
Содержание
Развитие эндосперма и зародыша у растений
Из оплодотворенной яйцеклетки, или зиготы, путем последовательных делений развивается зародыш, из оплодотворенной центральной клетки или клетки зачатка эндосперма развивается эндосперм.
После слияния спермия с яйцеклеткой, т. е. мужской и женской гаплоидных гамет, в оплодотворенной яйцеклетке восстанавливается диплоидное число хромосом. Первичное ядро эндосперма, являясь обычно продуктом слияния трех ядер — двух полярных и одного спермия — имеет триплоидное число хромосом. В редких случаях, когда эндосперм образуется без оплодотворения, его клетки имеют диплоидное число хромосом, а иногда и гаплоидное (представители семейства Onagraceae). Наконец, у тех растений, у которых при образовании вторичного ядра сливаются не два, а несколько ядер (3, 4, 7, 8 и даже 14), эндосперм может быть более или менее полиплоидным. После оплодотворения яйцеклетка и центральная клетка зародышевого мешка становятся способными к активному развитию. Двойное оплодотворение отсутствует у всех архегониальных растений и свойственно лишь покрытосеменным.
Двойное оплодотворение отсутствует у всех архегониальных растений и свойственно лишь покрытосеменным.
Во время развития зародыша и эндосперма зародышевый мешок нуждается в особенно обильном притоке питательных веществ, который осуществляется за счет извлечения их из окружающих материнских тканей, а также благодаря непосредственному потреблению зародышевым мешком тканей нуцеллуса и покровов семяпочки. Этому способствуют различного типа гаустории (присоски), развитые у многих растений. Под воздействием интенсивного обмена веществ размеры зародышевого мешка значительно увеличиваются. Наряду с этим стимулируется дальнейший рост и развитие семяпочки.
Период времени, протекающий от оплодотворения до образования зародыша и эндосперма, у разных растений неодинаков, колеблясь от 10—15 дней (у кок-сагыза, салата и др.) до нескольких недель (у злаков, хлопчатника и др.) и даже до нескольких месяцев (у черного саксаула и др.).
После двойного оплодотворения чаще всего первым делится первичное ядро эндосперма (продукт слияния вторичного — центрального ядра зародышевого мешка со спермием). Эндосперм у разных растений формируется неодинаково. Различают три основных типа развития эндосперма: ядерный, или нуклеарный, клеточный, или целлюлярный, и базальный, или тип Helobiae. В каждом из этих типов имеется ряд более или менее значительных модификаций.
Эндосперм у разных растений формируется неодинаково. Различают три основных типа развития эндосперма: ядерный, или нуклеарный, клеточный, или целлюлярный, и базальный, или тип Helobiae. В каждом из этих типов имеется ряд более или менее значительных модификаций.
Ядерный тип образования эндосперма характеризуется тем, что вслед за делением первичного ядра эндосперма не происходит немедленного образования клеточных перегородок, а возникает большее или меньшее число ядер, свободно лежащих в общей цитоплазме.
Формирование клеточных перегородок при ядерном типе развития эндосперма наступает позднее, начинаясь либо в микропилярной части зародышевого мешка, либо по всей периферии его. В редких случаях при ядерном типе эндосперма клеточные оболочки вообще не закладываются. Клеточный тип, напротив, характеризуется тем, что после первого и каждого из последующих делений ядер эндосперма немедленно появляются клеточные перегородки.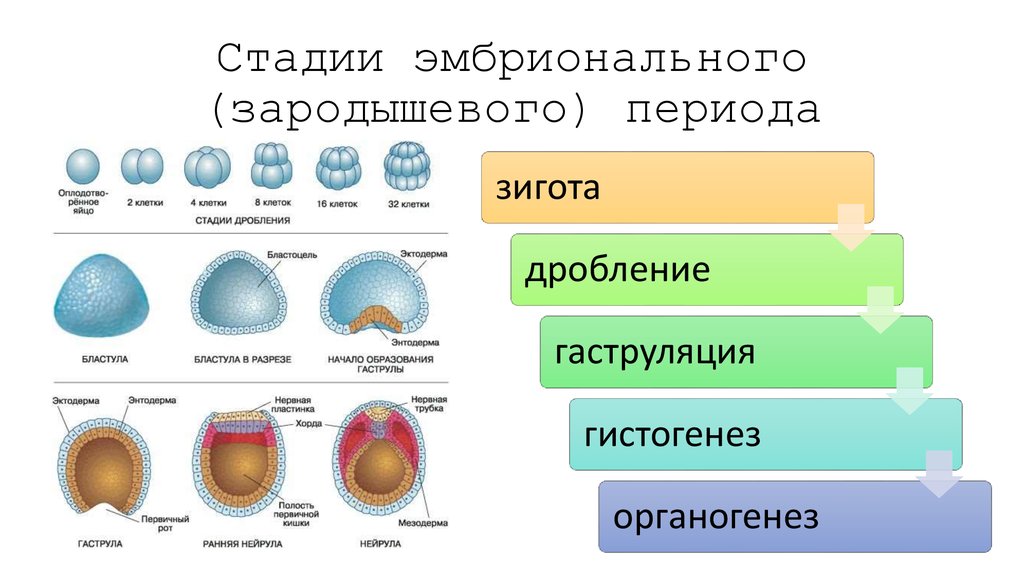
Базальный тип имеет промежуточный характер между ядерным и клеточным. Он отличается тем, что возникающая сразу после первого деления ядра эндосперма клеточная перегородка делит центральную клетку на две неравные части: большую верхнюю, или микропилярную, и маленькую нижнюю, или базальную. В микропилярной части эндосперма сначала образуется ряд свободных ядер и только позднее между ядрами появляются клеточные перегородки; в нижней же части либо образуются две клетки, либо небольшое число свободных ядер, либо делений вообще больше не происходит. Эта антиподальная часть эндосперма, или его базальный аппарат, функционирует часто как гаусторий.
Эндоспермальные гаустории формируются также при клеточном, а иногда и при ядерном типе развития эндосперма, возникая как в микропилярной, так и в халазальной и даже в латеральной части зародышевого мешка.
Гаусториями называются образования, состоящие из одно- или многоядерных клеток с густым содержимым и часто с гипертрофированными клеточными ядрами.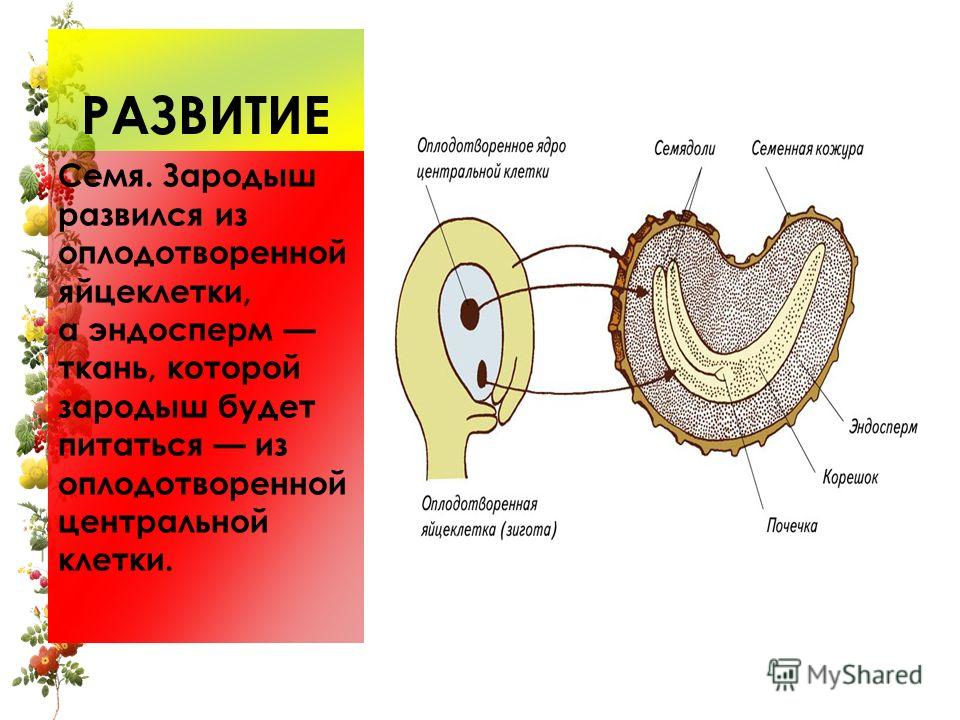 В одних случаях гаустории состоят из нескольких более или менее разросшихся и разветвленных клеток и даже целой ткани, в других — только из одной клетки, сильно разрастающейся и дающей боковые отростки.
В одних случаях гаустории состоят из нескольких более или менее разросшихся и разветвленных клеток и даже целой ткани, в других — только из одной клетки, сильно разрастающейся и дающей боковые отростки.
Наряду с эндоспермальными гаусториями в семяпочках покрытосеменных известны и другие типы гаусторий, образованных самим зародышевым мешком, антиподами, синергидами, подвеском зародыша. Гаустории весьма сильно варьируют по своему облику и строению, но роль их, по-видимому, одна и та же; они приспособлены к выполнению питательной и секреторной функции. Проникая своими отростками и боковыми ветвями глубоко в ткани семяпочки, плаценты и даже завязи, гаустории извлекают из материнского растения питательные вещества для роста и формирования зародыша. Эндоспермальные гаустории, как и гаустории любого другого типа, способствуют повышению интенсивности обмена веществ во время роста и развития зародыша, так как облегчают приток к нему питательных веществ из окружающих тканей материнского растения.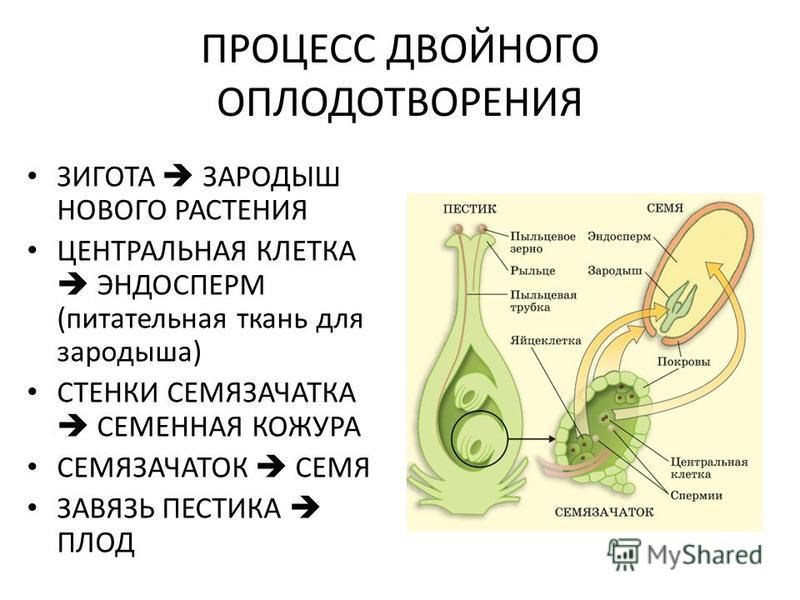
Примером гаусторий антиподального происхождения могут служить гаустории злаков. Комплекс антипод образует своеобразное подобие ткани, внедряющейся в нуцеллус. Клетки нуцеллуса, непосредственно примыкающие к клеткам антиподального комплекса, разрушаются.
