
Содержание
Палеонтологи уточнили время и место появления сосудистых растений
В Саудовской Аравии в отложениях возрастом 444–450 млн лет (конец ордовикского периода) обнаружены споры сосудистых растений — группы, к которой относятся псилофиты, хвощи, плауны, папоротники, голосеменные и цветковые, то есть все высшие растения, кроме мхов. До сих пор бесспорные ископаемые остатки сосудистых растений были известны лишь начиная с середины силура (около 425 млн лет назад). Находка не только отодвинула в прошлое момент появления сосудистых растений, но и указала на место этого события — северную часть суперконтинента Гондвана, откуда сосудистые растения позже распространились по всему миру.
Наземные растения подразделяются на две большие группы: бриофиты (моховидные) и сосудистые. Последние, в свою очередь, делятся на споровые (хвощи, плауны, папоротники, вымершие риниофиты) и семенные (голосеменные, покрытосеменные).
Первые наземные растения появились не позднее середины ордовикского периода (около 465 млн лет назад). Их предками, скорее всего, были харовые водоросли. Древнейшие наземные растения почти не оставили следов в палеонтологической летописи, если не считать характерных спор, получивших название «криптоспоры» (cryptospores). Похожие споры ныне производят самые примитивные представители мохообразных — печеночники. На основе этого сходства, а также ряда других фактов, считается, что первые наземные растения были близки к мохообразным.
Их предками, скорее всего, были харовые водоросли. Древнейшие наземные растения почти не оставили следов в палеонтологической летописи, если не считать характерных спор, получивших название «криптоспоры» (cryptospores). Похожие споры ныне производят самые примитивные представители мохообразных — печеночники. На основе этого сходства, а также ряда других фактов, считается, что первые наземные растения были близки к мохообразным.
Криптоспоры широко распространены в отложениях ордовикского и силурийского периодов по всему миру. При этом они имеют весьма однообразное строение. Это говорит о том, что древнейшие мохообразные имели обширные ареалы и низкий уровень видового разнообразия. Это были эврибионтные (экологически непритязательные) растения, которые могли расти практически везде, где было достаточно влажно. По-видимому, влажные участки всех континентов в то время покрылись однообразной порослью мелких моховидных растений. Эта первичная наземная флора просуществовала без особых изменений свыше 30 млн лет — до тех пор, пока ей на смену не пришли новые растительные сообщества, в которых преобладали сосудистые растения.
Появление сосудистых растений, которые, в отличие от мохообразных, способны гораздо более эффективно противостоять высыханию, было одним из важнейших событий в эволюции наземной биоты. До сих пор считалось, что первые сосудистые растения появились в середине силурийского периода, примерно 420–430 млн лет назад. Об этом свидетельствует появление в палеонтологической летописи нового типа спор — так называемых трилетных спор с плотной орнаментированной оболочкой, а также первых фрагментарных остатков самих сосудистых растений. Трилетные споры отличаются от криптоспор, во-первых, тем, что они распространяются поодиночке (криптоспоры всегда образуют диады или тетрады), во-вторых, трилетные споры имеют характерную тетраэдрическую форму и Y-образную складку на поверхности (см. Спорангии и споры). В отличие от криптоспор и тех спор, которые производятся современными мохообразными, трилетные споры способны эффективно противостоять высыханию. Более или менее целые отпечатки примитивных сосудистых растений — риниофитов, или псилофитов, — известны начиная с конца силура и становятся многочисленными в раннем девоне (примерно 410–415 млн лет назад).
Новое открытие, о котором палеонтологи из Бельгии, Франции, Великобритании и Саудовской Аравии сообщили в последнем номере журнала Science, отодвигает момент появления сосудистых растений на 20 млн лет в прошлое. Исследователи обнаружили многочисленные трилетные споры, в том числе с орнаментированной оболочкой, в позднеордовикских отложениях на севере Саудовской Аравии, которая в ордовике входила в состав суперконтинента Гондваны. Споры обнаружены в 37 образцах, полученных в ходе бурения нефтяных скважин. Возраст самых древних из этих образцов составляет около 450, самых молодых — около 444 млн лет.
Ранее немногочисленные трилетные споры (правда, более примитивные — с гладкой, а не орнаментированной оболочкой) были найдены в отложениях конца ордовикского периода (444 млн лет назад) в Турции, которая тоже находилась в то время в северной части Гондваны. Турецкие находки сами по себе еще не позволяли с абсолютной уверенностью говорить о существовании сосудистых растений в ордовике: во-первых, эти находки были единичными, во-вторых, не было найдено спор со сложной орнаментированной оболочкой. Теперь же стало ясно, что настоящие сосудистые растения (вероятно, примитивные риниофиты) существовали на севере Гондваны задолго до конца ордовикского периода. По всей видимости, этот регион был центром происхождения сосудистых растений. В отличие от своих предполагаемых предков — мохообразных, размножавшихся при помощи криптоспор, — первые сосудистые растения быстро достигли высокого уровня видового разнообразия (и, следовательно, экологической специализации). Об этом свидетельствует разнообразие трилетных спор, найденных в Саудовской Аравии.
