Содержание
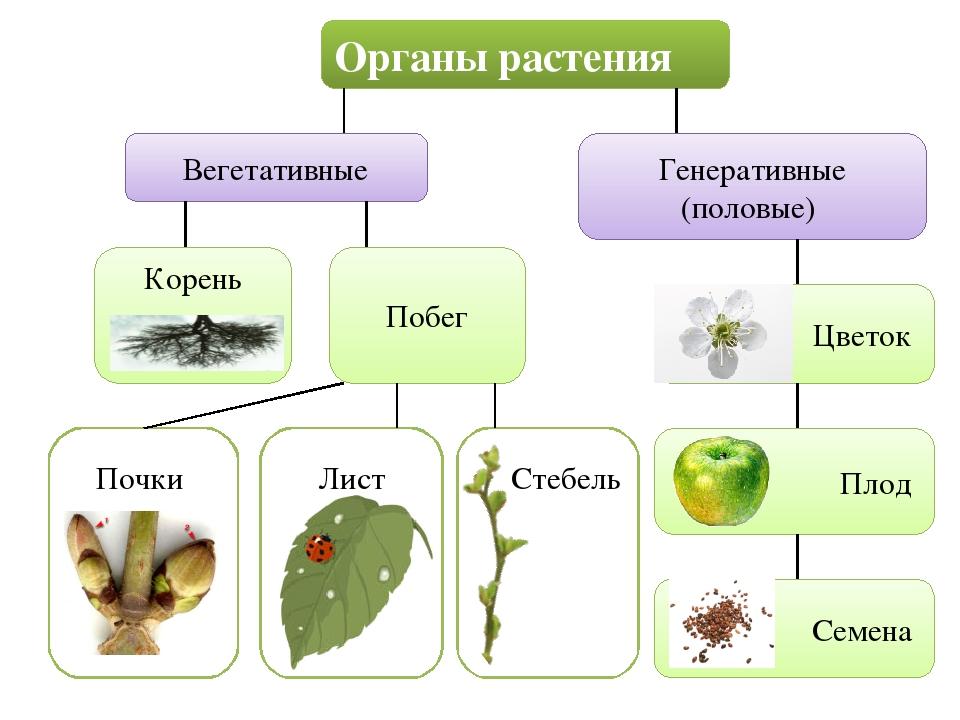
Вегетативные органы растений
Министерство
сельского хозяйства РФ
Департамента
научно-технологической
образования
ФГБОУ ВПС
Волгоградская
ГАУ
Кафедра:
ТПППХиТ
Дисциплина:
Анатомия
пищевого
сырья
РЕФЕРАТ
на тему:
«Вегетативные органы растений»
Выполнил:
Сочина Д.А.
ТОВ-23
Проверил:
доцент
Радзиевский
Е.Б.
Волгоград 2012
Содержание
Введение
Глава1.Корень
- Основные функции корня
- Происхождение корня
- Видоизменения и специализация корней
- Особенности строения корней
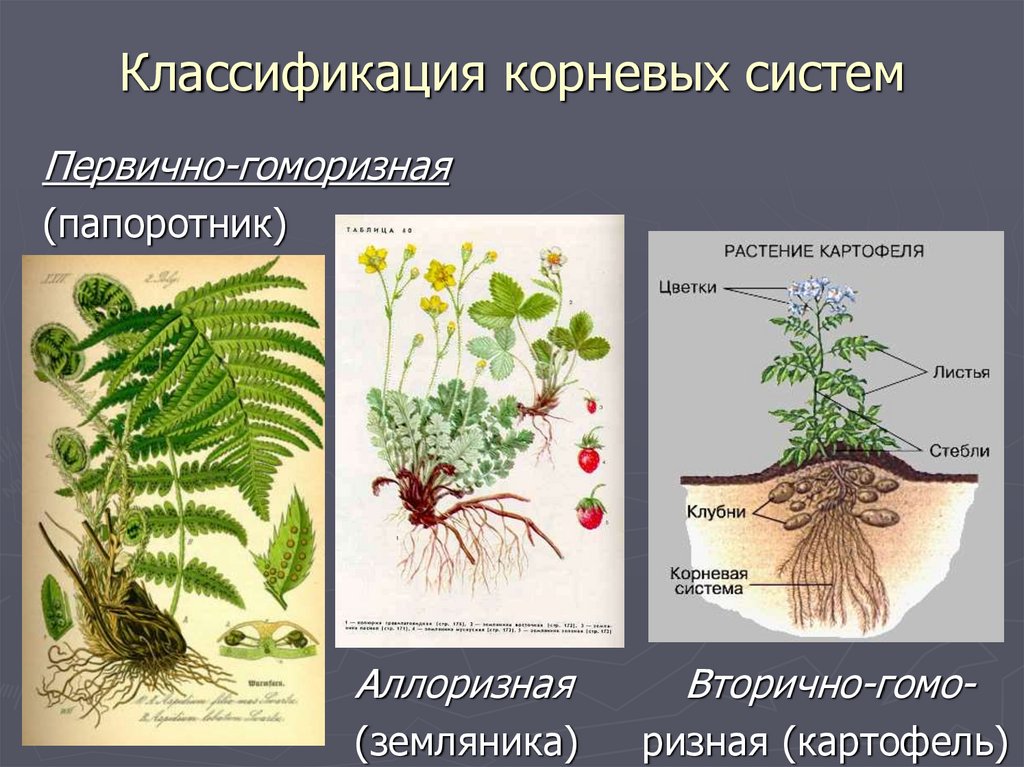
- Типы корневых систем
- Зоны молодого корневого окончания
- Анатомическое строение корня
Глава2.
Побег
- Филогенез побега
- Онтогенез побега
- Основные органы побега
- Особенности роста побегов
- Ветвление побегов
- Видоизменения побегов (метаморфоз)
Список литературы
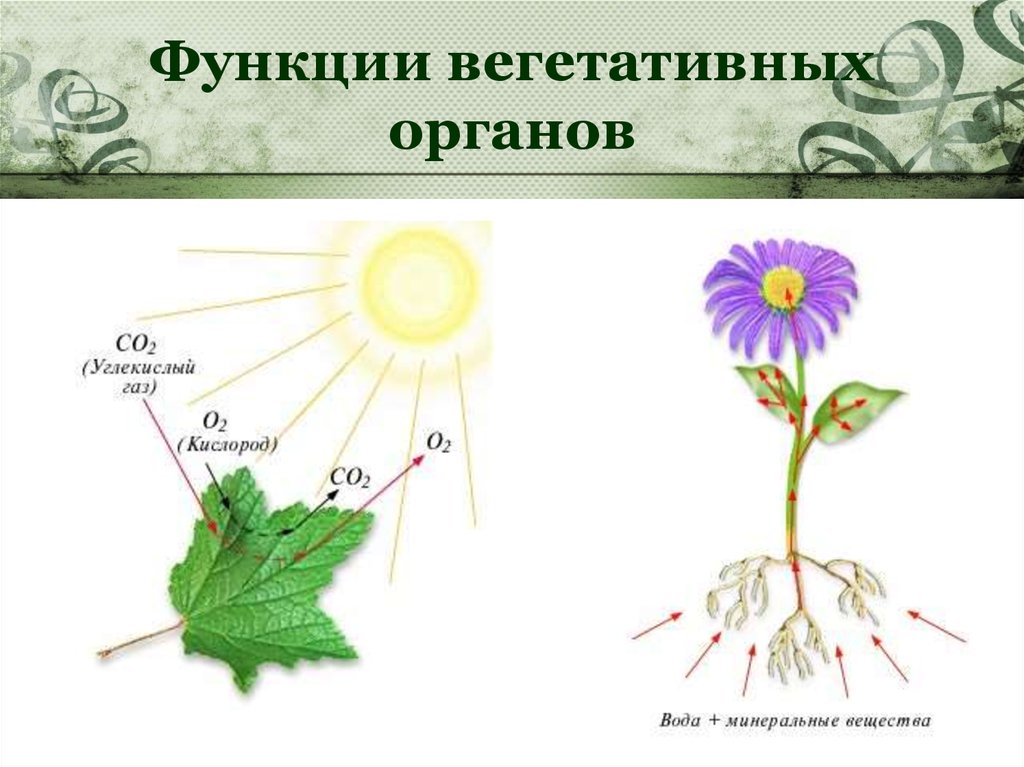
Введение
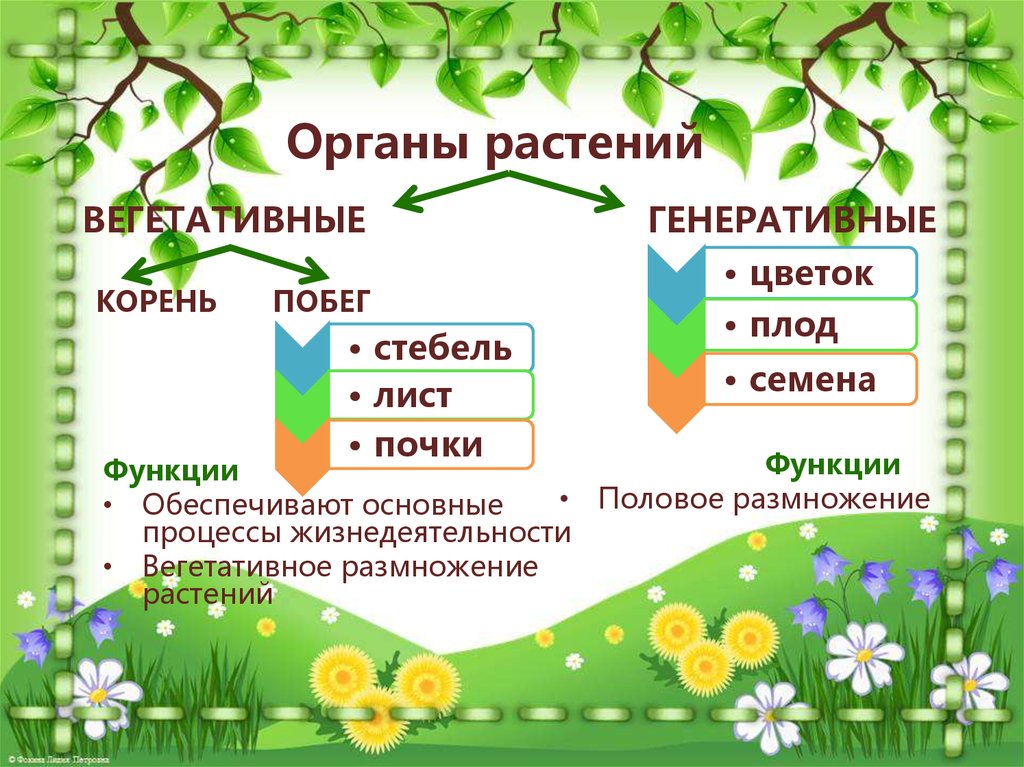
У многих представителей
царства растений функций питания
и размножения распределены между
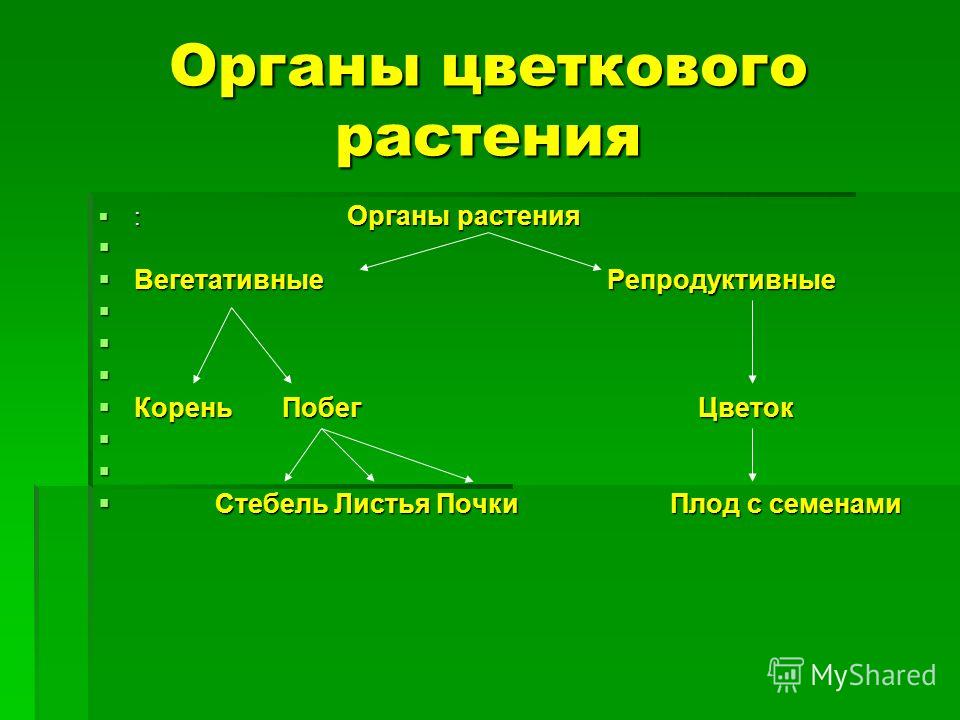
разными органами растения. Органы
растения, выполняющие функции питания
и обмена веществ с окружающей
средой, называют вегетативными (лат. «вегетабилис»
– растительный). Все вместе они образуют
вегетативное тело растения.
