
Содержание
Кукушкин лен: строение и размножение
Кукушкин лен: строение и размножение
Кукушкинский лен — растение, которое на территории Российской Федерации наиболее распространено в лесах северной и центральной зоны. Благоприятные условия для этого наблюдаются в заболоченных лесах тайги, болотах и сырых лугах. Растение относится к роду листовых мхов, к отделу мохообразных. На планете встречается более сотни его разновидностей. Кукушкинское полотно, образующее подушкообразные пучки, часто встречается в тундре и высокогорье. Наибольшее распространение на территории стран СНГ получил политрихум обыкновенный (второе название растения).
Содержание
- 1 Большое значение солнца
- 2 Описание
- 3 Происхождение названия
- 4 Строение кукушкина льна
- 5 Функции стебля
- 6 Размножение и развитие
- 7 Использование в различных целях
- 8 Природный утеплитель
- 9 Мох сфагнум
- 10 Чем полезен сфагнум?
Большое значение солнца
Кукушкинское белье очень любит свет. Вот почему в темных пихтовых лесах, даже если почва влажная и плодородная, она будет ограничена в росте и развитии. При достаточном количестве солнечного света растение быстро тянется, активно захватывая новые участки и покрывая землю плотным ковром. Земля под кукушкой сохнет намного медленнее, поэтому ее рост постепенно приводит к заболачиванию местности.
Вот почему в темных пихтовых лесах, даже если почва влажная и плодородная, она будет ограничена в росте и развитии. При достаточном количестве солнечного света растение быстро тянется, активно захватывая новые участки и покрывая землю плотным ковром. Земля под кукушкой сохнет намного медленнее, поэтому ее рост постепенно приводит к заболачиванию местности.
Описание
Кукушечный льняной мох отличается довольно высокими стеблями (длина их 10-15 сантиметров, но встречаются и сорок сантиметровые растения). Проводящая система обеспечивает движение воды и питательных веществ по стеблю.
Происхождение названия
У описываемого растения прямые коричневатые стебли. На них небольшие листочки темно-зеленого оттенка, напоминающие миниатюрный лен. Но коробочки, которые появляются на женских растениях, вызывают ассоциации с кукушкой, сидящей на каком-то шесте.
Строение кукушкина льна
Рассматриваемое растение относят к многолетним листовым мхам. Его размеры большие, в нижней части стебля расположены ризоиды — примитивные аналоги корней. На первичном горизонтальном стебле листьев нет. Вторичный стебель может быть простым или разветвленным. Он прямостоячий, средней длины пятнадцать сантиметров. Каждый лист имеет большую главную жилку. Кукушкинский лен, строение которого достаточно простое, имеет чешуйчатые нижние листья.
На первичном горизонтальном стебле листьев нет. Вторичный стебель может быть простым или разветвленным. Он прямостоячий, средней длины пятнадцать сантиметров. Каждый лист имеет большую главную жилку. Кукушкинский лен, строение которого достаточно простое, имеет чешуйчатые нижние листья.
Функции стебля
Основная роль этой части растения — опора. Не менее важна проводящая способность стебля. Он действует как связующее звено между листьями и корневой системой. Шток также выполняет некоторые второстепенные функции. Среди них — сохранение запаса питательных веществ.
Размножение и развитие
Размножается растение следующими способами: половым (гаметы) и бесполым (споры, побеги). Они чередуются.
Как именно размножается льняное растение с кукушкой? Споры, которые образует растение, находятся в спорангиях (капсуле) на стебле. После созревания они появляются из этого естественного месторождения. При благоприятных условиях споры образуют многоклеточную нить, а из нее, в свою очередь, появляются несколько гаметофитов (происходит это почкованием). Зеленый многолетний побег с листьями и ризоидами (корневидными образованиями) считается гаметофитом. Последние берут из почвы соль и йод. Клетки листа обеспечивают синтез всех других незаменимых веществ. Исходя из этого, можно утверждать, что гаметофит — самостоятельный организм.
Зеленый многолетний побег с листьями и ризоидами (корневидными образованиями) считается гаметофитом. Последние берут из почвы соль и йод. Клетки листа обеспечивают синтез всех других незаменимых веществ. Исходя из этого, можно утверждать, что гаметофит — самостоятельный организм.
Через некоторое время гаметофит перестает расти. Затем кукушка-лен начинает размножаться. В центре розетки листьев (расположенной в верхней части стебля) развиваются мужские и женские половые органы. Первые представлены антеридиями (название происходит от греческого слова «антерос», что означает «цветение»), у которых цикл развития подвижных гамет — сперматозоидов, а также архегоний — женских половых органов, ответственных за формирование неподвижная женская гамета — яйцо.
Мужские растения характеризуются наличием более крупных желто-коричневых листьев. У женских экземпляров таких листьев нет.
С наступлением сезона дождей или наводнения сперматозоиды (мужские клетки) могут подплывать к яйцеклетке. В результате они объединяются. В конце процесса оплодотворения появляется зигота (это слово происходит от греческого «zygotos», что переводится как «связанный»). Это первый этап развития эмбриона. На следующий год из оплодотворенной зиготы развивается капсула (спорогон), помещенная на довольно длинный стебель без листьев. В будущем ящик станет местом развития спор. Это природное месторождение очень хрупкое. Также качается от легкого ветерка. После того, как шляпка отпадет и споры отпадут, наблюдается прорастание зеленой ветвящейся нити — предварительный рост. Учтите, что для положительного результата необходимо, чтобы споры попадали в благоприятную для них среду, в таком случае кукушка-лен будет размножаться.
В результате они объединяются. В конце процесса оплодотворения появляется зигота (это слово происходит от греческого «zygotos», что переводится как «связанный»). Это первый этап развития эмбриона. На следующий год из оплодотворенной зиготы развивается капсула (спорогон), помещенная на довольно длинный стебель без листьев. В будущем ящик станет местом развития спор. Это природное месторождение очень хрупкое. Также качается от легкого ветерка. После того, как шляпка отпадет и споры отпадут, наблюдается прорастание зеленой ветвящейся нити — предварительный рост. Учтите, что для положительного результата необходимо, чтобы споры попадали в благоприятную для них среду, в таком случае кукушка-лен будет размножаться.
На предростках образуются почки, из которых выходят женские и мужские экземпляры растения. Таким образом, можно видеть, что жизненный цикл развития мхов включает последовательное чередование бесполого и полового поколений. В ходе эволюции этот признак развился у многих растений, в том числе у кукушечного льна.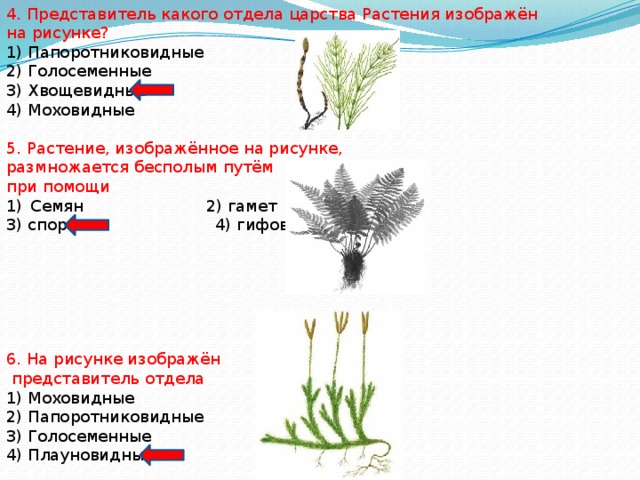
Вегетативное размножение этого вида мха позволяет легко получить на приусадебном участке толстый зеленый ковер. Просто поместите небольшой кусочек мха во влажное место. Однако следует учитывать способность этого растения застаиваться на площади своего обитания.
Использование в различных целях
Если удалить листья с кукушек льна, можно получить гибкие и жесткие нити, образованные центральными стеблями. Наши предки использовали этот натуральный материал для изготовления щеток и веников. После замачивания и чистки стебли стали отличной основой для циновок, ковриков, корзин и затемненных штор. Интересно, что при раскопках римского форта в Англии были обнаружены остатки корзин из кукушечного полотна. Продукция датируется 86 годом нашей эры.
Раньше кукушечный лен широко использовался при изготовлении плащей для воинов и путешественников. Полученная в результате одежда была особенно прочной. Они также имели декоративное значение.
Целители рекомендуют использовать этот вид мха для оздоровления пищеварительной системы, устранения желудочных колик и растворения камней в почках и желчном пузыре.
Лен кукушкин, структура которого позволяет использовать его в садоводстве в декоративных целях, положительно влияет на почву. Таким образом, это растение способно нормализовать показатели кислотности почвы до двух сезонов. Тогда любое садовое растение можно успешно выращивать на восстановленной почве. Мертвый мох — отличное удобрение.
Самым необычным является использование кукурузного льна в качестве солода при производстве виски.
Природный утеплитель
Кукушкинское белье способно эффективно защищать конструкцию от проникновения холода и влаги. Очень ценится то, что мох не гниет. Его размещение между бревнами сруба обеспечивает естественную вентиляцию. Для этого используют мох в свежем виде. Перед укладкой натурального утеплителя его необходимо тщательно очистить от веток, палочек, шишек, травы и других включений.