Теперь же стало ясно, что настоящие сосудистые растения (вероятно, примитивные риниофиты) существовали на севере Гондваны задолго до конца ордовикского периода. По всей видимости, этот регион был центром происхождения сосудистых растений. В отличие от своих предполагаемых предков — мохообразных, размножавшихся при помощи криптоспор, — первые сосудистые растения быстро достигли высокого уровня видового разнообразия (и, следовательно, экологической специализации). Об этом свидетельствует разнообразие трилетных спор, найденных в Саудовской Аравии.
В течение ордовика сосудистым растениям не удавалось выйти за пределы своей «исторической родины» (поскольку в других регионах в ордовикских отложениях трилетные споры не обнаружены, несмотря на хорошую изученность этих отложений).
В самом конце ордовика имело место одно из крупнейших в истории Земли оледенений. Ледники покрыли большую часть Гондваны, но сосудистые растения сумели пережить этот неблагоприятный период. В раннем силуре, после отступления ледников, они начали распространяться за пределы родного континента, постепенно вытесняя первичную флору их моховидных растений, размножавшихся при помощи криптоспор. Однако занять по-настоящему господствующее положение в растительных сообществах сосудистым растениям удалось лишь много позже, в конце силура — начале девона.
Однако занять по-настоящему господствующее положение в растительных сообществах сосудистым растениям удалось лишь много позже, в конце силура — начале девона.
Источник: Philippe Steemans, Alain Le Hérissé, John Melvin, Merrell A. Miller, Florentin Paris, Jacques Verniers, Charles H. Wellman. Origin and Radiation of the Earliest Vascular Land Plants // Science. 2009. V. 324. P. 353.
См. также:
Evolutionary history of plants.
Александр Марков
|
Эволюция царства растений началась с архейской эры (около 3500 млн. лет назад). В этот период на Земле отмечено появление синезеленых водорослей. Эти водоросли принадлежат к группе цианобактерий, поскольку в их клетках отсутствует оформленные ядра. Таким образом, их можно отнести к прокариотам (доядерным организмам). Среди синезеленых водорослей были одно- и многоклеточные организмы, имеющие возможность осуществлять фотосинтез.
Позднее в протерозойской эре (около 2600 млн. лет назад) Землей завладели красные и зеленые водоросли. Их господство распространилось и на палеозойскую эру (примерно 570 млн. лет назад). Только в поздний палеозой (силурийский период) отмечена жизнедеятельность на планете древнейших высших растений – риниофитов, или псилофитов. У этих растений были побеги, но отсутствовали корни и листья. Размножение риниофитов происходило спорами. Они обитали на суше или частично в воде.В существовании нашей планеты новая эра началась с появления высших, или наземных растений. Около 400- 360 млн. лет назад в девонском периоде палеозойской эры на фоне преобладания на Земле риниофитов и водорослей появились первые папоротникообразные (папоротники, хвощи, плауны) и моховидные растения. Они относятся к высшим споровым растениям. Благодаря распространению растений на суше, появились и новые наземные виды животных.
В каменноугольном периоде, который длился с 360 до 280 млн. лет назад, доказано появление на нашей планете семенных папоротников, которые в дальнейшем стали родоначальниками всех голосеменных растений. В это время полностью исчезают риниофиты из-за невозможности конкурировать с более развитыми растениями. А господствовашие тогда огромнейшие древовидные папоротникообразные после отмирания образовали залежи каменного угля.
В следующем пермском периоде палеозойской эры на Земле появились древние голосеменные растения. Древовидные папоротникообразные постепенно вымирают, а им на смену приходят семенные и травянистые папоротники, завладевая сушей. Особенностью голосеменных растений является то, что размножение их осуществляется семенами, не имеющими защиты в виде стенок плода, так как цветков и плодов эти растения не образуют.
Мезозойская эра наступила около 240 млн. лет назад. В триасовом периоде мезозоя появились современные голосеменные, а в юрском периоде зародились первые покрытосеменные растения. Но господствующие позиции на планете сохранились за голосеменными растениями. Это эра вымирания древних папоротникообразных, не выдерживающих естественного отбора. В процессе появления покрытосеменных растений произошла череда ароморфозов. Во-первых, сформировался цветок – трансформированный побег, приспособленный для образования спор и гамет. Опыление, оплодотворение и образование зародыша и плода происходило непосредственно в цветке. Во-вторых, для лучшей защиты и распространения семена покрытосеменных растений были окружены околоплодником.
В кайнозойской эре, наступившей приблизительно 70 млн. лет назад, на нашей планете стали господствовать существующие и в настоящее время покрытосеменные и голосеменные растения, тогда как высшие споровые растения регрессировали.
Сейчас на Земле произрастают более 350 видов растений, среди них встречаются цветковые, мохообразные растения, папоротники, водоросли.
|
 Благодаря процессу фотосинтеза, в атмосферу нашей планеты начал поступать кислород, необходимый для жизнедеятельности аэробов.