Высшие растения
имеют вегетативное тело, разделенное
на 2 основные части: корень и побег,
которые помогают растению осваивать
две разные среды для добычи пищи
– почвенную и воздушную. Корень
и побег – главные вегетативные
органы растения.
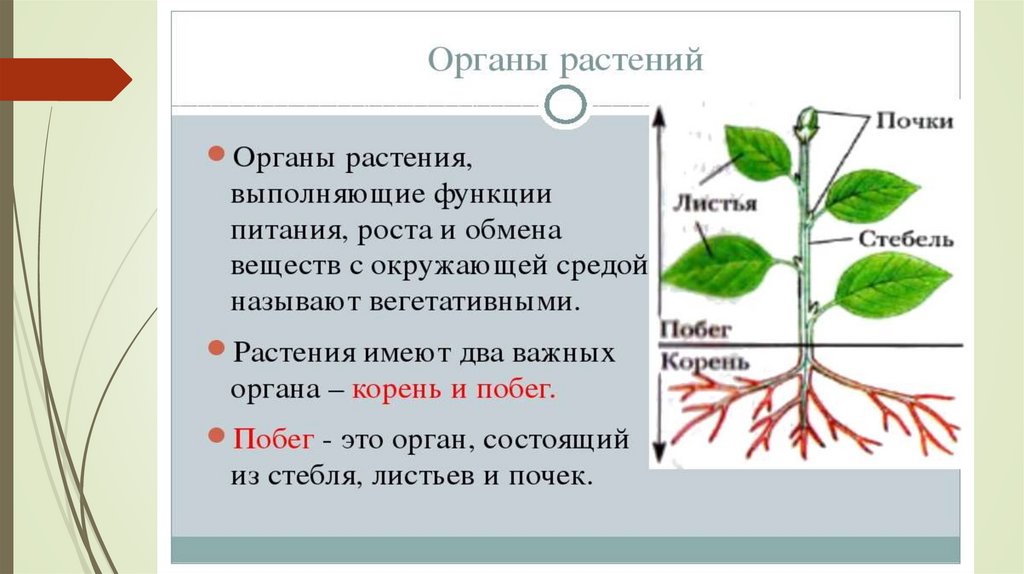
Побег в отличие
от корня сложный орган. В нем
различают стебель, листья и почки.
Вот почему побег можно рассматривать
как особую систему, состоящую из
отдельных, но взаимосвязанных частей.
Хотя стебель, листья и почки –
это части побега, их часто тоже
называют органами растения.
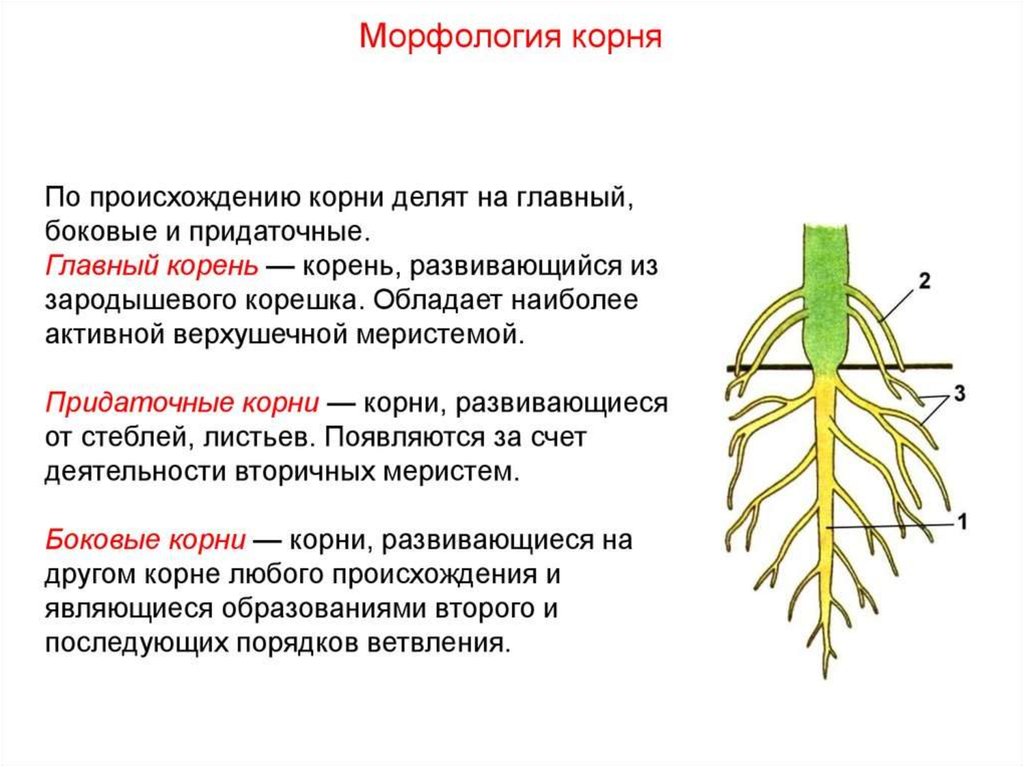
Корень
Ко́рень (лат.
radix) — осевой, обычно подземный вегетативный
орган высших растений (сосудистых растений),
обладающий неограниченным ростом в длину
и положительным геотропизмом. Корень
осуществляет закрепление растения в
почве и обеспечивает поглощение и проведение
воды с растворёнными минеральными веществами
к стеблю и листьям.
На корне
нет листьев, в клетках корня
нет хлоропластов.
Кроме основного
корня, многие растения имеют боковые
и придаточные корни. Совокупность
всех корней растения называют корневой
системой. В случае, когда главный
корень незначительно выражен, а
придаточные корни выражены значительно,
корневая система называется мочковатой.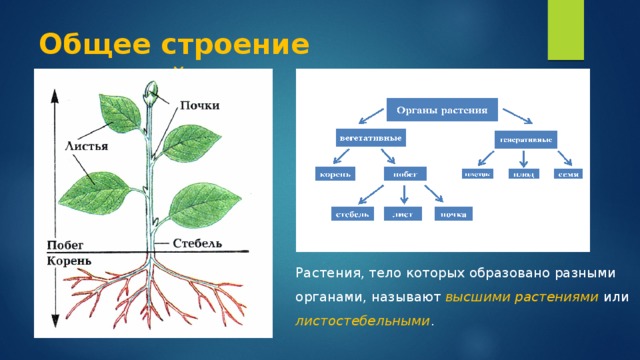
Если главный корень выражен значительно,
корневая система называется стержневой.
Некоторые растения
откладывают в корне запасные
питательные вещества, такие образования
называют корнеплодами.
Основные
функции корня
- Закрепление растения в субстрате.
- Всасывание, проведение воды и минеральных веществ.
- Запас питательных веществ в главном корне.
- Взаимодействие с корнями других растений (симбиоз), грибами, микроорганизмами, обитающими в почве (микориза, клубеньки представителей семейства Бобовые).
- Вегетативное размножение.
- Синтез биологически активных веществ.
У многих растений
корни выполняют особые функции
(воздушные корни, корни-присоски).
Происхождение
корня
Тело первых
вышедших на сушу растений ещё не было
расчленено на побеги и корни. Оно
состояло из ответвлений, одни из которых
поднимались вертикально, а другие
прижимались к почве и
воду и питательные вещества. Несмотря
Несмотря
на примитивное строение, эти растения
были обеспечены водой и питательными
веществами, так как имели небольшие
размеры и жили около воды.