Мох сфагнум
Это растение относится к роду белого мха (торфа). Награждено 320 видов. Сфагнум преимущественно представлен болотными мхами, которые образуют плотные скопления, образующие большие подушки или толстые ковры в сфагновых болотах.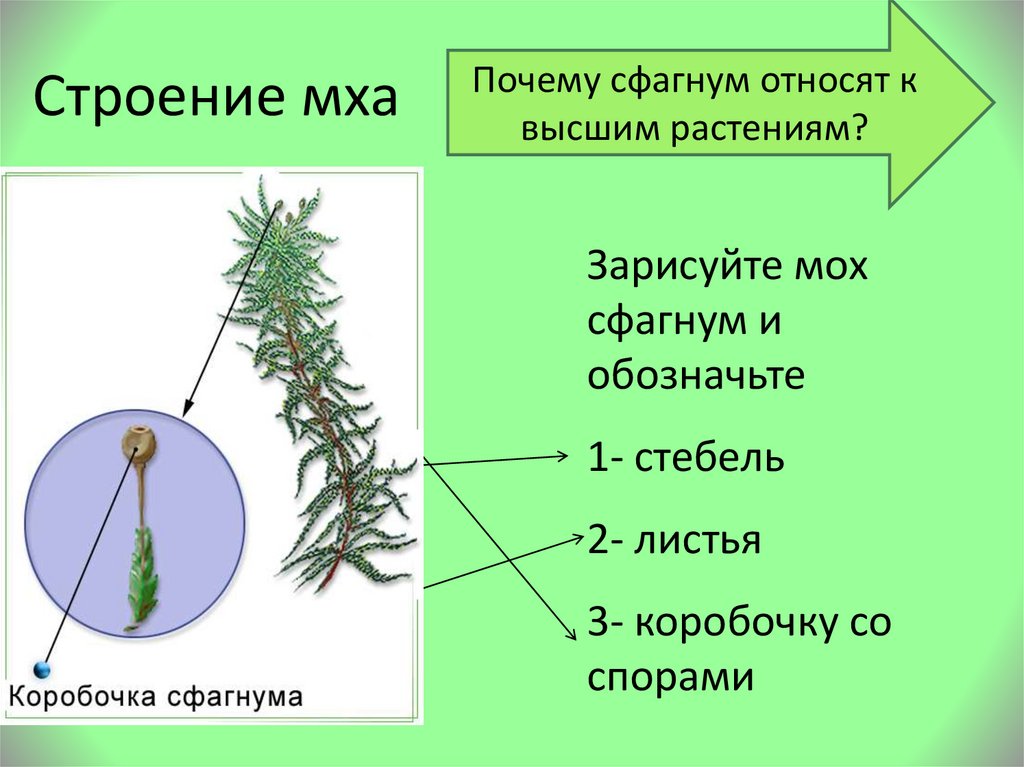 Но во влажных лесах сфагнум встречается гораздо реже. Льняной Кукушкин это растение напоминает своим прямостоячим стеблем, достигающим десяти-двадцати сантиметров в высоту. Листья сфагнума однослойные, расположены на пучковидных ветвях. В листьях много водяных клеток с порами, которые активно впитывают воду. Этот факт определяет повышенную влажность растения. В местах появления этих мхов быстро развиваются болота.
Но во влажных лесах сфагнум встречается гораздо реже. Льняной Кукушкин это растение напоминает своим прямостоячим стеблем, достигающим десяти-двадцати сантиметров в высоту. Листья сфагнума однослойные, расположены на пучковидных ветвях. В листьях много водяных клеток с порами, которые активно впитывают воду. Этот факт определяет повышенную влажность растения. В местах появления этих мхов быстро развиваются болота.
Ежегодно стебли в нижней части растения отмирают. Они образуют торф. Дальнейший рост стебля обеспечивают верхушечные ветви.
Отметим, что сфагнумы играют важную роль в формировании и существовании торфяных болот. Как было сказано выше, участки мертвого мха образуют торфяные отложения. Образование торфа возможно из-за застоя воды, вклада кислых мхов и недостатка кислорода. В этих условиях не происходит процессов распада, сфагнум не разлагается. Торф — ценный продукт, из которого получают воск, аммиак, парафин, спирт и др. он широко применяется в медицинской практике, строительстве. Мох действует как эффективное биотопливо и удобрение.
Мох действует как эффективное биотопливо и удобрение.
Чем полезен сфагнум?
Этот компонент включен во многие рецепты народной и официальной медицины. А все потому, что мох сфагнум — отличный антисептик и надежный перевязочный материал. Помогает заживлять гнойные раны благодаря способности впитывать большое количество влаги. По этому показателю сфагнум превосходит лучшие сорта впитывающей ваты. Этот мох обладает бактерицидным действием благодаря наличию сфагнума, особого фенолоподобного вещества, подавляющего развитие и жизнедеятельность кишечной палочки, холерного вибриона, золотистого стафилококка, сальмонеллы и некоторых других патогенных микроорганизмов.
Флористы активно используют сфагнум для выращивания комнатных растений. Он является составной частью субстрата, слоя мульчи или выполняет дренажные функции. Мох не богат питательными веществами, но придает почве необходимую рыхлость. Отличная гигроскопичность сфагнума обусловлена его способностью равномерно распределять влагу. Наличие сфагноло определяет бактерицидные свойства описываемого вида мха, что позволяет эффективно вылечить корни основного растения, предотвращая развитие болезней и кариеса.
Наличие сфагноло определяет бактерицидные свойства описываемого вида мха, что позволяет эффективно вылечить корни основного растения, предотвращая развитие болезней и кариеса.
Поделиться:
- Предыдущая записьГеографическое положение тайги. Особенности географического положения тайги
- Следующая записьКак понять, почему «плюс» на «минус» дает «минус» ?
×
Рекомендуем посмотреть
Adblock
detector
Мхи — торфообразователи
К мхам относятся низкие (высотой до 10—20 см) растения, состоящие из одного или нескольких стебельков со спирально расположенными на них листьями.
Корней мхи не имеют: у некоторых из них корни заменяются волосковидными многоклеточными образованиями, так называемыми ризоидами.
Размножаются мхи вегетативно или спорами, созревающими в коробочках. Спора, прорастая, образует предросток, или протонему, в виде сплетения зеленых нитей или в виде пластинки. Предросток развивает многочисленные ризоиды, прикрепляющие его к почве, и почку, из которой вырастает потом молодое растеньице.
Спора, прорастая, образует предросток, или протонему, в виде сплетения зеленых нитей или в виде пластинки. Предросток развивает многочисленные ризоиды, прикрепляющие его к почве, и почку, из которой вырастает потом молодое растеньице.
Вегетативное размножение у мхов происходит путем ежегодного возникновения в их головках ветви, развивающейся в новый стебель, принимающий в процессе роста направление главного стебля. Растение кажется вильчато-разделенным до тех пор, пока при постепенном отмирании стебля в нижней его части молодой стебель не обособится от старого в отдельное растение.
На болотах встречаются мхи двух порядков: зеленые (бурые) мхи (Bryales) и сфагновые (белые торфяные) мхи (Sphagnales).
Зеленые (бурые) мхи (Bryales). В порядок зеленых мхов входит несколько семейств. Название одного из этих семейств Hypnaceae часто относят ко всему порядку зеленых мхов, называя его порядком гипновых мхов.
Это — большая, богатая видами группа мхов. Зеленые мхи встречаются на минеральных и торфяных почвах, где они образуют сплошные дернины в виде зеленых или бурых ковров или подушек. По окраске и строению зеленые мхи весьма разнообразны. Высота растения обычно около 10 см, но у отдельных видов она достигает иногда 15—35 см (кукушкин лен). Стебельков чаще несколько и они иногда ветвисты. Форма стебля округлая или (редко) трехгранная. Ризоиды в виде войлока опушают нижнюю часть стебля. Ими растение укрепляется в почве и поглощает воду и растворенные в ней вещества. На стебельках спирально располагаются листья различных оттенков (зеленые, желто-золотистые и бронзовые). Форма листьев для разных семейств и родов довольно разнообразна: от круглой до заостренной, иногда заостренной с загнутой верхушкой.
По окраске и строению зеленые мхи весьма разнообразны. Высота растения обычно около 10 см, но у отдельных видов она достигает иногда 15—35 см (кукушкин лен). Стебельков чаще несколько и они иногда ветвисты. Форма стебля округлая или (редко) трехгранная. Ризоиды в виде войлока опушают нижнюю часть стебля. Ими растение укрепляется в почве и поглощает воду и растворенные в ней вещества. На стебельках спирально располагаются листья различных оттенков (зеленые, желто-золотистые и бронзовые). Форма листьев для разных семейств и родов довольно разнообразна: от круглой до заостренной, иногда заостренной с загнутой верхушкой.
Строение листа довольно простое: пластина его на большей площади однослойна и состоит из вытянутых, округлых или многогранных клеток. В середине пластинки нередко встречается пучок более сближенных, многослойно расположенных клеток — это так называемая жилка. У некоторых родов зеленых мхов эта жилка занимает большую часть пластинки (например, у Polytrichum). У многих родов в углах при основании листа округлые прозрачные клетки образуют в совокупности так называемые ушки.
В растительном покрове торфяных месторождений находится целый ряд семейств, принадлежащих к порядку зеленых мхов. Главнейшие из них: Hypnaceae, Meesiaceae и Polytrichaceae.
Из наиболее часто встречающихся на торфяных месторождениях видов к семейству Hypnaceae относятся: Drepanocladus vernicosus, D. sendtneri, D. aduncus, D. fluitans, Calliergon giganteum, C. slramineum, C. cordifolium, Calliergonella cuspidata, Tomenthypnum nitens; к семейству Meesiaceae относятся виды Paludella squarrosa, Meesia triquetra, к семейству Polytrichaceae — Polvtrichum strictum, P. commune.
Основным систематическим отличием между отдельными родами и видами зеленых мхов являются форма и характер строения листовой пластинки.
Для Drepanocladus характерны вытянутые и загнутые вверху листья с ясно выраженной посередине жилкой, для Calliergon — сердцевидно-яйцевидная или продолговато-языковидная форма листа. Жилка и ушки выражены у Calliergon giganteum, С. stramineum и С. cordifolium; у Calliergonella cuspidata жилка почти не выражена. Для Tomenthypnum характерны вытянутые узкие, заостренные листья с продольными складками.
