетки, подвергнутые гипоксии, демонстрируют аналогичные метаболические эффекты (Licausi, 2011). Действительно, наиболее очевидным следствием нехватки кислорода является дефицит энергии. Однако у растений энергетическая депривация является обычным следствием почти всех абиотических стрессов, отрицательно влияющих на фотосинтез и/или дыхание. В этих условиях клетки все еще способны производить АТФ посредством окислительного фосфорилирования, но скорость потребления кислорода снижается, и клетки оптимизируют использование энергии путем реорганизации метаболических потоков. С этой точки зрения метаболические изменения, происходящие при дефиците Fe, аналогичны изменениям, вызываемым гипоксией. Тем не менее, метаболические изменения, индуцируемые гипоксией, у растений опосредуются арсеналом транскрипционных факторов, отличных от системы HIF-P4DH, действующей в раковых клетках (Licausi et al.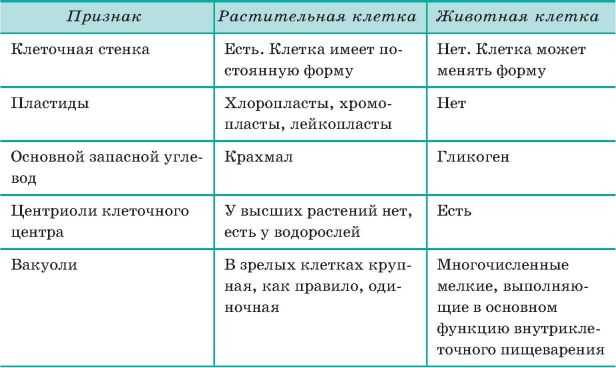 , 2012).
, 2012).
Интересно, что HIF-1 сотрудничает с геном c-myc, чтобы стимулировать аэробный гликолиз (Dang et al., 2008). C-myc является протоонкогеном, который регулирует экспрессию нескольких молекул, участвующих в гомеостазе Fe, включая переносчик Fe DMT1 и фратаксин. Следовательно, в клетках животных эффекты HIF-1 связаны с метаболизмом Fe. Действительно, высокая скорость пролиферации опухолевых клеток приводит к гипоксии и/или дефициту минералов в этих клетках. Фактически, в ответ на гипоксию способность эритроцитов транспортировать кислород повышается за счет экспрессии генов, участвующих в метаболизме Fe: гипоксия активирует трансферрин (Tf), который транспортирует Fe9.0163 3+ в клетки и церулоплазмин (известный также как ферроксидаза), который необходим для окисления ионов двухвалентного железа (Fe 2+ ) до ионов трехвалентного (Fe 3+ ). Повышение экспрессии этих генов поддерживает поступление Fe в клетку (Ke and Costa, 2006). Исследования, проведенные Callapina et al.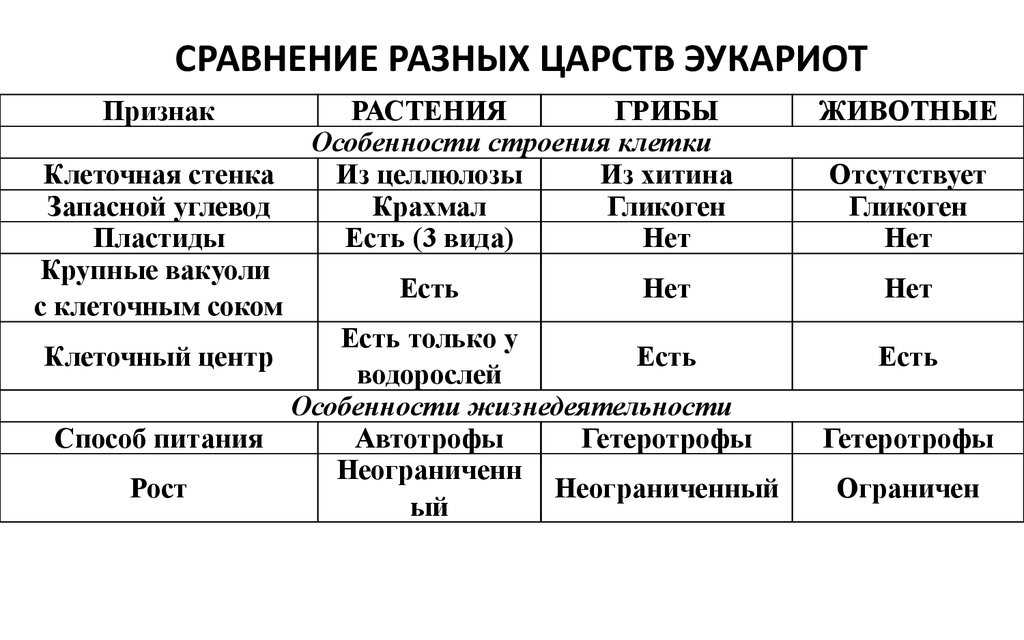 (2005) показали, что снижение уровня Fe также может имитировать состояние гипоксии. Поскольку Fe требуется для ферментативной активности пролилгидроксилазы, истощение внутриклеточного Fe приводит к инактивации фермента. Неактивный PHD не способен гидроксилировать HIF-1α, поэтому его последующая деградация не происходит, что приводит к транскрипции генов-мишеней HIF-1. Следовательно, в клетках животных дефицит Fe приводит к нарушению работы митохондрий вместе с увеличением как гликолиза, так и продукции лактата (Finch et al., 19).79). Собрав вместе все эти данные, становится ясно, что дефицит Fe вызывает сходные метаболические адаптации как в клетках растений, так и в клетках животных.