В ходе дальнейшей
эволюции некоторые ответвления
стали углубляться в почву
и дали начало корням, приспособленным
к более совершенному почвенному
питанию. Это сопровождалось глубокой
перестройкой их структуры и появлением
специализированных тканей. Образование
корней было крупным эволюционным достижением,
благодаря которому растения смогли
осваивать более сухие почвы
и образовывать крупные побеги, поднятые
вверх к свету. Например, у мохообразных
настоящих корней нет, их вегетативное
тело небольших размеров — до 30 см, обитают
мхи во влажных местах. У папоротникообразных
появляются настоящие корни, это приводит
к увеличению размеров вегетативного
тела и к расцвету этой группы в каменноугольный
период.
Видоизменения
и специализация корней
Корни некоторых
строений имеют склонность к метаморфозу.
Видоизменения корней:
- Корнеплод — утолщённый придаточный корень.
 В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка). - Корневые клубни (корневые шишки) образуются в результате утолщения боковых и придаточных корней. С их помощью растение цветёт быстрее.
- Корни-зацепки — своеобразные придаточные корни. При помощи этих корней растение «приклеивается» к любой опоре.
- Ходульные корни — выполняют роль опоры.
- Воздушные корни — боковые корни, растут в надземной части. Поглощают дождевую воду и кислород из воздуха. Образуются у многих тропических растений в условиях повышенной влажности.
- Микориза — сожительство корней высших растений с гифами грибов. При таком взаимовыгодном сожительстве, называемом симбиозом, растение получает от гриба воду с растворёнными в ней питательными веществами, а гриб — органические вещества. Микориза характерна для корней многих высших растений, особенно древесных. Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.
- Бактериальные клубеньки на корнях высших растений — сожительство высших растений с азотфиксирующими бактериями — представляют собой видоизменённые боковые корни, приспособленные к симбиозу с бактериями. Бактерии проникают через корневые волоски внутрь молодых корней и вызывают у них образование клубеньков. При таком симбиотическом сожительстве бактерии переводят азот, содержащийся в воздухе, в минеральную форму, доступную для растений. А растения, в свою очередь, предоставляют бактериям особое местообитание, в котором отсутствует конкуренция с другими видами почвенных бактерий. Бактерии также используют вещества, находящиеся в корнях высшего растения. Чаще других бактериальные клубеньки образуются на корнях растений семейства Бобовые. В связи с этой особенностью семена бобовых богаты белком, а представителей семейства широко используют в севообороте для обогащения почвы азотом.
- Дыхательные корни — у тропических растений — выполняют функцию дополнительного дыхания.
 В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).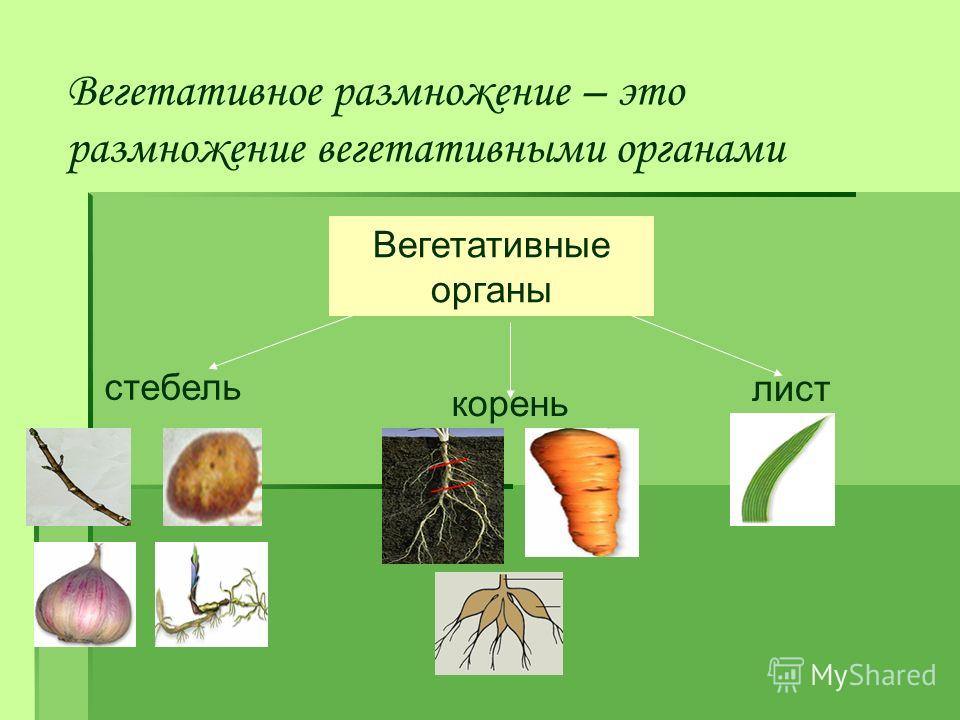 Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.
Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.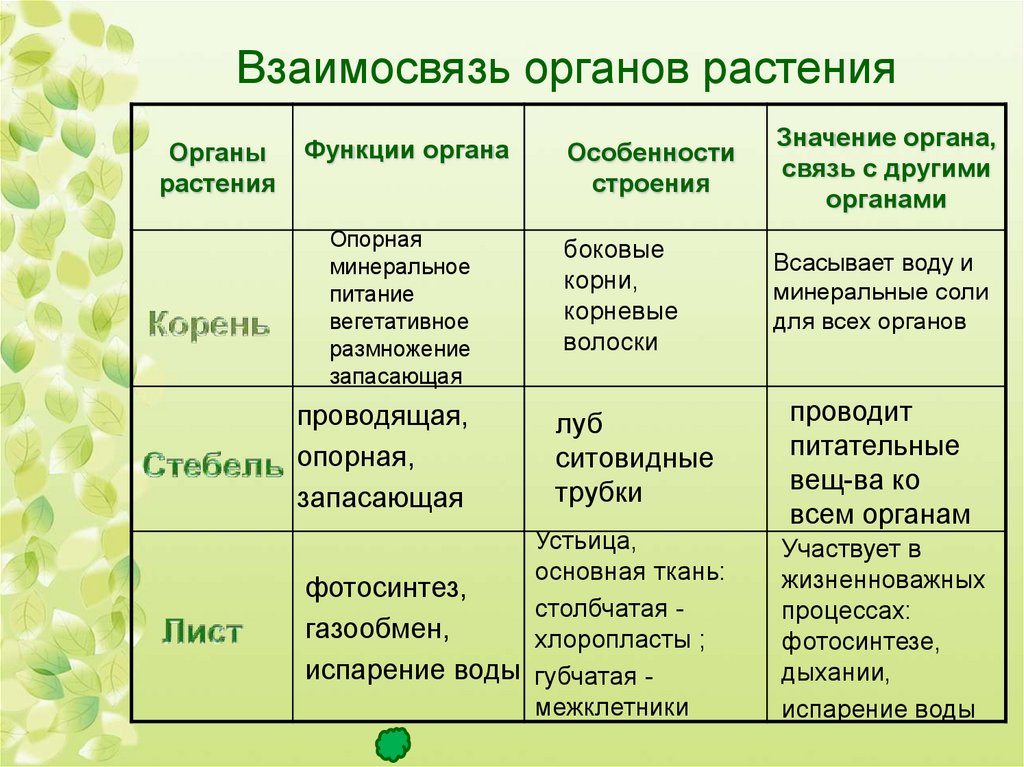
Особенности
строения корней
Совокупность
корней одного растения называют корневой
системой.
В состав корневых
систем входят корни различной природы.
Различают:
- главный корень,
- боковые корни,
- придаточные корни.
Главный корень
развивается из зародышевого корешка.
Боковые корни возникают на любом
корне в качестве бокового ответвления.
Придаточные корни образованы побегом
и его частями.
Части
корня
- Точка роста. Участок непосредственно за кончиком корня, где клетки делятся, обеспечивая новый рост.
- Зона растяжения. Участок новых клеток, образованных в точке роста и расположенных непосредственно за ней. Клетки, пока их клеточные стенки не станут жёсткими, растягиваются в длину при всасывании воды. Это растяжение толкает кончик корня дальше в почву.
- Слой корневых волосков. Наиболее молодая часть эпидермиса или внешней кожицы корня. Это зона, где образуются корневые волоски. Она расположена непосредственно за зоной растяжения. Как только стенки вытянувшихся клеток становятся жёстче, самые крайние из них превращаются в слой корневых волосков. Более старая часть этого слоя (выше по корню) медленно отделяется, замещаясь слоем жёстких клеток, входящих в экзодерму (самый наружный слой коры).
- Корневые волоски. Длинные выросты клеток слоя корневых волосков. Они всасывают воду и минеральные вещества.
- Корневой чехлик. Слой клеток, защищающих кончик корня при росте.
 Наиболее молодая часть эпидермиса или внешней кожицы корня. Это зона, где образуются корневые волоски. Она расположена непосредственно за зоной растяжения. Как только стенки вытянувшихся клеток становятся жёстче, самые крайние из них превращаются в слой корневых волосков. Более старая часть этого слоя (выше по корню) медленно отделяется, замещаясь слоем жёстких клеток, входящих в экзодерму (самый наружный слой коры).
Наиболее молодая часть эпидермиса или внешней кожицы корня. Это зона, где образуются корневые волоски. Она расположена непосредственно за зоной растяжения. Как только стенки вытянувшихся клеток становятся жёстче, самые крайние из них превращаются в слой корневых волосков. Более старая часть этого слоя (выше по корню) медленно отделяется, замещаясь слоем жёстких клеток, входящих в экзодерму (самый наружный слой коры).Типы
корневых систем
- В стержневой корневой системе главный корень сильно развит и хорошо заметен среди других корней (характерна для двудольных). Разновидность стержневой корневой системы — ветвистая корневая система: состоит из нескольких боковых корней, среди которых не различают главный корень; характерна для деревьев.
- В мочковатой корневой системе на ранних этапах развития главный корень, образованный зародышевым корешком, отмирает, а корневая система составляется придаточными корнями (характерна для однодольных). Стержневая корневая система проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая система лучше оплетает прилегающие частицы грунта.
- Придаточные корни растут непосредственно из стебля. Они отрастают от луковицы (представляющей собой особый стебель) или от садовых черенков.
- Воздушные корни. Корни, которые растут от стебля. но не проникают в землю. Они используются лазящими растениями для закрепления, как например, у плюща.
- Опорные (ходульные) корни. особый тип воздушных корней. Они отрастают от стебля и затем проникают в землю, которая может быть покрыта водой. Они поддерживают тяжёлые растения, например мангры.
 Стержневая корневая система проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая система лучше оплетает прилегающие частицы грунта.
Стержневая корневая система проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая система лучше оплетает прилегающие частицы грунта.Зоны
молодого корневого окончания
Различные части
корня выполняют неодинаковые функции
и различаются по внешнему виду.