Для Tomenthypnum характерны вытянутые узкие, заостренные листья с продольными складками.
Из семейства Meesiaceae роды Meesia и Paludella имеют языковидно-ланцетные листья, отогнутые назад, с отчетливым килем, по которому лист сложен, и пильчатым краем. Отличие Meesia от Paludella выражается наличием в клетках верхней части листа Paludella выростов (сосочков), отсутствующих у Meesia.
Для мхов семейства Polytrichaceae характерна линейно-ланцетная листовая пластинка, почти на всем протяжении многослойная.
Зеленые мхи весьма требовательны к условиям минерального питания, поэтому в растительном покрове они встречаются главным образом на низинных болотах, где грунтовые воды несут повышенное содержание минеральных соединений. На верховых болотах встречаются менее прихотливые к условиям минерального питания Drepanocladus fluitans и Polytrichum strictum. Зеленые мхи принимают большое участие в заболачивании минеральных почв.
В прошлые эпохи зеленые мхи играли значительно большую роль как в растительном покрове, так и в сложении торфяных залежей.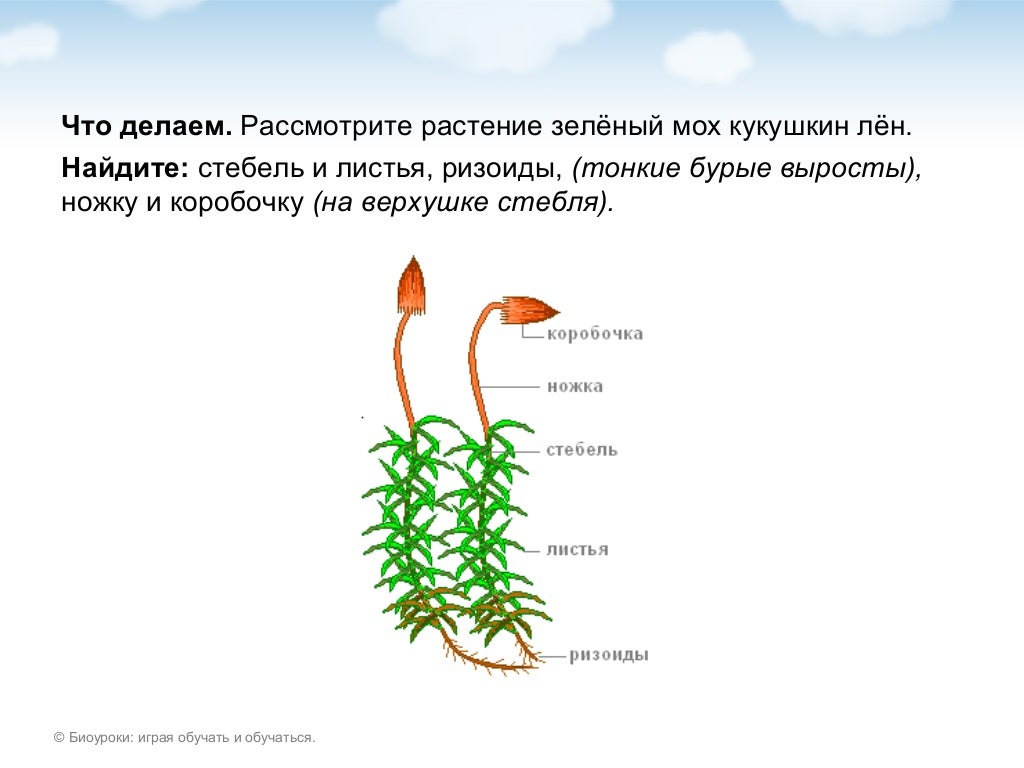 Об этом свидетельствуют довольно мощные слои гипнового торфа в основании низинных и верховых торфяников и слои межледникового торфа, почти нацело сложенные зелеными мхами.
Об этом свидетельствуют довольно мощные слои гипнового торфа в основании низинных и верховых торфяников и слои межледникового торфа, почти нацело сложенные зелеными мхами.
Сфагновые (торфяные) мхи(Sphagnales). Все сфагновые мхи (в СССР встречается около 40 разновидностей) принадлежат к одному семейству Sphagnaceae, объединяющему один только род Sphagnum. Сфагновые мхи наиболее распространены на торфяных субстратах, которые они часто затягивают сплошными плотными коврами беловатого цвета с различными оттенками: буроватым, зеленоватым, розоватым. Этот ковер состоит из сближенных, сцепленных своими веточками стебельков многих экземпляров растений. Выделенное из дернины растеньице сфагнового мха состоит из прямого тонкого олиственного стебелька, лишенного корневых волосков и ризоидов и несущего вверху растрепанную головку из кучно сближенных на стебле веточек. В процессе роста стебля вверх расстояние между веточками по стеблю увеличивается, веточки разветвляются у основания и образуют пучки ветвей, расположенных по стеблю спирально, всегда рядом с каждым четвертым стеблевым листом.
Каждый пучок состоит из 2—6, иногда до 13, веточек двух видов: торчащих и свисающих. Торчащие, более мощные веточки отстоят от стебля в различных направлениях и способствуют сцеплению отдельных стебельков в дернине. Более слабые, свисающие прилегают к стеблю и служат для проведения воды вверх по стеблю. Веточки сплошь покрыты черепитчаторасположенными листочками, лучше развитыми и гуще растущими на торчащих веточках и более слабыми, бледными и реже растущими — на свисающих.
Стебелек сфагнума состоит из тканей трех видов: коры, центрального цилиндра и сердцевины. Кора стебля (гиалодермис) состоит из одного или нескольких слоев бесцветных пустых тонкостенных (с широкими просветами в плане) клеток, в оболочках которых у отдельных видов можно наблюдать округлые отверстия (поры), а у мхов секции Sphagnum, кроме того, нежные спиральные утолщения. Центральный цилиндр образован толстостенными (с узким просветом в плане) продольно вытянутыми прозенхимными клетками, стенки которых окрашены в розовый, желтый или бурый цвет. Сердцевина стебля состоит из тонкостенных бесцветных паренхимных клеток. Строение веточек такое же, как стебля, но кора их однослойна, реже двухслойна.
Сердцевина стебля состоит из тонкостенных бесцветных паренхимных клеток. Строение веточек такое же, как стебля, но кора их однослойна, реже двухслойна.
Веточные листья у разных видов сфагновых мхов по форме различны: от ланцетной до яйцевидно-округлой, и отличаются от стеблевых листьев, имеющих большей частью также различную форму, постоянную и характерную для каждого вида: от треугольной до языковидной. Веточные и стеблевые листья однослойны, лишены жилки и состоят из двух видов клеток: живых хлорофиллоносных, ассимилирующих углерод, и мертвых гиалиновых, лишенных плазматического содержимого и хлорофилла.
Хлорофиллоносные клетки имеют узкую удлиненную, несколько извилистую форму и, соприкасаясь своими концами, образуют сетку, ячейки которой заполнены бесцветными гиалиновыми широкими клетками, червеобразной или ромбической: формы. Спиральные и кольчатые утолщения в тонких стенках гиалиновых клеток позволяют им сохранить форму при высыхании. Почти всегда оболочки гиалиновых клеток листьев обнаруживают отверстия (поры) различной формы и расположения.
Род Sphagnum подразделяется на секции, которые отличаются по внешнему виду объединяемых ими мхов, форме стеблевых и веточных листьев и анатомическому строению: форме и положению хлорофиллоносных клеток на поперечном срезе листа и расположению и форме пор в стенках гиалиновых клеток. Каждая секция объединяет ряд более или менее близких между собой видов. Главнейшие секции — это Sphagnum, Squarrosa, Subsecunda, Cuspidata и Acuiifolia. Мхи этих секции отличаются следующими систематическими признаками.
Секция Sphagnum — веточные листья вогнутые, тупые, округло-яйцевидные. Гиалиновые клетки короткие и широкие. Поры круглые (по две-три), располагаются в углах гиалиновых клеток на наружной стороне листа.
Секция Squarrosa — веточные листья яйцевидно-ланцетной формы, иногда с отогнутой назад верхушкой. Крупные округлые поры в большом числе с обеих сторон листа.
Секция Subsecunda — веточные листья округло-овальные или продолговато-яйцевидные. Мелкие поры в большом числе располагаются в виде цепочек вдоль комиссур по наружной стороне листа.
Секция Cuspidata — веточные листья ланцетные или ланцетно-линейные. Гиалиновые клетки узкие и длинные.
Секция Acutifolia — веточные листья мелкие ланцетные или овально-ланцетные. Комиссуральные поры гиалиновых клеток на наружной стороне листа имеют округлую или полуэллиптическую форму.
В отдельные секции входят следующие главнейшие виды: секция Sphagnum — Sph. magellanicum (Sph. medium), Sph. centrale, Sph. papillosum, Sph. palustre, Sph. imbricatum; секция Squarrosa — Sph. squarrosum, Sph. teres; секция Subsecunda — Sph. subsecundum, Sph. contortum, Sph. platyphyllum, Sph. orientale; секция Cuspidata — Sph. lindbergii, Sph. obtusum, Sph. majus (Sph, dusenii), Sph. balticum, Sph. fallax (Sph. apiculatum), Sph. angusti folium, Sph. cuspidatum, Sph. lenense, Sph. riparium; секция Acutifolia — Sph. girgensohnii, Sph. fuscum, Sph. rubellum, Sph. warnstorfii, Sph. nemoreum (Sph. acutifolium), Sph. russowii.
Сфагновые мхи наиболее распространены на торфяных субстратах с низким содержанием минеральных веществ. Малая потребность в минеральном питании дает им преимущество в борьбе с более требовательными к минеральному режиму растениями и обеспечивает им господство в растительном покрове.