Благодаря процессу фотосинтеза, в атмосферу нашей планеты начал поступать кислород, необходимый для жизнедеятельности аэробов. Сочетанное изменение в ходе эволюции форм растений и животных обусловило огромнейшее разнообразие жизни на Земле. Облик планеты изменился коренным образом. Прикрепленный образ жизни растения на суше привел к появлению расчленения растения на корень, стебель и лист, а также к возникновению опорных тканей и сосудистой проводящей системы. Самые первые наземные растения были маленьких размеров. Они поглощали воду посредством ризоидов, как сохранившиеся по сей день на Земле мхи. В цикле их развития преобладало гаплоидное поколение (гаметофит). Постепенно появлялись более крупные формы растений — папоротникообразные, у которых образовались сложные специализированные органы – корни с корневыми волосками. В цикле развития этих растений на первый план выступает диплоидная фаза — спорофит, являющийся непосредственно самим растением, тогда как гаметофит – это заросток, который выглядит как клубенек у хвощей и плаунов и как небольшая пластинка в форме сердца у папоротников. Так осуществлялся постепенный переход от гаплоидного поколения к более совершенному — диплоидному.
Сочетанное изменение в ходе эволюции форм растений и животных обусловило огромнейшее разнообразие жизни на Земле. Облик планеты изменился коренным образом. Прикрепленный образ жизни растения на суше привел к появлению расчленения растения на корень, стебель и лист, а также к возникновению опорных тканей и сосудистой проводящей системы. Самые первые наземные растения были маленьких размеров. Они поглощали воду посредством ризоидов, как сохранившиеся по сей день на Земле мхи. В цикле их развития преобладало гаплоидное поколение (гаметофит). Постепенно появлялись более крупные формы растений — папоротникообразные, у которых образовались сложные специализированные органы – корни с корневыми волосками. В цикле развития этих растений на первый план выступает диплоидная фаза — спорофит, являющийся непосредственно самим растением, тогда как гаметофит – это заросток, который выглядит как клубенек у хвощей и плаунов и как небольшая пластинка в форме сердца у папоротников. Так осуществлялся постепенный переход от гаплоидного поколения к более совершенному — диплоидному. В палеозойскую эру папоротникообразные были громадными растениями, господствовавшими на суше. Однако для их размножения была необходима вода, что ограничивало территорию их существования местностями с повышенной влажностью.
В палеозойскую эру папоротникообразные были громадными растениями, господствовавшими на суше. Однако для их размножения была необходима вода, что ограничивало территорию их существования местностями с повышенной влажностью. Половое размножение этих растений осуществлялось независимо от капельно-водной среды. А их появление в ходе эволюционных метаморфоз было обусловлено перепадами влажности и температуры и изменением рельефа Земли вследствие поднятия суши, то есть появления горных массивов.
Половое размножение этих растений осуществлялось независимо от капельно-водной среды. А их появление в ходе эволюционных метаморфоз было обусловлено перепадами влажности и температуры и изменением рельефа Земли вследствие поднятия суши, то есть появления горных массивов. Для этих растений характерно половое размножение. К покрытосеменным относятся травянистые растения, деревья и кустарники. Разнообразные видоизменения вегетативных органов (корня, стебля, листа) отмечаются у разных видов растений. Эволюционные изменения покрытосеменных растений происходили за относительно короткий срок, поэтому для них свойственна высокая эволюционная пластичность. Огромное значение в протекании эволюционных преобразований сыграли насекомые-опылители. Покрытосеменные растения более продуктивно осваивают окружающую среду и завоевывают новые территории, благодаря своим особенностям строения и способности формировать сложные многоярусные сообщества.
Для этих растений характерно половое размножение. К покрытосеменным относятся травянистые растения, деревья и кустарники. Разнообразные видоизменения вегетативных органов (корня, стебля, листа) отмечаются у разных видов растений. Эволюционные изменения покрытосеменных растений происходили за относительно короткий срок, поэтому для них свойственна высокая эволюционная пластичность. Огромное значение в протекании эволюционных преобразований сыграли насекомые-опылители. Покрытосеменные растения более продуктивно осваивают окружающую среду и завоевывают новые территории, благодаря своим особенностям строения и способности формировать сложные многоярусные сообщества.
Первые наземные растения и грибы изменили климат Земли, проложив путь для взрывной эволюции наземных животных, новое исследование генов предполагает
Лишайники и мох, покрывающие скалы в Пенсильвании. Так ли выглядела докембрийская земля? Фото: Дэвид Гейзер, штат Пенсильвания
Крупнейшее генетическое исследование, когда-либо проводившееся для выяснения того, когда наземные растения и грибы впервые появились на Земле, выявило правдоподобную биологическую причину двух основных климатических явлений: эпохи снежного кома Земли, когда лед периодически охватывала земной шар, и наступила эпоха, названная Кембрийским взрывом, в результате которого появились первые окаменелости почти всех основных категорий животных, живущих сегодня.