(2005) показали, что снижение уровня Fe также может имитировать состояние гипоксии. Поскольку Fe требуется для ферментативной активности пролилгидроксилазы, истощение внутриклеточного Fe приводит к инактивации фермента. Неактивный PHD не способен гидроксилировать HIF-1α, поэтому его последующая деградация не происходит, что приводит к транскрипции генов-мишеней HIF-1. Следовательно, в клетках животных дефицит Fe приводит к нарушению работы митохондрий вместе с увеличением как гликолиза, так и продукции лактата (Finch et al., 19).79). Собрав вместе все эти данные, становится ясно, что дефицит Fe вызывает сходные метаболические адаптации как в клетках растений, так и в клетках животных.
Понимание гомеостаза Fe в растениях значительно расширилось за последние годы. За последние несколько лет был достигнут значительный прогресс в понимании важности белков bHLH, таких как FIT, BHLH038, bHLH039 и PYE, как транскрипционных регуляторов ответов на дефицит железа (Ivanov et al., 2012).
А как насчет регуляции сложного метаболического перепрограммирования? Регулируются ли метаболические изменения в Fe-дефицитных растениях так же, как метаболические изменения, происходящие в опухолевых клетках? Несколько PHD-подобных изоформ (например, P4Hs) были клонированы и охарактеризованы в растениях (Vlad et al.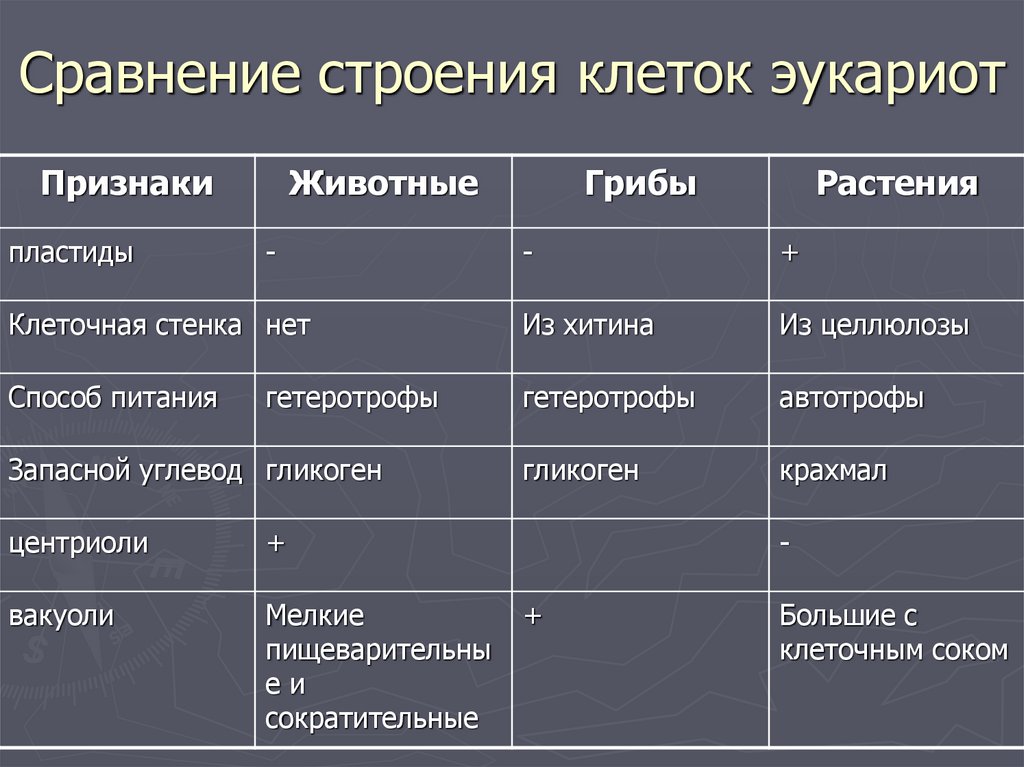 , 2007a). Хотя нет данных о присутствии гомологов HIF-1 в растениях, нельзя исключать возможность того, что P4Hs играют роль, сходную с ролью HIF-P4Hs в ответ на абиотические стрессы (Vlad et al., 2007b). Действительно, рекомбинантный 9Было обнаружено, что 0159 Arabidopsis P4H (At-P4H-1) гидроксилирует синтетические пептиды, представляющие две гидроксилированные последовательности фактора транскрипции животных HIF-1α, что