Малая потребность в минеральном питании дает им преимущество в борьбе с более требовательными к минеральному режиму растениями и обеспечивает им господство в растительном покрове.
Засасывая воду через поры гиалиновых клеток своих листьев и коры стебля, они в состоянии поглотить ее примерно в 20 раз больше собственной массы в воздушно-сухом состоянии. Произрастание сплошными коврами, состоящими из сближенных стебельков многих экземпляров растений, позволяет сфагновым мхам свести испарение воды к минимуму и удерживать ее на долгое время. При подсыхании гиалиновые клетки отдают воду и остаются пустыми, но растение не теряет из-за этого своей жизнеспособности.
Различные виды сфагновых мхов расселяются в разных условиях местообитания. Такие виды, как Sph. fuscum, Sph. magellanicum, Sph. rubellum, Sph. angustifolium, Sph. balticum, Sph. majus, Sph. cuspidatum, в массовых количествах заселяют верховые болота, питаемые в основном бедными атмосферными водами. Другие виды сфагновых мхов произрастают в условиях более богатого минерального питания — на переходных и низинных болотах.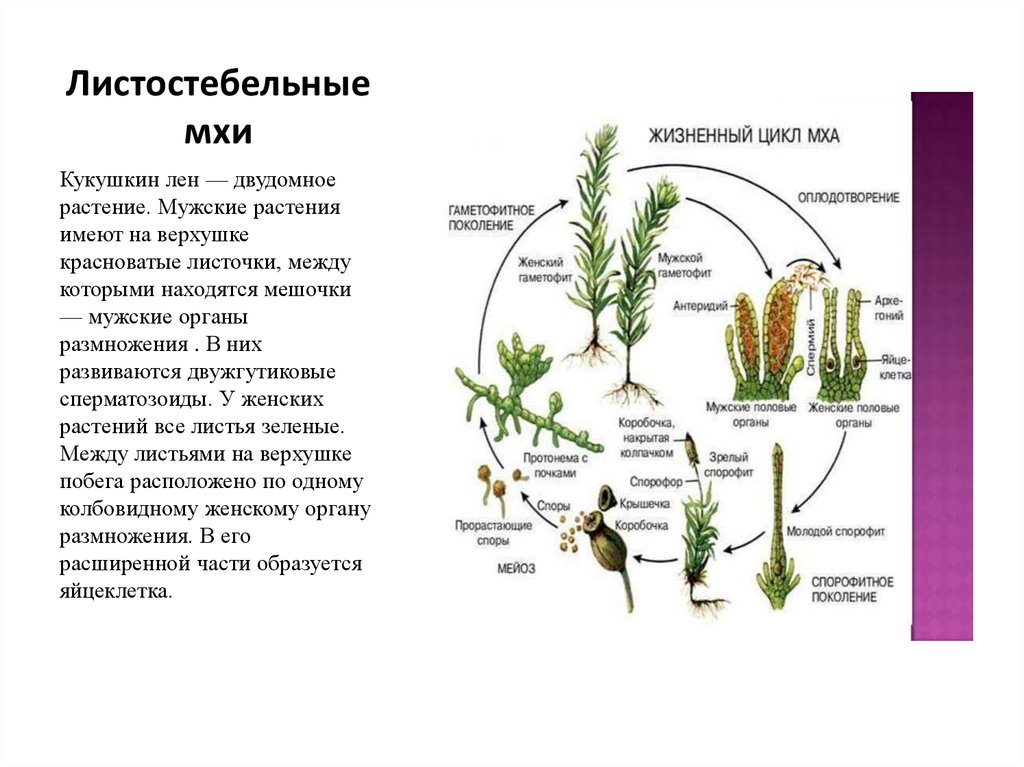 Сюда относятся: Sph. warnstorfii, Sph. teres, Sph. squarrosum, Sph. obtusum, Sph. subsecundum, Sph. contortum, Sph. centrale, Sph. papillosum.
Сюда относятся: Sph. warnstorfii, Sph. teres, Sph. squarrosum, Sph. obtusum, Sph. subsecundum, Sph. contortum, Sph. centrale, Sph. papillosum.
Требовательность различных видов сфагновых мхов к влажности среды также неодинакова: Sph. cuspidatum, Sph. majus растут погруженными в воду; Sph. angustifolium, Sph. obtusum и Sph. subsecundum, произрастая обычно на сильно обводненных участках, встречаются иногда и в условиях пониженной влажности; Sph. warnstorfii, Sph. centrale, Sph. magellanicum, Sph. fuscum, Sph. palustre обычно значительно приподняты над уровнем воды, образуя кочки и бугры; Sph. squarrosum, Sph. nemoreum, Sph. girgensohnii заселяют преимущественно слабо увлажненные участки на лесных заболачивающихся минеральных почвах.
Для местообитании сфагновых мхов, особенно верхового типа, характерна повышенная кислотность; они обладают способностью сами подкислять среду.
Сфагновые мхи верхового типа не переносят даже незначительного количества углекислого кальция — Sph.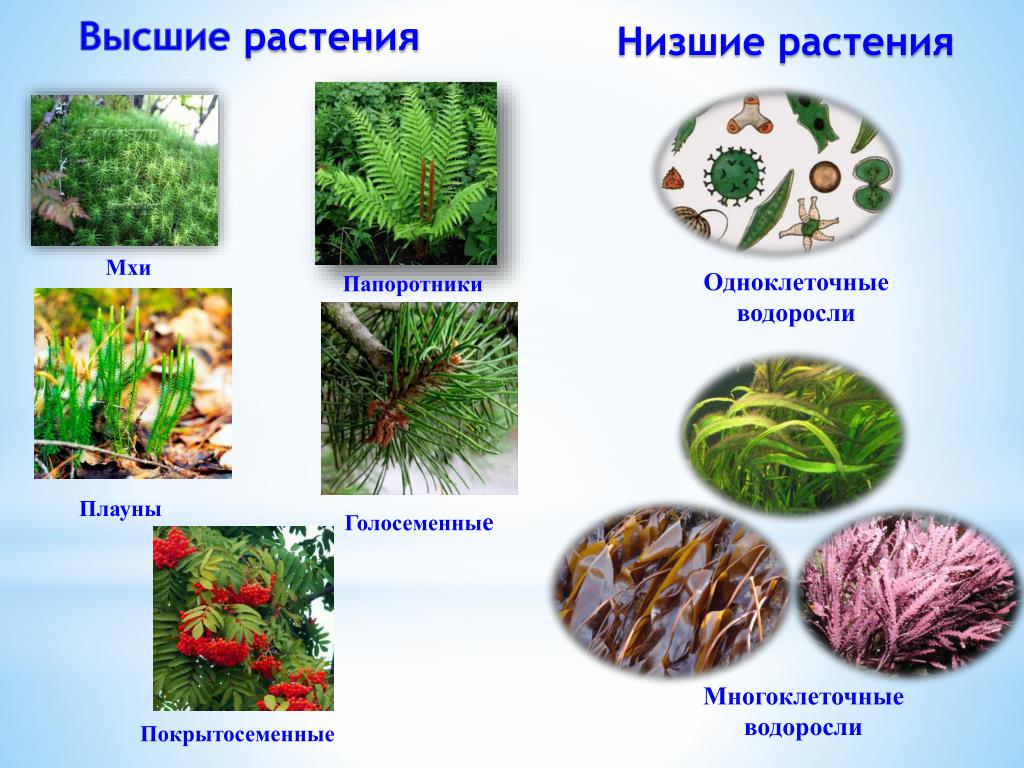 rubellum отмирает при содержаний в воде 77 мг углекислого кальция на 1 л, Sph. magellanicum — при 134 мг.
rubellum отмирает при содержаний в воде 77 мг углекислого кальция на 1 л, Sph. magellanicum — при 134 мг.
Мхи (тип Bryophyta) – мохообразные
Классификация и идентификация
Отправной точкой является схема классификации мхов, изложенная в документе, приведенном в следующей кнопке «Ссылка». Эта классификация основана на сочетании молекулярных, морфологических (макроскопических и микроскопических) данных и данных о клеточной структуре и для краткости будет называться на этой веб-странице классификацией 2000 . Он обеспечивает хорошую основу для построения краткого изложения принципов классификации мхов и сопоставления некоторых результатов более поздних исследований.
В схеме 2000 Классификации Phylum Bryophyta разделен на шесть классов: Takakiopsida , Sphagnopsida , Andreaeopsida , Andreaeopryopsida , Polytrichopsida и Breaopsida , Polytrichopsida и Breaopsida , . Ряд бриологов утверждают, что Takakiopsida и Sphagnopsida настолько непохожи на другие мхи, что эти два класса фактически должны составлять отдельные типы. Те, кто придерживается этой точки зрения, будут иметь два дополнительных типа, Takakiophyta и Sphagnophyta , с типом Bryophyta, уменьшенным с шести до четырех классов. Обратите внимание, что все согласны с тем, что группы «Takakia» и «Sphagnum» заметно отличаются от всех других мохообразных. Споры ведутся о том, требуют ли эти две группы разделения. В этих дебатах заложены два разных взгляда на эволюционную историю.
Ряд бриологов утверждают, что Takakiopsida и Sphagnopsida настолько непохожи на другие мхи, что эти два класса фактически должны составлять отдельные типы. Те, кто придерживается этой точки зрения, будут иметь два дополнительных типа, Takakiophyta и Sphagnophyta , с типом Bryophyta, уменьшенным с шести до четырех классов. Обратите внимание, что все согласны с тем, что группы «Takakia» и «Sphagnum» заметно отличаются от всех других мохообразных. Споры ведутся о том, требуют ли эти две группы разделения. В этих дебатах заложены два разных взгляда на эволюционную историю.