По мнению авторов исследования, которое будет опубликовано в выпуске журнала Science от 10 августа 2001 года, растений проложили путь эволюции наземных животных, одновременно увеличивая процентное содержание кислорода в атмосфере Земли и уменьшая процент углекислого газа, мощного парникового газа.
«Наше исследование показывает, что наземные растения и грибы эволюционировали намного раньше, чем считалось ранее — до событий Земли-снежка и Кембрийского взрыва — что позволяет предположить, что их присутствие могло оказать глубокое влияние на климат и эволюцию жизни на Земле», — говорится в сообщении 9.0017 Блэр Хеджес , биолог-эволюционист и руководитель исследовательской группы штата Пенсильвания, проводившей исследование.
Исследователи обнаружили, что наземные растения появились на Земле около 700 миллионов лет назад, а наземные грибы — около 1300 миллионов лет назад — намного раньше, чем предыдущие оценки около 480 миллионов лет назад, основанные на самых ранних окаменелостях этих организмов. . До этого исследования считалось, что ландшафт Земли в то время был покрыт бесплодными скалами, на которых не было ничего, кроме некоторых бактерий и, возможно, некоторых водорослей. По словам Хеджеса, в горных породах, образовавшихся в докембрийский период, не было обнаружено бесспорных окаменелостей самых ранних наземных растений и грибов, возможно, потому, что их примитивные тела были слишком мягкими, чтобы превратиться в окаменелости.
Раннее появление на земле грибов и растений позволяет предположить их вероятную роль как в загадочном понижении температуры поверхности Земли во время серии событий Земли-снежка примерно от 750 до 580 миллионов лет назад, так и во внезапном появлении многих новых видов ископаемые животные в эпоху кембрийского взрыва примерно 530 миллионов лет назад. «Как понижение температуры поверхности Земли, так и эволюция многих новых видов животных могли быть результатом уменьшения содержания углекислого газа в атмосфере и повышения содержания кислорода, вызванного присутствием на земле в это время лишайниковых грибов и растений, что подтверждается нашими исследованиями. предлагает», — говорит Хеджес.
«Увеличение численности наземных растений могло произойти непосредственно перед периодом, известным как Кембрийский взрыв, когда следующий период Земли-снежка не наступил, потому что температура не стала достаточно низкой», — говорит Хеджес. «Растения, по-видимому, повысили уровень кислорода в атмосфере настолько, что у животных развился скелет, они стали крупнее и разнообразнее».
Считается, что лишайники были первыми грибами, объединившимися с фотосинтезирующими организмами, такими как цианобактерии и зеленые водоросли. Лишайники могут месяцами жить без дождя, обеспечивая защиту для фотосинтезирующих организмов, которые производят кислород и выделяют его в атмосферу. Исследователи предполагают, что грибы-лишайники-первопроходцы, которые производят кислоты, достаточно сильные для растворения камней, также могли помочь уменьшить выбросы углекислого газа. При вымывании дождевой водой кальций, высвобождаемый из скал, покрытых лишайником, в конечном итоге образует известняк из карбоната кальция в океане, не позволяя атомам углерода образовывать парниковый газ, двуокись углерода, в атмосфере.
Наземные растения также могут снижать уровень углекислого газа в атмосфере. У них есть молекулы, называемые лигнинами, которые содержат углерод, но плохо разлагаются. После того, как растение умирает, часть его углерода остается запертой в лигнинах и может быть похоронена в Земле в результате геологических процессов, предотвращая возвращение этих атомов углерода в атмосферу и эффективно снижая содержание углекислого газа в атмосфере.
Лишайники и мох, покрывающие скалы в Пенсильвании. Кредит Фотографии: Дэвид Гейзер, Penn State
«Земля охлаждается, когда вы удаляете углекислый газ», — говорит Хеджес. «Другие факторы, такие как расположение континентов, могли иметь некоторое влияние на охлаждение атмосферы и создание периодов Земли-снежка, но я подозреваю, что самый большой охлаждающий эффект был вызван сокращением углекислого газа в атмосфере грибами и растениями, которые мы показали, что жили на земле в то время».
Ископаемые виды топлива, такие как уголь и нефть, производятся из растительного материала, содержащего углерод, который был взят из атмосферы и захоронен в болотах миллионы лет назад. Согласно многим исследованиям, выброс тех же самых атомов углерода обратно в атмосферу при сжигании ископаемого топлива, по-видимому, заставляет Землю снова нагреваться.
Хеджес и его исследовательская группа сделали удивительные открытия о раннем появлении на Земле первых наземных растений и грибов, изучив как можно больше генов их потомков — видов растений и грибов, живущих сегодня. Они начали с просеивания своих молекулярных отпечатков пальцев — уникальных последовательностей строительных блоков аминокислот — во многих тысячах генов сотен видов, заархивированных в общедоступных базах данных последовательностей генов.
Они начали с просеивания своих молекулярных отпечатков пальцев — уникальных последовательностей строительных блоков аминокислот — во многих тысячах генов сотен видов, заархивированных в общедоступных базах данных последовательностей генов.