При гибридизации и самоопылении перекрестников, а также под воздействием неблагоприятных внешних условий развитие эндосперма может преждевременно приостанавливаться или с самого начала происходит ненормально. Это, в свою очередь, сразу сказывается на формировании зародыша и приводит к образованию щуплых семян с недоразвитым зародышем.
В сформированном виде эндосперм зрелого семени (тех растений, у которых он сохраняется) представлен тканью, состоящей из более или менее крупных клеток, заполненных различными питательными веществами — крахмалом, маслом, сахарами, белком и т. д. Кроме того, в клетках эндосперма могут присутствовать дубильные вещества, алкалоиды, кристаллы оксалата кальция и т. д. Оболочки эндоспермальных клеток обычно тонкие, целлюлозные. У некоторых растений, у которых основное запасное вещество откладывается в оболочках, последние значительно утолщаются.
У некоторых растений, у которых основное запасное вещество откладывается в оболочках, последние значительно утолщаются.
По своей консистенции эндосперм у одних растений более или менее жидкий или студенистый, у других — в той или иной степени твердый или даже роговой. Эндосперм бывает либо бесцветным, либо различно пигментированным.
В дальнейшем эндосперм претерпевает различные превращения. У одних растений он полностью ассимилируется зародышем в процессе развития последнего и в зрелом семени или отсутствует полностью, или представлен одним-двумя слоями клеток. К таким растениям принадлежат бобовые, тыквенные, сложноцветные и другие. В этих случаях запасные питательные вещества, необходимые для прорастания зародыша, накапливаются в клетках семядолей самого зародыша, и последний заполняет собой почти весь объем семени.
У других растений эндосперм сохраняется до созревания семени и потребляется зародышем лишь при его прорастании (злаковые, ситниковые, магнолиевые и др.).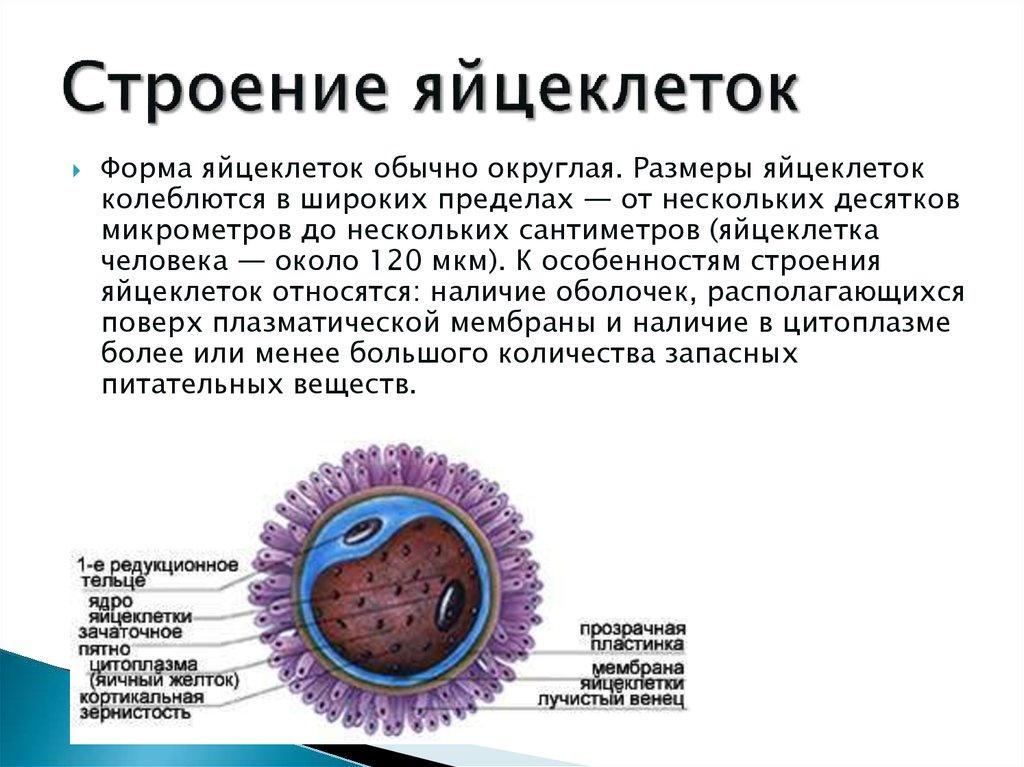 В этих случаях зародыши малы по объему, а основную часть семени занимает эндосперм. Между этими крайними примерами существует, конечно, много переходных форм, среди которых встречаются и такие, у которых объемы эндосперма и зародыша в зрелом семени одинаковые.
В этих случаях зародыши малы по объему, а основную часть семени занимает эндосперм. Между этими крайними примерами существует, конечно, много переходных форм, среди которых встречаются и такие, у которых объемы эндосперма и зародыша в зрелом семени одинаковые.
У некоторых растений (например, из семейства гвоздичных, перечноцветных) запасные вещества откладываются в нуцеллусе. Подобного рода запасная ткань называется периспермом.
Оплодотворенная яйцеклетка, или зигота, может делиться одновременно с делением первичного ядра эндосперма, но, как правило, зигота делится несколько позже (иногда значительно позже) — лишь после образования нескольких, а в некоторых случаях большого числа ядер или клеток эндосперма. Во время созревания зиготы она сама и ее ядро значительно увеличиваются в размерах, вакуоля постепенно исчезает, и вся клетка заполняется более или менее густым содержимым, концентрирующимся главным образом в апикальной ее части. По окончании периода созревания зигота начинает делиться, давая начало зародышу.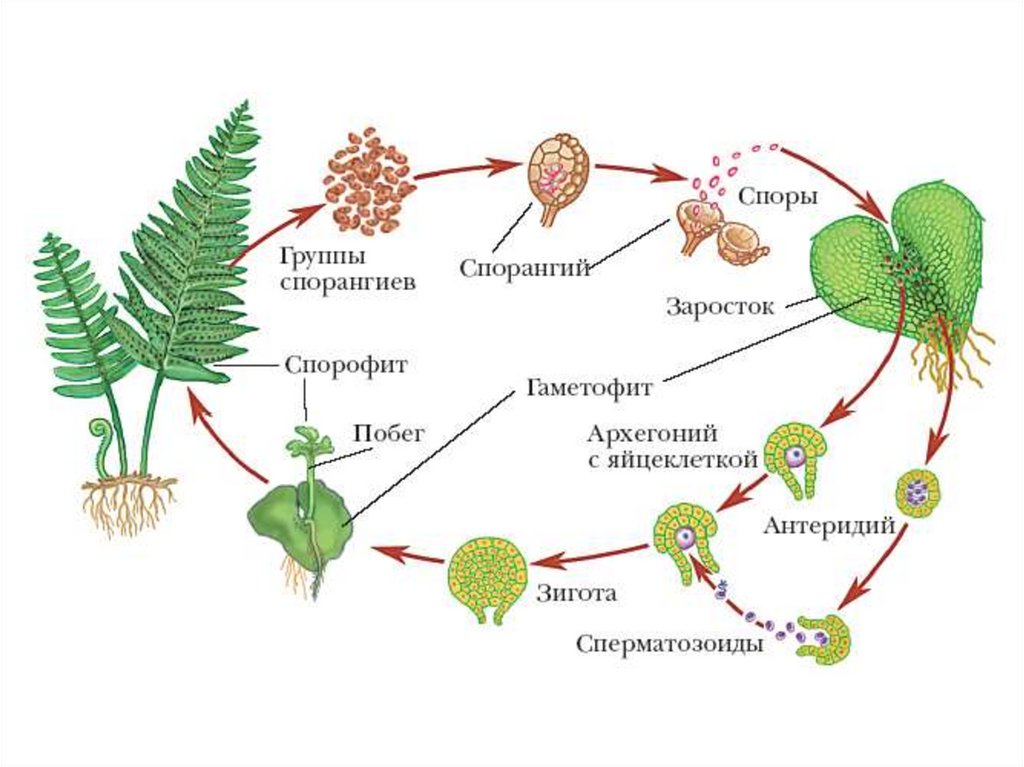
Имеется несколько классификаций типов развития зародыша, в основе которых лежат следующие признаки: способ заложения первой перегородки, делящей зиготу на две клетки, направления перегородок при последующих делениях, а также участие тех или иных клеток начинающего развиваться зародыша в формировании его органов.
При развитии зиготы покрытосеменного растения возникает клеточное тело, которое обычно уже на начальных фазах своего развития расчленено на базальную часть — подвесок, или суспензор, и терминальную — тело зародыша. Независимо от способа заложения клеточных перегородок зародыши покрытосеменных растений на самых ранних стадиях внешне довольно единообразны и являются радиально симметричными. В этот период нет различий даже между зародышами двудольных и однодольных растений, эти различия появляются позже. У двудольных растений тело зародыша постепенно принимает форму шара, который в дальнейшем несколько уплощается, после чего происходит дифференциация на семядоли и осевую часть. Через такой зародыш можно теперь провести только две плоскости симметрии.