Классы в классификации 2000 года сильно различаются по размеру. Около 95 % всех видов мхов относятся к классу Bryopsida, тогда как Takakiopsida содержит только один род, Takakia всего с двумя видами. Неудивительно, что большинство мхов, попадающихся на глаза людям, относятся к классу моховидных. Внутри класса Polytrichopsida члены класса Polytrichales обычно имеют очень прочные гаметофиты, которые легко заметить. Хотя Polytrichales намного меньше по численности, чем Bryopsida, они широко распространены. Следующие два раздела приведут несколько примеров Bryopsida и Polytrichales, показывая вам некоторое разнообразие в каждой группе, а также некоторые классификационные признаки, которые можно найти в этих группах. Страница заканчивается кратким изложением и некоторыми комментариями обо всех классах, составляющих тип Bryophyta.
Хотя Polytrichales намного меньше по численности, чем Bryopsida, они широко распространены. Следующие два раздела приведут несколько примеров Bryopsida и Polytrichales, показывая вам некоторое разнообразие в каждой группе, а также некоторые классификационные признаки, которые можно найти в этих группах. Страница заканчивается кратким изложением и некоторыми комментариями обо всех классах, составляющих тип Bryophyta.
Класс Bryopsida
Goniomitrium acuminatum |
Учитывая большой размер этого класса, неудивительно, что существует значительное разнообразие как гаметофитов, так и спорофитов его членов.
Гаметофиты Goniomitrium acuminatum (Pottiaceae, Pottiales) (справа) вырастают всего до нескольких миллиметров в высоту, а сферические капсулы со спорами на коротких ножках имеют диаметр всего 1-2 миллиметра. На сопроводительном фото преобладают незрелые спорофиты. В отличие от видов Papillaria (Meteoriaceae, Hypnales) может вырастать в виде куртин и достигать более метра в длину.
В отличие от видов Papillaria (Meteoriaceae, Hypnales) может вырастать в виде куртин и достигать более метра в длину.
Гаметофиты мхов семейства Pottiaceae обычно представляют собой короткостебельные, опушенные растения, часто растущие стадно и в суровых, незащищенных местообитаниях. Примерами, кроме Goniomitrium , являются Tortula Princeps и Trichostomum brachydontium . Это не означает, что все короткостебельные, пушистые мхи в суровых условиях обитания являются представителями Pottiaceae. Gigaspermum repens (Gigaspermaceae, Funariales) является хорошим контрпримером. Также не факт, что все представители Pottiaceae имеют короткие пучковые гаметофиты. Стебли Triquetrella papillata могут вырастать до нескольких сантиметров в длину и разветвляться, так что гаметофиты растут в виде ковриков на земле. Однако длинные стебли в матообразном росте гораздо более характерны для порядков Hookeriales и особенно Hypnales. Последний представляет собой крупный отряд, насчитывающий несколько тысяч видов. Название отряда происходит от рода Hypnum (из семейства Hypnaceae) и здесь небольшой, но плотный мат Hypnum cupressiforme . На этой фотографии показан рыхлый коврик из Brachythecium salebrosum (Brachytheciaceae, Hypnales), растущий на пышной лужайке у Старого здания парламента в Канберре, а здесь хорошо развитый коврик из Thuidium cymbifolium (Thuidiaceae, Hypnales), растущий на придорожной насыпи. на острове Норфолк. На примере Papillaria вы уже видели, что члены Hypnales могут расти не только как мат, но и как занавески.
Название отряда происходит от рода Hypnum (из семейства Hypnaceae) и здесь небольшой, но плотный мат Hypnum cupressiforme . На этой фотографии показан рыхлый коврик из Brachythecium salebrosum (Brachytheciaceae, Hypnales), растущий на пышной лужайке у Старого здания парламента в Канберре, а здесь хорошо развитый коврик из Thuidium cymbifolium (Thuidiaceae, Hypnales), растущий на придорожной насыпи. на острове Норфолк. На примере Papillaria вы уже видели, что члены Hypnales могут расти не только как мат, но и как занавески.
У каждого из Gemmabryum dichotomum (Bryaceae, Bryales) и Tayloria gunnii (Splachnaceae, Splachnales) устье споровой капсулы окружено перистомными зубцами , которые могут способствовать РАСПРОСТРАНЕНИЮ спор. Большинство представителей Bryopsida имеют споровые капсулы с перистомными зубцами. У Gemmabryum dichotomum зубцы хорошо развиты, а у Tayloria gunnii они довольно мелкие. Напротив, споровая капсула Physcomitrium pyriforme (Funariaceae, Funariales) перистомные зубы отсутствуют. Классификация 2000 года использует характер (или отсутствие) перистомных зубов, чтобы помочь определить отряды мхов. Одна из капсул со спорами Gemmabryum dichotomum все еще находится в процессе сбрасывания крышки , крышки, закрывающей устье капсулы в развивающихся капсулах со спорами. Вы можете видеть кольцо , соединительное кольцо ткани между жаберной крышкой и капсулой, в процессе сбрасывания. 9Классификация 0008 2000 явно использует наличие хорошо дифференцированного кольца, чтобы помочь определить некоторые отряды мхов.
Напротив, споровая капсула Physcomitrium pyriforme (Funariaceae, Funariales) перистомные зубы отсутствуют. Классификация 2000 года использует характер (или отсутствие) перистомных зубов, чтобы помочь определить отряды мхов. Одна из капсул со спорами Gemmabryum dichotomum все еще находится в процессе сбрасывания крышки , крышки, закрывающей устье капсулы в развивающихся капсулах со спорами. Вы можете видеть кольцо , соединительное кольцо ткани между жаберной крышкой и капсулой, в процессе сбрасывания. 9Классификация 0008 2000 явно использует наличие хорошо дифференцированного кольца, чтобы помочь определить некоторые отряды мхов.
Класс Polytrichopsida, отряд Polytrichales
Как отмечалось выше, гаметофиты представителей рода Polytrichales, как правило, довольно крепкие. Polytrichaceae — единственное семейство в этом отряде. Стебли гаметофитов могут быть от нескольких сантиметров в высоту до более 60 сантиметров у некоторых видов. Стебли некоторых водных мхов могут быть довольно длинными, но Polytrichales — это наземные мхи с отдельно стоящими стеблями. Вот фото нескольких растений рода Доусония . Вот масса из растений Dawsonia , с человеком для масштаба. Есть некоторые крепкие мхи, которых нет в Polytrichales. Одним из таких является тропический род Spiridens (Spiridentaceae, Rhizogoniales, Bryopsida), стебли которого могут достигать более 30 сантиметров в длину. Однако стебли Spiridens растут от горизонтального до слегка висячего, в отличие от вертикального роста у Polytrichales. Spiridens иногда растет на почве, но гораздо чаще встречается на растениях.
Стебли некоторых водных мхов могут быть довольно длинными, но Polytrichales — это наземные мхи с отдельно стоящими стеблями. Вот фото нескольких растений рода Доусония . Вот масса из растений Dawsonia , с человеком для масштаба. Есть некоторые крепкие мхи, которых нет в Polytrichales. Одним из таких является тропический род Spiridens (Spiridentaceae, Rhizogoniales, Bryopsida), стебли которого могут достигать более 30 сантиметров в длину. Однако стебли Spiridens растут от горизонтального до слегка висячего, в отличие от вертикального роста у Polytrichales. Spiridens иногда растет на почве, но гораздо чаще встречается на растениях.
Нервы присутствуют в листьях Polytrichales и, как правило, вдоль длинного нерва проходят стенообразные наросты, называемые пластинками . На прилагаемой диаграмме показано поперечное сечение листа вида Aloina Rigida . Центральная коричневая часть, толщиной в несколько клеток, представляет собой нерв. С каждой стороны от нерва листок имеет толщину всего в одну клетку и изогнут внутрь. Каждая зеленая «столбик» обозначает одну пластину. Помните, что это поперечное сечение, так что каждая зеленая «столбик» продолжается назад в экран и вперед из экрана, образуя пластину, подобную стене. Среди Polytrichales есть различия в таких особенностях, как форма листьев, курчавость листьев, форма и расположение ламелей. Они помогают различать виды. Диаграмма взята из Тафеля 16 в книге В. Мигулы 9.0008 Kryptogamen-Flora von Deutschland, Deutsch- Österreich und der Schweiz: Band 1: Moose , опубликовано в Берлине в 1904 году.
С каждой стороны от нерва листок имеет толщину всего в одну клетку и изогнут внутрь. Каждая зеленая «столбик» обозначает одну пластину. Помните, что это поперечное сечение, так что каждая зеленая «столбик» продолжается назад в экран и вперед из экрана, образуя пластину, подобную стене. Среди Polytrichales есть различия в таких особенностях, как форма листьев, курчавость листьев, форма и расположение ламелей. Они помогают различать виды. Диаграмма взята из Тафеля 16 в книге В. Мигулы 9.0008 Kryptogamen-Flora von Deutschland, Deutsch- Österreich und der Schweiz: Band 1: Moose , опубликовано в Берлине в 1904 году.
Споровые капсулы Polytrichales имеют перистомные зубцы, но не всегда простые. У некоторых родов, таких как Polytrichum , верхушки перистомных зубов прикреплены к белой дисковидной эпифрагме . У Dawsonia зубцы перистома длинные и несколько волосовидные.
Polytrichum juniperinum капсулы с эпифрагмами | Доусония sp. |
 капсула с волосовидными перистомными зубцами
капсула с волосовидными перистомными зубцамиКраткое изложение классификации мхов
В следующей таблице приведены краткие сведения о классах мхов согласно классификации 2000 года . После названия каждого класса серый текст дает некоторую информацию о характеристиках членов этого класса. Серый текст никогда не является полным описанием класса, но для первых четырех классов вы можете увидеть некоторые легко описываемые различия внутри этой группы из четырех и между этими четырьмя и оставшимися двумя. Для каждого класса (кроме мохообразных, у которых слишком много отрядов) есть также список отрядов и сводка номеров семейств, родов и видов. Более подробное обсуждение классификации мхов см. на странице мхов на веб-сайте «Древо жизни».