В конце концов, они нашли 119гены, общие для живых видов грибов, растений и животных, которые соответствовали строгим критериям исследователей для использования в качестве «молекулярных часов». В предыдущих исследованиях использовался один ген. Путем детального сравнения аминокислотных последовательностей отдельных генов среди множества видов ученые выявили те гены, которые накапливали мутации с довольно постоянной скоростью по отношению друг к другу в ходе своей эволюции. «Поскольку мутации в этих генах начинают происходить через равные промежутки времени, как только появляется новый вид — как тиканье часов — мы можем использовать их, чтобы проследить эволюционную историю вида до момента его происхождения», — объясняет Хеджес.
Ученые откалибровали каждый из своих генных часов с эволюционными событиями, хорошо установленными в исследованиях окаменелостей, в первую очередь в истории животных. Используя эти известные даты в качестве надежных калибровочных точек, а скорость мутаций для каждого из генов с постоянной скоростью в качестве устройства измерения времени, исследователи смогли определить, как давно возник каждый из видов.
Используя эти известные даты в качестве надежных калибровочных точек, а скорость мутаций для каждого из генов с постоянной скоростью в качестве устройства измерения времени, исследователи смогли определить, как давно возник каждый из видов.
Хеджес говорит, что его исследование может помочь в поисках жизни на других планетах, установив связь между различными стадиями эволюции жизни на Земле и синхронизацией событий химической эволюции земной атмосферы, таких как увеличение содержания кислорода. «Возможно, ранняя история жизни на Земле может дать нам ключ к предсказанию видов форм жизни, которые могут существовать на планетах в других солнечных системах, исходя из химического состава их атмосфер», — говорит Хеджес.
Помимо Хеджеса, исследовательская группа штата Пенсильвания включает Дэниела С. Хекмана , студента бакалавриата, чья дипломная работа с отличием стала частью этого исследования; Дэвид М. Гейзер , доцент кафедры патологии растений; и студенты бакалавриата Брук Р. Эйделл ; Ребекка Л. Штауффер ; и Натали Л. Кардос. Это исследование было частично поддержано Национальным управлением по аэронавтике и исследованию космического пространства через Исследовательский центр астробиологии штата Пенсильвания.
Эйделл ; Ребекка Л. Штауффер ; и Натали Л. Кардос. Это исследование было частично поддержано Национальным управлением по аэронавтике и исследованию космического пространства через Исследовательский центр астробиологии штата Пенсильвания.
КОНТАКТЫ:
Блэр Хеджес, 814-865-9991, [email protected]
Барбара К. Кеннеди, 814-863-4682, [email protected]
ИЗОБРАЖЕНИЯ:
дополнительных высококачественных цветных изображения лишайники можно получить непосредственно у Стивена Шарноффа, , , профессионального фотографа и коллеги исследователей, по электронной почте [email protected], по телефону 510-548-9189 или по почте 2406 Roosevelt Ave., Berkeley, CA. 94703.
Фрагменты древнейших наземных растений
- Опубликовано:
- Чарльз Х.
 Веллман 1 ,
Веллман 1 , - Питер Л. Остерлофф 2 nAff3 и
- Узма Мохиуддин 2
 Веллман 1 ,
Веллман 1 ,79
Природа
том 425 , страницы 282–285 (2003 г.)Процитировать эту статью
4280 доступов
406 цитирований
36 Альтметрический
Сведения о показателях
Abstract
Самые ранние окаменелости наземных растений происходят из микроскопических рассеянных спор 1,2,3 . Эти микрофоссилии многочисленны и широко распространены в отложениях, и самые ранние общепринятые сообщения относятся к породам среднего ордовикского возраста (Лланвирн, 475 миллионов лет назад) 4 . Хотя распределение, морфология и ультраструктура спор указывают на то, что они произошли от наземных растений, возможно, ранних родственников мохообразных, эта интерпретация остается спорной 9.0083 5 , так как прямых доказательств родительских растений немного. Дополнительным осложняющим фактором является то, что между появлением первых рассеянных спор и окаменелостей относительно полных наземных растений (мегафоссилий) 6 существует значительный промежуток времени: споры предшествуют самым ранним мегафоссилиям (поздний силур, 425 миллионов лет назад) примерно на 50 лет вперед. миллион лет 7 . Здесь мы приводим описание споросодержащих фрагментов растений из ордовикских пород Омана. Эти окаменелости являются прямым свидетельством природы растений, производящих споры.