Эти части получили название зон.
Кончик корня
снаружи всегда прикрыт корневым
чехликом, защищающим нежные клетки меристемы.
Чехлик состоит из живых клеток,
которые постоянно обновляются.
Клетки корневого чехлика выделяют
слизь, она покрывает поверхность
молодого корня. Благодаря слизи
снижается трение о почву, её частицы
легко прилипают к корневым окончаниям
и корневым волоскам. В редких случаях
корни лишены корневого чехлика
(водные растения, некоторые растения-паразиты).
Под чехликом располагается зона
деления, представленная образовательной
тканью — меристемой. Если эта апикальная
меристема обособлена и образует
только клетки корневого чехлика (как
у большинства однодольных
её называют калиптрогеном. У большинства
двудольных меристематическая ткань кончика
корня сливается с меристемой, образующей
зону всасывания, и называется дерматокалиптрогеном.[источник
не указан 797 дней]
Клетки зоны
деления тонкостенные и заполнены
цитоплазмой, вакуоли отсутствуют.
Зону деления можно отличить на живом
корешке по желтоватой окраске, длина
её около 1 мм. Вслед за зоной деления
располагается зона растяжения. Она
также невелика по протяжённости: составляет
всего несколько миллиметров, выделяется
светлой окраской и как бы прозрачна.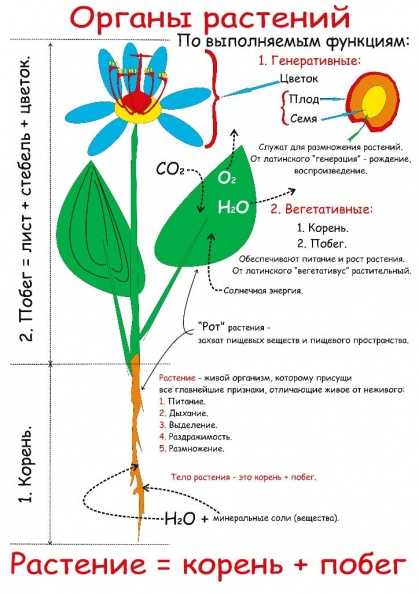
Клетки зоны роста уже не делятся,
но способны растягиваться в продольном
направлении, проталкивая корневое
окончание вглубь почвы. В пределах
зоны роста происходит разделение клеток
на ткани.
Окончание зоны
роста хорошо заметно по появлению
многочисленных корневых волосков. Корневые
волоски располагаются в зоне
всасывания, функция которой понятна
из её названия. Длина её от нескольких
миллиметров до нескольких сантиметров.
В отличие от зоны роста участки
этой зоны уже не смещаются относительно
частиц почвы. Основную массу воды и
питательных веществ молодые
корни всасывают с помощью
корневых волосков.
Корневые
волоски — это выросты клеток
поверхностной ткани поглощающей зоны
корня растения. Они увеличивают всасывающую
поверхность корня, выделяют продукты
обмена; находятся чуть выше корневого
чехлика. Все вместе они создают впечатление
белого пушка вокруг корня. У растения,
только что вынутого из почвы, всегда можно
увидеть прилипшие к корневым волоскам
комочки почвы. Они содержат слой протоплазмы,
Они содержат слой протоплазмы,
ядро, крупную вакуоль; их тонкие, легко
проницаемые для воды оболочки плотно
склеиваются с комочками почвы. Корневые
волоски выделяют в почву различные вещества.
Длина варьируется у разных видов растений
от 0,06 до 10 мм. С увеличением влажности
почвы образование замедляется; не образуются
они и в очень сухой почве.
Этап урока | Деятельность учителя | Деятельность ученика | Результат | Планируемые результаты |
| 1. Организационный (1-2 мин.) | Приветствие, высказывает добрые пожелания детям; | Приветствие. Организация учащимися рабочего места. | Организация учащимися рабочего места. | Метапредметные: умение организовывать рабочее место, самоорганизовываться. Личностные: ответственное отношение к учению, самодисциплина. |
| 2. Актуализация знаний (10 мин.) | 1. Проверка домашнего задания (устный опрос) 2. Актуализация знаний 3. Постановка учебной задачи 4. Открытие новых знаний – Давайте вспомним что такое царство? – Как называются все растения вместе взятые? (Слайд 3) – А что характерно для царства растений? – Из чего состоит цветковое растение? Проблемный вопрос: Могут ли растения жить без органов? Сформулируйте тему урока. Некоторые растения имеют на побегах специальные образования, в которых развиваются особые мелкие клетки — споры, с помощью которых растения (водоросли, мхи, папоротники) размножаются вегетативным (бесполым) путем и расселяются по земной поверхности. Кроме вегетативных органов растения имеют специальные органы, служащие для полового размножения. Их называют половыми, пли генеративными, органами (от лат. 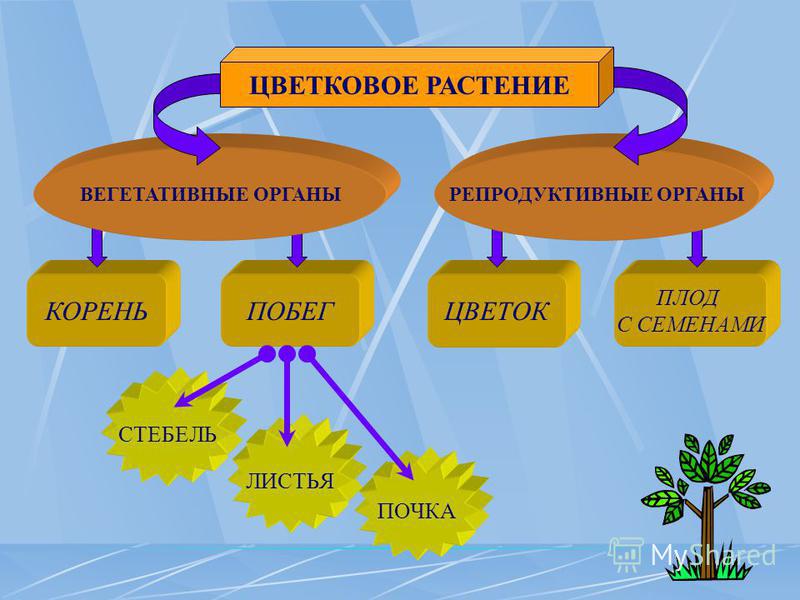 Генераре — «рожать», «производить»). К генеративным органам относят, например, цветок, образующийся из него плод и заключенные в нем семена. Генераре — «рожать», «производить»). К генеративным органам относят, например, цветок, образующийся из него плод и заключенные в нем семена. Чем отличаются рассмотренные вами гербарии? | Мир растений огромен и удивителен. Органы растений — это части организма растения, выполняющие в нем определенные функции и имеющие особое строение. У растений обычно различают органы, выполняющие функции питания и функции размножения. Органы растения, выполняющие функции питания, обмена веществ с окружающей средой и роста, называют вегетативными (от лат. Вегетабилис – «растительный»). Все вместе они образуют вегетативное тело растения. Высшие растения имеют вегетативное тело, разделенное на две основные части: корень и побег, которые помогают растению осваивать две разные среды для получения пищи — почвенную и воздушную. Корень и побег — главные вегетативные органы растения. Побег — это орган, в котором различают стебель, листья и почки. Вот почему побег можно рассматривать как особую систему, состоящую из отдельных, но взаимосвязанных частей. Хотя стебель, листья и почки — это части побега, их часто тоже называют органами растения. Растения с помощью отдельных частей вегетативных органов могут осуществлять рост и вегетативное размножение. Среди растений есть такие, тело которых не расчленено на отдельные органы (от греч. органон — «орудие», «инструмент»). Поэтому их называют низшими растениями. К низшим растениям относятся, например, водоросли). Вот почему побег можно рассматривать как особую систему, состоящую из отдельных, но взаимосвязанных частей. Хотя стебель, листья и почки — это части побега, их часто тоже называют органами растения. Растения с помощью отдельных частей вегетативных органов могут осуществлять рост и вегетативное размножение. Среди растений есть такие, тело которых не расчленено на отдельные органы (от греч. органон — «орудие», «инструмент»). Поэтому их называют низшими растениями. К низшим растениям относятся, например, водоросли). Растения, тело которых образовано органами (прежде всего стеблями с листьями), называют высшими растениями, или листостебельными. К ним относятся мхи, папоротники, хвощи, плауны, семенные растения. | Сообщение учащихся – органы растений у растений с.Загарино: лиственница, сосна обыкновенная, ель голубая
Просмотр видеоурока «Растения» | Регулятивные УУД: Коммуникативные УУД: Личностные УУД: Познавательные УУД: |
3. Создание проблемной ситуации (20 мин. ) ) | Лабораторная работа. Инструктаж по ТБ работы с гербарным материалом.
В тетрадях запишите тему, цель и оборудование работы. Перед вами два растения: одно цветковое, другое хвойное (показываю черенки сосны и березы) (Слайд8). Они совершенно разные. Что вам сразу бросилось в глаза? У сосны нет листьев, но есть хвоинки. Сейчас осень, поэтому листьев нет и на цветковом растении, но мы знаем, что они были. А куда они делись? Растение сбросило их, готовясь к зиме, и это явление называется листопад. Но мы можем посчитать сколько листьев было на этом побеге. А сделаем мы это по листовым следам (показываю). Кроме того, хвойное растение отличается от лиственного наличием шишки. Посмотрите где она расположена. Это орган размножения растения, в нем содержатся семена. А у лиственного растения органом размножения является цветок. Вы хорошо поработали, а теперь время немного отдохнуть! (физкультпауза)(Слайд 10) | Изучают инструкцию по выполнению лабораторной работы Делают необходимые записи в тетрадях 5. Заполните схему. | Формулируют тему и цели урока. Тема: «Органы растений». Выполняют лабораторную работу. Только у покрытосеменных присутствуют все органы. У водорослей органов нет.  Знания об уровневой организации растений. Знания о вегетативных и генеративных органах, их значении для растений. Умение выполнять лабораторную работу, работать по инструкции, умение определять, находить и обозначать на схематическом рисунке органы растений. Определять к какой группе органов они относятся. Знание правил оформления лабораторной работы. | Предметные: комментировать содержание рисунка, предлагающего использование имеющихся знаний в новой ситуации Регулятивные: Познавательные: Коммуникативные: Личностные: |
| 4. Закрепление нового материала (7 мин.) | Проводит фронтальный опрос.
Корректирует выводы учащихся. | Подведем итоги по уроку. Отвечают на вопросы Формулируют вывод. Записывают вывод. | Уметь сверять свои действия с целью и при необходимости исправлять ошибки самостоятельно | Предметные: подводить итоги работы, формулировать вывод. Метапредметные: адекватно использовать речевые средства для дискуссии и аргументации своей позиции, отстаивать свою позицию. Личностные: демонстрировать доброжелательное отношение к мнению другого человека. |
| 5. Контроль (5 мин.) | Вставьте пропущенные слова. Часть растения, имеющую определенное строение и выполняющую определенные функции – это … . У растений различают … и … органы. | Вставляют пропуски. Проверяют всем классом, обозначая правильные и неправильные ответы знаками «+» и «–». | Выставляют оценки по следующей шкале: 9-10 плюсов – 5 7-8 плюсов – 4 5-6 плюсов – 3 | Регулятивные УУД: осуществление учебных действий, выполнение заданий в соответствии с поставленной целью, адекватно оценивать полученные в ходе урока знания.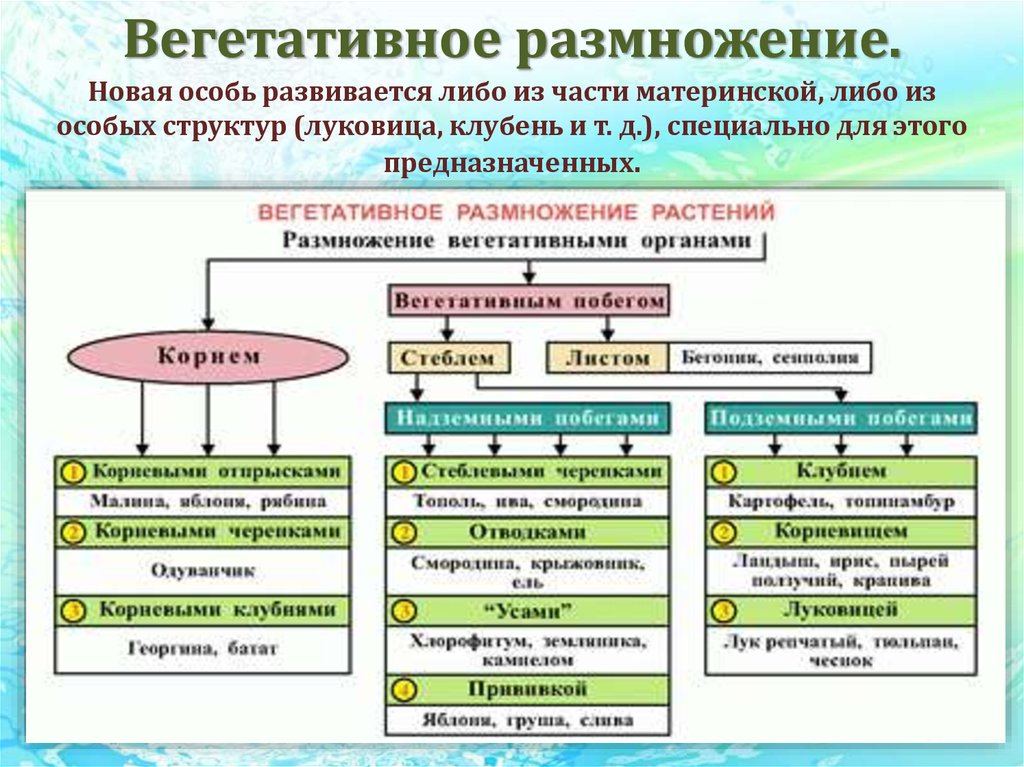 |
| 6. Рефлексия (2 мин.) | – Мы достигли цели урока? – Что нового вы сегодня узнали в ходе урока? – Почему? – Вспомните вашу гипотезу. – Совпала ваша гипотеза с полученным выводом? Ответить на вопросы: | Регулятивные УУД: осуществление учебных действий, выполнение заданий в соответствии с поставленной целью. Познавательные УУД: систематизация информации, формулировка ответов на вопросы учителя Коммуникативные УУД: строить понятные монологические высказывания; умение активно слушать одноклассников. | ||
| 7. Домашнее задание (3 мин.) | Обязательное: параграф учебника 11, термины, пересказ. Дополнительное: прочитать стихотворение Бальмонта К. Д. «Растение». Найдите слова автора, которыми он описывает значение органов для растения.  | Записывают домашнее задание в дневник. | Регулятивные УУД: осуществление учебных действий, выполнение заданий в соответствии с поставленной целью. Познавательные УУД: осуществление поиска и отбора необходимой информации, систематизация информации Коммуникативные УУД: планирование учебного сотрудничества с родителями, формулировка вопросов |
 Инструктивная карточка. Тест. Индивидуальное тестирование-компьютер. Презентация, Приложение 1, Приложение 2, Приложение 3, Приложение 4.
Инструктивная карточка. Тест. Индивидуальное тестирование-компьютер. Презентация, Приложение 1, Приложение 2, Приложение 3, Приложение 4.
 Именно поэтому лиственные растения еще называют цветковыми. Теперь самостоятельно рассмотрите оба растения, зарисуйте в тетради, сделайте соответствующие обозначения (Слайд 9).
Именно поэтому лиственные растения еще называют цветковыми. Теперь самостоятельно рассмотрите оба растения, зарисуйте в тетради, сделайте соответствующие обозначения (Слайд 9).

Вегетативный рост — биология развития
Когда побег появляется из почвы, большая часть плана тела спорофита еще предстоит разработать. показаны основные части зрелого растения-спорофита, которые образуются из меристем.
Рисунок 20.20
Морфология генерализованного покрытосеменного спорофита.
Меристемы
Как уже упоминалось, меристемы представляют собой скопления клеток, которые позволяют повторять и расширять основной рисунок тела, установленный во время эмбриогенеза, после прорастания.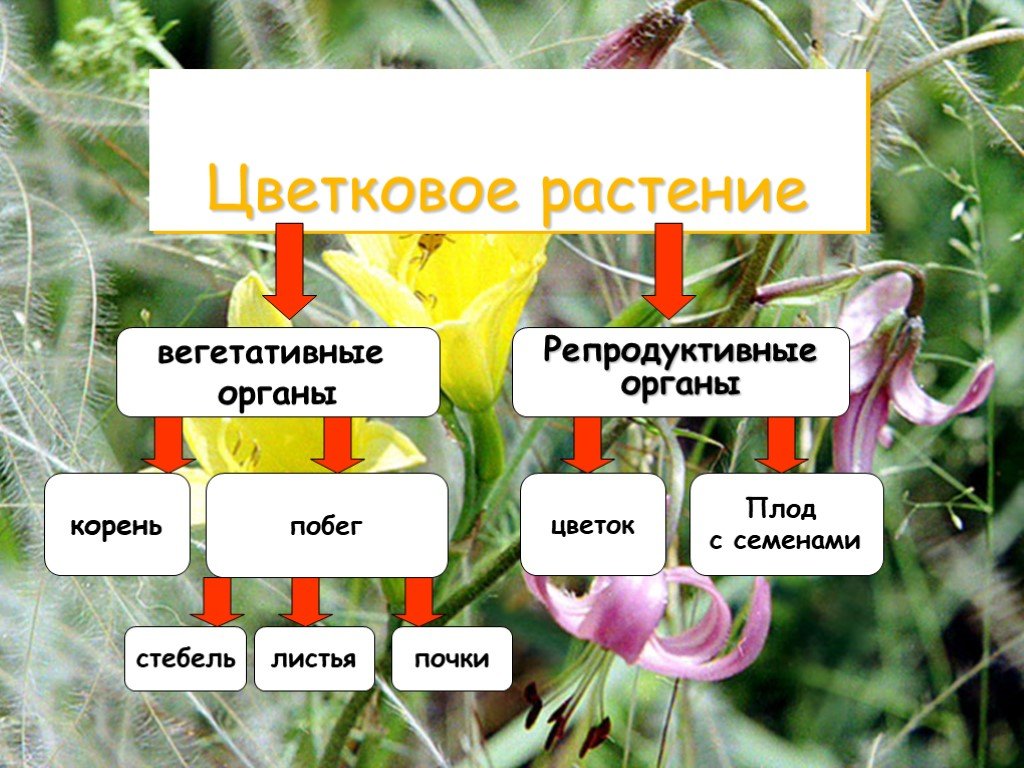 Меристематические клетки сходны со стволовыми клетками животных. * Они делятся, давая одну дочернюю клетку, которая продолжает быть меристемной, и другую, которая дифференцируется. Меристемы делятся на три категории: верхушечные, латеральные и интеркалярные.
Меристематические клетки сходны со стволовыми клетками животных. * Они делятся, давая одну дочернюю клетку, которая продолжает быть меристемной, и другую, которая дифференцируется. Меристемы делятся на три категории: верхушечные, латеральные и интеркалярные.
Апикальные меристемы встречаются на кончиках растущих побегов и корней (). Верхушечные меристемы корня образуют корневой чехлик, который состоит из смазанных клеток, которые отслаиваются, когда меристема проталкивается через почву за счет деления клеток и удлинения более проксимальных клеток. Апикальная меристема корня также дает начало дочерним клеткам, которые производят три системы тканей корня. Новые корневые апикальные меристемы инициируются из ткани в сердцевине корня и выходят через основную ткань и кожную ткань. Корневые меристемы также могут быть получены вторично из стебля растения; в случае кукурузы это основной источник корневой массы.
Рисунок 20.21
Побеги и корневые меристемы. И побеги, и корни развиваются из апикальных меристем, на концах которых сгруппированы недифференцированные клетки. В корнях также образуется корневой чехлик, защищающий меристему при ее прорастании через почву. Боковые органы побега (подробнее…)
И побеги, и корни развиваются из апикальных меристем, на концах которых сгруппированы недифференцированные клетки. В корнях также образуется корневой чехлик, защищающий меристему при ее прорастании через почву. Боковые органы побега (подробнее…)
Верхушечная меристема побега образует стебли, листья и репродуктивные структуры. В дополнение к апикальной меристеме побега, заложенной в эмбриогенезе, в пазухах (углы между листом и стеблем) образуются производные от исходной апикальные меристемы побегов (пазушные почки; см.). В отличие от новых корневых меристем, они возникают из поверхностных слоев меристемы.
Апикальные меристемы покрытосеменных состоят из трех слоев клеток (обозначенных L1, L2 и L3) на поверхности растения (). Одним из способов изучения вклада различных слоев в структуру растения является создание химер. Химеры растений состоят из слоев, имеющих различные генотипы с различимыми маркерами. Когда, например, L2 имеет генотип, отличный от L1 или L3, вся пыльца будет иметь генотип L2, что указывает на то, что пыльца происходит от L2. Химеры также использовались для демонстрации классической индукции у растений, у которых, как и при развитии животных, один слой влияет на путь развития соседнего слоя.