тип:Мохообразные класс: Такакиопсида отряд: Takakiales (1 семейство, 1 род, 2 вида) класс: Sphagnopsida отряд: Sphagnales (1 семейство, 1 род, от 100 до 300 видов) отряд: Ambuchananiales (1 семейство, 1 род, 1 вид) класс: Andreaeopsida отряд: Andreaeales (1 семейство, 2 рода, до 100 видов) класс: Andreaeobryopsida отряд: Andreaeobryales (1 семейство, 1 род, 1 вид) класс: Polytrichopsida отряд: Tetraphidales (3 семейства, 4 рода, не более примерно 50 видов) отряд: Polytrichales (1 семейство, 23 рода, несколько сотен видов) класс: Bryopsida 16 отрядов, 107 семейств, около 880 родов, около 12 000 видов
|



Нематодонты и артродонты
артродонтальный перистом |
Перистомальные зубы нематодонтов состоят из цельных мертвых клеток и равномерно утолщены, тогда как перистомальные зубы артродонтов дифференциально утолщены и образованы из частей мертвых клеток,
Большинство мхов имеют членистоногие зубы перистома, и пример членистоногого перистома показан на прилагаемой фотографии. Вот крупный план одного зуба. Артродонтические перистомированные зубы обладают значительной гибкостью и могут скручиваться или раскручиваться в ответ на изменения влажности.
Вот крупный план одного зуба. Артродонтические перистомированные зубы обладают значительной гибкостью и могут скручиваться или раскручиваться в ответ на изменения влажности.
Предлагаемая редакция
Основываясь на обзоре морфологических данных и данных ДНК, авторы статьи, приведенной в следующей ссылке, предложили пересмотренную классификацию мхов. Большинство предлагаемых изменений относятся к уровню порядка или ниже в классе Bryopsida. Однако они предложили следующие изменения в классе Polytrichopsida. Семейство Buxbaumiacae (только с одним родом Buxbaumia ) будет помещен в отдельный отряд Buxbaumiales в классе Bryopsida. В классификации -2000 это семейство относилось к отряду Tetraphidales. Каждая из оставшихся двух семей Tetraphidales образует новый класс, создавая таким образом:
Класс
: Эдиподиопсида
Класс
: Tetraphidopsida
Это оставило бы класс Polytrichopsida только с одним порядком Polytrichales и одним семейством Polytrichaceae. В 2000 классификация класс Polytrichopsida содержал четыре семейства с устьями споровых капсул, обладающих следующими характеристиками:
В 2000 классификация класс Polytrichopsida содержал четыре семейства с устьями споровых капсул, обладающих следующими характеристиками:
Oedipodiaceae Перистомальные зубцы отсутствуют, устье капсулы гладкое
Tetraphidaceae Четыре передних зуба, расположенные по одной круговой линии
Polytrichaceae 16-64 перистомных зуба, расположенных по одной круговой линии
Buxbaumiaceae Один или несколько рядов зубцов, концентрических вокруг усеченного складчатого конуса
В пересмотренной классификации этот смешанный пакет был разделен на более однородные группы.
Структура устья капсулы Buxbaumiaceae является одной из самых сложных среди мхов, а виды семейства Diphysciaceae имеют подобные устья капсулы. Интересно, что в классификации 2000 семейство Diphysciaceae было помещено в порядок Diphysciales внутри класса Bryopsida. Это размещение было основано на тщательном изучении как тонкой структуры перистома, так и молекулярных данных. В прошлом веке велись серьезные споры о классификации Buxbaumiaceae и Diphysciaceae, при этом споры были сосредоточены на том, является ли сложный перистом более нематодонтным, более артродонтным или промежуточной формой. Обратите внимание, что в пересмотренной классификации, хотя Buxbaumiaceae и Diphysciaceae помещены в класс Bryopsida, они помещены в разные подклассы.
Это размещение было основано на тщательном изучении как тонкой структуры перистома, так и молекулярных данных. В прошлом веке велись серьезные споры о классификации Buxbaumiaceae и Diphysciaceae, при этом споры были сосредоточены на том, является ли сложный перистом более нематодонтным, более артродонтным или промежуточной формой. Обратите внимание, что в пересмотренной классификации, хотя Buxbaumiaceae и Diphysciaceae помещены в класс Bryopsida, они помещены в разные подклассы.
Авторы пересмотренной классификации также предложили пять надклассов , на данный момент пронумерованных римскими цифрами и определенных следующим образом:
Надкласс I (содержащий класс Takakiopsida)
Надкласс II (содержащий класс Sphagnopsida)
Надкласс III (содержащий класс Andreaeopsida)
Надкласс IV (содержащий класс Andreaeobryopsida
Суперкласс V
Класс
: Эдиподиопсида
Класс
: Политрихопсида
Класс
: Tetraphidospida
класс
: моховидные
Это ясно показывает, что, по мнению авторов, классы Oedipodiopsida, Polytrichopsida, Tetraphidospida и Bryopsida более тесно связаны друг с другом, чем любой из них с любым из Takakiopsida, Sphagnopsida, Andreaeopsida или Andreaeobryopsida.
Мох / Мхи — Biocyclopedia.com
Мхи — это небольшие мягкие растения, обычно 1–10 см (0,4–4 дюйма) в высоту, хотя некоторые виды намного крупнее. Обычно они растут близко друг к другу, образуя скопления или коврики во влажных или тенистых местах. У них нет цветов или семян, а их простые листья покрывают тонкие проволочные стебли. В определенное время мхи производят капсулы со спорами, которые могут выглядеть как клювовидные капсулы, поднимающиеся вверх на тонких стеблях.
Около 12 000 видов мхов относятся к мохообразным. В отдел Bryophyta раньше входили не только мхи, но также печеночники и роголистники. Эти две другие группы мохообразных теперь часто помещают в отдельные отделы.
Содержимое
» Физические характеристики
» Описание
» Жизненный цикл
» Классификация
» Геологическая история
» Среда обитания
» Культивирование
» Ингибирование роста мха
» Mossery
» Коммерческое использование
» Ссылки
Описание
«Muscinae» из книги Эрнста Геккеля Kunstformen der Natur , 1904
С ботанической точки зрения мхи являются мохообразными или несосудистыми растениями. Их можно отличить от внешне похожих печеночников (Marchantiophyta или Hepaticae) по их многоклеточным ризоидам. Другие отличия не являются универсальными для всех мхов и всех печеночников, но наличие четко дифференцированных «стебля» и «листьев», отсутствие глубоко лопастных или сегментированных листьев, а также отсутствие листьев, расположенных в три ряда, все указывает на растение. будучи мхом.
Их можно отличить от внешне похожих печеночников (Marchantiophyta или Hepaticae) по их многоклеточным ризоидам. Другие отличия не являются универсальными для всех мхов и всех печеночников, но наличие четко дифференцированных «стебля» и «листьев», отсутствие глубоко лопастных или сегментированных листьев, а также отсутствие листьев, расположенных в три ряда, все указывает на растение. будучи мхом.
В дополнение к отсутствию сосудистой системы, мхи имеют жизненный цикл с преобладанием гаметофитов, т. е. клетки растения являются гаплоидными на протяжении большей части его жизненного цикла. Спорофиты (то есть диплоидное тело) недолговечны и зависят от гаметофита. Это контрастирует с тем, что демонстрируют большинство «высших» растений и большинство животных. У семенных растений, например, гаплоидное поколение представлено пыльцой и семязачатком, а диплоидное поколение — привычное цветковое растение.
Жизненный цикл
Большинство видов растений, за исключением водорослей и мохообразных, имеют два набора хромосом в своих вегетативных клетках и называются диплоидными, т. е. у каждой хромосомы есть партнер, который содержит одинаковую или подобную генетическую информацию. Мхи (и другие мохообразные) имеют только один набор хромосом (гаплоидный, т. е. каждая хромосома существует в единственной копии внутри клетки). В жизненном цикле мхов бывают периоды, когда они имеют полный двойной набор парных хромосом, но только на стадии спорофита.
е. у каждой хромосомы есть партнер, который содержит одинаковую или подобную генетическую информацию. Мхи (и другие мохообразные) имеют только один набор хромосом (гаплоидный, т. е. каждая хромосома существует в единственной копии внутри клетки). В жизненном цикле мхов бывают периоды, когда они имеют полный двойной набор парных хромосом, но только на стадии спорофита.
Жизненный цикл типичного мха
Жизнь мха начинается с гаплоидной споры, которая прорастает с образованием протонемы, которая представляет собой либо массу нитей, либо таллоид (плоский и похожий на слоевище). Это переходный этап в жизни мха. Из протонемы вырастает гаметофор («носитель гамет»), который дифференцируется на стебли и листья. Из кончиков стеблей или ветвей развиваются половые органы мхов. Женские органы известны как архегонии (sing. archegonium) и защищены группой модифицированных листьев, известных как перихетум (множественное число, перихеты). У архегоний есть шейки, называемые вентерами, по которым спускаются мужские сперматозоиды. Мужские органы известны как антеридии (единственное число антеридий) и окружены видоизмененными листьями, называемыми перигониями (множественное число, перигония).
Мужские органы известны как антеридии (единственное число антеридий) и окружены видоизмененными листьями, называемыми перигониями (множественное число, перигония).