Эти микрофоссилии многочисленны и широко распространены в отложениях, и самые ранние общепринятые сообщения относятся к породам среднего ордовикского возраста (Лланвирн, 475 миллионов лет назад) 4 . Хотя распределение, морфология и ультраструктура спор указывают на то, что они произошли от наземных растений, возможно, ранних родственников мохообразных, эта интерпретация остается спорной 9.0083 5 , так как прямых доказательств родительских растений немного. Дополнительным осложняющим фактором является то, что между появлением первых рассеянных спор и окаменелостей относительно полных наземных растений (мегафоссилий) 6 существует значительный промежуток времени: споры предшествуют самым ранним мегафоссилиям (поздний силур, 425 миллионов лет назад) примерно на 50 лет вперед. миллион лет 7 . Здесь мы приводим описание споросодержащих фрагментов растений из ордовикских пород Омана. Эти окаменелости являются прямым свидетельством природы растений, производящих споры. Они подтверждают, что самые ранние споры в большом количестве развивались в спорангиях, предоставляя убедительные доказательства того, что они являются окаменелыми остатками настоящих наземных растений. Кроме того, анализ ультраструктуры стенок спор подтверждает сходство с печеночниками.
Они подтверждают, что самые ранние споры в большом количестве развивались в спорангиях, предоставляя убедительные доказательства того, что они являются окаменелыми остатками настоящих наземных растений. Кроме того, анализ ультраструктуры стенок спор подтверждает сходство с печеночниками.
Это предварительный просмотр содержимого подписки, доступ через ваше учреждение
Соответствующие статьи
Статьи открытого доступа со ссылками на эту статью.
Молекулярная делимитация европейских листовых печеночников рода Calypogeia на основе пластидных суперштрих-кодов
- Моника Слипико
- , Камиль Мыщинский
- … Якуб Савицкий
BMC Биология растений
Открытый доступ
28 мая 2020 г.

Варианты доступа
Подписаться на журнал
Получите полный доступ к журналу на 1 год
199,00 €
всего 3,90 € за выпуск
Подписаться
Расчет налогов будет завершен во время оформления заказа.
Купить статью
Получите ограниченный по времени или полный доступ к статье на ReadCube.
$32,00
Купить
Все цены указаны без учета стоимости.
Рисунок 1: Фрагменты ископаемых растений из скважины Габа-1, керн 21. Рисунок 2: Образец CW47c.
Ссылки
Грей, Дж. и Буко, А. Дж. Раннесилурийские тетрады спор из Нью-Йорка: самые ранние свидетельства существования сосудистых растений в Новом Свете? Наука 173 , 918–921 (1971)
Статья
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Грей, Дж.
Микрофоссилии ранних наземных растений: достижения в понимании ранней земной жизни, 1970–1984 гг. Фил. Транс. Р. Соц. Лонд. В 309 , 167–195 (1985)Статья
ОБЪЯВЛЕНИЯGoogle ученый
Wellman, CH & Gray, J. Летопись микрофоссилий ранних наземных растений. Фил. Транс. Р. Соц. Лонд. B 355 , 717–732 (2000)
Статья
КАСGoogle ученый
Стротер, П.К., Аль-Хаджри, С. и Траверс, А. Новые данные о наземных растениях нижнего среднего ордовика Саудовской Аравии. Геология 24 , 55–58 (1996)
Статья
ОБЪЯВЛЕНИЯGoogle ученый
Banks, H. P. Самые старые сосудистые наземные растения: предостережение. Преподобный Палеобот. Палинол. 20 , 13–25 (1975)
Статья
Google ученый
Эдвардс, Д.
и Фихан, Дж. Записи о спорангиях типа Cooksonia из поздних слоев Венлока в Ирландии. Природа 287 , 41–42 (1980)Статья
ОБЪЯВЛЕНИЯGoogle ученый
Эдвардс, Д. и Веллман, С. Х. в Plants Invade the Land (ред. Гензель, П. Г. и Эдвардс, Д.) 3–28 (Columbia Univ. Press, Нью-Йорк, 2001)
Google ученый
Дросте, HHJ Стратиграфия нижнепалеозойской надгруппы Хайма в Омане. ГеоАрабия 2 , 419–468 (1997)
Google ученый
Мохиуддин У. . Количественный анализ палиноморфов ордовика Омана Диссертация (Университет Шеффилда)
Эдвардс, Д., Селден, П.А., Ричардсон, Дж. Б. и Акс, Л. Копролиты как свидетельство взаимодействия растений и животных в силуро-девонских наземных экосистемах.
Природа 377 , 329–331 (1995)Статья
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Тейлор, В. А. Исследования ультраструктуры криптоспор: изменчивость рода тетрад Tetrahedraletes и типовой материал диады Dyadospora murusattenuata . Преподобный Палеобот. Палинол. 119 , 325–334 (2002)
Статья
Google ученый
Пачини, Э. в Микроспоры: эволюция и онтогенез (ред. Блэкмор, С. и Нокс, Р. Б.) 213–237 (Академический, Лондон, 1990)
Книга
Google ученый
Эдвардс, Д. Новый взгляд на ранние наземные экосистемы: взгляд на мир лилипутов. Преподобный Палеобот. Палинол. 90 , 159–174 (1996)
Статья
Google ученый
Ричардсон, Дж.