Через такой зародыш можно теперь провести только две плоскости симметрии.
При развитии зародыша однодольных растений шарообразная стадия отсутствует, а зародыш из радиально симметричного становится билатерально симметричным, т. е. через него можно провести лишь одну плоскость симметрии. В некоторых случаях, например у злаков, это объясняется тем, что здесь очень рано нарушается равномерность ритма деления клеток зародыша— на вентральной его стороне деления задерживаются, в то время как на противоположной дорзальной стороне и в апикальной части зародыша они протекают интенсивно. В результате зародыш изгибается, а это в свою очередь обусловливает иной по сравнению с двудольными характер заложения органов. Единственная семядоля закладывается терминально, а точка роста занимает боковое положение. В других случаях не исключены и другие пути, приводящие к становлению своеобразного зародыша однодольных.
Несколько особняком в отношении формирования зародыша среди покрытосеменных растений стоит семейство Леоновых.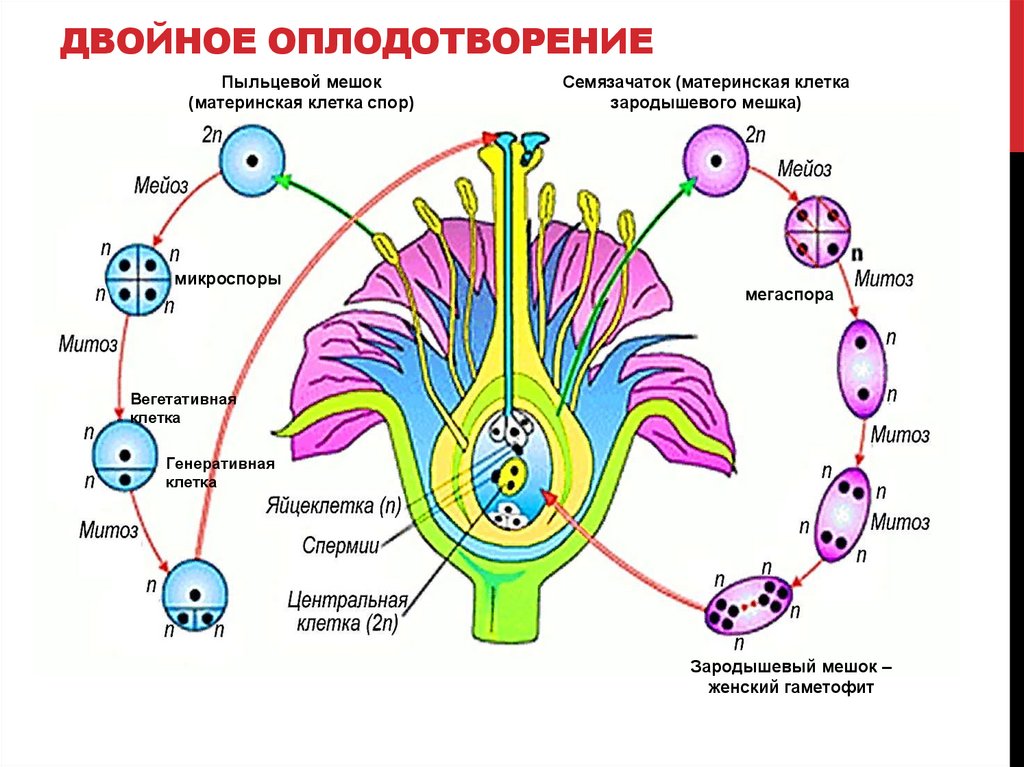 У его представителей деление зиготы и последующие деления производных ядер не сопровождаются заложением клеточных перегородок. В результате образуется предзародышевая ценоцитная структура, очень сходная с тем, что имеет место у голосеменных растений. Лишь позже между ядрами такого ценоцита закладываются перегородки и возникают меристематические очаги, дающие начало зародышам. Рост и дифференциация зародыша происходят в процессе активного обмена веществ, осуществляющегося между ним и окружающей его средой. Выше уже говорилось о том, что этому способствуют различного рода гаусториальные образования, формируемые различными структурами зародышевого мешка, в первую очередь эндоспермом. Такого рода гаусториальные функции свойственны также и самому зародышу, в частности его подвеску. При помощи подвеска зародыш продвигается в ткань эндосперма и получает наиболее благоприятные возможности для его использования; при помощи подвеска зародыш может получать питательные вещества и непосредственно из материнских тканей.
У его представителей деление зиготы и последующие деления производных ядер не сопровождаются заложением клеточных перегородок. В результате образуется предзародышевая ценоцитная структура, очень сходная с тем, что имеет место у голосеменных растений. Лишь позже между ядрами такого ценоцита закладываются перегородки и возникают меристематические очаги, дающие начало зародышам. Рост и дифференциация зародыша происходят в процессе активного обмена веществ, осуществляющегося между ним и окружающей его средой. Выше уже говорилось о том, что этому способствуют различного рода гаусториальные образования, формируемые различными структурами зародышевого мешка, в первую очередь эндоспермом. Такого рода гаусториальные функции свойственны также и самому зародышу, в частности его подвеску. При помощи подвеска зародыш продвигается в ткань эндосперма и получает наиболее благоприятные возможности для его использования; при помощи подвеска зародыш может получать питательные вещества и непосредственно из материнских тканей.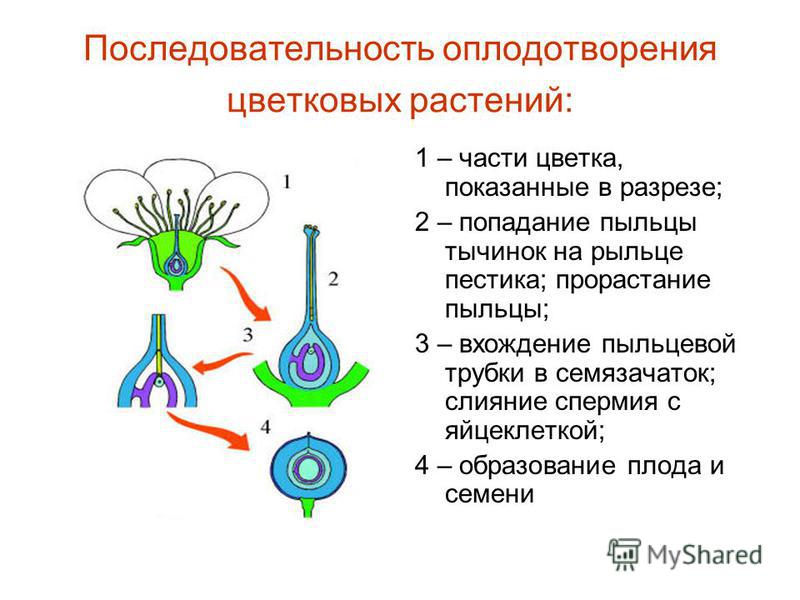 Облегчая поступление к зародышу питательных веществ из окружающих его тканей семяпочки, подвесок способствует повышению интенсивности обмена веществ на ранних стадиях роста и развития зародыша.
Облегчая поступление к зародышу питательных веществ из окружающих его тканей семяпочки, подвесок способствует повышению интенсивности обмена веществ на ранних стадиях роста и развития зародыша.
Подвески зародышей бывают одноклеточными и многоклеточными, клетки в них могут быть одноядерными или многоядерными. В некоторых случаях подвесок сильно разрастается и разветвляется, образуя подвесочный гаусторий. Иногда же подвесок совершенно не развивается или развивается крайне слабо.
Половые клетки человека, хромосомы, оплодотворение — статья МЦРМ
Половые клетки — гаметы (от греч. gametes — «супруг») можно обнаружить уже у двухнедельного эмбриона человека. Их называют первичными половыми клетками. В это время они совсем не похожи на сперматозоиды или яйцеклетки и выглядят абсолютно одинаковыми. Никаких различий, присущих зрелым гаметам, на этой стадии развития зародыша обнаружить у первичных половых клеток не удается. Это не единственная их особенность. Во-первых, первичные половые клетки появляются у зародыша гораздо раньше собственно половой железы (гонады), а во-вторых, они возникают на значительном удалении от того места, где эти железы сформируются позднее. В определенный момент происходит совершенно удивительный процесс — первичные половые клетки дружно устремляются к половой железе и заселяют, «колонизируют» ее.
Во-первых, первичные половые клетки появляются у зародыша гораздо раньше собственно половой железы (гонады), а во-вторых, они возникают на значительном удалении от того места, где эти железы сформируются позднее. В определенный момент происходит совершенно удивительный процесс — первичные половые клетки дружно устремляются к половой железе и заселяют, «колонизируют» ее.
После того, как будущие гаметы попали в половые железы, они начинают интенсивно делиться, и количество их увеличивается. На этом этапе половые клетки содержат пока то же количество хромосом, что и «телесные» (соматические) клетки — 46. Однако для успешного осуществления своей миссии половые клетки должны иметь в 2 раза меньше хромосом. В противном случае после оплодотворения, то есть слияния гамет, клетки зародыша будут содержать не 46, как установлено природой, а 92 хромосомы. Нетрудно догадаться, что в следующих поколениях их число прогрессивно бы увеличивалось. Чтобы избежать такой ситуации формирующиеся половые клетки проходят специальное деление, которое в эмбриологии называется мейоз (греч. meiosis — «уменьшение»). В результате этого удивительного процесса диплоидный (от греч. diploos — «двойной»), набор хромосом как бы «растаскивается» на составляющие его одинарные, гаплоидные наборы (от греч. haploos — одиночный). В результате из диплодной клетки с 46 хромосомами получаются 2 гаплоидные клетки с 23 хромосомами. Вслед за этим наступает завершающий этап формирования зрелых половых клеток. Теперь в гаплоидной клетке копируются имеющиеся 23 хромосомы и эти копии используются для образования новой клетки. Таким образом, в результате описанных двух делений из одной первичной половой клетки образуется 4 новых.
meiosis — «уменьшение»). В результате этого удивительного процесса диплоидный (от греч. diploos — «двойной»), набор хромосом как бы «растаскивается» на составляющие его одинарные, гаплоидные наборы (от греч. haploos — одиночный). В результате из диплодной клетки с 46 хромосомами получаются 2 гаплоидные клетки с 23 хромосомами. Вслед за этим наступает завершающий этап формирования зрелых половых клеток. Теперь в гаплоидной клетке копируются имеющиеся 23 хромосомы и эти копии используются для образования новой клетки. Таким образом, в результате описанных двух делений из одной первичной половой клетки образуется 4 новых.
Причем, в сперматогенезе (греч. genesis — зарождение, развитие) в результате мейоза появляется 4 зрелых сперматозоида с гаплоидным набором хромосом, а в процессе формирования яйцеклетки — в оогенезе (от греч. oon — «яйцо») только одна. Это происходит потому, что образовавшийся в результате мейоза второй гаплоидный набор хромосом яйцеклетка не использует для формирования новой зрелой половой клетки — ооцита, а «выбрасывает» их, как «лишние», наружу в своеобразном «мусорном контейнере», который называется полярным тельцем. Первое деление хромосомного набора завершается в оогенезе выделением первого полярного тельца непосредственно перед овуляцией. Второе репликационное деление происходит только после проникновения сперматозоида внутрь яйцеклетки и сопровождается выделением второго полярного тельца. Для эмбриологов полярные тельца — очень важные диагностические показатели. Есть первое полярное тельце, значит яйцеклетка зрелая, появилось второе полярное тельце — оплодотворение произошло.
Первое деление хромосомного набора завершается в оогенезе выделением первого полярного тельца непосредственно перед овуляцией. Второе репликационное деление происходит только после проникновения сперматозоида внутрь яйцеклетки и сопровождается выделением второго полярного тельца. Для эмбриологов полярные тельца — очень важные диагностические показатели. Есть первое полярное тельце, значит яйцеклетка зрелая, появилось второе полярное тельце — оплодотворение произошло.
Первичные половые клетки, оказавшиеся в мужской половой железе, до поры до времени не делятся. Их деление начинается только в период полового созревания и приводит к образованию когорты так называемых стволовых диплоидных клеток, из которых и формируются сперматозоиды. Запас стволовых клеток в яичках постоянно пополняется. Здесь уместно напомнить описанную выше особенность сперматогенеза — из одной клетки образуется 4 зрелых сперматозоида. Таким образом, после полового созревания у мужчины в течение всей жизни формируются сотни миллиардов новых сперматозоидов.
Формирование яйцеклеток протекает иначе. Едва заселив половую железу, первичные половые клетки начинают интенсивно делиться. К 5 месяцу внутриутробного развития их количество достигает 6-7 миллионов, но затем происходит массовая гибель этих клеток. В яичниках новорожденной девочки их остается не более 1-2 миллионов, к 7-летнему возрасту — всего лишь около 300 тысяч, а в период полового созревания 30 —50 тысяч. Общее же число яйцеклеток, которые достигнут зрелого состояния за период половой зрелости, будет еще меньше. Хорошо известно, что в течение одного менструального цикла в яичнике обычно созревает лишь один фолликул. Нетрудно подсчитать, что в течение репродуктивного периода, продолжающегося у женщин 30 — 35 лет, образуется около 400 зрелых яйцеклеток.
Если мейоз в сперматогенезе начинается в период полового созревания и повторяется миллиарды раз в течение жизни мужчины, в оогенезе формирующиеся женские гаметы вступают в мейоз еще в периоде внутриутробного развития. Причем начинается этот процесс почти одновременно у всех будущих яйцеклеток. Начинается, но не заканчивается! Будущие яйцеклетки доходят только до середины первой фазы мейоза, а дальше процесс деления блокируется на 12 — 50 лет! Лишь с приходом половой зрелости мейоз в оогенезе продолжится, причем не всех клеток сразу, а лишь для 1- 2 яйцеклеток ежемесячно. Полностью же процесс мейотического деления яйцеклетки завершится, как уже было сказано выше, только после ее оплодотворения! Таким образом, сперматозоид проникает в яйцеклетку, еще не завершившую деление, имеющую диплоидный набор хромосом!
Начинается, но не заканчивается! Будущие яйцеклетки доходят только до середины первой фазы мейоза, а дальше процесс деления блокируется на 12 — 50 лет! Лишь с приходом половой зрелости мейоз в оогенезе продолжится, причем не всех клеток сразу, а лишь для 1- 2 яйцеклеток ежемесячно. Полностью же процесс мейотического деления яйцеклетки завершится, как уже было сказано выше, только после ее оплодотворения! Таким образом, сперматозоид проникает в яйцеклетку, еще не завершившую деление, имеющую диплоидный набор хромосом!
Сперматогенез и оогенез — очень сложные и во многом загадочные процессы. Вместе с тем очевидна подчиненность их законам взаимосвязи и обусловленности природных явлений. Для оплодотворения одной яйцеклетки in vivo (лат. в живом организме) необходимы десятки миллионов сперматозоидов. Мужской организм вырабатывает их в гигантских количествах практически всю жизнь.
Вынашивание и рождение ребенка является чрезвычайно тяжелой нагрузкой на организм. Врачи говорят, что беременность — это проба на здоровье. Каким родится ребенок — напрямую зависит от состояния здоровья матери. Здоровье, как известно, не вечно. Старость и болезни, к сожалению, неотвратимы. Природа дает женщине строго ограниченное невосполнимое число половых клеток. Снижение способности к деторождению развивается медленно, но постепенно по наклонной. Наглядное доказательство того, что это действительно так, мы получаем, ежедневно оценивая результаты стимуляции яичников в программах ВРТ. Большая часть яйцеклеток обычно израсходована к 40 годам, а к 50 годам весь их запас полностью исчерпывается. Нередко так называемое истощение яичников наступает значительно раньше. Следует также сказать, что яйцеклетка подвержена «старению», с годами ее способность к оплодотворению снижается, процесс деления хромосом все чаще нарушается. Заниматься деторождением в позднем репродуктивном возрасте рискованно из-за возрастающей опасности рождения ребенка с хромосомной патологией.
Врачи говорят, что беременность — это проба на здоровье. Каким родится ребенок — напрямую зависит от состояния здоровья матери. Здоровье, как известно, не вечно. Старость и болезни, к сожалению, неотвратимы. Природа дает женщине строго ограниченное невосполнимое число половых клеток. Снижение способности к деторождению развивается медленно, но постепенно по наклонной. Наглядное доказательство того, что это действительно так, мы получаем, ежедневно оценивая результаты стимуляции яичников в программах ВРТ. Большая часть яйцеклеток обычно израсходована к 40 годам, а к 50 годам весь их запас полностью исчерпывается. Нередко так называемое истощение яичников наступает значительно раньше. Следует также сказать, что яйцеклетка подвержена «старению», с годами ее способность к оплодотворению снижается, процесс деления хромосом все чаще нарушается. Заниматься деторождением в позднем репродуктивном возрасте рискованно из-за возрастающей опасности рождения ребенка с хромосомной патологией. Типичным примером является синдром Дауна, который возникает из-за оставшейся при делении третьей лишней 21 хромосомы. Таким образом, ограничив репродуктивный период, природа охраняет женщину и заботится о здоровом потомстве.
Типичным примером является синдром Дауна, который возникает из-за оставшейся при делении третьей лишней 21 хромосомы. Таким образом, ограничив репродуктивный период, природа охраняет женщину и заботится о здоровом потомстве.
По каким законам происходит деление хромосом? Как передается наследственная информация? Для того чтобы разобраться с этим вопросом, можно привести простую аналогию с картами. Представим себе молодую супружескую пару. Назовем их условно — Он и Она. В каждой его соматической клетке находятся хромосомы черной масти — трефы и пики. Набор треф от шестерки до туза он получил от своей мамы. Набор пик — от своего папы. В каждой ее соматической клетке хромосомы красной масти — бубны и червы. Набор бубен от шестерки до туза она получила от своей мамы. Набор червей — от своего папы.
Для того чтобы получить из диплоидной соматической клетки половую клетку, число хромосом должно быть уменьшено вдвое. При этом половая клетка обязательно должна содержать полный одинарный (гаплоидный) набор хромосом. Ни одна не должна потеряться! В случае карт такой набор можно получить следующим образом. Взять наугад из каждой пары карт черной масти по одной и таким образом сформировать два одиночных набора. Каждый набор будет включать все карты черной масти от шестерки до туза, однако, какие именно это будут карты (трефы или пики) определил случай. Например, в одном таком наборе шестерка может быть пиковой, а в другом — трефовой. Нетрудно прикинуть, что в примере с картами при таком выборе одиночного набора из двойного мы можем получить 2 в девятой степени комбинаций — более 500 вариантов!
Ни одна не должна потеряться! В случае карт такой набор можно получить следующим образом. Взять наугад из каждой пары карт черной масти по одной и таким образом сформировать два одиночных набора. Каждый набор будет включать все карты черной масти от шестерки до туза, однако, какие именно это будут карты (трефы или пики) определил случай. Например, в одном таком наборе шестерка может быть пиковой, а в другом — трефовой. Нетрудно прикинуть, что в примере с картами при таком выборе одиночного набора из двойного мы можем получить 2 в девятой степени комбинаций — более 500 вариантов!
Точно также будем составлять одиночный набор из ее карт красной масти. Получим еще более 500 разных вариантов. Из его одиночного и ее одиночного набора карт составим двойной набор. Он получится мягко сказать «пестреньким»: в каждой паре карт одна будет красной масти, а другая — черной. Общее число таких возможных наборов 500×500, то есть 250 тысяч вариантов.