Химеры также использовались для демонстрации классической индукции у растений, у которых, как и при развитии животных, один слой влияет на путь развития соседнего слоя.
Рисунок 20.22
Организация верхушечной меристемы побега. (A) Меристемы покрытосеменных имеют два или три внешних слоя клеток, которые гистологически различны (здесь обозначены L1, L2 и L3). Хотя клетки в определенных слоях, как правило, имеют определенные судьбы, они не обязательно (подробнее…)
Размер апикальной меристемы побега точно контролируется межклеточными сигналами, скорее всего, между слоями меристемы (обзор Doerner 1999) . Мутации в Arabidopsis CLAVATA 9Например, гены 0034 приводят к увеличению размера меристемы и образованию дополнительных органов. † STM имеет противоположный эффект, и двойные мутантные фенотипы согласуются с гипотезой о том, что они работают вместе, чтобы поддерживать размер меристемы (Clark et al. 1996). Возможно, они уравновешивают скорость деления клеток (которая увеличивает меристему) и скорость дифференцировки клеток на периферии меристемы (которая уменьшает размер меристемы) (Meyerowitz 1997).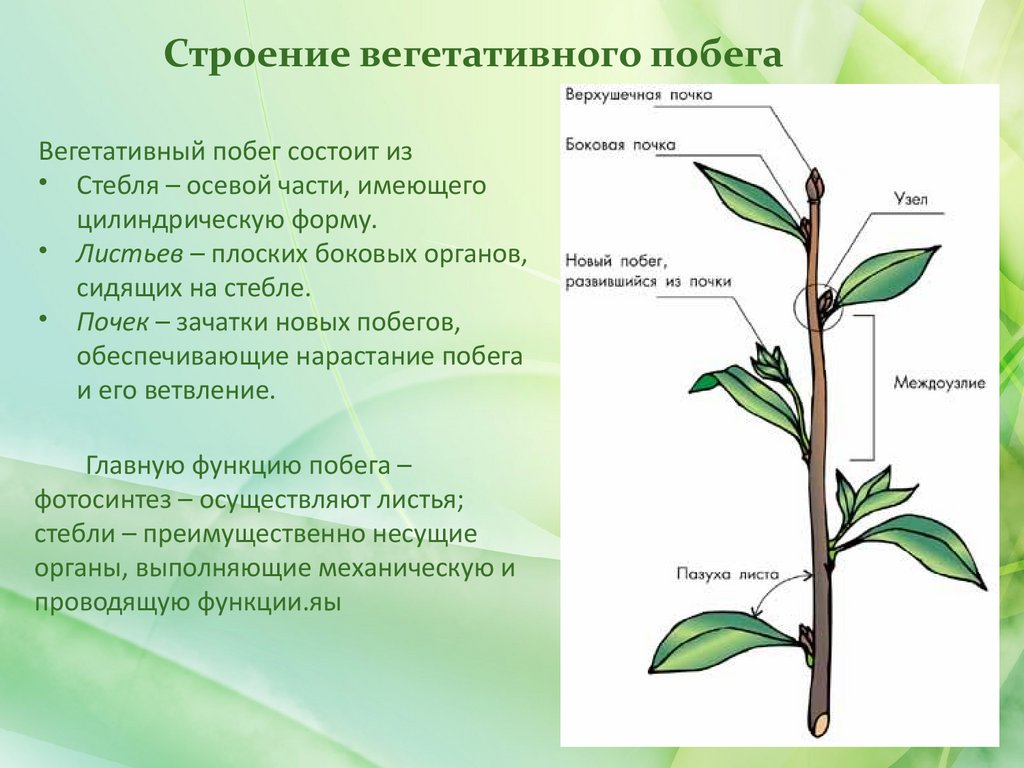
Боковые меристемы представляют собой цилиндрические меристемы, обнаруживаемые в побегах и корнях, которые приводят к вторичному росту (увеличению диаметра стебля и корня за счет образования сосудистых тканей). Стебли однодольных не имеют боковых меристем, но часто имеют интеркалярных меристем , встроенных в стебли между зрелыми тканями. Хлопающий звук, который вы можете услышать на кукурузном поле летней ночью, на самом деле вызван быстрым увеличением длины стебля за счет интеркалярных меристем.
Развитие корня
Формирование радиального и осевого паттерна в корнях начинается во время эмбриогенеза и продолжается на протяжении всего развития по мере роста первичного корня и появления боковых корней из клеток перицикла глубоко внутри корня. Эксперименты по лазерной абляции, устраняющие отдельные клетки, и клональный анализ продемонстрировали, что клетки пластичны и что их положение является основной детерминантой судьбы в раннем развитии корней. Анализ мутантов радиальной организации корней выявил гены со специфичной для слоя активностью (Scheres et al. 19).95; Шерес и Хейдстра, 1999). Мы проиллюстрируем эти результаты, рассмотрев два гена Arabidopsis , которые регулируют судьбу основной ткани.
Анализ мутантов радиальной организации корней выявил гены со специфичной для слоя активностью (Scheres et al. 19).95; Шерес и Хейдстра, 1999). Мы проиллюстрируем эти результаты, рассмотрев два гена Arabidopsis , которые регулируют судьбу основной ткани.
У дикого типа Arabidopsis, имеется два слоя основной ткани корня. Внешний слой становится корой, а внутренний слой становится эндодермой, которая образует трубку вокруг ядра сосудистой ткани. Гены SCARECROW ( SCR ) и SHORT-ROOT ( SHR ) имеют мутантные фенотипы с одним, а не с двумя слоями корневой ткани (Benfey et al. 19).93). Ген SCR необходим для асимметричного клеточного деления в начальном слое клеток с образованием энтодермальной клетки меньшего размера и клетки коры большего размера (10). Мутант scr экспрессирует маркеры как для клеток коры, так и для клеток энтодермы, указывая на то, что дифференцировка прогрессирует в отсутствие клеточного деления (Di Laurenzio 1996).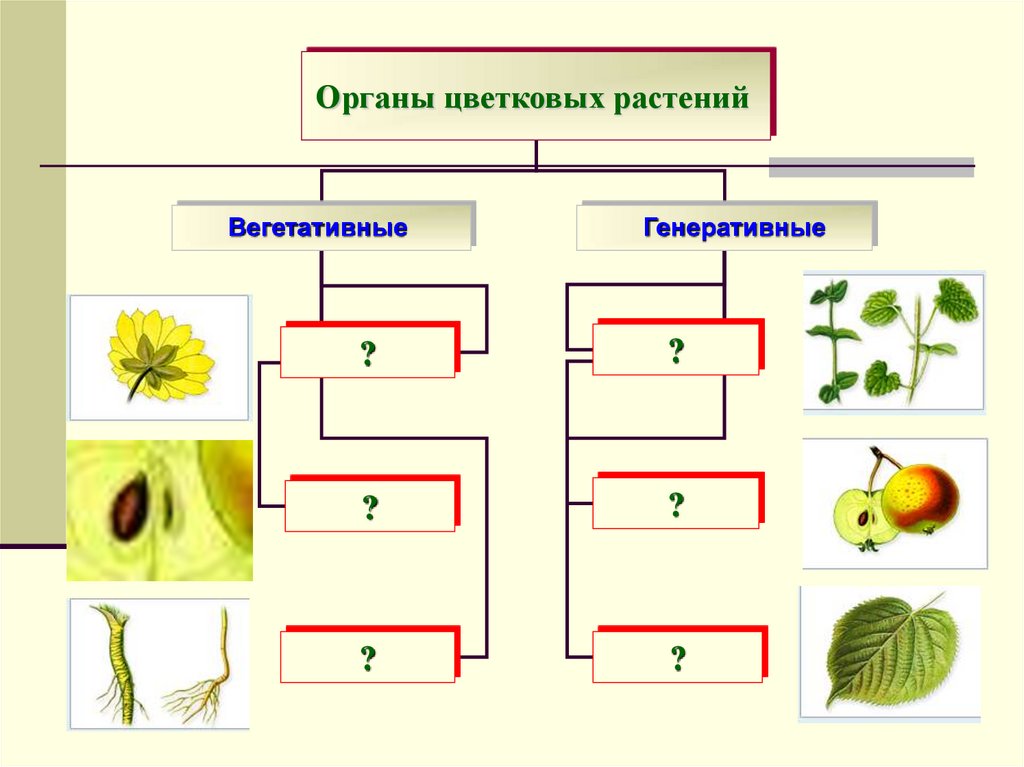 SHR отвечает за спецификацию эндодермальных клеток. Клетки мутанта shr не имеют энтодермальных признаков.
SHR отвечает за спецификацию эндодермальных клеток. Клетки мутанта shr не имеют энтодермальных признаков.
Рисунок 20.23
SCR и SHR регулируют энтодермальную дифференцировку в радиальном развитии корня. (A) Схема нормального деления клеток с образованием клеток коры и энтодермы. SCR регулирует это асимметричное деление клеток. (B, C) Экспрессия SCR в корнях и побегах. Промотор SCR (подробнее…)
Формирование осевого паттерна в корнях может зависеть от морфогена, параллельно с некоторыми аспектами развития животных. Различные эксперименты установили, что распределение растительного гормона ауксин организует осевой рисунок. Пик концентрации ауксина на кончике корня должен восприниматься как нормальный осевой паттерн (Sabatini et al. 1999).