Мхи могут быть либо двудомными (сравните двудомные у семенных растений), либо однодомными (сравните однодомные). У двудомных мхов как мужские, так и женские половые органы находятся на разных растениях-гаметофитах. У однодольных (также называемых аутоидными) мхов они переносятся на одном и том же растении. В присутствии воды сперматозоиды из антеридий переплывают к архегониям, и происходит оплодотворение, приводящее к образованию диплоидного спорофита. Сперматозоиды мхов двужгутиковые, то есть у них есть два жгутика, которые способствуют движению. Поскольку сперматозоиды должны доплыть до архегония, оплодотворение не может происходить без воды. После оплодотворения незрелый спорофит выталкивается из вентральной части архегониума. Для созревания спорофита требуется от четверти до полугода.
Тело спорофита состоит из длинной ножки, называемой щетинкой, и капсулы, увенчанной крышечкой, называемой крышечкой. Капсула и жаберная крышка, в свою очередь, покрыты гаплоидной калиптрой, которая представляет собой остатки архегониального вентрального отдела. Калиптра обычно отпадает, когда капсула созреет. Внутри капсулы клетки, производящие споры, подвергаются мейозу с образованием гаплоидных спор, после чего цикл может начаться снова. Устье капсулы обычно окружено зубцами, называемыми перистомом. Это может отсутствовать у некоторых мхов.
Капсула и жаберная крышка, в свою очередь, покрыты гаплоидной калиптрой, которая представляет собой остатки архегониального вентрального отдела. Калиптра обычно отпадает, когда капсула созреет. Внутри капсулы клетки, производящие споры, подвергаются мейозу с образованием гаплоидных спор, после чего цикл может начаться снова. Устье капсулы обычно окружено зубцами, называемыми перистомом. Это может отсутствовать у некоторых мхов.
В некоторых мхах, напр. Ulota phyllantha , зеленые вегетативные структуры, называемые геммами, образуются на листьях или ветвях, которые могут отрываться и образовывать новые растения без необходимости проходить цикл оплодотворения. Это способ бесполого размножения, и генетически идентичные единицы могут привести к образованию клональных популяций.
Классификация
Мхи традиционно объединяли с печеночниками и роголистниками в отдел Bryophyta (мохообразные), внутри которого мхи составляли класс Musci. Эта группа, однако, является парафилетической и в настоящее время имеет тенденцию к разделению. В такой системе отдел Bryophyta относится именно к мхам. Они, по-видимому, являются ближайшими живыми родственниками сосудистых растений.
В такой системе отдел Bryophyta относится именно к мхам. Они, по-видимому, являются ближайшими живыми родственниками сосудистых растений.
The mosses are grouped as a single division, now named Bryophyta, and divided into six classes:
» Takakiopsida
» Sphagnopsida
» Andreaeopsida
» Andreaeobryopsida
» Polytrichopsida
» Bryopsida
Ствол дерева окружен двумя разными видами мхов (и лишайником в самой маленькой коробке).
Мох в Национальном лесу Аллегейни, Пенсильвания, США.
Andreaeopsida и Andreaeobryopsida отличаются двурядными (два ряда клеток) ризоидами, многорядной (много рядов клеток) протонемой и спорангием, расщепляющимся вдоль продольных линий. У большинства мхов есть капсулы, которые открываются вверху.
Sphagnopsida, торфяные мхи, включают два живых рода Ambuchanania и Sphagnum, а также ископаемые таксоны.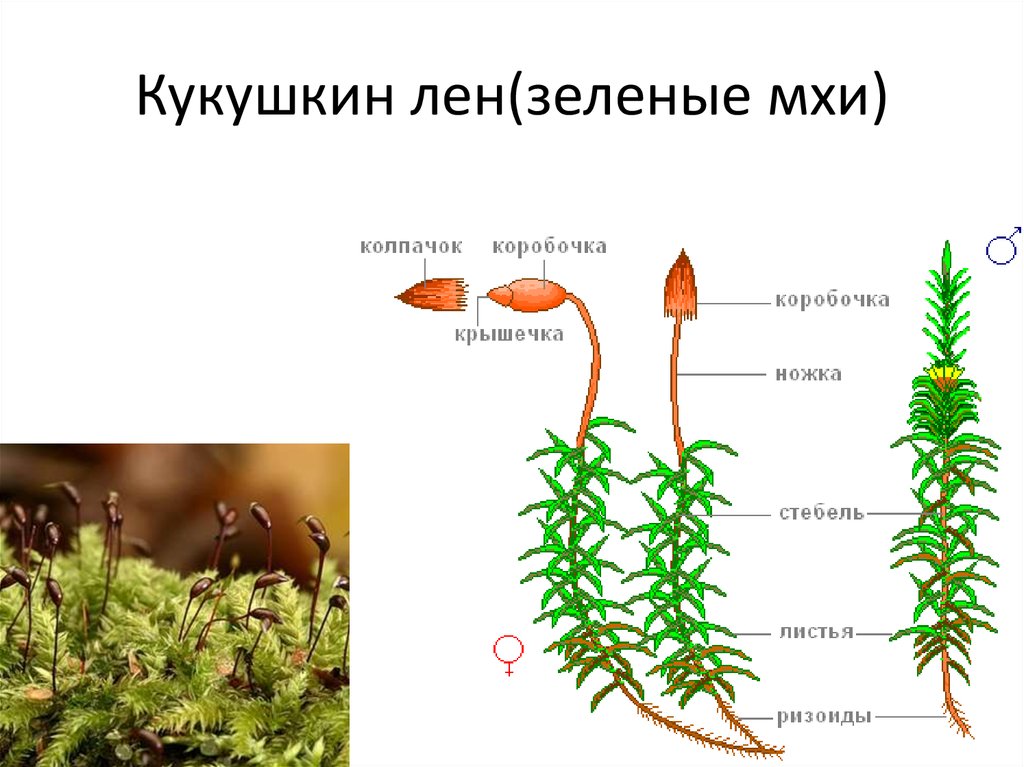 Эти крупные мхи образуют на торфяных болотах обширные кислые болота. Листья сфагнума имеют крупные мертвые клетки, чередующиеся с живыми фотосинтетическими клетками. Мертвые клетки помогают запасать воду. Помимо этого признака, уникальное ветвление, таллозная (плоская и расширенная) протонема и взрывно разрывающийся спорангий отличают его от других мхов.
Эти крупные мхи образуют на торфяных болотах обширные кислые болота. Листья сфагнума имеют крупные мертвые клетки, чередующиеся с живыми фотосинтетическими клетками. Мертвые клетки помогают запасать воду. Помимо этого признака, уникальное ветвление, таллозная (плоская и расширенная) протонема и взрывно разрывающийся спорангий отличают его от других мхов.
Polytrichopsida имеют листья с наборами параллельных ламелей, лоскутами клеток, содержащих хлоропласты, которые выглядят как плавники на радиаторе. Они осуществляют фотосинтез и могут способствовать сохранению влаги, частично закрывая поверхности газообмена. Polytrichopsida также отличаются от других мхов другими деталями своего развития и анатомии, а также могут становиться больше, чем большинство других мхов, например, с Polytrichum commune образует подушки высотой до 40 см (16 дюймов). Самый высокий наземный мох, член Polytrichidae, вероятно, Dawsonia superba, произрастающий в Новой Зеландии и других частях Австралазии.
Мохообразные — самая разнообразная группа; более 95% видов мхов относятся к этому классу.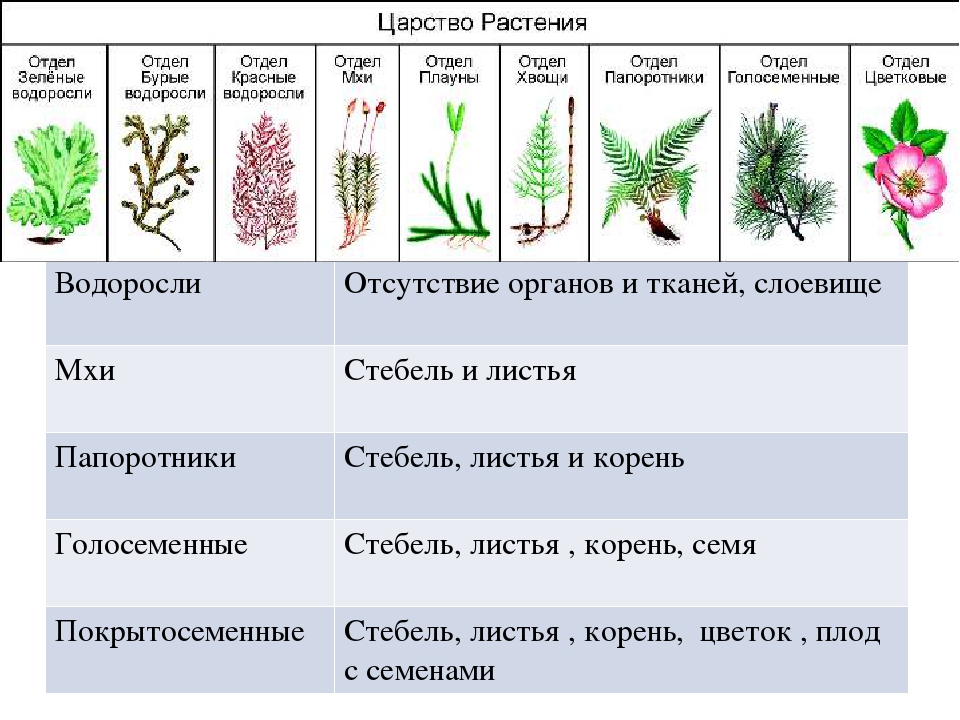
Archidiidae отличаются чрезвычайно крупными спорами и способом развития спорангиев.
Геологическая история
Летопись окаменелостей мхов скудна из-за их мягких стенок и хрупкости. Однозначные окаменелости мхов были обнаружены еще в пермском периоде Антарктиды и России, и выдвигаются аргументы в пользу каменноугольных мхов. Кроме того, утверждалось, что трубчатые окаменелости силурийского периода представляют собой размокшие остатки моховых калиптр.
Капсулы красного мха, зимний уроженец вересковой пустоши Йоркшир-Дейлс.
Среда обитания
Мхи встречаются в основном в сырых и малоосвещенных местах. Мхи распространены в лесистых местностях и по краям ручьев. Также мхи встречаются в щелях между брусчаткой на сырых городских улицах. Некоторые виды приспособились к городским условиям и встречаются только в городах. Некоторые виды полностью водные, такие как Fontinalis antipyretica, а другие, такие как Sphagnum, населяют болота, болота и очень медленно движущиеся водотоки. Такие водные или полуводные мхи могут значительно превышать нормальный диапазон длины, наблюдаемый у наземных мхов. Например, отдельные растения длиной 20–30 см (8–12 дюймов) и более обычны для видов Sphagnum.
Такие водные или полуводные мхи могут значительно превышать нормальный диапазон длины, наблюдаемый у наземных мхов. Например, отдельные растения длиной 20–30 см (8–12 дюймов) и более обычны для видов Sphagnum.
Везде, где они встречаются, мхам для выживания требуется влага из-за небольшого размера и тонкости тканей, отсутствия кутикулы (воскового покрытия для предотвращения потери воды) и потребности в жидкой воде для полного оплодотворения. Некоторые мхи могут пережить высыхание, возвращаясь к жизни в течение нескольких часов после регидратации.
В северных широтах на северной стороне деревьев и скал обычно в среднем больше мха, чем на других сторонах (хотя выходы на южную сторону известны). Предполагается, что это связано с отсутствием достаточного количества воды для размножения на обращенной к солнцу стороне деревьев. К югу от экватора верно обратное. В густых лесах, куда не проникает солнечный свет, мхи одинаково хорошо растут со всех сторон ствола дерева.
Выращивание
Густые моховые колонии в прохладном прибрежном лесу
Крупный план мха на скале
Молодые спорофиты обыкновенного мха Tortula muralis (настенный винтовой мох)
Небольшой комок мха
Мох считается сорняком на травяных газонах, но его намеренно поощряют выращивать в соответствии с эстетическими принципами, примером которых является японское садоводство. В садах старых храмов мох может покрыть лесную сцену. Считается, что мох добавляет ощущение спокойствия, возраста и тишины в сцену в саду. Правила выращивания широко не установлены. Коллекции мхов нередко начинают с образцов, пересаженных из дикой природы в водоудерживающий мешок. Тем не менее, некоторые виды мхов может быть чрезвычайно трудно содержать вдали от их естественных местообитаний из-за их уникального сочетания света, влажности, укрытия от ветра и т. д.
Выращивание мха из спор еще менее контролируемо. Споры мха выпадают под постоянным дождем на открытые поверхности; те поверхности, которые подходят для определенных видов мха, обычно заселяются этим мхом в течение нескольких лет после воздействия ветра и дождя. Пористые и влагоудерживающие материалы, такие как кирпич, дерево и некоторые крупнозернистые бетонные смеси, благоприятны для мха. Поверхности также можно подготовить с помощью кислых веществ, включая пахту, йогурт, мочу и мягко протертые смеси образцов мха, воды и верескового компоста.
Пористые и влагоудерживающие материалы, такие как кирпич, дерево и некоторые крупнозернистые бетонные смеси, благоприятны для мха. Поверхности также можно подготовить с помощью кислых веществ, включая пахту, йогурт, мочу и мягко протертые смеси образцов мха, воды и верескового компоста.
Ингибирование роста мха
Рост мха можно затормозить несколькими способами:
» Уменьшение доступности воды за счет дренажа или прямого внесения изменений.
» Увеличение прямого солнечного света.
» Увеличение количества и ресурсов, доступных для конкурентоспособных растений, таких как травы.
» Повышение pH почвы с применением извести.
Интенсивное движение транспорта или ручное вскапывание моховой подстилки граблями также препятствует росту мха.
Применение продуктов, содержащих сульфат железа или сульфат железа и аммония, убивает мох, эти ингредиенты обычно входят в состав коммерческих продуктов для борьбы со мхом и удобрений.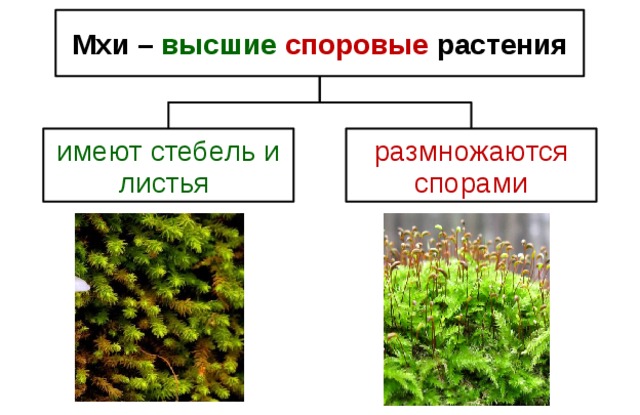 Сера и железо являются важными питательными веществами для некоторых конкурирующих растений, таких как травы. Уничтожение мхов не предотвратит повторный рост, если не будут изменены условия, благоприятные для их роста.
Сера и железо являются важными питательными веществами для некоторых конкурирующих растений, таких как травы. Уничтожение мхов не предотвратит повторный рост, если не будут изменены условия, благоприятные для их роста.
Моховой
Преходящая мода на сбор мха в конце 19 века привела к созданию мхов во многих британских и американских садах. Моховик обычно строится из реечного дерева с плоской крышей, открытой на северную сторону (для сохранения тени). Образцы мха устанавливали в щели между деревянными планками. Затем весь мох регулярно увлажняют для поддержания роста.
Коммерческое использование
Моховой фотобиореактор с Physcomitrella patens
Существует значительный рынок мхов, собранных в дикой природе. Использование неповрежденного мха в основном в цветочной торговле и для украшения дома. Разлагающийся мох рода Sphagnum также является основным компонентом торфа, который «добывается» для использования в качестве топлива, в качестве добавки к почве для садоводства и в курительном солоде при производстве шотландского виски.
Мох сфагнум, обычно виды cristatum и subnitens, собирают во время роста и сушат для использования в питомниках и садоводстве в качестве среды для выращивания растений. Заготовку мохового торфа не следует путать с заготовкой мохового торфа.
Торфяной мох можно заготавливать на устойчивой основе и управлять им так, чтобы был разрешен повторный рост, в то время как заготовка торфяного мха, как правило, считается наносящей значительный ущерб окружающей среде, поскольку торф удаляется с небольшим шансом на восстановление или вообще без него.
Во время Второй мировой войны сфагновые мхи использовались в качестве первой помощи при ранениях солдат, так как эти мхи хорошо впитывают влагу и обладают мягкими антибактериальными свойствами. Некоторые ранние люди использовали его в качестве подгузника из-за его высокой впитывающей способности.
В сельской местности Великобритании Fontinalis antipyretica традиционно использовался для тушения пожаров, поскольку его можно было найти в значительных количествах в медленно текущих реках, а мох удерживал большие объемы воды, которые помогали тушить пламя. Это историческое использование отражено в его конкретном латинско-греческом названии, примерное значение которого — «против огня».
Это историческое использование отражено в его конкретном латинско-греческом названии, примерное значение которого — «против огня».
В Финляндии торфяные мхи использовались для приготовления хлеба во время голода.
В Мексике мох используется в качестве рождественского украшения.
Моховой фотобиореактор с Physcomitrella patensPhyscomitrella patens все чаще используется в биотехнологии. Яркими примерами являются идентификация генов мхов, имеющих значение для улучшения урожая или здоровья человека, и безопасное производство сложных биофармацевтических препаратов в биореакторе со мхом, разработанном Ральфом Рески и его сотрудниками.
Ссылки
- Гоффине, Бернар; Уильям Р. Бак (2004). «Систематика мохообразных (мхов): от молекул к пересмотренной классификации». Монографии по систематической ботанике (издательство Missouri Botanical Garden Press) 98: 205–239.
- Бак, Уильям Р. и Бернард Гоффине. 2000. «Морфология и классификация мхов», страницы 71-123 в А.
 Джонатан Шоу и Бернард Гоффине (ред.), Биология мохообразных. (Кембридж: Издательство Кембриджского университета).
Джонатан Шоу и Бернард Гоффине (ред.), Биология мохообразных. (Кембридж: Издательство Кембриджского университета). - Томас, Б.А. (1972). «Вероятный мох из нижнего карбона в лесу Дин, Глостершир». Анналы ботаники 36 (1): 155–161.
- Коднер, Р. Б.; Грэм, Л. Э. (2001). «Высокотемпературные кислотно-гидролизованные остатки Polytrichum (Musci, Polytrichaceae) напоминают загадочные силурийско-девонские трубчатые микрофоссилии». Американский журнал ботаники 88 (3): 462–466.
- Стив Уитчер, главный садовник (1996). «Борьба с мхом на газонах» (Интернет). Садоводство в Западном Вашингтоне. Университет штата Вашингтон.
- Ральф Рески и Вольфганг Франк (2005): Функциональная геномика мха (Physcomitrella patens) – открытие генов и разработка инструментов с последствиями для сельскохозяйственных культур и здоровья человека. Брифинги по функциональной геномике и протеомике 4, 48-57.
- Ева Л. Декер и Ральф Рески (2007 г.): Моховые биореакторы, производящие улучшенные биофармацевтические препараты.
 Джонатан Шоу и Бернард Гоффине (ред.), Биология мохообразных. (Кембридж: Издательство Кембриджского университета).
Джонатан Шоу и Бернард Гоффине (ред.), Биология мохообразных. (Кембридж: Издательство Кембриджского университета).