Примерно также, по закону случайной выборки, поступает и природа с хромосомами в процессе мейоза.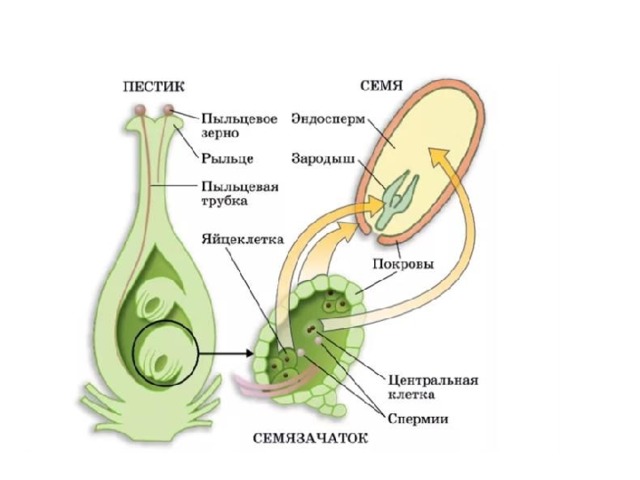 В результате из клеток с двойным, диплоидным набором хромосом получаются клетки, каждая из которых содержит одиночный, гаплоидный полный набор хромосом. Предположим, в результате мейоза в вашем теле образовалась половая клетка. Сперматозоид или яйцеклетка — в данном случае не важно. Она обязательно будет содержать гаплоидный набор хромосом — ровно 23 штуки. Что именно это за хромосомы? Рассмотрим для примера хромосому № 7. Это может быть хромосома, которую вы получили от отца. С равной вероятностью она может быть хромосомой, которую вы получили от матери. То же самое справедливо для хромосомы № 8, и для любой другой.
В результате из клеток с двойным, диплоидным набором хромосом получаются клетки, каждая из которых содержит одиночный, гаплоидный полный набор хромосом. Предположим, в результате мейоза в вашем теле образовалась половая клетка. Сперматозоид или яйцеклетка — в данном случае не важно. Она обязательно будет содержать гаплоидный набор хромосом — ровно 23 штуки. Что именно это за хромосомы? Рассмотрим для примера хромосому № 7. Это может быть хромосома, которую вы получили от отца. С равной вероятностью она может быть хромосомой, которую вы получили от матери. То же самое справедливо для хромосомы № 8, и для любой другой.
Поскольку у человека число хромосом гаплоидного набора равно 23, то число возможных вариантов половых гаплоидных клеток, образующихся из диплоидных соматических, равно 2 в степени 23. Получается более 8 миллионов вариантов! В процессе оплодотворения две половые клетки соединяются между собой. Следовательно, общее число таких комбинаций будет равно 8 млн. х 8 млн. = 64000 млрд. вариантов! На уровне пары гомологичных хромосом основа этого разнообразия выглядит так. Возьмем любую пару гомологичных хромосом вашего диплоидного набора. Одну из таких хромосом вы получили от матери, но это может быть хромосома либо вашей бабушки, либо вашего дедушки по материнской линии. Вторую гомологичную хромосому вы получили от отца. Однако она опять-таки может быть независимо от первой либо хромосомой вашей бабушки, либо вашего дедушки уже по отцовской линии. А таких гомологических хромосом у вас 23 пары! Получается невероятное число возможных комбинаций. Неудивительно, что при этом у одной пары родителей, рождаются дети, которые отличаются друг от друга и внешностью, и характером.
вариантов! На уровне пары гомологичных хромосом основа этого разнообразия выглядит так. Возьмем любую пару гомологичных хромосом вашего диплоидного набора. Одну из таких хромосом вы получили от матери, но это может быть хромосома либо вашей бабушки, либо вашего дедушки по материнской линии. Вторую гомологичную хромосому вы получили от отца. Однако она опять-таки может быть независимо от первой либо хромосомой вашей бабушки, либо вашего дедушки уже по отцовской линии. А таких гомологических хромосом у вас 23 пары! Получается невероятное число возможных комбинаций. Неудивительно, что при этом у одной пары родителей, рождаются дети, которые отличаются друг от друга и внешностью, и характером.
Кстати, из приведенных выше расчетов следует простой, но важный вывод. Каждый человек, ныне здравствующий, или когда-либо живший в прошлом на Земле, абсолютно уникален. Шансы появления второго такого же практически равны нулю. Поэтому не надо себя ни с кем сравнивать. Каждый из вас неповторим, и тем уже интересен!
Однако вернемся к нашим половым клеткам. Каждая диплоидная клетка человека содержит 23 пары хромосом. Хромосомы с 1 по 22 пару называются соматическим и по форме они одинаковы. Хромосомы же 23-й пары (половые хромосомы) одинаковы только у женщин. Они и обозначаются латинскими буквами ХХ. У мужчин хромосомы этой пары различны и обозначаются ХY. В гаплоидном наборе яйцеклетки половая хромосома всегда только Х, сперматозоид же может нести или Х или Y хромосому. Если яйцеклетку оплодотворит Х сперматозоид, родится девочка, если Y сперматозоид — мальчик. Все просто!
Каждая диплоидная клетка человека содержит 23 пары хромосом. Хромосомы с 1 по 22 пару называются соматическим и по форме они одинаковы. Хромосомы же 23-й пары (половые хромосомы) одинаковы только у женщин. Они и обозначаются латинскими буквами ХХ. У мужчин хромосомы этой пары различны и обозначаются ХY. В гаплоидном наборе яйцеклетки половая хромосома всегда только Х, сперматозоид же может нести или Х или Y хромосому. Если яйцеклетку оплодотворит Х сперматозоид, родится девочка, если Y сперматозоид — мальчик. Все просто!
Почему мейоз у яйцеклетки так долго растянут во времени? Каким образом ежемесячно происходит выбор когорты фолликулов, которые начинают свое развитие и как из них выделяется лидирующий, доминантный, овуляторный фолликул, в котором созреет яйцеклетка? На все эти непростые вопросы у биологов нет пока однозначных ответов. Процесс формирования зрелых яйцеклеток у человека ждет новых исследователей!
Образование и созревание сперматозоидов, как уже было сказано, происходит в семенных канальцах мужской половой железы — яичках.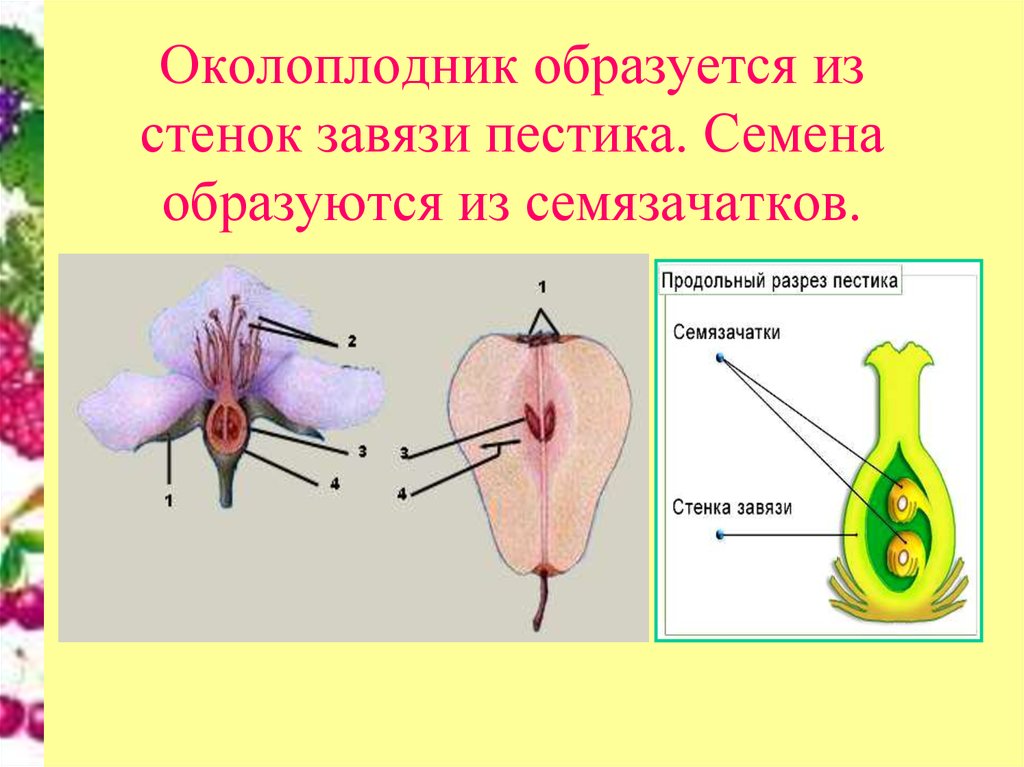 Сформированный сперматозоид имеет длину около 50-60 микрон. Ядро сперматозоида находится в его головке. Оно содержит отцовский наследственный материал. За головкой располагается шейка, в которой имеется крупная извитая митохондрия — органоид, обеспечивающий движения хвоста. Иначе говоря, это своеобразная «энергетическая станция». На головке сперматозоида есть «шапочка». Благодаря ей форма головки — овальная. Но, дело не в форме, а в том, что содержится под «шапочкой». «Шапочка» эта на самом деле является контейнером и называется акросомой, а содержатся в ней ферменты, которые способны растворять оболочку яйцеклетки, что необходимо для проникновения сперматозоида внутрь — в цитоплазму яйцеклетки. Если у сперматозоида нет акросомы, головка у него не овальная, а круглая. Эта патология сперматозоидов называется глобулоспермия (круглоголовые сперматозоиды). Но, беда опять не в форме, а в том, что такой сперматозоид не может оплодотворить яйцеклетку, и мужчина с таким нарушением сперматогенеза до начала 90-х прошлого столетия был обречен на бездетность.
Сформированный сперматозоид имеет длину около 50-60 микрон. Ядро сперматозоида находится в его головке. Оно содержит отцовский наследственный материал. За головкой располагается шейка, в которой имеется крупная извитая митохондрия — органоид, обеспечивающий движения хвоста. Иначе говоря, это своеобразная «энергетическая станция». На головке сперматозоида есть «шапочка». Благодаря ей форма головки — овальная. Но, дело не в форме, а в том, что содержится под «шапочкой». «Шапочка» эта на самом деле является контейнером и называется акросомой, а содержатся в ней ферменты, которые способны растворять оболочку яйцеклетки, что необходимо для проникновения сперматозоида внутрь — в цитоплазму яйцеклетки. Если у сперматозоида нет акросомы, головка у него не овальная, а круглая. Эта патология сперматозоидов называется глобулоспермия (круглоголовые сперматозоиды). Но, беда опять не в форме, а в том, что такой сперматозоид не может оплодотворить яйцеклетку, и мужчина с таким нарушением сперматогенеза до начала 90-х прошлого столетия был обречен на бездетность. Сегодня благодаря ВРТбесплодие у этих мужчин может быть преодолено, но об этом мы расскажем позднее в главе, посвященной микроманипуляциям, в частности, ИКСИ.
Сегодня благодаря ВРТбесплодие у этих мужчин может быть преодолено, но об этом мы расскажем позднее в главе, посвященной микроманипуляциям, в частности, ИКСИ.
Перемещение сперматозоида осуществляется за счет движения его хвостика. Скорость движения сперматозоида не превышает 2-3 мм в минуту. Казалось бы, немного, однако, за 2-3 часа в женском половом тракте сперматозоиды проходят путь, в 80000 раз превышающий их собственные размеры! Будь на месте сперматозоида в этой ситуации человек, ему пришлось бы двигаться вперед со скоростью 60-70 км/час — то есть со скоростью автомобиля!
Сперматозоиды, находящиеся в яичке, неподвижны. Способность к движению они приобретают лишь, проходя по семявыводящим путям под воздействием жидкостей семявыводящих протоков и семенных пузырьков, секрета предстательной железы. В половых путях женщины сперматозоиды сохраняют подвижность в течение 3 — 4 суток, но оплодотворить яйцеклетку они должны в течение 24 часов. Весь процесс развития от стволовой клетки до зрелого сперматозоида длится примерно 72 дня. Однако, поскольку сперматогенез происходит непрерывно и в него одномоментно вступает громадное число клеток, то в яичках всегда есть большое количество спермиев, находящихся на разных этапах сперматогенеза, а запас зрелых сперматозоидов постоянно пополняется. Активность сперматогенеза индивидуальна, но с возрастом снижается.
Однако, поскольку сперматогенез происходит непрерывно и в него одномоментно вступает громадное число клеток, то в яичках всегда есть большое количество спермиев, находящихся на разных этапах сперматогенеза, а запас зрелых сперматозоидов постоянно пополняется. Активность сперматогенеза индивидуальна, но с возрастом снижается.
Как мы уже говорили, яйцеклетки находятся в фолликулах яичника. В результате овуляции яйцеклетка попадает в брюшную полость, откуда она «вылавливается» фимбриями маточной трубы и переносится в просвет ее ампулярного отдела. Именно здесь происходит встреча яйцеклетки со сперматозоидами.
Какое же строение имеет зрелая яйцеклетка? Она довольно крупная и достигает 0,11-0,14 мм в диаметре. Сразу после овуляции яйцеклетка окружена скоплением мелких клеток и желатинообразной массой (так называемым лучистым венцом). Видимо, в таком виде фимбриям маточной трубы удобнее захватывать яйцеклетку. В просвете маточной трубы с помощью ферментов и механического воздействия (биения ресничек эпителия), происходит «очистка» яйцеклетки от лучистого венца. Окончательно освобождение яйцеклетки от лучистого венца происходит после встречи ее со сперматозоидами, которые буквально облепляют яйцеклетку. Каждый сперматозоид выделяет из акросомы фермент, растворяющий не только лучистый венец, но и действующий на оболочку самой яйцеклетки. Эта оболочка называется блестящей, так она выглядит под микроскопом. Выделяя фермент, все сперматозоиды стремятся оплодотворить яйцеклетку, но блестящая оболочка пропустит лишь один из них. Получается, что устремляясь к яйцеклетки, воздействуя на нее коллективно, сперматозоиды «расчищают дорогу» только для одного счастливчика. Отбором сперматозоида роль блестящей оболочки не ограничивается, на ранних стадиях развития эмбриона она поддерживает упорядоченное расположение его клеток (бластомеров). В какой-то момент блестящая оболочка становится тесной, она разрывается и происходит хетчинг (от анг.
В просвете маточной трубы с помощью ферментов и механического воздействия (биения ресничек эпителия), происходит «очистка» яйцеклетки от лучистого венца. Окончательно освобождение яйцеклетки от лучистого венца происходит после встречи ее со сперматозоидами, которые буквально облепляют яйцеклетку. Каждый сперматозоид выделяет из акросомы фермент, растворяющий не только лучистый венец, но и действующий на оболочку самой яйцеклетки. Эта оболочка называется блестящей, так она выглядит под микроскопом. Выделяя фермент, все сперматозоиды стремятся оплодотворить яйцеклетку, но блестящая оболочка пропустит лишь один из них. Получается, что устремляясь к яйцеклетки, воздействуя на нее коллективно, сперматозоиды «расчищают дорогу» только для одного счастливчика. Отбором сперматозоида роль блестящей оболочки не ограничивается, на ранних стадиях развития эмбриона она поддерживает упорядоченное расположение его клеток (бластомеров). В какой-то момент блестящая оболочка становится тесной, она разрывается и происходит хетчинг (от анг.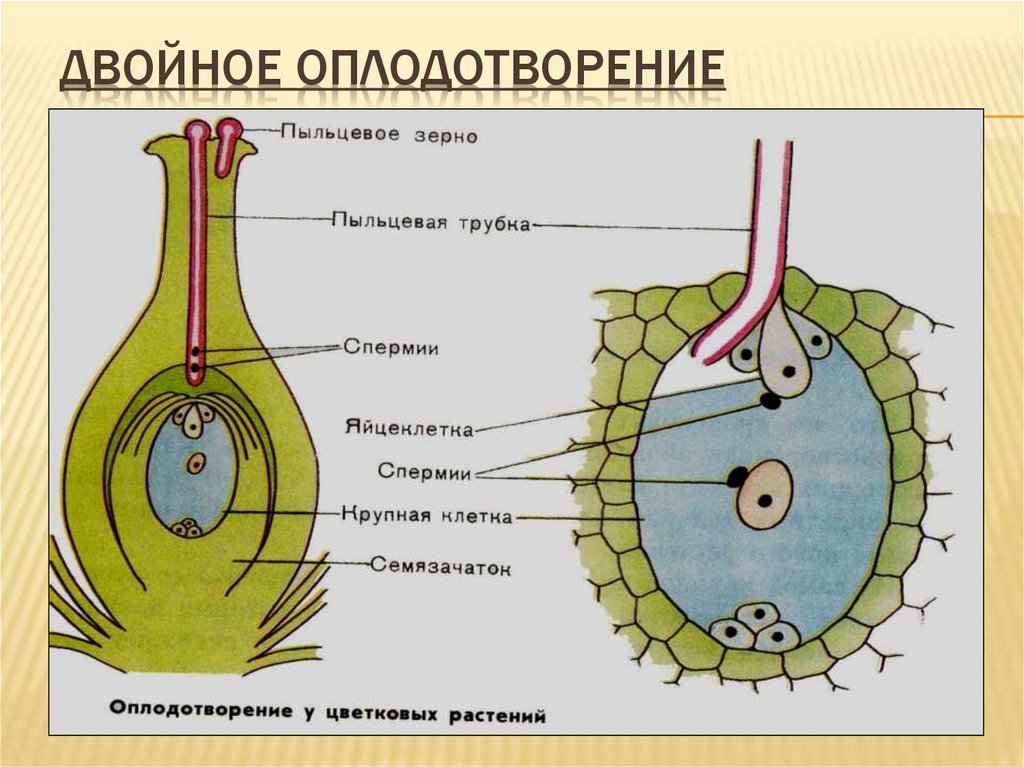 hatching — «вылупление») — вылупление эмбриона. Эмбрион готов к имплантации в эндометрий.
hatching — «вылупление») — вылупление эмбриона. Эмбрион готов к имплантации в эндометрий.
Оплодотворение | Шаги, процесс и факты
сперма
Посмотреть все СМИ
- Ключевые люди:
- Чарльз Дарвин
Кристиан Конрад Шпренгель
Эдуард ван Бенеден
Дама Энн Макларен
Фрэнк Рэттрей Лилли
- Похожие темы:
- двойное оплодотворение
перекрестное опыление
самооплодотворение
полиспермия
цитогамия
Просмотреть весь соответствующий контент →
Резюме
Прочтите краткий обзор этой темы
оплодотворение , слияние ядра сперматозоида отцовского происхождения с ядром яйцеклетки материнского происхождения с образованием первичного ядра эмбриона. У всех организмов суть оплодотворения состоит в слиянии наследственного материала двух различных половых клеток, или гамет, каждая из которых несет половину числа хромосом, типичного для данного вида. Самая примитивная форма оплодотворения, встречающаяся у микроорганизмов и простейших, состоит в обмене генетическим материалом между двумя клетками.
Самая примитивная форма оплодотворения, встречающаяся у микроорганизмов и простейших, состоит в обмене генетическим материалом между двумя клетками.
Первым важным событием при оплодотворении является слияние мембран двух гамет, в результате чего образуется канал, позволяющий проходить материалу из одной клетки в другую. Оплодотворению у развитых растений предшествует опыление, во время которого пыльца переносится на женскую гамету или макроспору и устанавливает с ней контакт. Слияние у продвинутых животных обычно сопровождается проникновением в яйцеклетку одного сперматозоида. Результатом оплодотворения является клетка (зигота), способная к клеточному делению с образованием новой особи.
Слияние двух гамет инициирует несколько реакций в яйцеклетке. Один из них вызывает изменение оболочки (оболочек) яйцеклетки, так что прикрепление и проникновение более чем одного сперматозоида не может произойти. У видов, у которых в норме в яйцеклетку входит более одного сперматозоида (полиспермия), только одно ядро сперматозоида фактически сливается с ядром яйцеклетки. Важнейшим результатом оплодотворения является активация яйцеклетки, которая позволяет яйцеклетке подвергнуться клеточному делению. Однако активация не обязательно требует вмешательства сперматозоида; во время партеногенеза, при котором оплодотворения не происходит, активация яйцеклетки может быть достигнута за счет вмешательства физических и химических агентов. Беспозвоночные, такие как тли, пчелы и коловратки, обычно размножаются партеногенезом.
Важнейшим результатом оплодотворения является активация яйцеклетки, которая позволяет яйцеклетке подвергнуться клеточному делению. Однако активация не обязательно требует вмешательства сперматозоида; во время партеногенеза, при котором оплодотворения не происходит, активация яйцеклетки может быть достигнута за счет вмешательства физических и химических агентов. Беспозвоночные, такие как тли, пчелы и коловратки, обычно размножаются партеногенезом.