указывает на специфичность субстрата для фактора транскрипции из гетерологичного организма (Hieta and Myllyharju, 2002). Однако, даже если нет четких доказательств наличия HIF-1-P4H-подобной системы у растений, косвенные данные не позволяют исключить наличие таких белков у растений.
, 2007a). Хотя нет данных о присутствии гомологов HIF-1 в растениях, нельзя исключать возможность того, что P4Hs играют роль, сходную с ролью HIF-P4Hs в ответ на абиотические стрессы (Vlad et al., 2007b). Действительно, рекомбинантный 9Было обнаружено, что 0159 Arabidopsis P4H (At-P4H-1) гидроксилирует синтетические пептиды, представляющие две гидроксилированные последовательности фактора транскрипции животных HIF-1α, что указывает на специфичность субстрата для фактора транскрипции из гетерологичного организма (Hieta and Myllyharju, 2002). Однако, даже если нет четких доказательств наличия HIF-1-P4H-подобной системы у растений, косвенные данные не позволяют исключить наличие таких белков у растений.
Подводя итог, я показал здесь интересное сходство между растениями и животными, чтобы спровоцировать открытое обсуждение новой интерпретации метаболического перепрограммирования Fe-дефицитных растительных клеток как эффекта Варбурга. Метаболические изменения, происходящие в опухолевых клетках, рассматриваются как отличительный признак рака, и их понимание приводит к характеристике ахиллесовой пяты опухоли, стимулируя определение арсенала терапевтических соединений для использования во время химиотерапии.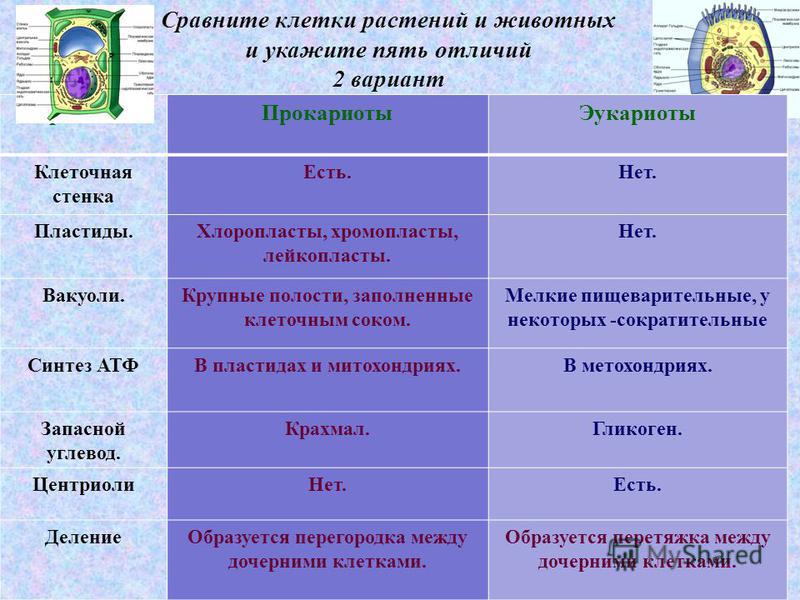 Можно ли с этой точки зрения считать метаболическое перепрограммирование, происходящее при дефиците Fe у растений, отличительной чертой адаптационного механизма, выработанного растениями для преодоления нарушения питания?