Б. в Подповерхностная палиностратиграфия северо-востока Ливии (ред. Эль-Арнаути, Б., Оуэнс, Б. и Тусу, Б.) 89–109 (Университет Гариунис, Бенгази, Ливия, 1988 г.)Google ученый
Steemans, P. Палеодиверсификация спор и криптоспор де L’Ordovician au Dévonien inferieur. Геобиос 32 , 341–352 (1999)
Статья
Google ученый
Taylor, WA Споры в самых ранних наземных растениях. Природа 373 , 391–392 (1995)
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Эдвардс, Д., Дакетт, Дж. Г. и Ричардсон, Дж. Б. Печеночные признаки у самых ранних наземных растений. Природа 374 , 635–636 (1995)
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Wellman, C.
H., Edwards, D. & Axe, L. Ультраструктура в лавигатных спорах хилатов из спорангиев и споровых масс из верхнего силура и нижнего девона Уэльской границы. Фил. Транс. Р. Соц. Лонд. B 153 , 1983–2004 (1998)Артикул
Google ученый
Веллман, С. Х., Эдвардс, Д. и Акс, Л. Постоянные диады в спорангиях и споровых массах из нижнего девона Уэльской границы. Бот. Дж. Линн. соц. 127 , 117–147 (1998)
Статья
Google ученый
Эдвардс, Д., Веллман, С. Х. и Акс, Л. Тетрады в спорангиях и споровых массах формируют верхний силур и нижний девон валлийской границы. Бот. Дж. Линн. соц. 130 , 111–156 (1999)
Статья
Google ученый
Graham, L.E. Происхождение наземных растений (Wiley, New York, 1993)
Google ученый
Brown, R.
C. & Lemmon, B.E. в Microspores: Evolution and Ontogeny (ред. Blackmore, S. & Knox, R.B.) 55–94 (Academic, London, 1990)Книга
Google ученый
Кенрик, П. Отношения сосудистых растений. Фил. Транс. Р. Соц. Лонд. B 355 , 847–855 (2000)
Статья
КАСGoogle ученый
Kenrick, P. & Crane, P. R. The Origin and Early Diversification of Land Plants (Смитсоновский институт, Вашингтон, 1997)
Google ученый
Nickrent, D.L. et al. Мультигенная филогения наземных растений с особым упором на мохообразные и самые ранние наземные растения. Мол. биол. Эвол. 17 , 1885–1895 (2000)
Артикул
КАСGoogle ученый
Цю, Ю.
Л. и Ли, Дж. Переход к наземной флоре: молекулярная филогенетическая перспектива. J. Phycol. 36 , 799–802 (2000)Артикул
КАСGoogle ученый
Крокен, С. Б., Грэм, Л. Э. и Кук, М. Э. Возникновение и эволюционное значение устойчивых клеточных стенок у харофитов и мохообразных. утра. Дж. Бот. 83 , 1241–1254 (1996)
Артикул
Google ученый
 Микрофоссилии ранних наземных растений: достижения в понимании ранней земной жизни, 1970–1984 гг. Фил. Транс. Р. Соц. Лонд. В 309 , 167–195 (1985)
Микрофоссилии ранних наземных растений: достижения в понимании ранней земной жизни, 1970–1984 гг. Фил. Транс. Р. Соц. Лонд. В 309 , 167–195 (1985)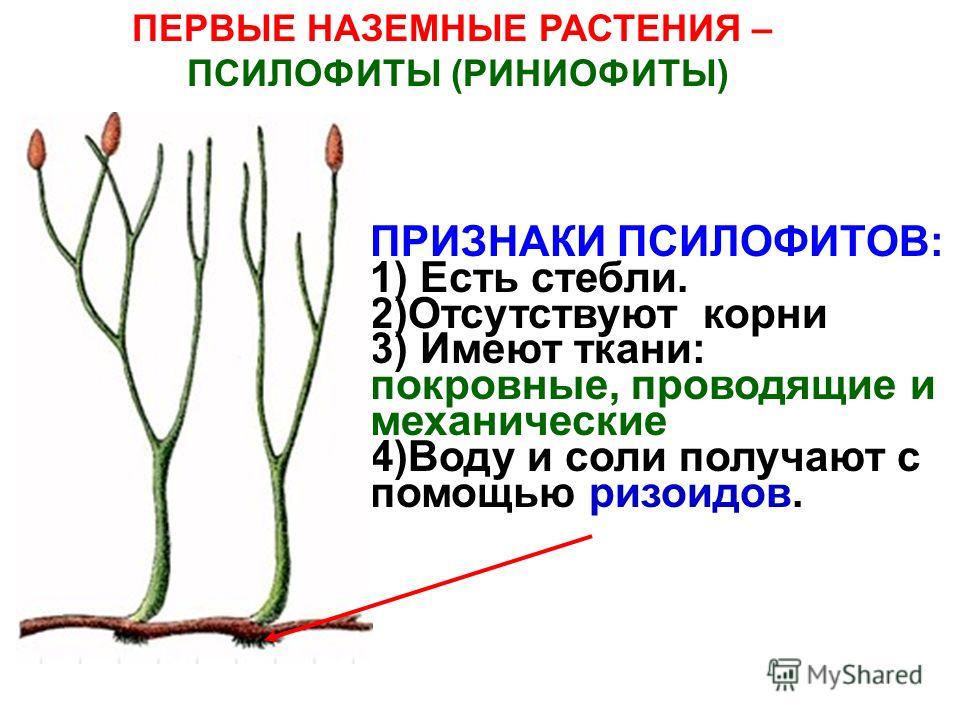 и Фихан, Дж. Записи о спорангиях типа Cooksonia из поздних слоев Венлока в Ирландии. Природа 287 , 41–42 (1980)
и Фихан, Дж. Записи о спорангиях типа Cooksonia из поздних слоев Венлока в Ирландии. Природа 287 , 41–42 (1980) Природа 377 , 329–331 (1995)
Природа 377 , 329–331 (1995) Б. в Подповерхностная палиностратиграфия северо-востока Ливии (ред. Эль-Арнаути, Б., Оуэнс, Б. и Тусу, Б.) 89–109 (Университет Гариунис, Бенгази, Ливия, 1988 г.)