Как обсуждалось ранее, были идентифицированы отдельные гены, определяющие формирование меристем корней и побегов; однако развитие корней и побегов может иметь общие группы генов, которые регулируют клеточную судьбу и формирование паттерна (Benfey 1999).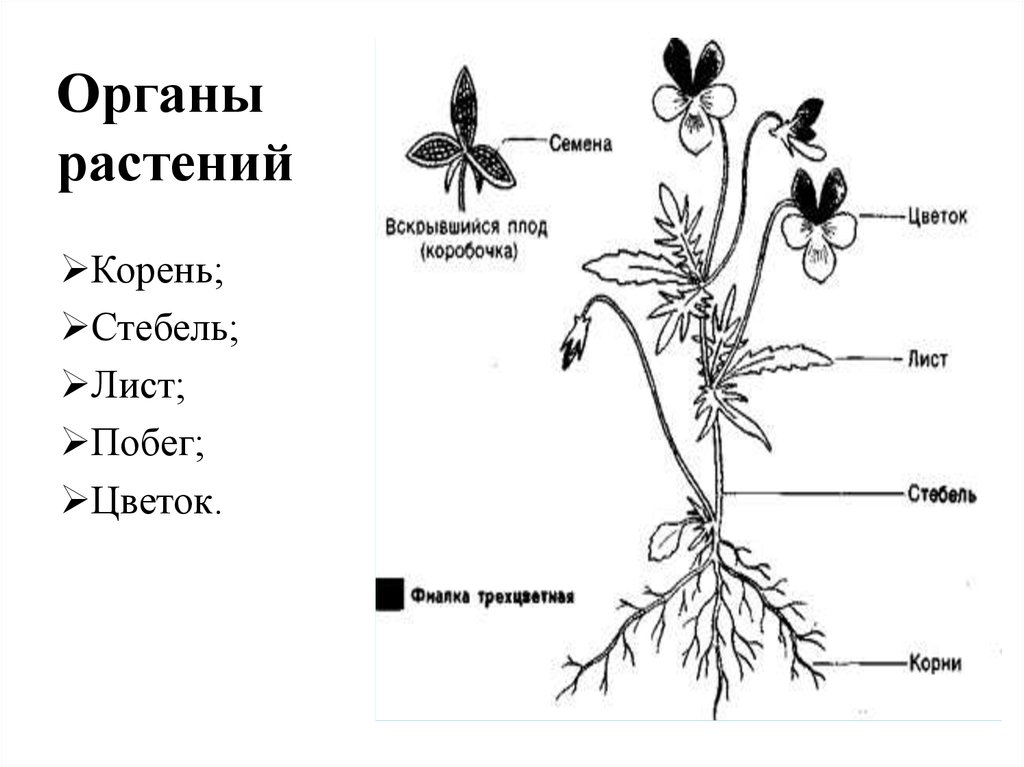 По-видимому, это относится к генам SCR и SHR . В побеге эти гены необходимы для нормального гравитропного ответа, который зависит от нормального образования эндодермы (дефект у мутантов обоих генов; см.). Важно иметь в виду, что существует ряд шагов между установлением основного паттерна и преобразованием этого паттерна в анатомическую и морфологическую структуру. Раскрытие основных механизмов контроля, вероятно, будет наиболее продуктивной стратегией в понимании того, как развиваются корни и побеги.
По-видимому, это относится к генам SCR и SHR . В побеге эти гены необходимы для нормального гравитропного ответа, который зависит от нормального образования эндодермы (дефект у мутантов обоих генов; см.). Важно иметь в виду, что существует ряд шагов между установлением основного паттерна и преобразованием этого паттерна в анатомическую и морфологическую структуру. Раскрытие основных механизмов контроля, вероятно, будет наиболее продуктивной стратегией в понимании того, как развиваются корни и побеги.
Развитие побегов
Уникальная надземная структура различных видов растений берет свое начало в меристемах побегов. Архитектура побегов зависит от количества отростков пазушных почек. Характер ветвления регулируется верхушкой побега — явление, называемое верхушечным доминированием, и, по-видимому, за это ответственны растительные гормоны. Ауксин вырабатывается молодыми листьями и транспортируется к основанию листа. Может подавлять рост пазушных почек. Выпас и цветение часто освобождают почки от верхушечного доминирования, и в это время происходит ветвление.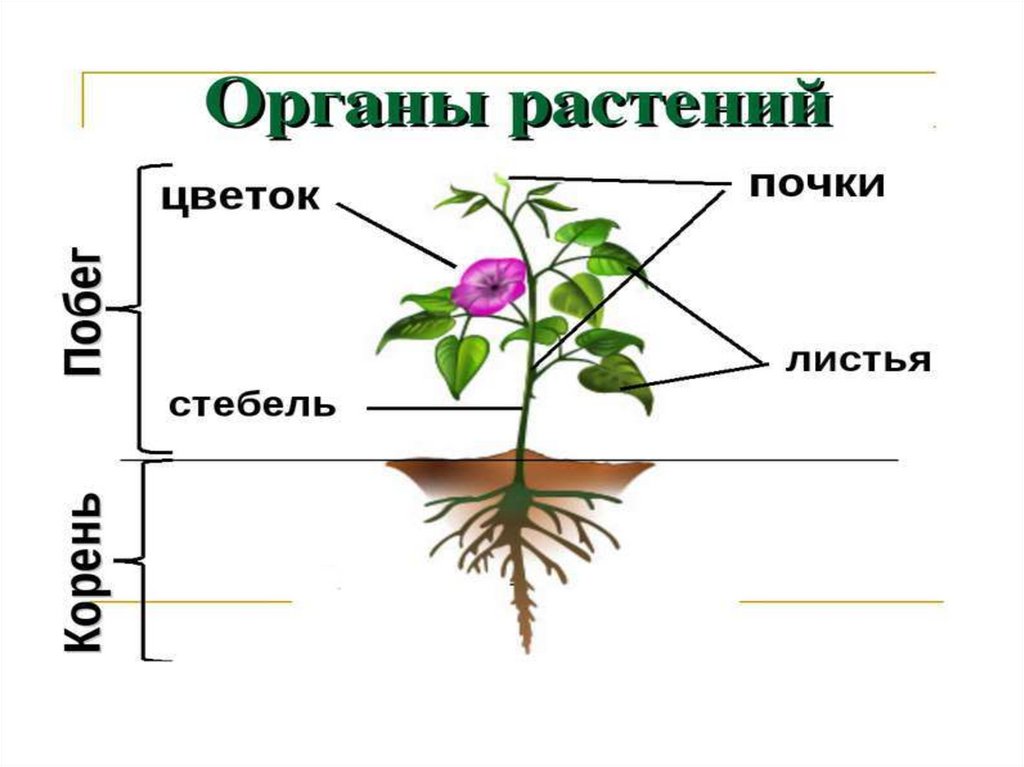 Цитокинины также могут освобождать почки от апикального доминирования. Подмышечные почки могут инициировать свои собственные пазушные почки, поэтому схемы ветвления могут быть довольно сложными. Шаблоны ветвления могут регулироваться сигналами окружающей среды, так что обширный навес на открытой местности максимально улавливает свет. Асимметричные кроны деревьев образуются, когда два дерева растут очень близко друг к другу. В дополнение к пластичности окружающей среды архитектура побегов регулируется генетически. У нескольких видов в настоящее время идентифицированы гены, которые регулируют паттерны ветвления.
Цитокинины также могут освобождать почки от апикального доминирования. Подмышечные почки могут инициировать свои собственные пазушные почки, поэтому схемы ветвления могут быть довольно сложными. Шаблоны ветвления могут регулироваться сигналами окружающей среды, так что обширный навес на открытой местности максимально улавливает свет. Асимметричные кроны деревьев образуются, когда два дерева растут очень близко друг к другу. В дополнение к пластичности окружающей среды архитектура побегов регулируется генетически. У нескольких видов в настоящее время идентифицированы гены, которые регулируют паттерны ветвления.
Зачатки листьев (скопления клеток, которые образуют листья) закладываются на периферии меристемы побега (см. ). Соединение листа и стебля называется узлом , а ткань стебля между узлами называется междоузлием (см. ). Проще говоря, зрелый спорофит создается путем сложения узлов и междоузлий вместе. Филлотаксия , расположение листьев на стебле, включает связь между существующими и вновь образующимися листовыми зачатками.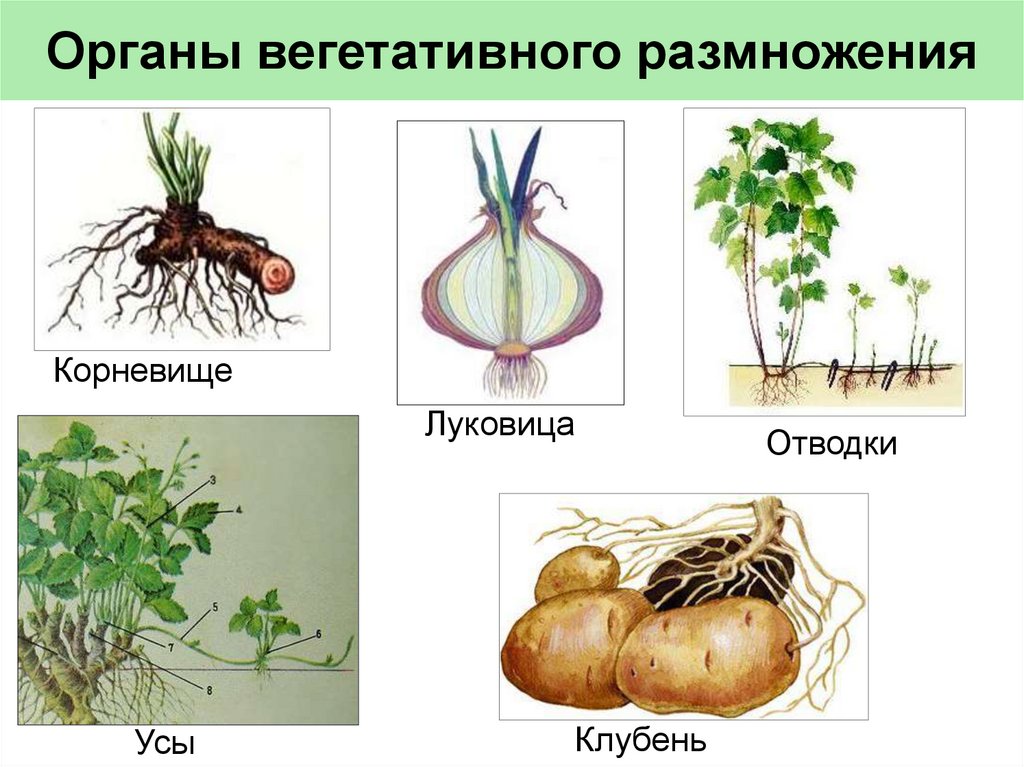 Листья могут располагаться по разным схемам, в том числе по спирали, 180-градусному чередованию одиночных листьев, пар и мутовок из трех и более листьев в узле (Жан и Барабе 19).98). Эксперименты выявили ряд механизмов для поддержания геометрически правильного расстояния между листьями на растении, включая химические и физические взаимодействия новых листовых зачатков с верхушкой побега и с существующими зачатками (Steeves and Sussex, 1989).
Листья могут располагаться по разным схемам, в том числе по спирали, 180-градусному чередованию одиночных листьев, пар и мутовок из трех и более листьев в узле (Жан и Барабе 19).98). Эксперименты выявили ряд механизмов для поддержания геометрически правильного расстояния между листьями на растении, включая химические и физические взаимодействия новых листовых зачатков с верхушкой побега и с существующими зачатками (Steeves and Sussex, 1989).