Можно ли с этой точки зрения считать метаболическое перепрограммирование, происходящее при дефиците Fe у растений, отличительной чертой адаптационного механизма, выработанного растениями для преодоления нарушения питания?
В целом, было бы интересно сосредоточить исследования на регуляции процессов метаболической адаптации, чтобы идентифицировать другие гены-кандидаты, способные определять толерантность к дефициту Fe у растений.
Благодарности
Я хочу поблагодарить профессора Грациано Зокки и доктора Марту Делл’Орто за критическое прочтение рукописи. Я также хочу поблагодарить редактора и рецензентов за их ценные комментарии по улучшению рукописи. Автора поддержала «Dote Ricerca»: FSE, Regione Lombardia.
Ссылки
Брахими-Хорн, М.С., Чиче, Дж., и Пуйессегюр, Дж. (2007). Сигнализация гипоксии контролирует метаболические потребности. Курс. мнение Клеточная биол. 19, 223–229.
Опубликован Аннотация | Опубликован полный текст | CrossRef Полный текст
Каллапина М.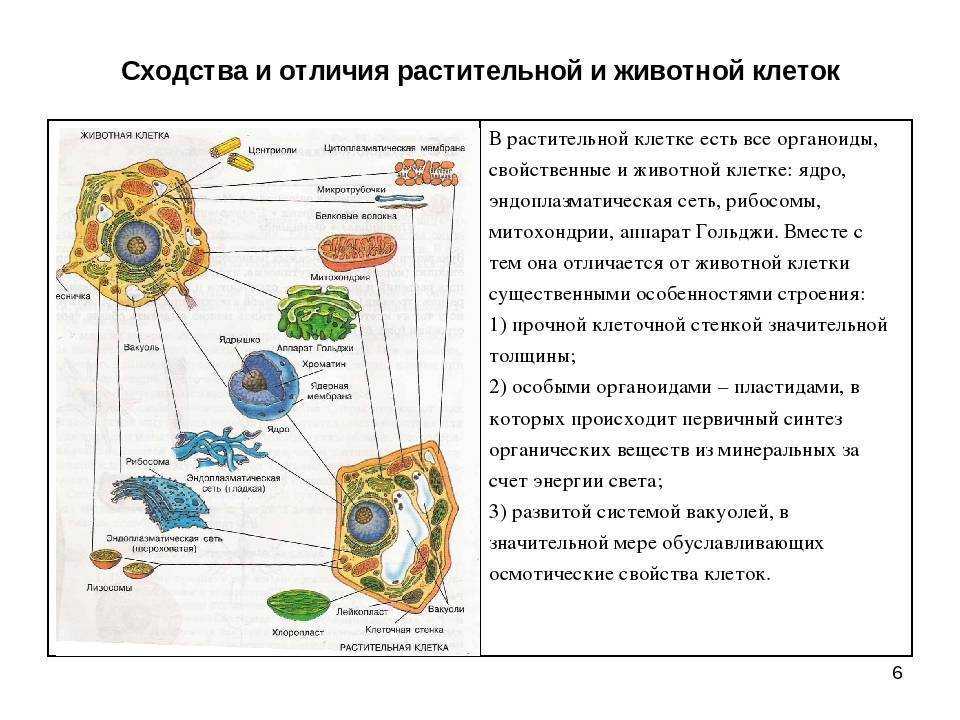 , Чжоу Дж., Шнитцер С., Метцен Э., Лор К., Дейтмер Дж. В. и Брюн Б. (2005). Оксид азота обращает накопление HIF-1 альфа, вызванное десферриоксамином и гипоксией, что влияет на активность пролилгидроксилазы и железо. Экспл. Сотовый рез. 306, 274–284.
, Чжоу Дж., Шнитцер С., Метцен Э., Лор К., Дейтмер Дж. В. и Брюн Б. (2005). Оксид азота обращает накопление HIF-1 альфа, вызванное десферриоксамином и гипоксией, что влияет на активность пролилгидроксилазы и железо. Экспл. Сотовый рез. 306, 274–284.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Кэрью, Дж. С., и Хуанг, П. (2002). Митохондриальные дефекты при раке. BMC Мол. Можно. 1, 9.
CrossRef Полный текст
Данг, К.В., Кин, Дж.В., Гао, П., и Юстейн, Дж. (2008). Взаимодействие между MYC и HIF при раке. Нац. Преподобный Рак 8, 51–56.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Делл’Орто, М., Пировано, Л., Вильяльба, Дж. М., Гонсалес-Рейес, Дж. А., и Зокки, Г. (2002). Локализация Н+-АТФазы плазматической мембраны в Fe-дефицитных корнях огурца методом иммунодетекции. Почва для растений 241, 11–17.
CrossRef Полный текст
Доннини С., Принси Б. , Негри А.С., Вигани Г., Эспен Л. и Зокки Г. (2010). Протеомная характеристика реакции дефицита железа в корнях Cucumis sativus L.. BMC Растение Биол. 10, 268.
, Негри А.С., Вигани Г., Эспен Л. и Зокки Г. (2010). Протеомная характеристика реакции дефицита железа в корнях Cucumis sativus L.. BMC Растение Биол. 10, 268.
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Финч, К.А., Колник, П.Д., Хластала, М.П., Миллер, Л.Р., Диллманн, Э., и Маклер, Б. (1979). Лактоацидоз в результате дефицита железа. Дж. Клин. Инвестировать. 64 129–137.
Фишер, К., Хоффманн, П., Фёлькль, С., Мейденбауэр, Н., Аммер, Дж., Эдингер, М., Готфрид, Э., Шварц, С., Роте, Г., Ховес, С. ., Реннер К., Тимишль Б., Макенсен А., Кунц-Шугхарт Л., Андрисен Р., Краузе С.В. и Кройц М. (2007). Ингибирующее действие молочной кислоты, полученной из опухолевых клеток, на Т-клетки животных. Кровь 109, 3812–3819.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Гейтенби, Р. А., и Гиллис, И. П. (2004). Почему рак имеет высокий аэробный гликолиз? Нац. Преподобный Рак 4, 891–899.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Ханахан, Д., и Вайнберг, Р. А. (2011). Отличительные признаки рака: следующее поколение. Сотовый 144, 646–674.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Harris, AL (2002). Гипоксия – ключевой регуляторный фактор роста опухоли. Нац. Преподобный Рак 2, 38–47.
Опубликован Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Хиета, Р., и Мюллихарью, Дж. (2002). Клонирование и характеристика низкомолекулярной пролил-4-гидроксилазы из Arabidopsis thaliana . Эффективное гидроксилирование богатых пролином, коллагеноподобных и индуцируемых гипоксией факторов транскрипции альфа-подобных пептидов. Дж. Биол. хим. 277, 23965–23971.
Опубликован Аннотация | Опубликован полный текст | CrossRef Full Text
Иванов Р., Брумбарова Т. и Бауэр П. (2012). Приспособление к суровой реальности: регуляция реакции дефицита железа у двудольных растений. Мол Завод 5, 27–42.
Мол Завод 5, 27–42.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Ке, К., и Коста, М. (2006). Индуцируемый гипоксией фактор-1 (HIF-1). Мол. фарм. 70, 1469–1480.
CrossRef Full Text
Кукуракис М.И., Гиатроманолаки А., Харрис А.Л. и Сивридис Э. (2006). Сравнение метаболических путей между раковыми клетками и стромальными клетками при колоректальных карциномах: роль метаболического выживания стромы, связанной с опухолью. Рак рез. 66, 632–637.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Licausi, F. (2011). Регуляция молекулярного ответа на ограничение кислорода в растениях. Новый Фитол. 190, 550–555.
Опубликован Аннотация | Опубликован полный текст | CrossRef Full Text
Licausi, F., Kosmacz, M., Weits, D.A., Giuntoli, B., Giorgi, F.M., Voesenek, L.A., Perata, P., and van Dongen, J.T. (2012). Восприятие кислорода у растений опосредовано путем правила N-конца для дестабилизации белка.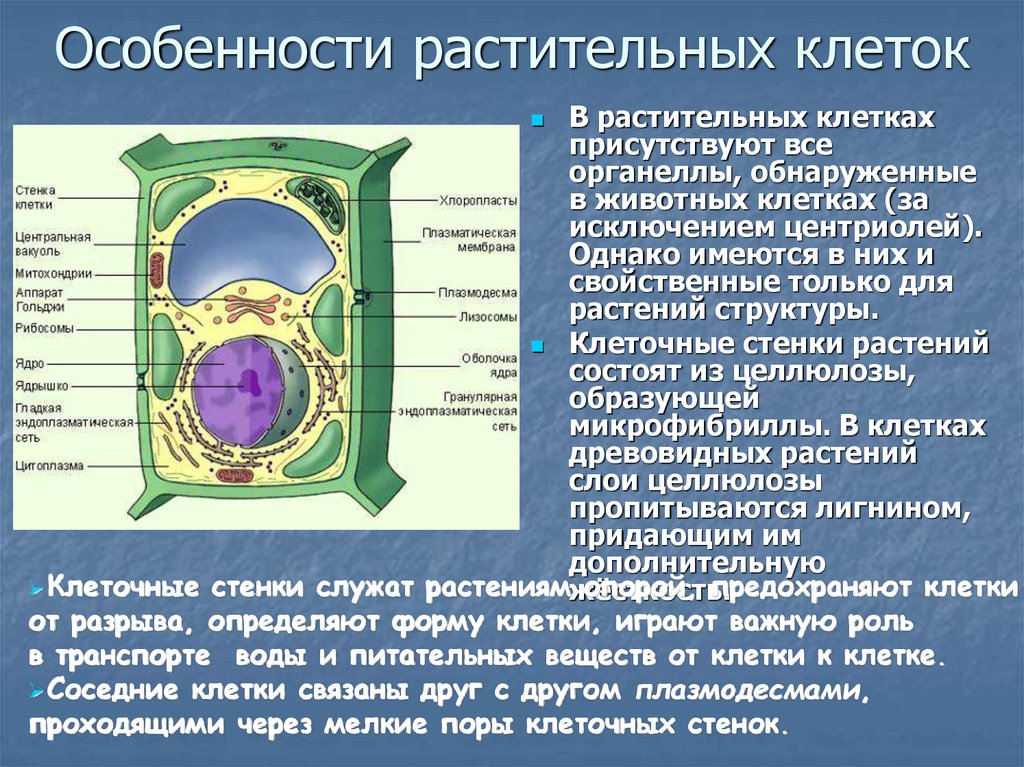 Природа 479, 419–422. (2000). Реакция корнеплодов сахарной свеклы на дефицит железа. Изменения в ассимиляции углерода и использовании кислорода. Завод физиол. 124, 885–897.
Природа 479, 419–422. (2000). Реакция корнеплодов сахарной свеклы на дефицит железа. Изменения в ассимиляции углерода и использовании кислорода. Завод физиол. 124, 885–897.
CrossRef Полный текст
Лопес-Миллан, А. Ф., Моралес, Ф., Гогорсена, Ю., Абадия, А., и Абадия, Дж. (2009). Метаболические реакции у растений томатов с дефицитом железа. J. Физиол растений. 166, 375–384.
CrossRef Full Text
Реллан-Альварес, Р., Андалус, С., Родригес-Сельма, Дж., Вольгемут, Г., Зокки, Г., Альварес-Фернандес, А., Фин, О., Лопес- Миллан, А.Ф., и Абадия, Дж. (2010). Изменения в протеомных и метаболических профилях кончиков корней Beta vulgaris в ответ на дефицит железа и пополнение запасов. BMC Растение Биол. 10, 120.
CrossRef Полный текст
Санти, С., и Шмидт, В. (2008). Анализ функциональной судьбы корневых волосков огурца, вызванных дефицитом железа, с помощью лазерной микродиссекции. Дж. Эксп. Бот. 59 697–704.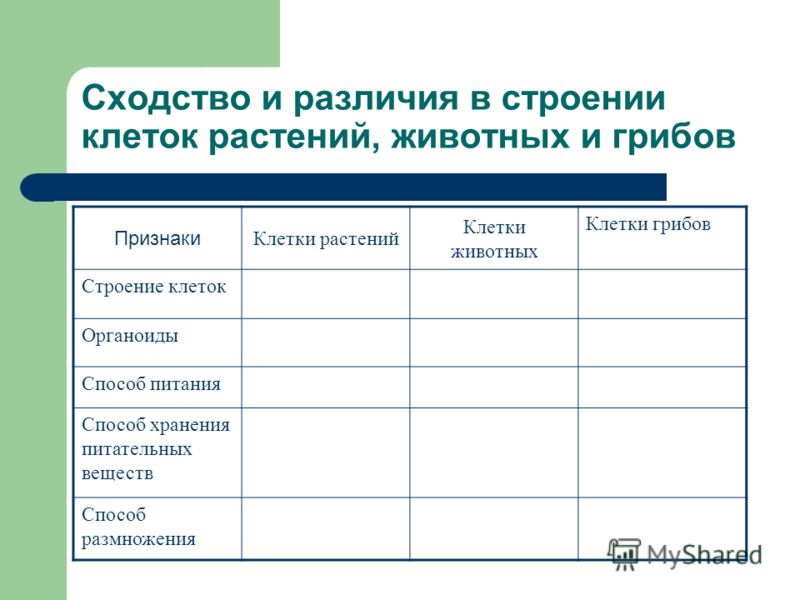
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Semenza, GL (2007). Путь фактора 1, индуцируемого гипоксией (HIF-1). Наука СТКЭ. 2007 г., см8.