Б. в Подповерхностная палиностратиграфия северо-востока Ливии (ред. Эль-Арнаути, Б., Оуэнс, Б. и Тусу, Б.) 89–109 (Университет Гариунис, Бенгази, Ливия, 1988 г.) H., Edwards, D. & Axe, L. Ультраструктура в лавигатных спорах хилатов из спорангиев и споровых масс из верхнего силура и нижнего девона Уэльской границы. Фил. Транс. Р. Соц. Лонд. B 153 , 1983–2004 (1998)
H., Edwards, D. & Axe, L. Ультраструктура в лавигатных спорах хилатов из спорангиев и споровых масс из верхнего силура и нижнего девона Уэльской границы. Фил. Транс. Р. Соц. Лонд. B 153 , 1983–2004 (1998) C. & Lemmon, B.E. в Microspores: Evolution and Ontogeny (ред. Blackmore, S. & Knox, R.B.) 55–94 (Academic, London, 1990)
C. & Lemmon, B.E. в Microspores: Evolution and Ontogeny (ред. Blackmore, S. & Knox, R.B.) 55–94 (Academic, London, 1990) Л. и Ли, Дж. Переход к наземной флоре: молекулярная филогенетическая перспектива. J. Phycol. 36 , 799–802 (2000)
Л. и Ли, Дж. Переход к наземной флоре: молекулярная филогенетическая перспектива. J. Phycol. 36 , 799–802 (2000)Ссылки на скачивание
Благодарности
Это исследование было поддержано грантом NERC. Мы хотели бы поблагодарить Petroleum Development Oman и Министерство нефти и газа Омана за предоставление материала для изучения и за разрешение на публикацию.
Информация о авторе
Авторы Примечания
Питер Л. Остерлофф
Настоящий адрес: Sarawak Shell Berhad, Locked Bag № 1, 98009, Miri, Sarawak, Malaysia
Авторы и Affiliations
44444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444449н. наук о животных и растениях, Шеффилдский университет, здание Альфреда Денни, Western Bank, S10 2TN, Sheffield, UK
наук о животных и растениях, Шеффилдский университет, здание Альфреда Денни, Western Bank, S10 2TN, Sheffield, UK
Charles H. Wellman
Petroleum Development Oman, PO Box 81, Muscat, 113, Sultanate of Oman
Peter L. Osterloff & Uzma Mohiuddin
Авторы
- Charles H. Wellman
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Peter L. Osterloff
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Узма Мохиуддин
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Академия
Автор, ответственный за переписку
Чарльз Х. Веллман.
Декларация этики
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих финансовых интересов.
Права и разрешения
Перепечатка и разрешения
Об этой статье
Эта статья цитируется
Простой протокол для генетической трансформации на основе культур таллома печеночника Marchantia polymorpha
- Дон Хе Со
- Соджон Ким
- У Тэк Ким
Журнал биологии растений (2022)
Настройка устойчивости к засухе с использованием специфичного для корней экспрессионного фактора транскрипции PdNF-YB21 в Arabidopsis thaliana
- Юэ Чжан
- Чао Шэнь
- Синьли Ся
Культура растительных клеток, тканей и органов (PCTOC) (2021)
Разработка нового метода одновременной экстракции хлорофиллов и каротиноидов из биомассы микроводорослей
- Ракель Гидетти Вендрусколо
- Андресса Сильва Фернандес
- Роджер Вагнер
Журнал прикладной психологии (2021)
Палеоэкология и сохранение исчезающих скрытых видов; пример печеночника Riella (Riellaceae)
- Мортеза Джамали
- Хосе Габриэль Сегарра-Морагес
Биоразнообразие и сохранение (2021)
Молекулярная делимитация европейских листовых печеночников рода Calypogeia на основе пластидных суперштрих-кодов
- Моника Слипико
- Камиль Мыщинский
- Якуб Савицкий
Биология растений BMC (2020)
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества.