
Содержание
Сравнительная характеристика высших и низших растений.
1) тело низших может быть одноклеточным
колониальным или многоклеточным и
называться таллом–
слоевище. Поэтому часто низшие называют
талломные или слоевищные.
2) низшие не имеют дифференциации клеток
на ткани.
3) тело низших растений не дифференцируются
на органы.
ИсторияЗемли
Земля возникла 4-5 млрд. лет тому назад.
Сине — зеленые водоросли 1 организмы.
Органическая жизнь возникла 2,5 млрд.
лет назад. Водные организмы – 600 млн.
лет назад. Растения вышли на сушу – 400
млн. лет назад. Покрытосемянные возникли
– 150-200 млн. лет назад. Человеческое
общество возникло 1 млн. лет назад.
Выход растений на сушу.
Риниофиты = псилофиты – первенцы высших
растений.
Жизнь зародилась 600 млн. лет назад в
протерозойскую эру. Выход растений на
сушу в силурийский период ………. эры
400 млн. лет назад.
Палеоботаники реконструировали облик:
Ныне живет бурая водоросль. pelvicia– живет на скалах. Первыми покинули
сушу риниофиты (псилофиты).
Существует современная система
высших растений (Тахтаджян А.Л.) в
основании родословного древа высших
растений находятся ископаемая группа
риниофиты, от которых отходят:
–– моховидны
–– плауновидные
–– хвощевидные
–– папоротниковидные
–– голосемянные и покрытосемянные
11.02.2005 Тема : Моховидные (Bryophyta )
Наука бриология.
Общая характеристика отдела моховидные.
Весь отдел насчитывает от 20-25 тыс.
видов. Они разделяются на 2 класс:
–– печеночники (слоевцовые)
–– листостебельные
Представитель: маршанция
Представитель: сорагнум, кукушкин
лен.
Структура
тела:
Моховидные представляют собой
многолетние растения, низкорослые
(10-15 см.) примитивно устроенные, имеют
чаще всего расчленения тела на стебель
и листья, а у маршанции только слоевище
(пластинчатая структура ), корней у мхов
нет. Вместо корней у них имеется
своеобразная структура называется
ризоиды . Не содержат хлоропластов
низшая часть стебля, участвует в
поглощении Н2О и минеральных
солей. У листостебельных стебель может
ветвится, и на нем расположены листья
(зеленые), которые имеют весьма примитивное
строение и их называютфиллодиями.
Эти “листья” одно-, двухслойные,
эпидермиса в проводящих тканях они не
имеют и в стеблях и в листьях можно
видеть следующие ткани:
–– запасающая паренхима
–– ассимиляционная паренхима
–– примитивно устроенные проводящие,
механические ткани.
Произрастают мхи не по одиночке, а
группами. Образуют более густые дернины.
Несмотря на примитивное морфологическое
строение, у этих растений имеется
сложно-устроенные половые органы.
Формируются половые органы у мхов на
верхушках или боковых веточках, среди
листочков, а у Маршанции формируется
особые структуры – подставки (и с нижней
стороны формируется половые органы.
Половые органы могут формироваться на
одном растении. Тогда они будут называться
однодомные, например: Маршанция) ; и
половые органы могут формироваться на
разных растениях, тогда в этом случае
растения будут называться двудомные:
например сфагнум, кукушкин лен.
Низшие растения, их отличия от высших растений. Царство грибы. Общая характеристика, классы. Распространение в природе. Практическое значение. Важнейшие представители
жүктеу/скачать 39.31 Kb.
|
1 2 3 4 5 6 7 8 9
Байланысты:
низшие растения
МУ к пр.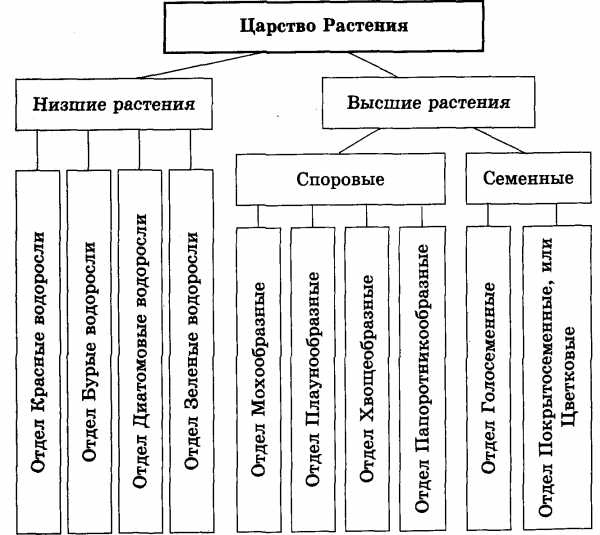 зан, Салалар тех.экол.м. пр, Лекции САПР каз, атмосфераның ластануы, низшие растения, Лекции по жиру и сыру, АҚН МУ лаб, 375972, ИП 19 8к Матем модел негіздері
зан, Салалар тех.экол.м. пр, Лекции САПР каз, атмосфераның ластануы, низшие растения, Лекции по жиру и сыру, АҚН МУ лаб, 375972, ИП 19 8к Матем модел негіздері
- Бұл бет үшін навигация:
- Низшие растения, их отличия от высших растений
- Царство Грибы
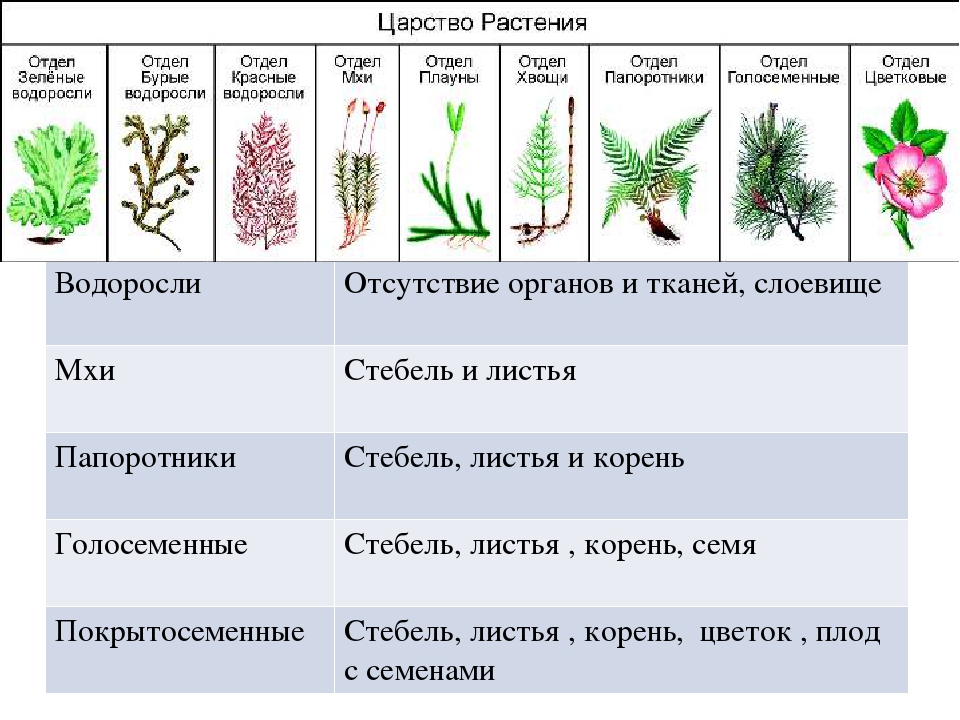
| Лекция 4 Тема: Низшие растения, их отличия от высших растений. Царство грибы. Общая характеристика, классы. Распространение в природе. Практическое значение. Важнейшие представители. Низшие растения, их отличия от высших растений К низшим растениям относятся наиболее просто устроенные представители растительного мира. Вегетативное тело низших растений не имеет расчленения на органы (стебель, лист) и представлено талломом – называют талломными Для низших растений характерно отсутствие сложной внутренней дифференцировки, у них нет анатомо-физиологической системы тканей, как у высших растений, органы полового размножения низших, одноклеточные (за исключением харовых и некоторых бурых водорослей К низшим растениям относятся Бактерии, водоросли, слизевики (миксомицеты), Грибы, лишайники.
Основное отличие низших растений от высших заключается в то, что тело низших растений (таллом, или слоевище) не расчленено на корень, стебель и лист. Царство Грибы
Общая характеристика. Грибы — царство живых организмов, которые сочетают в себе признаки растений и животных.
С растениями их сближает-. 1) наличие хорошо выраженной клеточной стенки; 2) неподвижность в вегетативном состоянии; 3) размножение спорами; 4) способность к синтезу витаминов; 5) поглощение пищи путем всасывания (адсорбции). Общим с животными является: 1) гетеротрофность; 2) наличие в составе клеточной стенки хитина, характерного для наружного скелета членистоногих; 3) отсутствие в клетках хлоропластов и фотосинтезирующих пигментов; 4) накопление гликогена как запасного вещества; 5) образование и выделение продукта метаболизма — мочевины. Эти особенности строения и жизнедеятельности грибов позволяют считать их одной их самых древних групп эукариотных организмов, не имеющих прямой эволюционной связи с растениями, как считалось ранее. Грибы и растения возникли независимо от разных форм микроорганизмов, обитавших в воде.
Известно более 100 тыс. видов грибов, причем предполагается, что реальное число их значительно больше — 250—300 тыс. и более. В мире ежегодно описывают более тысячи новых видов. Подавляющее большинство их обитает на суше, причем встречаются они практически повсеместно, где может существовать жизнь. Подсчитано, что в лесной подстилке 78—90% биомассы всех микроорганизмов приходится на долю грибной массы (примерно 5 т/га).
Грибы могут жить в различных средах — в почве, лесной подстилке, в воде, на разлагающихся и живых организмах. В зависимости от способа потребления органических веществ бывают: жүктеу/скачать 39.31 Kb. Достарыңызбен бөлісу: |
 Водоросли относятся к группе автотрофных организмов. Бактерии (за редким исключением), миксомицеты и грибы представляют собой гетеротрофные организмы, нуждающиеся в готовом органическом веществе. Те и другие как бы дополняют друг друга. Водоросли служат основными образователями органического вещества в водоемах. Разложение органических веществ и их минерализация осуществляются в результате деятельности гетеротрофных организмов: бактерий и грибов. Благодаря процессам разложения органических веществ атмосфера пополняется углекислым газом. Некоторые почвенные бактерии и сине-зеленые водоросли способны связывать свободный азот атмосферы. Таким образом, биологический круговорот веществ, совершаемый автотрофными и гетеротрофными организмами, немыслим без деятельности низших растений. По широкому распространению в природе и по численности низшие растения превосходят высшие.