В растениях некоторые химические вещества, вырабатываемые яйцеклеткой, могут привлекать сперматозоиды. У животных, за возможным исключением некоторых кишечнополостных (кишечнополостных), кажется вероятным, что контакт между яйцеклетками и сперматозоидами зависит от случайных столкновений. С другой стороны, желатиновые оболочки, окружающие яйцеклетку многих животных, улавливают сперматозоиды, увеличивая таким образом шансы на успешное взаимодействие сперматозоида с яйцеклеткой.
Яйца морских беспозвоночных, особенно иглокожих, являются классическими объектами для изучения оплодотворения. Эти прозрачные яйца ценны для изучения живых клеток, а также для биохимических и молекулярных исследований, поскольку время оплодотворения можно точно установить, развитие многих яиц происходит примерно с одинаковой скоростью в подходящих условиях, и можно получить большое количество яиц. Яйца некоторых костистых и амфибий также использовались с благоприятными результатами.
Эти прозрачные яйца ценны для изучения живых клеток, а также для биохимических и молекулярных исследований, поскольку время оплодотворения можно точно установить, развитие многих яиц происходит примерно с одинаковой скоростью в подходящих условиях, и можно получить большое количество яиц. Яйца некоторых костистых и амфибий также использовались с благоприятными результатами.
Созревание яйцеклетки
Созревание является заключительным этапом производства функциональных яйцеклеток (оогенеза), которые могут связываться со сперматозоидом и вызывать реакцию, препятствующую проникновению более чем одного сперматозоида. Кроме того, цитоплазма зрелой яйцеклетки может поддерживать изменения, приводящие к слиянию ядер сперматозоида и яйцеклетки и инициирующие эмбриональное развитие.
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Подпишитесь сейчас
Поверхность яйца
Некоторые компоненты поверхности яйца, особенно гранулы коры, связаны со зрелым состоянием. Кортикальные гранулы яиц морского ежа, расположенные под плазматической мембраной (тонкий, мягкий, податливый слой) зрелых яиц, имеют диаметр 0,8–1,0 мкм (0,0008–0,001 мм) и окружены мембраной, сходной по структуре с плазматической. мембрана, окружающая яйцо. Кортикальные гранулы образуются в клеточном компоненте, известном как комплекс Гольджи, из которого они мигрируют на поверхность созревающей яйцеклетки.
Кортикальные гранулы яиц морского ежа, расположенные под плазматической мембраной (тонкий, мягкий, податливый слой) зрелых яиц, имеют диаметр 0,8–1,0 мкм (0,0008–0,001 мм) и окружены мембраной, сходной по структуре с плазматической. мембрана, окружающая яйцо. Кортикальные гранулы образуются в клеточном компоненте, известном как комплекс Гольджи, из которого они мигрируют на поверхность созревающей яйцеклетки.
Поверхность яйца морского ежа обладает способностью неодинаково влиять на прохождение света в разных направлениях; это свойство, называемое двойным лучепреломлением, свидетельствует о том, что молекулы, составляющие поверхностные слои, расположены определенным образом. Поскольку двойное лучепреломление появляется по мере созревания яйцеклетки, вполне вероятно, что свойства мембраны зрелой яйцеклетки связаны со специфическим молекулярным расположением. Зрелая яйцеклетка способна поддерживать формирование ядра зиготы; т. е. результат слияния ядер сперматозоида и яйцеклетки. В большинстве яйцеклеток процесс редукции числа хромосом (мейоз) не завершается до оплодотворения. В таких случаях оплодотворяющий сперматозоид остается под поверхностью яйцеклетки до завершения мейоза в яйцеклетке, после чего происходят изменения и движения, ведущие к слиянию и образованию зиготы.
В большинстве яйцеклеток процесс редукции числа хромосом (мейоз) не завершается до оплодотворения. В таких случаях оплодотворяющий сперматозоид остается под поверхностью яйцеклетки до завершения мейоза в яйцеклетке, после чего происходят изменения и движения, ведущие к слиянию и образованию зиготы.
Оболочки яиц
Поверхность яиц большинства животных окружена оболочками, которые могут быть мягкими студенистыми оболочками (как у иглокожих и некоторых земноводных) или толстыми оболочками (как у рыб, насекомых и млекопитающих). Следовательно, чтобы достичь поверхности яйцеклетки, сперматозоиды должны проникнуть через эти оболочки; действительно, сперматозоиды содержат ферменты (органические катализаторы), которые их расщепляют. В некоторых случаях (например, у рыб и насекомых) в оболочке есть канал, или микропиле, по которому сперматозоид может попасть в яйцеклетку.
Желейная оболочка яиц иглокожих и амфибий состоит из сложных углеводов, называемых сульфатированными мукополисахаридами. Оболочка яйца млекопитающего более сложная. Яйцо окружено толстой оболочкой, состоящей из углеводно-белкового комплекса, называемого блестящей оболочкой. Зона окружена внешней оболочкой, corona radiata, которая состоит из многих слоев клеток и образована клетками фолликула, прилипающими к ооциту до того, как он покинет фолликул яичника.
Оболочка яйца млекопитающего более сложная. Яйцо окружено толстой оболочкой, состоящей из углеводно-белкового комплекса, называемого блестящей оболочкой. Зона окружена внешней оболочкой, corona radiata, которая состоит из многих слоев клеток и образована клетками фолликула, прилипающими к ооциту до того, как он покинет фолликул яичника.
Хотя когда-то постулировалось, что желеобразная оболочка яйца иглокожих содержит вещество (фертилизин), которое, как считается, играет важную роль не только в установлении взаимодействия сперматозоида с яйцеклеткой, но и в активации яйцеклетки, в настоящее время показано, что фертилизин идентичен гелеобразный материал, а не постоянно выделяемое из него вещество. Тем не менее есть свидетельства того, что оболочки яиц действительно играют роль в оплодотворении; т. е. контакт с оболочкой яйца вызывает акросомную реакцию (описанную ниже) у сперматозоидов.
Наконец-то, как растения откладывают яйца
Давняя загадка, связанная с фундаментальным процессом в биологии растений, раскрыта группой ученых из Калифорнийского университета в Дэвисе.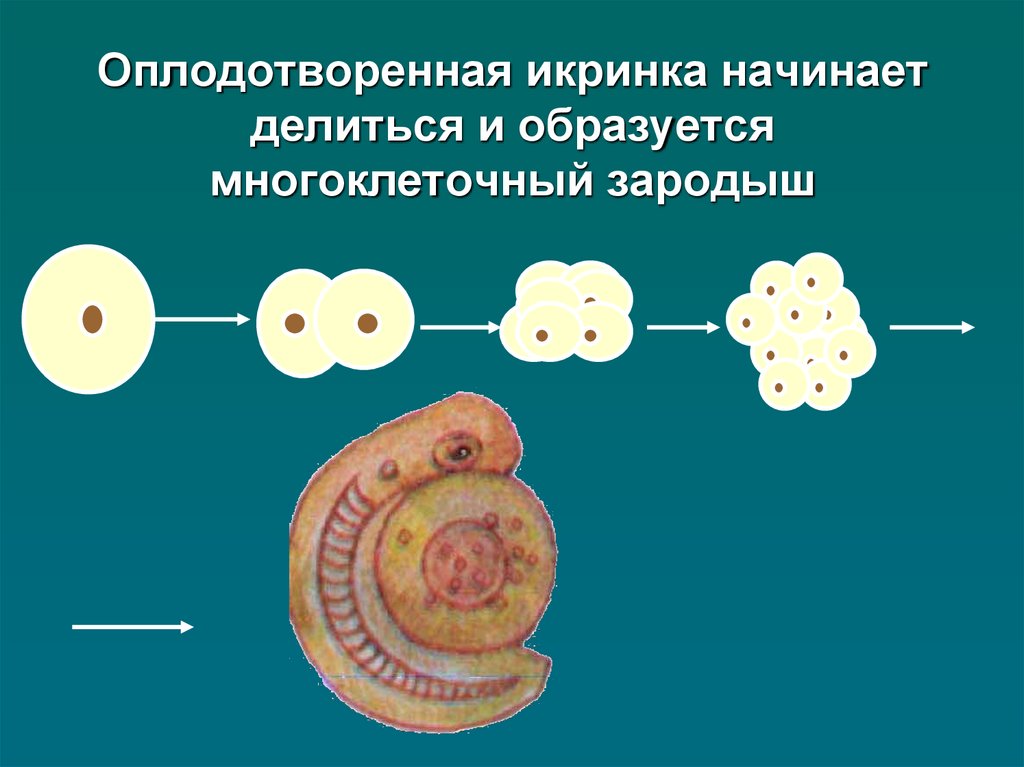
Новаторское открытие группы, что растительный гормон, называемый ауксином, отвечает за производство яиц, имеет несколько важных последствий.
Во-первых, это первый окончательный отчет о растительном гормоне, действующем как морфоген, то есть вещество, которое на основе его концентрации определяет модель развития клеток.
Кроме того, результаты исследования дают дразнящее новое понимание пути эволюции, который прошли цветковые растения 135 миллионов лет назад, когда они отделились от голосеменных, группы растений с «голыми семенами», в которую входят хвойные деревья, саговники и деревья гинкго.
Наконец, группа использовала свое открытие для создания дополнительных яйцеклеток в репродуктивных структурах растений, что повысило вероятность того, что эти методы когда-нибудь могут быть использованы для улучшения воспроизводства и плодородия сельскохозяйственных культур.
«Теперь последовательность становится ясной», — сказал Венкатесан Сундаресан, профессор биологии растений и наук о растениях Калифорнийского университета в Дэвисе, который руководил исследованием. «Растение запускает синтез ауксина на одном конце женской репродуктивной единицы, называемой зародышевым мешком, создавая градиент ауксина. Затем восемь ядер в мешочке подвергаются воздействию различных уровней ауксина, но только ядро в правильном положении в градиенте становится яйцеклеткой. И эта клетка впоследствии оплодотворяется, чтобы создать следующее поколение».
«Растение запускает синтез ауксина на одном конце женской репродуктивной единицы, называемой зародышевым мешком, создавая градиент ауксина. Затем восемь ядер в мешочке подвергаются воздействию различных уровней ауксина, но только ядро в правильном положении в градиенте становится яйцеклеткой. И эта клетка впоследствии оплодотворяется, чтобы создать следующее поколение».
Статья с описанием исследования была опубликована 4 июня на онлайн-сайте журнала Science, Science Express, до ее публикации в журнале в конце этого месяца.
Развитие сперматозоидов и яйцеклеток у растений
У людей и других животных зародышевые клетки для производства яйцеклеток и сперматозоидов формируются при рождении. Но клетки цветковых растений назначаются более или менее случайным образом, чтобы стать репродуктивными единицами, когда растение достигает половой зрелости. Внутри цветка сперматозоиды производятся пыльцой на кончиках тычинок, а яйцеклетки развиваются в семяпочках, крошечных структурах, встроенных в завязь у основания пестика.
В начале процесса развития яйцеклетки «материнская клетка» в семязачатке делится несколько раз в последовательности, включающей как мейозные, так и митотические деления. Эти деления приводят к созданию продолговатой клеточной структуры, называемой зародышевым мешком, который содержит восемь ядер, три из которых сгруппированы возле открытого конца семязачатка.
В течение нескольких часов начинают формироваться клеточные мембраны, в результате чего образуются семь клеток: самая важная яйцеклетка рядом с отверстием семязачатка, куда попадает пыльца, и шесть других поддерживающих клеток, выполняющих важные функции для образования семян.
«Большой вопрос в нашей области в течение последних 50 или более лет заключался в следующем: как этот процесс происходит в такой прекрасно организованной схеме?» — сказал Сундаресан. «Было ясно, что здесь есть программа, которая точно говорит растениям, что делать, и что она работает не с клетками, а с ядрами».
Концентрация ауксина определяет судьбу ядер
Два года назад Сундаресан и научный сотрудник его лаборатории Габриэла Пагнуссат использовали генетические инструменты для смещения положения одного ядра на одном конце зародышевого мешка у растения Arabidopsis. Когда они исследовали зрелый мешок, они обнаружили, что он произвел две яйцеклетки вместо одной.
Когда они исследовали зрелый мешок, они обнаружили, что он произвел две яйцеклетки вместо одной.
Сундаресан обнаружил, что подобное изменение паттерна было похоже на реакцию, о которой сообщалось двумя десятилетиями ранее у плодовых мушек Drosophila в экспериментах, которые предоставили первые прямые доказательства существования морфогенов.
Это побудило его начать поиск вещества в арабадопсисе, которое могло действовать как морфоген. Когда группа обнаружила, что ауксин накапливается на открытом конце семязачатка, они обратили свое внимание на этот вездесущий гормон, который, как известно, играет множество сигнальных ролей в процессах роста и поведения растений. (О существовании этого гормона впервые догадался Чарльз Дарвин, когда изучал, как растения растут к свету.)
После многих тестов Сундаресан и его группа обнаружили, что во время формирования зародышевого мешка концентрации ауксина действительно следуют градиенту, при этом самые высокие уровни наблюдаются в семязачатке в конце зародышевого мешка, куда входит пыльца, а самые низкие уровни наблюдаются в конце зародышевого мешка.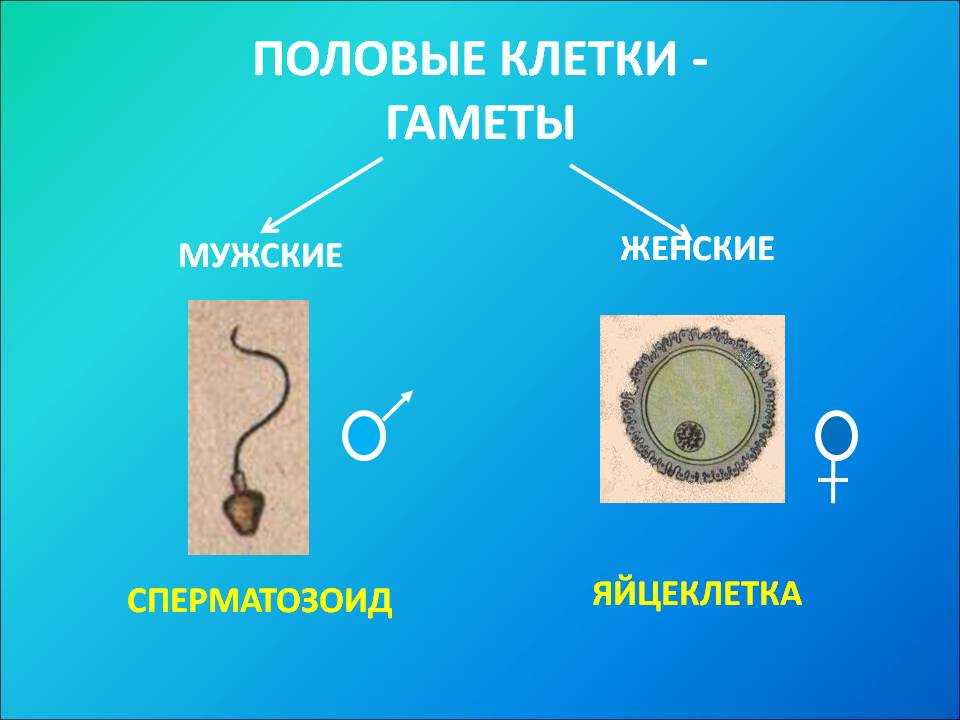 противоположный конец мешка.
противоположный конец мешка.
Чтобы проверить теорию о том, что этот градиент определяет судьбу ядер в мешочке, Сундаресан и его группа создали серию генетически модифицированных растений арабадопсиса. У некоторых растений они усиливали выработку ауксина в зародышевом мешке, а у других снижали чувствительность мешка к ауксину, вызывая тот же эффект, что и уменьшение ауксина.
Когда они исследовали эти экспериментальные растения, их гипотеза подтвердилась: концентрация ауксина определяет судьбу ядер. Зная, были ли уровни ауксина высокими или низкими, стало возможным предсказать появление или исчезновение яйцеклеток в разных местах внутри зародышевого мешка.
Наконец, группа использовала длинную серию биоманипулятивных методов, чтобы определить, что градиент ауксина, который они обнаружили в зародышевом мешке, был обусловлен синтезом на месте, а не транспортом из источника вне мешка.
«То, что мы узнали о том, как работает здесь ауксин, просто поразительно, — сказал Сундаресан.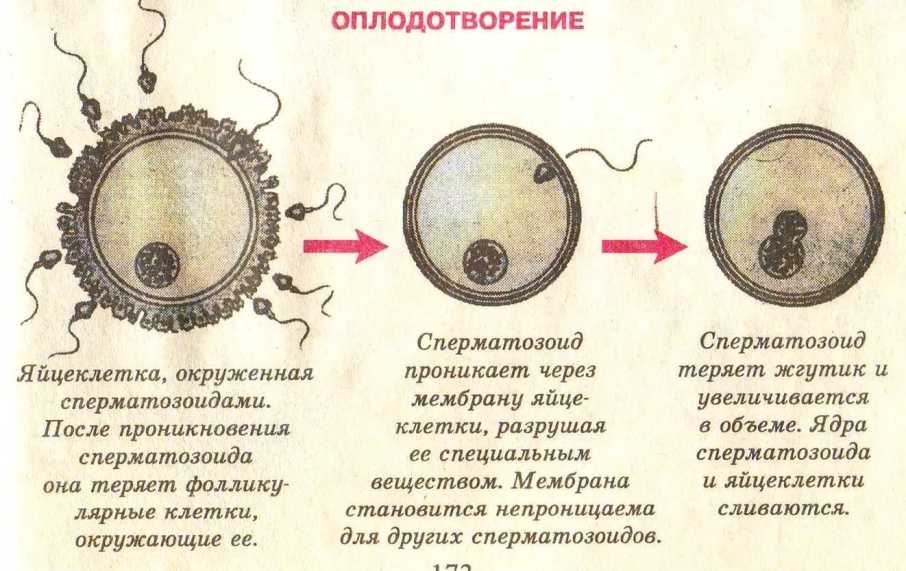 «Идея о том, что у вас может быть такая маленькая молекула, которая поддерживается в градиенте внутри этой восьмиядерной структуры только за счет синтеза, ошеломляет».
«Идея о том, что у вас может быть такая маленькая молекула, которая поддерживается в градиенте внутри этой восьмиядерной структуры только за счет синтеза, ошеломляет».
Значение для эволюции цветковых растений
Развитие зародышевого мешка, возможно, является ключевым элементом в эволюции от голосеменных к цветковым растениям, также известным как покрытосеменные.
Тем не менее летопись окаменелостей очень мало говорит о стадиях, которые привели от производства семян голосеменных к производству семян покрытосеменных, когда переход произошел около 135 миллионов лет назад. Быстрое распространение цветковых растений и их окончательное господство над земной растительностью Дарвин назвал «отвратительной тайной».
Выяснив механизм развития зародышевого мешка, Сундаресан и его команда открыли двери для новой работы по изучению эволюционного пути между этими двумя основными группами растений. Открытие подтверждает так называемую модульную теорию, которая утверждает, что первые покрытосеменные претерпели резкое сокращение женской репродуктивной единицы по сравнению с голосеменными, что позволило цветковым растениям воспроизводиться более эффективно и в конечном итоге вытеснить своих предков с голыми семенами.
Наиболее примечательно, пожалуй, то, что новая работа предполагает, что восемь ядер зародышевого мешка покрытосеменных сохранили пластичность развития в своей эволюции от голосеменных. «Удивительно, что хотя раскол предположительно произошел более ста миллионов лет назад, — сказал Сундаресан, — все эти ядра все еще способны стать яйцеклетками».
В исследовании принимали участие ведущий автор Габриэла Пагнуссат и Моника Аландете-Саез, которые были исследователями с докторской степенью в Sundaresan, когда они выполняли эту работу, и Джон Л. Боуман, профессор биологии растений в Калифорнийском университете в Дэвисе на момент исследования. сейчас в Университете Монаша в Мельбурне, Австралия.
Работа выполнена при поддержке грантов Национального научного фонда.
О Калифорнийском университете в Дэвисе
В течение 100 лет Калифорнийский университет в Дэвисе занимается обучением, исследованиями и общественной работой, которые важны для Калифорнии и преображают мир.