CrossRef Full Text
Swietach, P., Vaughan-Jones, R.D., and Harris, A.L. (2007). Регуляция pH опухоли и роль карбоангидразы 9. Cancer Metastasis Rev. 26, 299–310.
Опубликован Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Тейлор, К.Т., и Пуиссегюр, Дж. (2007). Кислород, гипоксия и стресс. Энн. Н. Я. акад. науч. 1113, 87–94.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Тимм О., Эссигманн Б., Клоска С., Альтманн Т. и Бакхаут Т. Дж. (2001). Реакция арабидопсиса на стресс дефицита железа, выявленная микрочиповым анализом. Завод физиол. 127, 1030–1043.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Вандер Хейден, М. Г., Кэнтли, Л.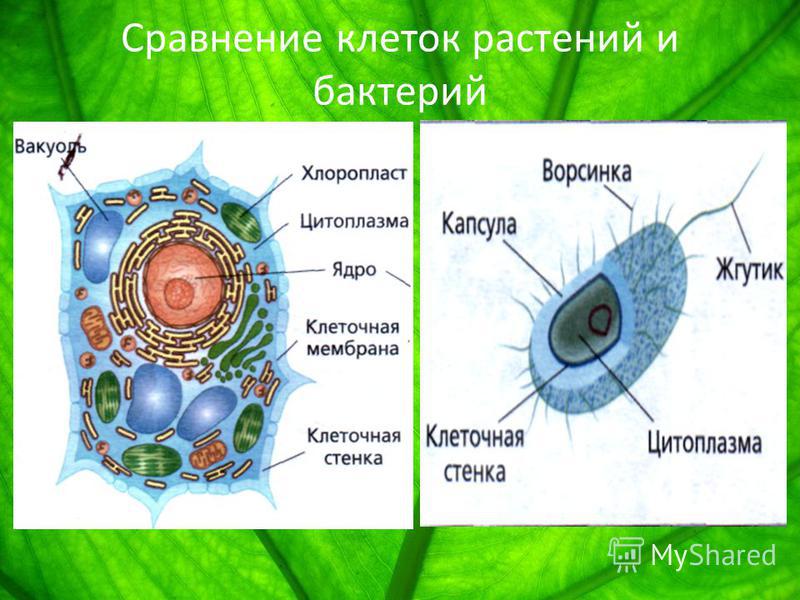 К., и Томпсон, К. Б. (2009 г.). Понимание эффекта Варбурга: метаболические потребности пролиферации клеток. Наука 324, 1029–1033.
К., и Томпсон, К. Б. (2009 г.). Понимание эффекта Варбурга: метаболические потребности пролиферации клеток. Наука 324, 1029–1033.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Вигани, Г. (2012). Обнаружение роли митохондрий в метаболических реакциях растений, вызванных дефицитом железа. J. Физиол растений. 168, 1–11.
CrossRef Полный текст
Влад, Ф., Спано, Т., Влад, Д., Буо Дахер, Ф., Уэлхадж, А., и Калаитцис, П. (2007a). Пролил-4-гидроксилазы арабидопсиса по-разному экспрессируются в ответ на гипоксию, аноксию и механические повреждения. Завод Физиол 130, 471–483.
CrossRef Полный текст
Влад, Ф., Спано, Т., Влад, Д., Буо Дахер, Ф., Уэльхадж, А., Фрагкостефанакис, С., и Калайцис, П. (2007b). Участие пролил-4-гидроксилаз арабидопсиса при гипоксии, аноксии и механических повреждениях. Поведение сигнала установки. 2, 638–369.
CrossRef Полный текст
Варбург, О.