Неясно, как запускается конкретный образец филлотаксии. Описательные математические модели могут воспроизводить наблюдаемые закономерности, но ничего не раскрывают о механизме. Биофизические модели (например, эффектов стресса/деформации на отложение материала клеточной стенки, что влияет на деление и удлинение клеток) пытаются восполнить этот пробел. Подходы генетики развития многообещающи, но идентифицировано несколько филлотактических мутантов. Один кандидат — 9Мутант 0033 терминального початка кукурузы с нерегулярной филлотаксией. Ген дикого типа экспрессируется в области подковообразной формы с промежутком, где будет инициироваться лист (Veit et al. 1998). Плоскость подковы перпендикулярна оси стебля.
1998). Плоскость подковы перпендикулярна оси стебля.
Развитие листа
Развитие листа включает в себя стремление стать листом, образование оси листа и морфогенез, приводящий к огромному разнообразию форм листа. Эксперименты с культурами оценивали, когда зачатки листьев становятся определяемыми для развития листьев. Исследования папоротников и покрытосеменных показывают, что самые молодые видимые зачатки листьев не предназначены для образования листа; скорее, эти молодые зачатки могут развиваться как побеги в культуре (Стивс 19).66; Смит 1984). Программирование развития листьев происходит позже. Радиальная симметрия зачатка листа становится дорсально-вентральной или уплощенной у всех листьев. Также устанавливаются две другие оси, проксимально-дистальная и латеральная. Уникальная форма листьев является результатом регуляции клеточного деления и расширения клеток по мере развития листовой пластинки. В некоторых случаях избирательная гибель клеток (апоптоз) участвует в формировании листа, но дифференциальный рост клеток, по-видимому, является более распространенным механизмом (Gifford and Foster 19). 89).
89).
Листья делятся на две категории: простые и сложные (см. обзор Sinha 1999). Существует большое разнообразие простой формы листьев, от листьев с гладкими краями до листьев дуба с глубокими лопастями. Сложные листья состоят из отдельных листочков (а иногда и усиков), а не из одной листовой пластинки. Вопрос о том, развиваются ли простые и сложные листья по одному и тому же механизму, остается открытым. Одна точка зрения состоит в том, что сложные листья представляют собой простые листья с большими лопастями. Альтернативная точка зрения состоит в том, что сложные листья представляют собой видоизмененные побеги. Считается, что предковое состояние семенных растений сложное, а у покрытосеменных — простое. Сложные листья возникали несколько раз у покрытосеменных, и неясно, являются ли они возвратом к исходному состоянию.
Рисунок 20.24
Простые и сложные листья.
Генетические подходы к развитию применяются к морфогенезу листьев. Гены класса I KNOX представляют собой гомеобоксные гены, которые включают STM и ген KNOTTED 1 ( KN1 ) кукурузы.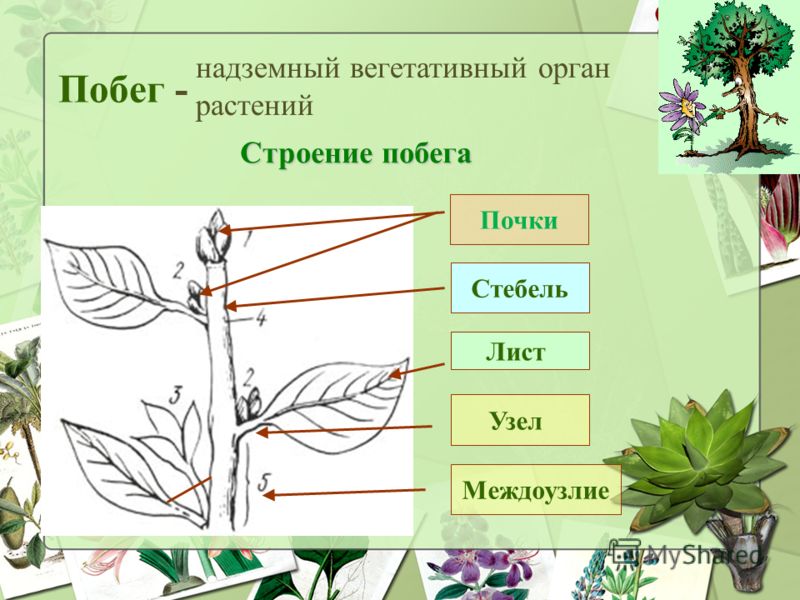 Мутации с приобретением функции KN1 вызывают образование меристемоподобных бугорков на листьях кукурузы. У растений дикого типа этот ген экспрессируется в меристемах. Когда KN1, или томатный гомолог Промотор LeT6, заменен промотором вируса мозаики цветной капусты и вставлен в геном томата, ген экспрессируется на высоком уровне по всему растению, и листья становятся «суперсоставными» (; Hareven et al. 1996). ; Янссен и др., 1998). Простые листья становятся более лопастными (но не составными) в ответ на сверхэкспрессию KN1, , что согласуется с гипотезой о том, что сложные листья могут быть крайним случаем лопастности простых листьев (Jackson 1996). Роль KN1 в меристеме побегов и развитии листьев, однако, согласуется с гипотезой о том, что сложные листья представляют собой видоизмененные побеги.
Мутации с приобретением функции KN1 вызывают образование меристемоподобных бугорков на листьях кукурузы. У растений дикого типа этот ген экспрессируется в меристемах. Когда KN1, или томатный гомолог Промотор LeT6, заменен промотором вируса мозаики цветной капусты и вставлен в геном томата, ген экспрессируется на высоком уровне по всему растению, и листья становятся «суперсоставными» (; Hareven et al. 1996). ; Янссен и др., 1998). Простые листья становятся более лопастными (но не составными) в ответ на сверхэкспрессию KN1, , что согласуется с гипотезой о том, что сложные листья могут быть крайним случаем лопастности простых листьев (Jackson 1996). Роль KN1 в меристеме побегов и развитии листьев, однако, согласуется с гипотезой о том, что сложные листья представляют собой видоизмененные побеги.
Рисунок 20.25
Сверхэкспрессия генов класса 1 KNOX у томата. На фотографии показаны отдельные листья (A) растения дикого типа, (B) мутантного мышиного уха с повышенной сложностью листа и (C) трансгенного растения, которое использует вирусный промотор для сверхэкспрессии гомолога томата (подробнее . ..)
..)
Второй ген, LEAFY, , который необходим для перехода от вегетативного к репродуктивному развитию, также, по-видимому, играет роль в развитии сложных листьев. Он был идентифицирован в Arabidopsis и львином зеве (в котором он называется FLORICAULA ), и имеет гомологи в других покрытосеменных растениях. Гомолог гороха ( UNIFOLIATA ) имеет мутантный фенотип, в котором сложные листья редуцированы до простых листьев (Hofer and Ellis 1998). Это открытие также свидетельствует о регуляторных отношениях между побегами и сложными листьями.
В некоторых сложных листьях в процессе развития также принимаются решения о формировании листа или усика. Мутации двух генов формы листа могут по отдельности и в совокупности резко изменить морфологию сложного листа гороха (10). Мутант акации ( tl ) превращает усики в листочки; afilia ( af ) превращает листовку в усики (Marx 1987). Двойной мутант af tl имеет сложную архитектуру и напоминает лист петрушки.
Рисунок 20.26
Мутанты морфологии листьев у гороха. (A) Горох дикого типа. (B) Мутант tl , у которого усики превращаются в листочки. (C) Мутант af , у которого листочки превращаются в усики. (D) Двойной мутант af tl , который приводит к «листу петрушки» (подробнее…)
На более микроскопическом уровне паттерн устьиц (отверстий для газообмена и водного обмена) и трихом (волосы) через лист также исследуется. У однодольных устьица формируются параллельными рядами, а у двудольных распределение кажется более случайным. В обоих случаях узоры обеспечивают максимальную равномерность распределения устьиц. Генетический анализ дает представление о механизмах, регулирующих это распространение. Общая группа генов, по-видимому, работает как в побегах, так и в корнях, влияя на характер распределения как трихом, так и корневых волосков (Benfey 19).99).
Сноски
- *
Сходство между клетками меристемы растений и стволовыми клетками животных может распространяться на молекулярный уровень, указывая на то, что стволовые клетки существовали до того, как растения и животные пошли разными филогенетическими путями.
Была обнаружена гомология между генами, необходимыми для сохранения меристем растений, и генами, экспрессируемыми в стволовых клетках зародышевой линии Drosophila (Cox et al. 1998).- †
Это явление, называемое фасциация, встречается у многих видов, включая горох и помидоры.
 Была обнаружена гомология между генами, необходимыми для сохранения меристем растений, и генами, экспрессируемыми в стволовых клетках зародышевой линии Drosophila (Cox et al. 1998).
Была обнаружена гомология между генами, необходимыми для сохранения меристем растений, и генами, экспрессируемыми в стволовых клетках зародышевой линии Drosophila (Cox et al. 1998).Что из следующего не является вегетативной частью растения?A. Карпель Б. Таламус С. ПрилистникиD. Calyx
Ответить
Проверено
216,3 тыс.+ просмотров
Подсказка: Вегетативные части не участвуют непосредственно в половом размножении. Они часто используются при бесполых формах размножения. Корни, стебель, побеги и листья являются вегетативными частями растения.
Полный ответ: Внешние структуры растений, например, листья, стебли, корни, плоды, семена известны как органы растений. Каждый орган представляет собой совокупность тканей, взаимодействующих для выполнения определенной функции. Эти структуры можно разделить на две группы: половое размножение и бесполое размножение (вегетативное размножение).
Эти структуры можно разделить на две группы: половое размножение и бесполое размножение (вегетативное размножение).
— Вегетативные части включают корни, стебли, побеги и листья, они не связаны с половым размножением. Вегетативные части часто используются в бесполых формах размножения, таких как черенки, окулировка или прививка.
— Плодолистики или гинецей — женская репродуктивная часть цветка. Плодолистик состоит из трех частей, которые представляют собой особое клеймо, столбик и завязь. В зависимости от количества плодолистиков гинецей может быть моноплодолистным, двуплодолистным или полиплодолистным.
— Таламус или вместилище представляет собой вегетативную часть, на которой находятся репродуктивные органы.
— Прилистник представляет собой небольшой отросток по обе стороны от основания черешка листа.
— Чашечка или чашелистики — самая внешняя мутовка, предназначенная для защиты других частей цветка. Чашелистики с оттенком, отличным от зеленого, называются лепестками.