Водоросли относятся к группе автотрофных организмов. Бактерии (за редким исключением), миксомицеты и грибы представляют собой гетеротрофные организмы, нуждающиеся в готовом органическом веществе. Те и другие как бы дополняют друг друга. Водоросли служат основными образователями органического вещества в водоемах. Разложение органических веществ и их минерализация осуществляются в результате деятельности гетеротрофных организмов: бактерий и грибов. Благодаря процессам разложения органических веществ атмосфера пополняется углекислым газом. Некоторые почвенные бактерии и сине-зеленые водоросли способны связывать свободный азот атмосферы. Таким образом, биологический круговорот веществ, совершаемый автотрофными и гетеротрофными организмами, немыслим без деятельности низших растений. По широкому распространению в природе и по численности низшие растения превосходят высшие.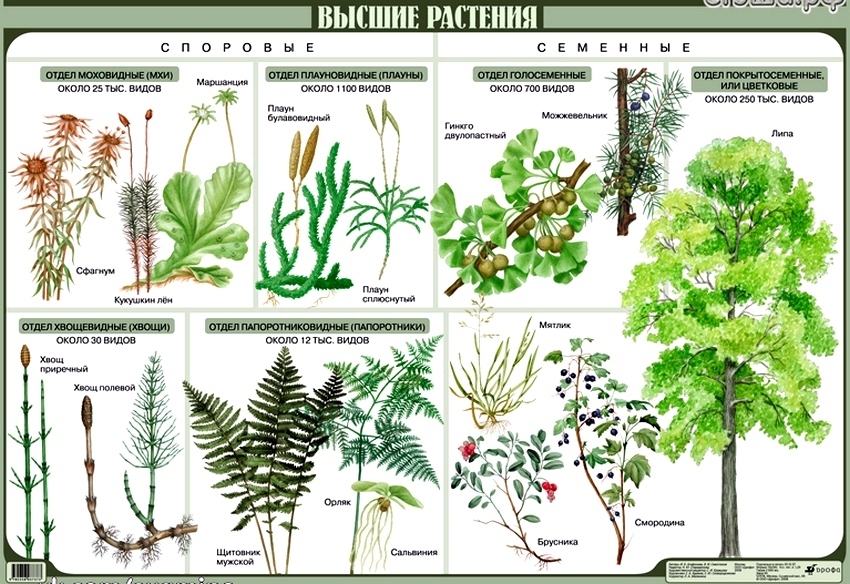

1 2 3 4 5 6 7 8 9
©melimde.com 2022
әкімшілігінің қараңыз
Характеристики водопользования растений пяти доминирующих видов кустарников в Нижней долине Рио-Гранде, штат Техас, США: значение для восстановления и сохранения кустарников
1. Арчер С. (1989) Были ли саванны южного Техаса преобразованы в леса в новейшей истории?
Арчер С. (1989) Были ли саванны южного Техаса преобразованы в леса в новейшей истории?
Я Нат
134: 545–561. [Google Scholar]
2. Archibald RD, Harper RJ, Fox JED, Silberstein RP. (2006) Производительность деревьев и накопление солей в корневой зоне на трех засушливых австралийских плантациях. Агрофорес Сист
66: 191–204. [Google Scholar]
3. Berger A, Grouzis M, Fournier C. (1996) Состояние воды шести древесных видов, сосуществующих в Сахеле (Ферло, Сенегал). Джей Троп Эколь
12: 607–627. [Google Scholar]
4. Боларин М.С., Фернандес Ф.Г., Крус В., Куартеро Дж. (1991) Устойчивость к засолению у четырех дикорастущих видов томатов с использованием вегетативных кривых урожай-засоленность. J Am Soc Hort Sci
116: 286–290. [Google Scholar]
5. Бойер Дж.С. (1995) Измерение водного статуса растений и почв. Академик Пресс, Сан-Диего. [Академия Google]
6. Браун Дж. Р., Арчер С. (1990) Водные отношения многолетних трав и сеянцев по сравнению со взрослыми древесными растениями в субтропической саванне, Техас, США. Ойкос
Ойкос
57: 366–374. [Google Scholar]
7. Браун Дж. Р., Арчер С. (1999) Вторжение кустарников в пастбища: пополнение происходит непрерывно и не регулируется биомассой и плотностью травянистых растений. Экология
80: 2385–2396. [Google Scholar]
8. Бруньоли Э., Бьоркман О. (1992) Рост хлопчатника в условиях постоянного засоления: влияние на структуру распределения, устьичные и неустьичные компоненты фотосинтеза и рассеивание избыточной световой энергии. Планта
187: 335–347. [PubMed] [Академия Google]
9. Bucci SJ, Scholz FB, Godstein G, Meinzer FC, Hinojosa JA, Hoffmann WA, Franco AC. (2004) Процессы, препятствующие ночному уравновешиванию водного потенциала листьев и почвы у древесных видов тропической саванны. Физиол дерева
24: 1119–1127. [PubMed] [Google Scholar]
10. Chaves MM, Maroco JP, Pereira JS. (2003) Понимание реакции растений на засуху — от генов до всего растения. Функциональная биология растений
30: 239–264. [Google Scholar]
11. Кондит Р.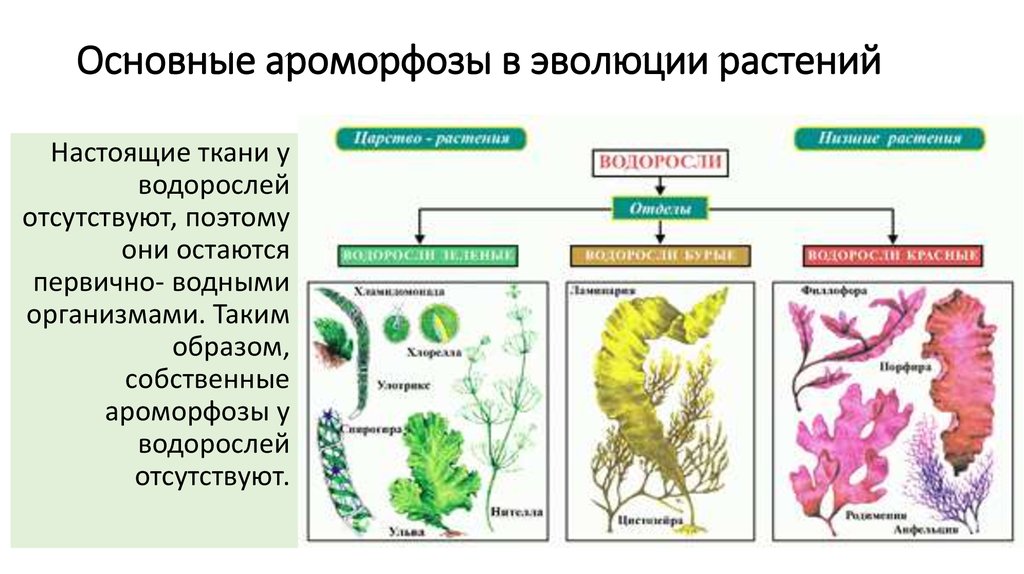 , Хаббелл С.П., Фостер Р.Б. (1995) Показатели смертности 205 видов неотропических деревьев и кустарников и последствия сильной засухи. Эколь Моногр
, Хаббелл С.П., Фостер Р.Б. (1995) Показатели смертности 205 видов неотропических деревьев и кустарников и последствия сильной засухи. Эколь Моногр
65: 419–439. [Google Scholar]
12. Dai Z, Edwards GE, Ku MSB. (1992) Контроль фотосинтеза и устьичной проводимости у Ricinus communis L. (клещевины) за счет дефицита давления паров воздуха между листьями и воздухом. Завод Физиол
99: 1426–1434. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Davies WJ. (1977) Реакция устьиц на водный стресс и свет у растений, выращенных в контролируемых условиях и в полевых условиях. Растениеводство
5: 735–740. [Академия Google]
14. Дойер Л.М., Стюарт Д.В. (1985) Кондиционирование кукурузы при водном стрессе ( Zea mays ) в поле и в теплице. Может Джей Бот
63: 704–710. [Google Scholar]
15. де Сойза А.Г., Киллингбек К.Т., Уитфорд В.Г. (2004) Водные отношения растений и фотосинтез во время и после засухи в арройо пустыни Чиуауа. J Засушливая среда
59: 27–39. [Google Scholar]
[Google Scholar]
16. Дитмарова Л., Куряк Д., Палмрот С., Кмет Дж., Стрелцова К. (2009) Физиологические реакции ели европейской ( Picea abies ) сеянцы к стрессу от засухи. Физиол дерева
30: 205–213. [PubMed] [Google Scholar]
17. Донаван Л.А., Элерингер Дж.Р. (1991) Экофизиологические различия между ювенильными и репродуктивными растениями нескольких древесных видов. Экология
86: 594–597. [PubMed] [Google Scholar]
18. Донаван Л.А., Элерингер Дж.Р. (1992) Сравнение моделей водопользования среди классов размера и жизненного цикла полузасушливого кустарника. Функция Ecol
6: 482–488. [Google Scholar]
19. Донован Л.А., Гриз Д.Дж., Уэст Дж.Б., Папперт Р.А., Олдер Н.Н., Ричардс Дж.Х. (1999) Предрассветное нарушение равновесия водного потенциала растений и почвы у двух холодных пустынных кустарников. Экология
120: 209–217. [PubMed] [Google Scholar]
20. Донован Л.А., Линтон М.Дж., Ричардс Дж.Х. (2001) Предрассветный водный потенциал растений не обязательно уравновешивается с водным потенциалом почвы при хорошем поливе. Экология
Экология
129: 328–335. [PubMed] [Google Scholar]
21. Донован Л.А., Линтон М.Дж., Ричардс Дж.Х. (2003) Величина и механизмы неравновесия между предрассветным потенциалом воды растений и почвы в кустарниках пустыни. Экология
84: 463–470. [Академия Google]
22. Эдди М.Р., Джадд Ф.В. (2003) Фенология Acacia berlandieri , A. minuata , A. Rigiula , A. schaffneri и Chloroleucon ebano в Нижней долине Рио-Гранде в Техасе во время засухи. Юго-Западный Нат
48: 321–332. [Google Scholar]
23. Escos J, Alados CL, Pugnaire FI, Puigdefabregas J, Elmen J. (2000) Стратегия стрессоустойчивости в засушливом кустарнике: взаимодействие между нестабильностью развития и фрактальной размерностью. J Засушливая среда
45: 325–336. [Академия Google]
24. Ewing K, Best C. (2004) Эксперимент по восстановлению колючих кустарников в Тамаулипане, Южный Техас, измеряет рост посаженной древесной растительности. Экол Рез
22: 11–17. [Google Scholar]
25. Фолтеску В.Л., Прайор С.К., Беннет С. (2005) Образование, рассеивание и удаление морской соли в региональном масштабе. Атмос Окружающая среда
Фолтеску В.Л., Прайор С.К., Беннет С. (2005) Образование, рассеивание и удаление морской соли в региональном масштабе. Атмос Окружающая среда
39: 2123–2133. [Google Scholar]
26. Franco JA, Fernández JA, Bañón S, Gonzalez A. (1997) Взаимосвязь между влиянием засоления на площадь листьев сеянцев и урожайность шести сортов дыни. Дж. Хорт Наука
32: 642–647. [Академия Google]
27. Gebrehiwot K, Muys B, Haile M, Mitloehner R. (2005) Использование водного баланса растений для характеристики видов деревьев и местообитаний в засушливых районах северной Эфиопии. J Засушливая среда
60: 581–592. [Google Scholar]
28. Гебрекирстос А., Текетай Д., Фетене М., Митлонер Р. (2006) Адаптация пяти совместно встречающихся видов деревьев и кустарников к водному стрессу и его влияние на восстановление деградированных земель. Для Экол Менеджмент
229: 259–267. [Google Scholar]
29. Голлан Т., Тернер Н.К., Шульце Э.Д. (1985) Реакция газообмена устьиц и листа на дефицит давления пара и содержание влаги в почве. III. У склерофильного древесного вида Nerium olaender . Экология
III. У склерофильного древесного вида Nerium olaender . Экология
65: 356–362. [PubMed] [Google Scholar]
30. Гонсалес-Родригес Х., Канту-Сильва I, Гомес-Меса М.В., Джордан В.Р. (2000) Сезонные водные отношения растений у Acacia berlandieri . Реабилитация засушливых почв
14: 343–357. [Google Scholar]
31. Гонсалес-Родригес Х., Канту-Сильва И.С., Гомес-Меса М.В., Лосано Р.Г.Р. (2004) Водные отношения между растениями и видами кустарников терновника, северо-восток Мексики. J Засушливая среда
58: 483–503. [Академия Google]
32. Гонсалес-Родригес Х., Канту-Сильва И., Рамирес-Лозано Р.Г., Гомес-Меса М.В., Саркис-Рамирес Х., Кориа-Хил Н., Сервантес-Монтойя Х.Р., Майти Р.К. (2011) Водный потенциал ксилемы местных кустарников с северо-востока Мексики. Acta Agr Scand, Раздел B — Наука о почвенных растениях
61: 214–219. [Google Scholar]
33. Gowing DJG, Jones HG, Davies WJ. (1993) Абсцизовая кислота, переносимая ксилемой: относительная важность ее массы и концентрации в контроле устьичного отверстия. Окружающая среда растительной клетки
Окружающая среда растительной клетки
16: 453–459. [Google Scholar]
34. Grime JP. (1977) Доказательства существования трех основных стратегий у растений и их отношение к экологической и эволюционной теории. Я Нат
111: 1169–1194. [Google Scholar]
35. Hendrickx JMH, Harrison JB, Beekma J, Rodribuez-Marin G. (1999) Управление соленостью в Рио-Гранде-Боске. Протокол Лесной службы Министерства сельского хозяйства США RMRS-P-7, стр.
68–71. [Google Scholar]
36. Ховард А.Р., Ван Ирсел М.В., Ричардс Дж.Х., Донован Л.А. (2009) Транспирация в ночное время может уменьшить гидравлическое перераспределение. Окружающая среда растительной клетки
32: 1060–1070. [PubMed] [Академия Google]
37. Iyengar ERR, Reddy MP. (1996) Фотосинтез у растений с высокой солеустойчивостью. В Пессаракли М., изд. Справочник по фотосинтезу. Marcel Decker, Inc., Батон Роуз, США, стр. 56–65. [Google Scholar]
38. Ярсдоерфер С.Е., Лесли Д.М. (1988) Кустарниковые угодья Тамаулипан в Нижней долине Рио-Гранде в Техасе: описание, воздействие человека и варианты управления.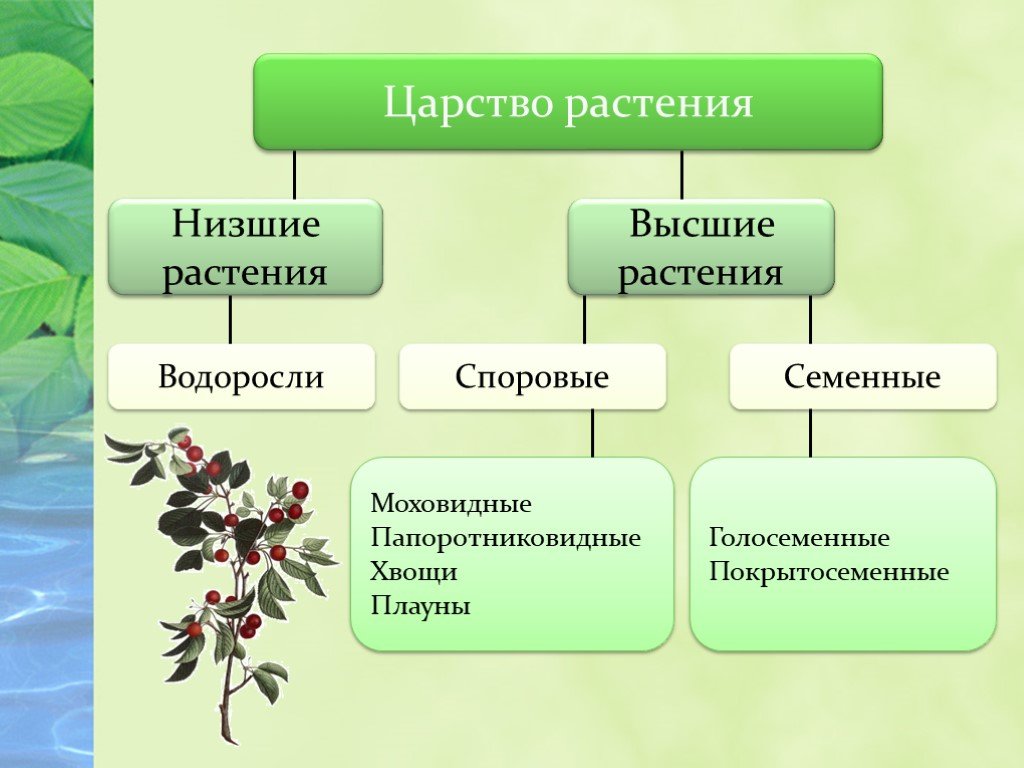 Биологический отчет 88
Биологический отчет 88
Служба рыболовства и дикой природы США, Министерство внутренних дел США. [Google Scholar]
39. Кнапп А.К., Фанесток Дж.Т. (1990) Влияние размера растения на соотношение углерода и воды у Cucurbita foetidissima HBK. Функция Ecol
4: 789–797. [Google Scholar]
40. Козловский Т.Т. (1997) Реакция древесных растений на затопление и засоление. Дерево Физиол Моногр
1: 1–29. [Google Scholar]
41. Козловский Т.Т., Палларди С.Г. (2002) Акклиматизация и адаптивные реакции древесных растений на стрессы окружающей среды. Бот Рев
68: 270–334. [Google Scholar]
42. Маас Э.В., Хоффман Г.Дж. (1977) Солеустойчивость сельскохозяйственных культур – текущая оценка. J Irrig Drain Eng
103: 115–134. [Академия Google]
43. Маккой Т.В. (1990) Оценка ресурсов подземных вод в Нижней долине Рио-Гранде, Техас. Texas Water Development Board, Report 316, 48 pp. [Google Scholar]
44. McDowell NG, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, et др.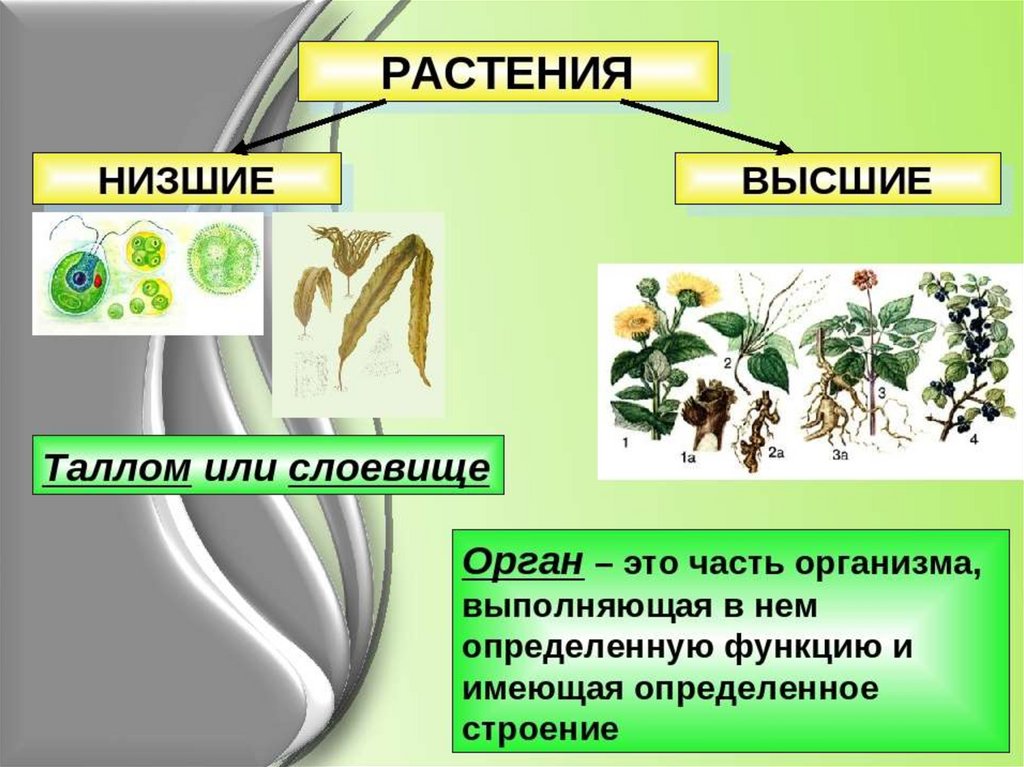 (2008) Механизмы выживания и гибели растений во время засухи: почему одни растения выживают, а другие погибают от засухи?
(2008) Механизмы выживания и гибели растений во время засухи: почему одни растения выживают, а другие погибают от засухи?
Новый Фитол
178: 719–739. [PubMed] [Google Scholar]
45. Medrano H, Escalona JM, Bota J, Gulías J, Flexas J. (2002) Регуляция фотосинтеза C 3 растений к прогрессирующей засухе: устьичная проводимость как эталонный параметр. Энн Бот
89: 895–905. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Montagu KD, Woo KC. (1999) Восстановление способности деревьев к фотосинтезу в результате сезонной засухи во влажно-сухих тропиках: роль филлодии и процессов полога у Acacia auriculiformis . Ауст Дж Завод Физиол
26: 135–145. [Google Scholar]
47. Morgan JA, LeCain DR, Pendall E, Blumenthal D, Kimball BA, Carrillo Y, Williams DG, Heisler-White J, Dijkstra FA, West M. (2011) C 4 травы процветают, поскольку углекислый газ устраняет высыхание на прогретых полузасушливых пастбищах. Природа
476: 202–205. [PubMed] [Google Scholar]
48. Морган Дж.М. (1984) Осморегуляция и стресс от засухи у высших растений. Анну Рев Растение Физиол
Морган Дж.М. (1984) Осморегуляция и стресс от засухи у высших растений. Анну Рев Растение Физиол
35: 299–319. [Google Scholar]
49. Майерс Б.А., Нилс Т.Ф. (1984) Сезонные изменения водного режима Eucalyptus behriana F. Muell и E. microcarpa (Maiden) в полевых условиях. Бот Aust J Plant
32: 495–510. [Академия Google]
50. Навар-Чайдес JDJ. (2008) Потоки углерода в результате изменений в землепользовании в терновнике Тамаулипан на севере Мексики. Управление углеродным балансом
3:6
doi:10.1186/1750-0680-3-6 [бесплатная статья PMC] [PubMed] [Google Scholar]
51. Ноорметс А., Собер А., Пелл Э.Дж., Диксон Р.Е., Подила Г.К., Собер Дж., Изебрандс Дж.Г., Карноски Д.Ф. . (2001) Устьичные и неустьичные ограничения фотосинтеза у двух клонов осины дрожащей ( Populus tremuloides Michx.), подвергшихся воздействию повышенного содержания CO 2 и/или О 3 . Окружающая среда растительной клетки
24: 327–336. [Google Scholar]
52. Otieno DO, Schmidt MWT, Adiku S, Tenhaunen J. (2005) Физиологические и морфологические реакции на водный стресс у двух видов Acacia из контрастных местообитаний. Физиол дерева
Otieno DO, Schmidt MWT, Adiku S, Tenhaunen J. (2005) Физиологические и морфологические реакции на водный стресс у двух видов Acacia из контрастных местообитаний. Физиол дерева
25: 361–371. [PubMed] [Google Scholar]
53. Ourcival JM, Floret C, Le Floch E, Pontanier R. (1994) Водные отношения между двумя многолетними видами в степях южного Туниса. J Засушливая среда
28: 333–350. [Академия Google]
54. Парида А.К., Дас А.Б., Моханти П. (2004) Защитные потенциалы против NaCl в мангровых зарослях, Bruguiera parviflora : дифференциальные изменения изоформ некоторых антиоксидантных ферментов. J Растение Физиол
161: 531–542. [PubMed] [Google Scholar]
55. Порпорато А., Лайо Ф., Ридольфи Л., Родригес-Итурбе И. (2001) Растения в водорегулирующих экосистемах: активная роль в гидрологических процессах и реакция на водный стресс III. Водный стресс растительности. Adv Water Resour
24: 725–744. [Академия Google]
56. Рейд Н., Маррокин Дж., Бейер-Мунцель П. (1990) Использование кустарников и деревьев для покоса, топливной древесины и древесины в терновниках Тамаулипана, северо-восток Мексики. Для Экол Менеджмент
(1990) Использование кустарников и деревьев для покоса, топливной древесины и древесины в терновниках Тамаулипана, северо-восток Мексики. Для Экол Менеджмент
36: 61–79. [Google Scholar]
57. Ritchie GA, Hinckley TM. (1975) Барокамера как инструмент экологических исследований. Ад Экол Рез
9: 165–254. [Google Scholar]
58. Saliendra NZ, Sperry JS, Comstock JP. (1995) Влияние состояния воды в листьях на реакцию устьиц на влажность, гидравлическую проводимость и засуху почвы в Бетула западная . Планта
196: 357–366. [Google Scholar]
59. Саллео С., Нардини А., Питт Ф., Ло Гулло М.А. (2000)Кавитация ксилемы и гидравлический контроль устьичной проводимости у лавра ( Laurus nobilis L.). Окружающая среда растительной клетки
23: 71–79. [Google Scholar]
60. Sellin A. (1996) Базовый водный потенциал Picea abies как характеристика водного статуса почвы. Растительная почва
184: 273–280. [Google Scholar]
61. Tardieu F, Davies WJ.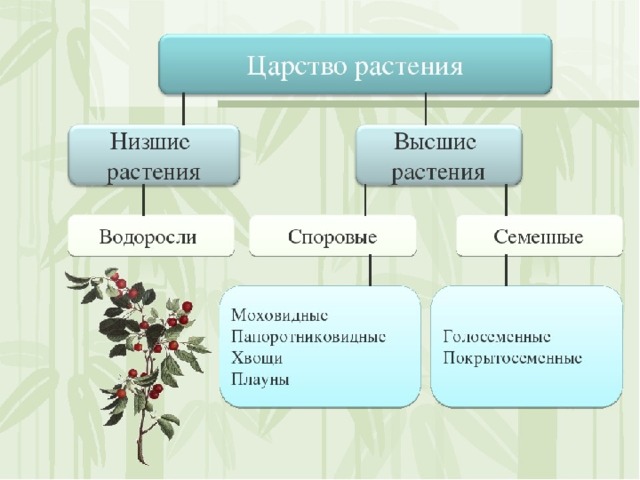 (1992) Реакция устьиц на АБК зависит от текущего состояния воды в растении. Завод Физиол
(1992) Реакция устьиц на АБК зависит от текущего состояния воды в растении. Завод Физиол
98: 540–545. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Тезара В., Митчелл В.Дж., Дрисколл С.Д., Лоулор Д.В. (1999) Водный стресс ингибирует фотосинтез растений за счет уменьшения фактора связи и АТФ. Природа
401: 914–917. [Google Scholar]
63. Trejo CL, Davies WJ, Ruiz L del MP. (1993) Чувствительность устьиц к абсцизовой кислоте: влияние мезофилла. Завод Физиол
102: 497–502. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Вертовец М., Сакчали С., Озтурк М., Саллео С., Джакомич П., Феоли Э., Нардини А. (2001) Диагностика водного статуса растений как инструмент количественной оценки водного стресса на региональной основе в засушливых районах Средиземноморья. Энн для науки
58: 113–125. [Google Scholar]
65. Wan C, Sosebee RE. (1991) Водные отношения и транспирация медового мескитового дерева на 2 участках в западном Техасе. J Управление диапазоном
44: 156–160. [Google Scholar]
[Google Scholar]
66. Уайт Дж. Д., Гуцвиллер К. Дж., Барроу К. В., Рэндалл Л. Дж., Свинт П. (2008) Моделирование механизмов изменения растительности из-за пожара в полузасушливой экосистеме. Эколь Модель
214: 181–200. [Академия Google]
67. Виденфельд Б. (2008) Влияние солености оросительной воды и электростатической обработки воды на производство сахарного тростника. Агроводоуправление
95: 85–88. [Google Scholar]
Мохообразные, папоротники и союзники папоротников
Мохообразные, папоротники и союзники папоротников
Лаборатория 8 — Примитивные растения —
Мохообразные,
Папоротники и союзники папоротников
Введение в мохообразные — мхи, печеночники,
и роголистники
При переходе от мхов к папоротникам мы видим постепенный переход от
примитивный
к современным чертам. Есть две основные тенденции, на которые следует обратить внимание.
сегодняшний
лаборатория Первое – переход в жизненных циклах, второе – смена
в
основная внутренняя структура.
Во-первых, все растения претерпевают смену поколений ,
между стадией гаплоидного гаметофита и диплоидным спорофитом
сцена. У самых примитивных растений, таких как мхи, гаметофит
доминирующий
(то есть он большой и зеленый). У высших растений, таких как папоротники и их союзники,
стадия спорофита является доминирующей. Гаметофиты образуют гаметы (сперматозоиды).
а также
яйца) в специальной структуре, называемой гаметангий (-ia), а
спорофиты
производят споры в особой структуре, называемой спорангием
(-я).
Во-вторых, все растения нуждаются в воде для своих клеток.
Примитивный
мохообразные
например, мхи и печеночники настолько малы, что могут полагаться на диффузию
для подачи и отвода воды из растения. У мхов есть несколько нитей
вода
проводящая ткань в их центральном стебле, но ничего похожего на большую и
хорошо организованная сеть трубок в трахеофиты , или «трубчатые
растения».
сосудистых тканей у более продвинутых папоротников и «папоротниковых
союзники»
состоят из ксилемы и флоэмы, проводящих воду,
питательные вещества,
и питание по всему телу растения. Мы рассмотрим эти ткани в
потом
лаборатория
Мохообразным для размножения также необходима влажная среда. Их
жгутик
сперматозоиды должны плыть по воде, чтобы добраться до яйцеклетки. Итак, мхи и
печеночники
ограничены влажными местообитаниями. В пустыне нет мхов.
Но
мхи на удивление устойчивы к высыханию и могут выживать под
очень
суровые условия. Мхи являются наиболее многочисленными растениями в обоих
Арктический
и Антарктика. Бесполое размножение у мохообразных осуществляется
по
фрагментацией или крошечными вегетативными «ростками», называемыми гемма ,
который
формируются в специальные небольшие структуры, называемые чашечками gemmae .
Мхи и печеночники объединены в мохообразные ,
растения
лишены настоящих сосудистых тканей и имеют ряд других примитивных
черты. У них также отсутствуют настоящие стебли, корни или листья, хотя они
У них также отсутствуют настоящие стебли, корни или листья, хотя они
клетки
которые выполняют эти общие функции. Листовое зеленое растение, которое мы видим
когда мы смотрим на мох или печеночницу, это действительно гаметофит ,
которая является доминирующей стадией у всех мохообразных. спорофиты
из
Мохообразные не ведут свободноживущего существования. Они растут прямо из
оплодотворенной яйцеклетки в архегониях и остаются зависимыми от
родитель
гаметофит для их питания.
Характеристики разделов
Отдел Bryophyta — (9500 sp.) — мхи, Mnium, Sphagnum
Мхи бывают двух основных типов: мягкий тип с прямостоячими стеблями,
и перистый тип, который образует уплощенные маты низменных и высоко расположенных
ветвистые моховые растения. В обоих случаях лиственные зеленые гаметофиты
раздельнополый
Это могут быть мужские растения с антеридиями на верхушке или
женский
растения с архегониями наверху. Помните, что эти гаметофиты
находятся
всегда гаплоидные (1N) растения.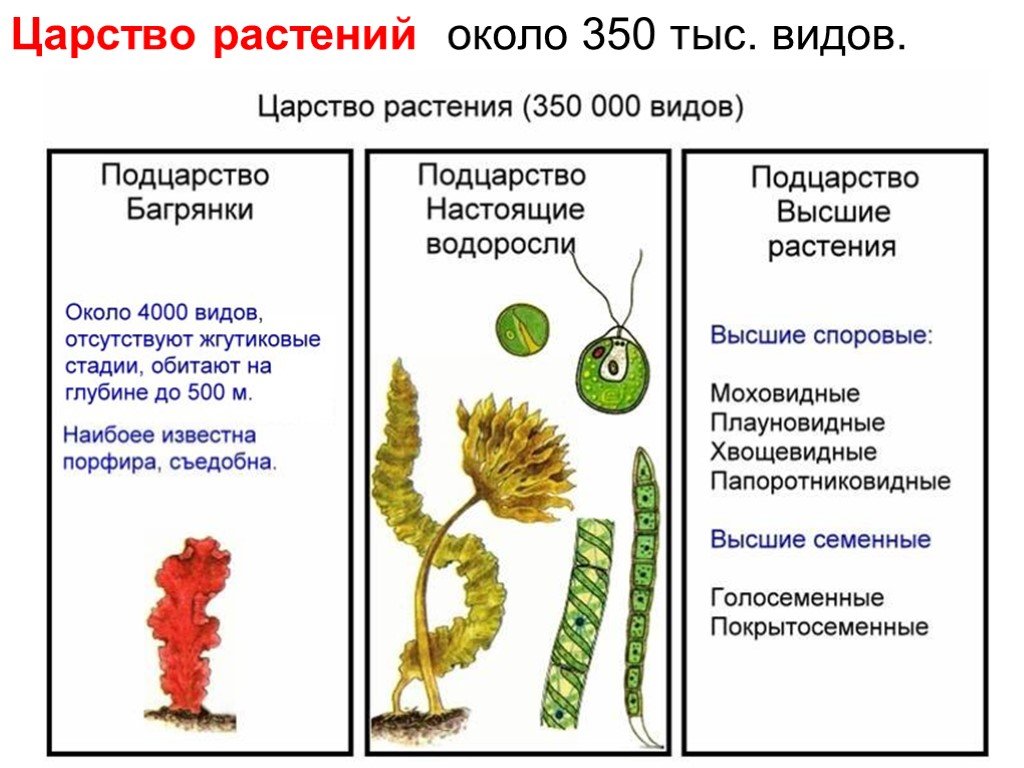
Сперматозоиды образуются в каждом антеридии , а яйцеклетка — в
каждый
архегоний .
Поскольку растение уже гаплоидно, эти гаметы могут быть созданы
митоз,
простое деление клеток. Сперматозоид подплывает к архегониям через тонкую
водная пленка, нарисованная химическим аттрактантом, вырабатываемым самкой
завод,
затем плывет по шее архегония к яйцу. доброе утро
роса
воды более чем достаточно для плавания сперматозоидов. Как только сперма
входит
архегоний, он сливается с яйцом. Зигота 2N развивается в
диплоидный
спорофитное растение, небольшой стебель, растущий прямо из верхушки
в
архегоний. Этот стебель изначально зеленый и фотосинтезирующий, но
потом
становится коричневым и становится паразитом на самке
гаметофит.
Спорофитное растение состоит из стебля и небольшой коробочки
на вершине. Внутри капсулы клетки подвергаются мейозу с образованием
тетрады
гаплоидных спор . Когда капсула созреет, ее откидная крышка или
жаберная крышка
раскрывается, и споры быстро разносятся ветром и водой.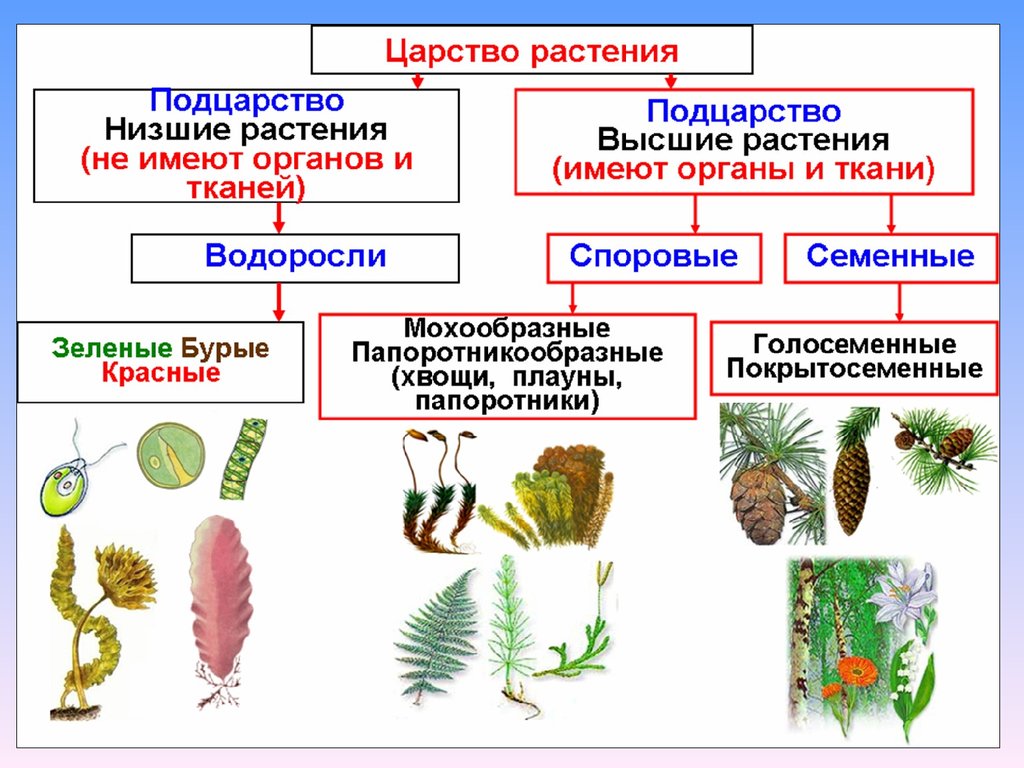
споры
прорастают в крошечную зеленую нить, которая выглядит как простая прядь
зеленые водоросли. Это сходство является еще одним признаком того, что мохообразные
спустился
из зеленых водорослей. Эта ранняя нитевидная стадия называется 9-й.0191 протонема
(= первая нить, множественное число = протонемы, такие как стигмы/стигматы). Новый
взрослый
гаметофиты вырастают из крошечной почки, которая развивается на протонеме.
В итоге
эти гаметофиты будут расти, чтобы производить гаметы, и весь цикл
будут
начать заново. Мхи также могут размножаться бесполым путем путем фрагментации.
или путем выращивания маленьких вегетативных почек, называемых геммой, которые могут отламываться.
и вырасти в новое растение.
Хотя мохообразные в целом более интересны, чем важны, в
обычном смысле, заметным исключением являются мхи рода Сфагнум .
Сфагнум
мох образует плотные маты, которые спрессовываются в торф, который можно
использовал
в качестве топлива, хотя и сильно дымит. Торф также содержит другие растения, такие как
как тростник, растущий среди сфагнума.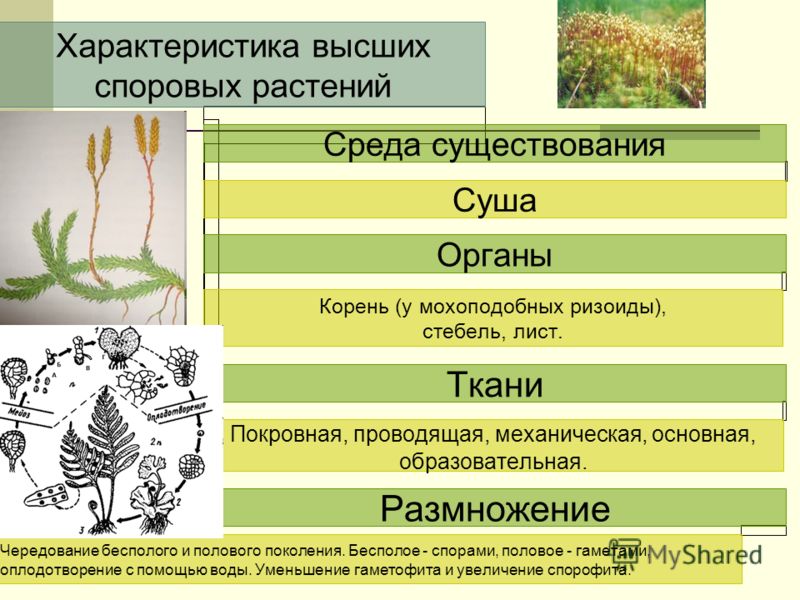 В высушенном виде торфяной мох
В высушенном виде торфяной мох
удивительно
абсорбент и, и использовался для подгузников, для обогащения бедного сада
почвы, а также в качестве полевой повязки на раны. В то время как хлопок впитывает 4-6
раз
своего сухого веса, высушенный сфагнум может поглощать в 20 раз больше собственного веса в
жидкости!
Торфяные болота представляют собой очень важные и интересные экосистемы. Сфагнум
мхи значительно повышают местную кислотность, выделяя ионы Н+, а рН
торфяных болот может упасть до 4 и ниже, что, возможно, является самой кислой природной
Окружающая среда.
Торфяные болота покрывают около 1% земной поверхности, площадь около
половина
размер Соединенных Штатов.
Отдел Hepaticophyta — (9000 экз.), печеночники, Marchantia,
Коноцефалум, Порелла
У печеночников самое простое тело из всех зеленых растений.
гаметофит,
доминантная стадия имеет вид плоского чешуйчатого листа с выступающими лопастями.
Он выглядит для всего мира как крошечная приплюснутая печень, отсюда
научный
название hepatico-phyta = растение печени.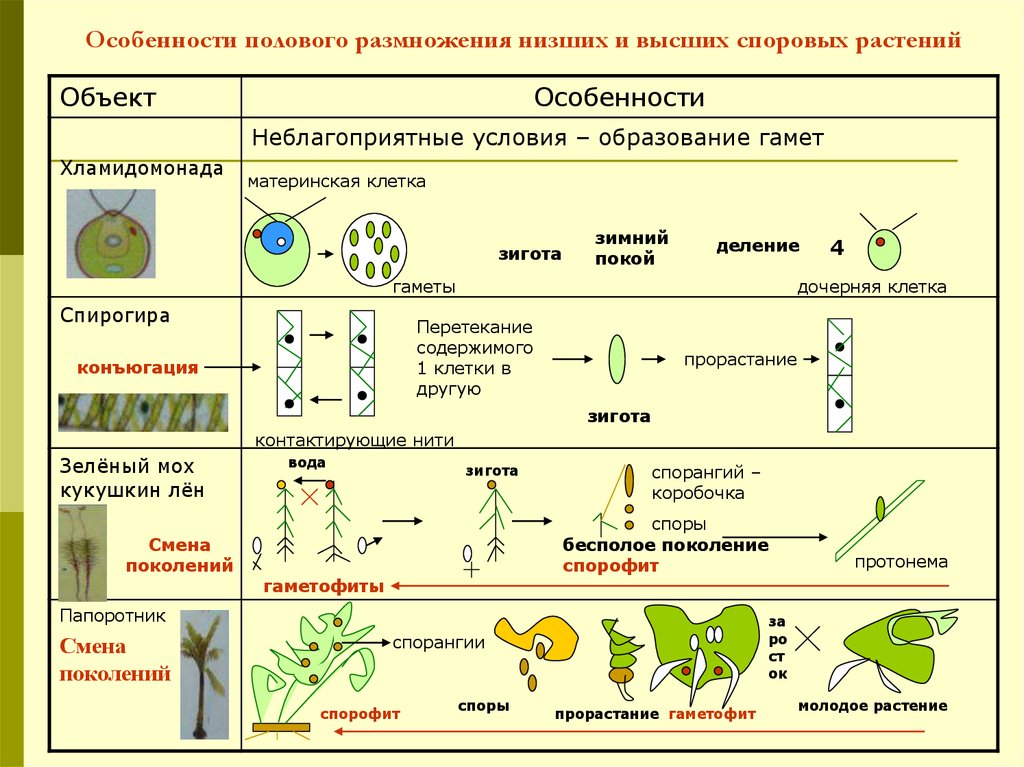 В средние века это
В средние века это
сходство
заставили врачей назначать печеночницу при заболеваниях печени.
В соответствии
к Учению Подписей Творец устроил всю природу,
включая
растения, с заботой о нашем благополучии. Люди верили, что растения
намеренно спроектированы так, чтобы напоминать органы тела, которыми они были
предполагаемый
Лечить! Отсюда печеночное сусло, а вирт — это англосаксонское слово, обозначающее траву.
форма печеночника была подписью Творца в природе. Можно
Вы догадались, что грецкие орехи должны были вылечить? (болезни
мозг)
Печеночники обладают общими свойствами мохообразных, но не
очень
близкие к мхам или роголистникам. Многие ботаники считают, что они могут
имеют
произошли независимо от другой группы зеленых водорослей. Для одного
вещь,
печеночники хранят пищу не в виде крахмала, а в виде масел. Если вы получите
водный
печеночница Porella в лаборатории, понюхайте банку, но не слишком
глубокий!
Пахнет прогорклыми маслами, маслами, которые немного испортились, пока растение
был отправлен. Еще одной характерной чертой печеночников является их
Еще одной характерной чертой печеночников является их
отсутствие устьиц, которые есть у всех других растений, в том числе у мхов
и роголистники.
У многих видов печеночников, таких как Marchantia , один
ты
скорее всего увижу в лаборатории, антеридии и архегонии не на
Топ
растения, но свисающие с нижней стороны странных маленьких
структуры
которые выглядят как крошечные зонтики. (Эти зонтиковидные структуры
называется
антеридиофор и архегониофор). Двужгутиковые сперматозоиды
плавает
к яйцеклетке, и происходит оплодотворение с образованием диплоидного (2N)
зигота.
Крошечные диплоидные спорофиты, которые остаются прикрепленными к родительскому
завод,
имеют очень простую структуру. Мейоз внутри спорофита дает
количество гаплоидных спор. Эти споры окружены любопытными длинными
а также
скрученные влажные клетки, называемые элатеры. Когда капсула высыхает и лопается,
элатеры крутятся и дергаются так, что споры рассеиваются в
все направления. Печеночники также могут размножаться бесполым путем с помощью
специальный
структуры под названием чашки из драгоценных камней . Эти маленькие чашки можно легко
Эти маленькие чашки можно легко
видимый
на поверхности растения. Каждая чашечка геммы содержит ряд крошечных
проростки
называются геммами, и одна капля воды рассеет их. Эти
маленький
затем вегетативные «клоны» вырастут в новый гаметофит.
Отдел Anthocerophyta — (100 видов), роголистники, Anthoceros
Зеленые гаметофиты роголистника очень похожи на
печеночница.
Но их мелкие спорофиты больше напоминают спорофиты мхов.
спорофиты вырастают из гаметофита и выглядят как немного прямостоячие
рог. Как и у мхов, у роголистников есть устьица, и поэтому они, вероятно, более
тесно
связаны со мхами и другими растениями, чем с печеночниками, которые они матируют
напоминать.
Эти растения симбиотичны с цианобактериями Носток .
цианобактерии
фиксирует азот для роголистника.
Таксономия
Царство растений
Мохообразные
Отдел Bryophyta — мхи ( Mnium, Sphagnum ; фр.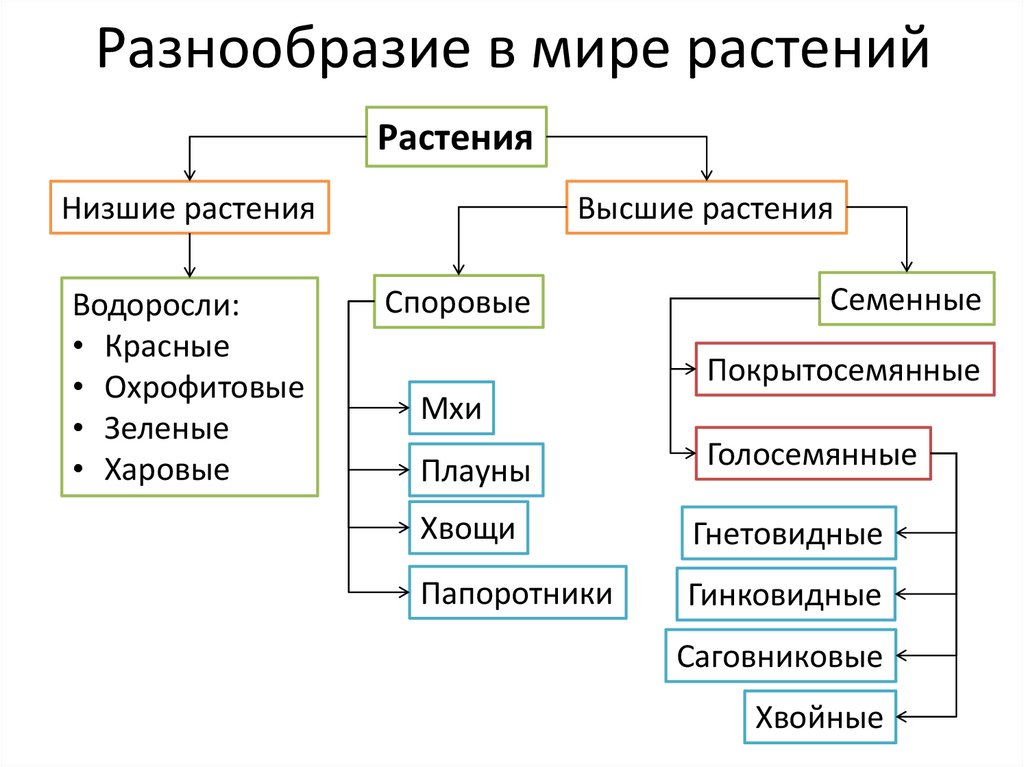 Гр.
Гр.
брион
= мох)
Отдел Hepaticophyta — печеночники ( Marchantia,
коноцефалум,
Порелла; фр.Гр. гепато = печень)
Отдел Anthocerophyta — роголистники (= Anthocerotophyta; Anthoceros;
фр.Гр
anthos = цветок, keras = рог)
Условия
- смена поколений
- гаметофит
- гаметангий (-ia)
- архегоний (-ia)
- антеридий (-ia)
- спорофит
- спорангий (-иа)
- спора
- капсула
- протонема
- сосудистая система
- драгоценные камни
- чашка из драгоценных камней
Делать и просматривать
Осмотрите живые мхи на витрине. Обратите внимание на маленькие капсулы .
на крошечных спорофитах. Мхи обычно растут в одном из двух
рост
типы: мягкий мох и перистый мох .
Изучите препараты антеридий и архегоний.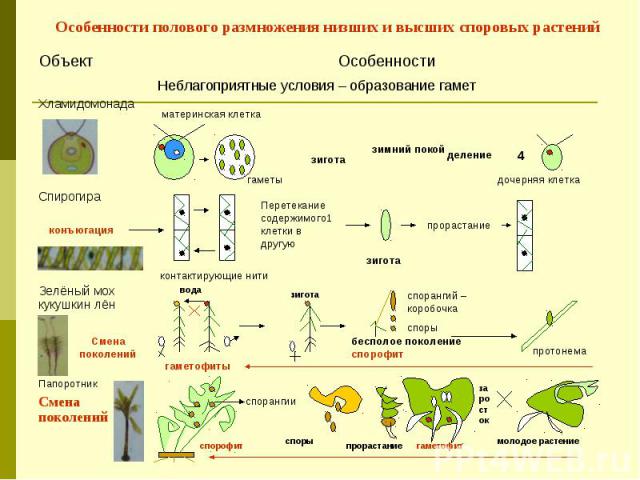 Колбаса
Колбаса
формы антеридии производят сперму, а форма колбы архегония
производит яйца.
Изучите слайды мха капсулы и определите споры .
Изучите препараты протонемы . Какой вид водорослей
делает
это вам напоминает? Это сходство является дополнительным свидетельством того, что зеленый
водоросли
дал начало всем высшим растениям.
Исследование наземных печеночников Marchantia и Conencephalum
(один
или оба должны быть на дисплее). Чем их рост отличается от
что из мхов? Ты видишь какие-нибудь чашки gemmae на верхней части
поверхность
этих растений?
Изучите препараты печеночника Marchantia.
Осмотрите водных печеночников , таких как Porella
а также
Ричча
(один или оба должны быть на дисплее). Обратите внимание, чем они отличаются от
более
наземные формы печеночника.
Посмотрите на сохранившиеся печеночники и обратите внимание на их отличительные
репродуктивные структуры (выглядят как маленькие зеленые зонтики). Как
Как
делает
их жизненный цикл отличается от мхов?
Изучите препараты роголистника Anthocerophyta .
Что нужно помнить
Знать жизненный цикл мха в деталях и уметь
распознавать
различные этапы.
Подсказка: Убедитесь, что вы понимаете общий жизненный цикл
растения,
и может сказать, какие стадии являются гаплоидными гаметофитами (1N) или диплоидными
спорофиты
(2Н). Мы изучим несколько жизненных циклов на лекции и в лаборатории (мох,
папоротник,
сосна, цветущее растение), но все они являются вариациями одного и того же
базовый
тема.
Экологическое, эволюционное и экономическое значение
Мхи важны в ландшафтном дизайне и садоводстве, особенно торф.
мох
( Сфагнум ).
Торфяной мох ( Sphagnum ) исторически использовался в качестве подкормки.
для ран.
Торфяной мох можно использовать в качестве топлива.
Мхи – самые примитивные живые наземные растения.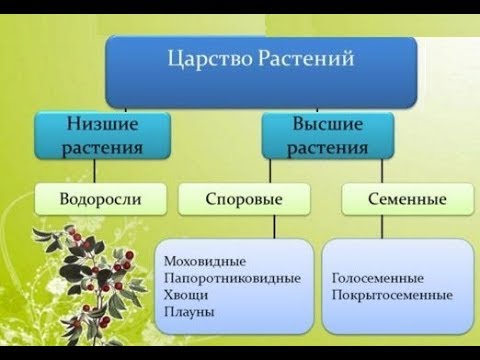
Роголистники содержат симбиотические колонии цианобактерий Nostoc .
сияющий клубный мох
Введение в трахеофиты — папоротники и их союзники
Трахеофиты ( сосудистых растения) завершили завоевание
земной поверхности, начатой более примитивными мохообразными. Так же, как
эволюция
спор был ключом к вторжению на поверхность земли
мохообразные,
изобретение сложных сосудистых тканей позволило трахеофитам завершить
в
завоевание суши. Существует около 250 000 видов сосудистых
растения,
сгруппированы в девять дивизий. Все трахеофиты имеют хорошо развитую
корневая поросль
система с высокоспециализированными корнями, стеблями и листьями, а также
специализированный
сосудистый
ткань (ксилема и флоэма), которые функционируют как миниатюрные трубки для
руководить
пища, вода и питательные вещества по всему растению. Потому что папоротник и папоротник
союзники обладают настоящими сосудистыми тканями, они могут увеличиваться в размерах
а также
толще, чем у мохообразных.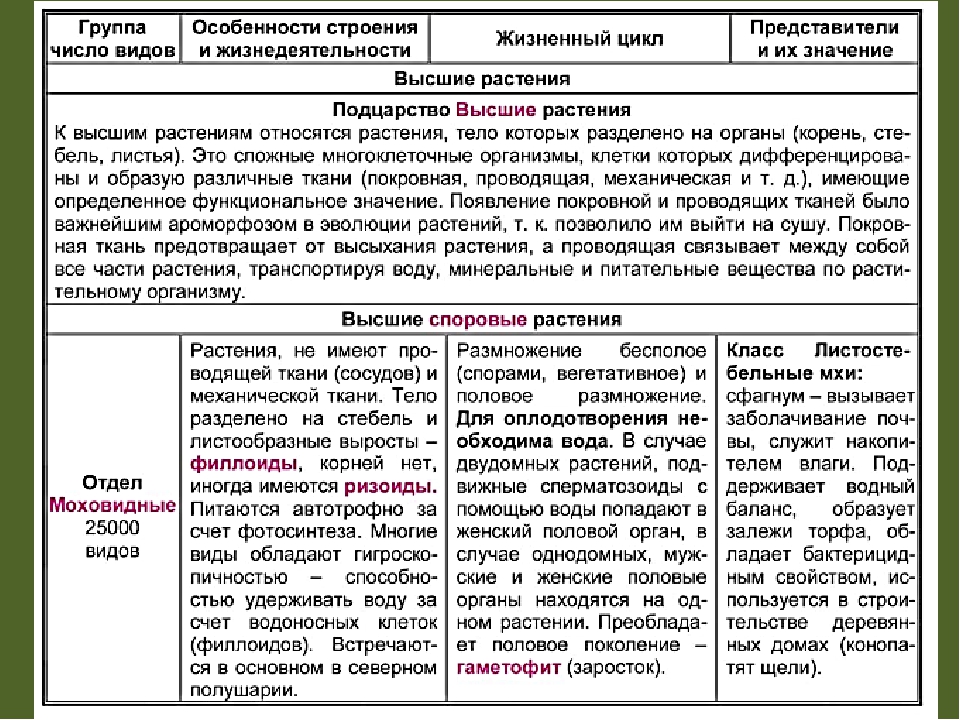
Папоротники и их союзники (несеменные трахеофиты) являются двумя основными
эволюционный
шаги. У этих и у всех более развитых растений диплоидный зеленый лист
спорофит теперь становится доминирующей стадией. Крошечный гаметофит
может быть либо автотрофным (как папоротник заросток ), либо
гетеротрофный
(как и гаметофиты некоторых плауновидных), и вообще свободноживущие
и независимо от родительского спорофита. В отличие от сосудистых
спорофиты,
гаметофиты вообще не имеют сосудистой ткани. Эти гаметофиты
поэтому они очень маленькие и лучше всего развиваются во влажных местах, где они могут
поглощать
воду прямо из окружающей среды.
Как и мохообразные, папоротники и их союзники по-прежнему ограничены
влажный
среда обитания. Их жгутиковым сперматозоидам нужна тонкая пленка воды, чтобы плавать.
между
антеридий и архегоний. А когда вырастет детеныш спорофита
вверх от гаметофита подвергается десикации (высыханию). Этот
общая стратегия свободно плавающего сперматозоида и неподвижной яйцеклетки
растениями, животными и водорослями. Это имеет смысл, потому что это означает только
Это имеет смысл, потому что это означает только
один
набор гамет должен совершить опасное путешествие за пределы
организм.
молотая сосна ( Lycopodium) со стробилами
Папоротники и их союзники прорастают из спор. Эти растения
в основном
односпоровый
— их споры идентичны, и вы не можете отличить, какие из них
превращаются в мужские или женские растения. Они же однодомные —
оба
архегонии и антеридии (мужские и женские репродуктивные структуры)
рождаются на одном и том же растении. Сравните эти примитивные сосудистые растения
с
более развитые семенные растения, голосеменные и покрытосеменные, которые
прорастать
из семян, а не из спор. Семенные растения все разноспоровые .
Более крупную женскую мегаспору легко отличить от
в
меньший самец микроспора . Сперматозоиды семенных растений не имеют
жгутики.
У них отсутствуют антеридии, и лишь у некоторых сохранились архегонии.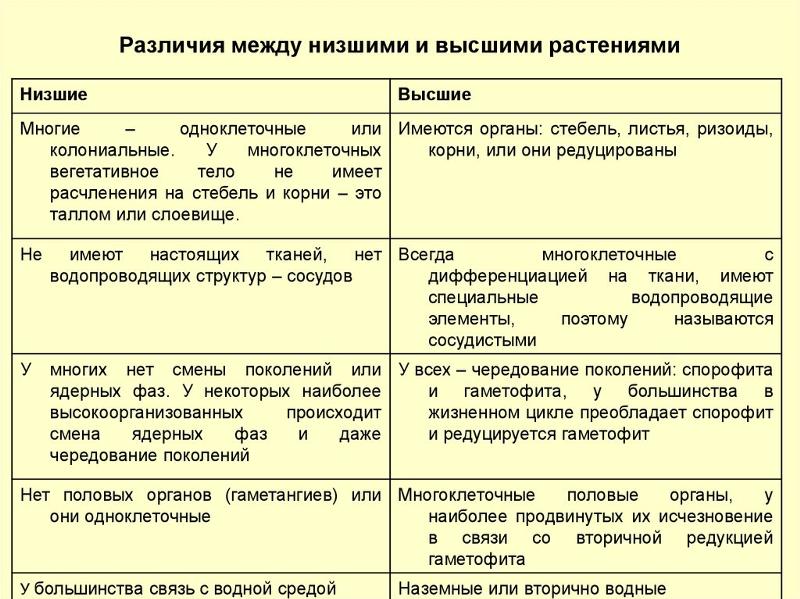 в отличие
в отличие
в
более примитивные папоротники и союзники папоротников, семенные растения в основном двудомные ,
наличие отдельных мужских и женских растений.
У многих из этих примитивных растений некоторые листья являются специализированными.
за
воспроизведение. Эти видоизмененные листья, или спорофиллы , несут
спорангии
на своих базах. Эти спорофиллы обычно отходят от укороченного
корень,
образуя булавовидную структуру, называемую стробилом . Сосна
конус
и цветок — сложные вариации этих примитивных стробилов.
Различают четыре отдела несеменных трахеофитов, сосудистых растений
которые размножаются с помощью спор , Psilophyta, Lycophyta,
Сфенофиты,
и Птерофиты. До появления этих несеменных трахеофитов
мохообразные
были господствующей формой растительной жизни. Эволюционное преимущество
более эффективная проводящая система и хорошо развитая корневая система
система
позволили им превзойти мохообразных. Если вам посчастливится увидеть
земля
сосновые или другие плауны, растущие в тени большой сосны,
считать
на мгновение о том, как эти крошечные растения когда-то были хозяевами
планета,
образуя обширные леса со стволами высотой от 20 до 100 футов!
Таксономия
Царство растений
Трахеофиты (сосудистая ткань, без семян)
Отдел Lycophyta — плаун, игольница ( Lycopodium,
Селагинелла )
Отдел Sphenophyta — хвощи ( Equisetum; fr.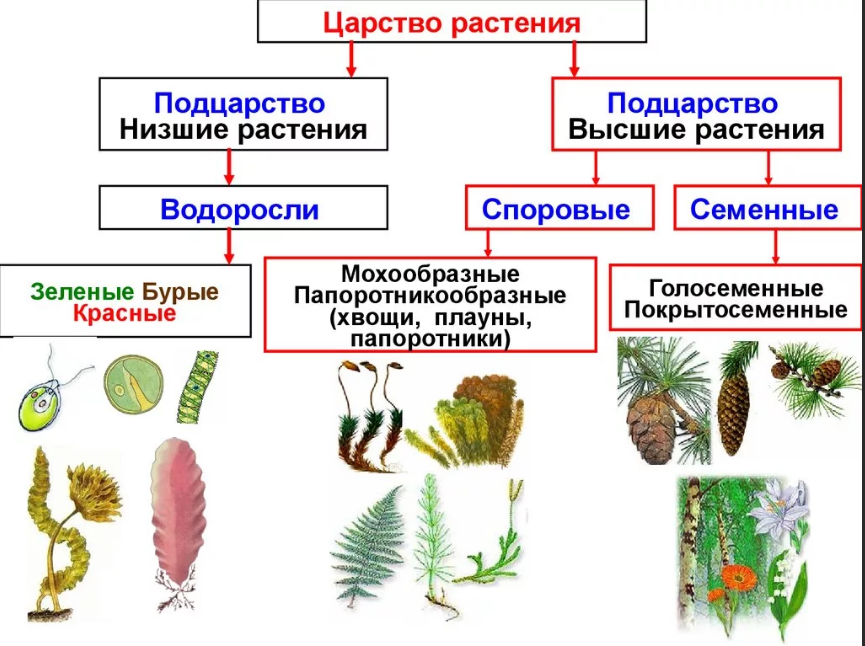 L.
L.
экю
= лошадь)
Отдел Psilophyta — папоротник венчик ( Psilotum )
Отдел Pterophyta — папоротники настоящие ( Pteris; fr.Gr.
птеридион
= маленькое крыло)
Условия
- односпоровый
- разноспоровый
- мегаспоры
- микроспоры
- корневища
- спорофиллы
- стробил (-i)
- сори
- индий (-ия)
- заросток
- фиддлхеды
Характеристики разделов
Отдел Psilophyta — (фр. греч. psilo = гладкий), венчик
папоротники,
Псилотум
Есть только два живых рода папоротников, единственные оставшиеся в живых
крупная и широко распространенная группа ранних наземных растений. В добавок к
жизнь
Отдел Psilophyta, псилопсиды, есть два вымерших отдела
первобытные сосудистые растения. Примитивные папоротники напоминают эти
вымерший
пионерами во многих отношениях.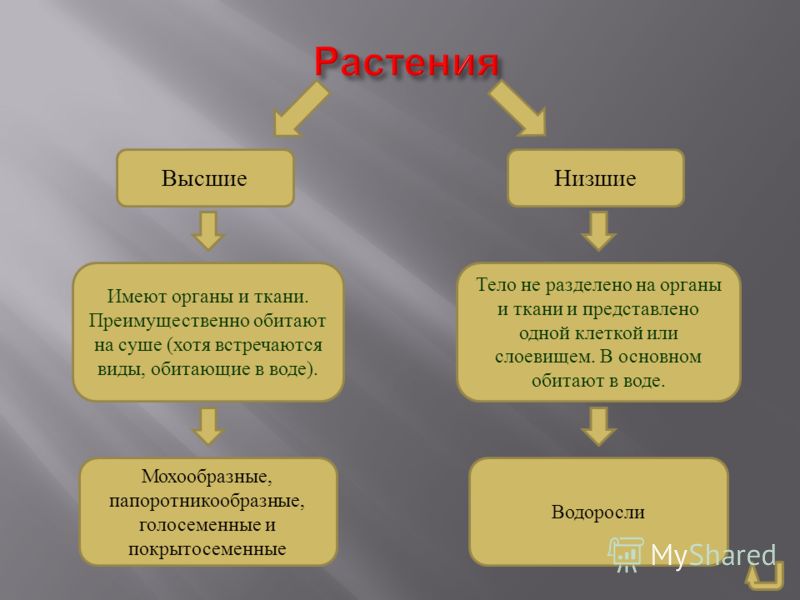 Это единственные живые сосудистые растения,
Это единственные живые сосудистые растения,
недостаток
система корневых побегов, характеристика, которую они разделяют с обоими вымершими
Подразделения
предков сосудистых растений. Некоторые недавние молекулярные данные свидетельствуют о том, что
что
один или даже оба из ныне живущих родов псилопсид на самом деле могут быть
более
тесно связаны с папоротниками, например папоротник, который превратился в более
примитивный
черты. Если это правда, то Psilophyta пополнит ряды
многочисленные
Вымершие отделы растений.
Псилопсиды встречаются в тропических и субтропических районах и встречаются
через
юг США. Однажды я нашел один, растущий на моем заднем крыльце под
листья
паутинного растения. Висячие папоротники — распространенный сорняк в теплицах всех
над
мир. Это простые зеленые прямостоячие стебли с дихотомическими
разветвление.
У них нет ни листьев, ни настоящих корней. Наружные ткани стебля
делать
все фотосинтезирующие. Часть стебля, называемая корневищем, проходит
по земле или чуть ниже нее. А корневище – горизонтальное
А корневище – горизонтальное
корень
который распространяет растение вокруг. Корни вырастают из нижней части
корневище,
и новое растение может возникнуть в той же точке сверху.
Зеленое стеблевидное растение представляет собой диплоидный спорофит, доминирующий
сцена
в жизненном цикле. В мелких спорангиях (ярко-желтого цвета), образующих
вместе
на верхних стеблях материнская клетка спор образует гаплоидные споры путем мейоза.
Их гаметофиты представляют собой крошечные нитевидные подземные растения,
лишены хлорофилла и живут как гетеротрофы в почве, выглядя и
действующий
очень похоже на крошечные грибы. На самом деле он содержит симбиотические грибы,
такой же
микоризы, обитающие в корневище взрослого спорофита.
Отдел Lycophyta — (1000 sp., фр. греч. lycos = волк), клуб
мхи,
quillworts, Lycopodium (подус = ступня)
Их всего пять живых родов плауновидных, но когда-то из
далеком девоне, около 400 млн лет назад, в каменноугольном периоде, они
были господствующей формой растительности на Земле. Теперь они
Теперь они
превращаются в тень своего славного прошлого, неприметные маленькие
растения
в подлеске леса. Тропические виды — мелкие эпифиты.
(растения
которые растут на других растениях).
Их корни растут из особых подземных стеблей, называемых корневищами.
делают большинство этих примитивных трахеофитов. У некоторых видов
спорофиллы
смешаны с чешуйчатыми листьями. У многих видов,
спорофиллы
организованы в стробилы, отсюда и общее название «плауны».
сперматозоиды спускаются по стробилу к архегониям, а зигота
формы
задерживается в шишке, которая созревает и падает на землю.
гаметофиты
независимы и свободно живут. Это любопытные существа, которые выглядят
а также
ведут себя совсем не так, как их родители-спорофиты. Они могут быть либо
гетеротрофный
или автотрофные, и обычно имеют симбиотические грибы, связанные с
их.
Многие ликопсиды гетероспоровые. Селагинелла хороший
пример
гетероспорового растения.
Отдел Sphenophyta — (15 видов, один род, фр. гр.
сфен = клин),
хвощи, Equisitum
На пустырях, нарушенных участках, таких как тропы и железнодорожные полотна, и
в дальних уголках полей и лесов можно найти еще одно маленькое растение
тихо мечтая о своем былом великолепии, хвоще.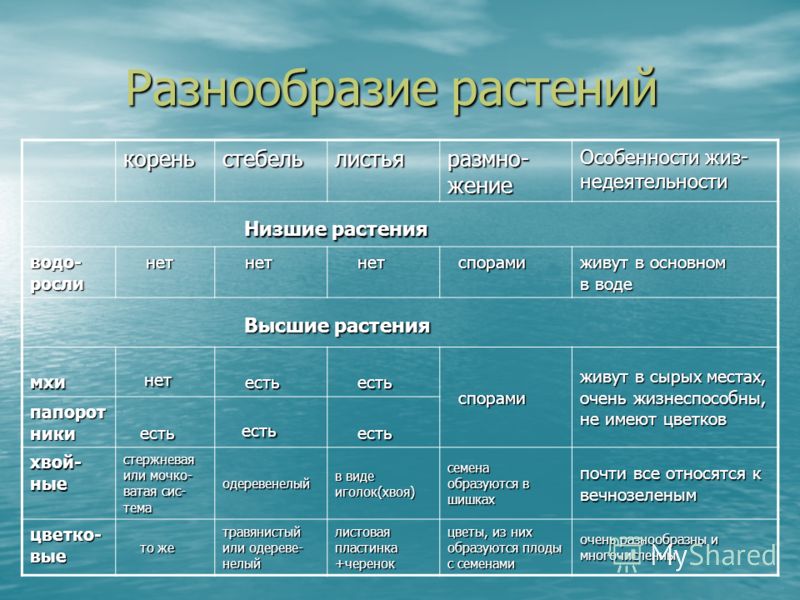 Хвощи
Хвощи
появился
в позднем девоне и были среди доминирующих лесных деревьев для
сотни
миллионов лет. До сих пор существует только один род Sphenophyta,
род
Эквисетум ,
и это может быть самый древний из ныне живущих родов растений на Земле. Хвощи
возвышался
среди каменноугольных лесов, достигая высоты 30-60 футов. Много
из
угольные месторождения, которые мы сегодня используем в качестве топлива, образовались из хвощей
и другие деревья в каменноугольном периоде, ближе к концу
Палеозой.
Хвощи имеют настоящие корни, стебли и листья, хотя листья
чуть больше, чем уплощенные стебли. Их полые ребристые стебли
соединенный,
вроде стебля бамбука, и на каждом из них возникает мутовка листьев.
соединение.
Растения размножаются вегетативно корневищами. Стебли чувствуют себя очень
грубый,
потому что эпидермальные ткани пропитаны мельчайшими зернами
кремнезем
(песок). Вероятно, это помогает защитить растение от травоядных. Эти
грубые стебли сделали это растение идеальным для использования женщинами-первопроходцами для
вытирание
кастрюли и сковородки, отсюда и его другое распространенное название «чистящая каша».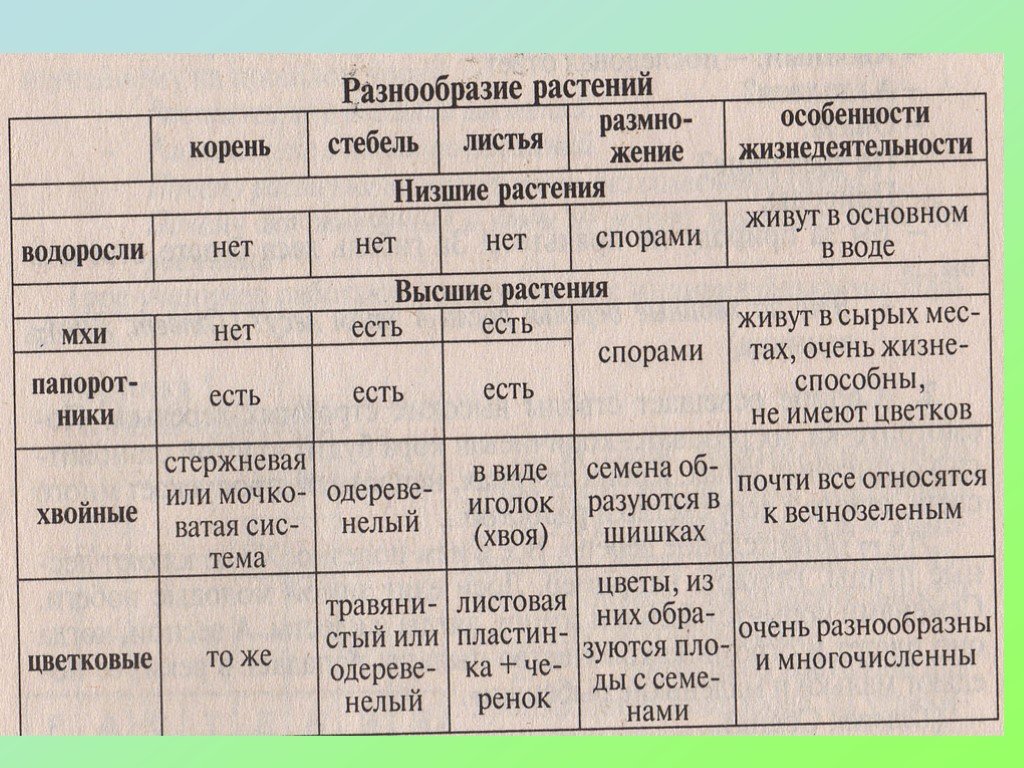
Зеленое растение, которое мы видим, представляет собой поколение диплоидного спорофита.
стебли
могут быть сильно разветвленными вегетативными стеблями, которые на самом деле выглядят как
лошадь
хвосты или прямые неразветвленные репродуктивные стебли с
с
крупный стробил со спорангиями. гомоспоровые споры
развивать
в крошечный зеленый гаметофит длиной всего несколько миллиметров, который выглядит
как
гаметофит папоротника. Гаметофит гаплоидный, свободноживущий.
автотрофный.
Подразделение Pterophyta — (12 000 экз., фр. греч. pteridion = маленький
крыло),
папоротники
Папоротники, вероятно, произошли от псилопсид где-то в
Девон,
относительно рано в эволюции наземных растений. Они очень многочисленны и
разнообразны, размером от одного сантиметра до деревьев 24 метра
высокий
с 5-метровыми ветвями. Папоротники были лучшими конкурентами с семенами
растения
чем другие бессемянные сосудистые растения, и являются заметной частью
ландшафт во всем мире, но особенно в тропиках, где
75%
из их 12 000 видов.
Папоротники — относительно развитые растения с настоящими корнями, стеблями и
листья.
Лист папоротника называется вайей, а маленькая особь
листовки
называются пиннами. У папоротников настоящие листья, что ботаники называют
макрофиллы.
В то время как листья более примитивных растений, которые называются
микрофиллы,
являются просто продолжением эпидермиса стебля, листьев папоротников
и высшие растения формировались в виде паутины ткани, натянутой между
маленький
конечные ветки. Листья высших растений, а также видоизмененные
листья, из которых состоят сосновая шишка и цветок.
Жизненный цикл папоротника типичен для других несеменных сосудистых
растения.
Листовое зеленое растение – спорофит. Развиваются плодородные листья
кластеры
из мелких спорангиев на нижней стороне вайи. Эти кластеры
спорангии
называются сори (песн. сорус). Сори часто защищены
крошечный
зонтиковидная кепка, называемая индузиум (-ia). Папоротники в основном
односпоровый,
хотя некоторые из них разноспоровые. Гетероспоровое состояние более
Гетероспоровое состояние более
передовой
состояние, которое, по-видимому, развилось независимо в нескольких группах
из
растения.
Гаплоидные споры образуются путем мейоза внутри спорангиев. Они
выбрасываются миниатюрным взрывом, вызванным неравномерным высыханием
в
чередующиеся толстостенные и тонкостенные клетки, выстилающие наружную поверхность.
верхняя часть медленно тянется назад, пока не достигнет критической точки, а затем защелкивается
вперед с невероятной скоростью. При таком масштабе изгнание
папоротник
споры — одно из самых взрывоопасных явлений в природе. Споры
прорастать
в крошечные гаметофиты. Гаметофит в форме маленького сердца называется
заросток ,
буквально «первое тело» (пл. prothalli). проталлус не имеет сосудов
салфетка.
Его небольшой размер позволяет полностью полагаться на диффузию. Его крошечные ризоиды
находятся
связаны с микоризными грибами. Маленький заросток зеленый, а
фотосинтезирует и несет либо антеридии и архегонии, либо
иногда
оба вместе, на его верхней поверхности (лабораторные предметные стекла имеют оба на одном и том же
проталлус).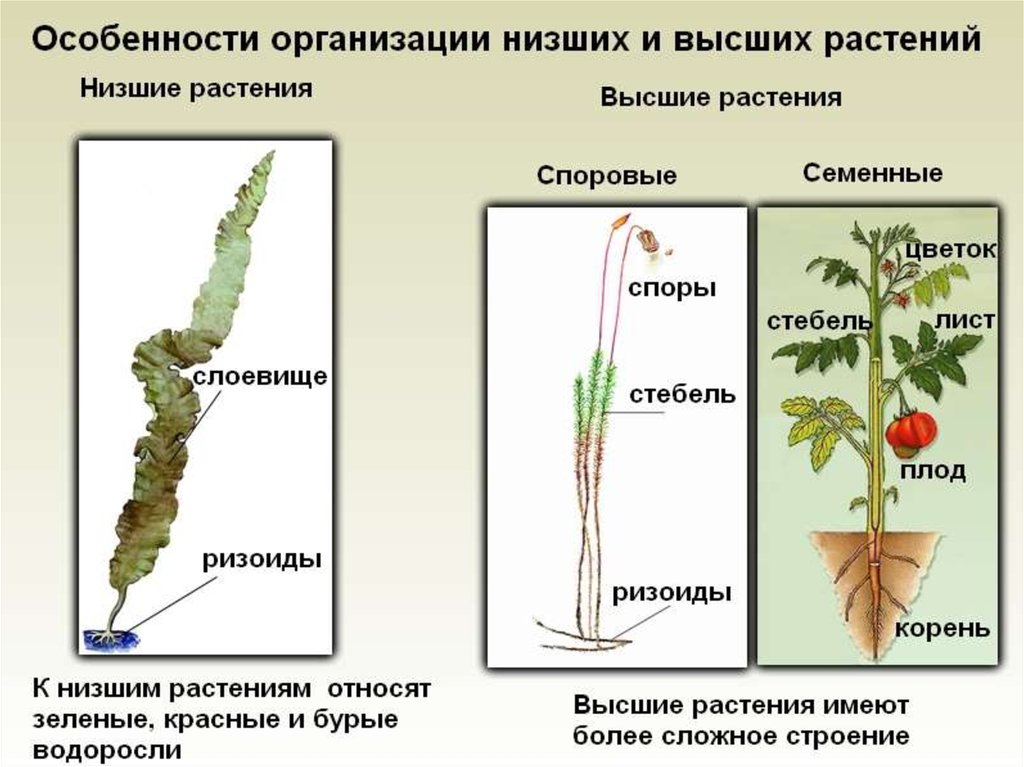
Архегонии всегда находятся на дуге сердца, а
антеридии
спрятаны среди крошечных ризоидов на другом конце. Сперма
плавает
к яйцеклетке, чтобы слиться в диплоидную зиготу. Новый спорофит вырастает
напрямую
из верхушки гаметофита. Когда он впервые начинает разворачиваться,
ветвь похожа на закрученный гриф скрипки или скрипки, и этот
сцена
развития называется фиддлхедом.
Делать и просматривать
Осмотрите живых ликопсидов на витрине. Почему их называют
клубные мхи? Обратите внимание, что иголки и Selaginella очень
другой
по внешнему виду из плауновых мхов.
Изучите препараты стробила селагинеллы . Определите мегаспору
и микроспор .
Осмотрите живые хвощи на витрине. Обратите внимание на
видный
стробили
из
репродуктивные стебли и кустистая форма роста вегетативных
стебли
(если доступно).
Осмотрите живые папоротники на витрине. псилопсиды
псилопсиды
имеют простой дихотомический тип ветвления. Вы можете увидеть крошечный желтый
спорангии
на ветвях. Метелки папоротника отсутствуют стробилы. Эти примитивные растения
тесно связаны с папоротниками.
Осмотрите живые папоротники на витрине. Вы видите какие-нибудь фиддлхеды ?
Ищите корневища . Корневища представляют собой видоизмененные горизонтальные стебли.
несущий
корни, идущие вдоль или прямо под землей, и распространяющие папоротники
а также
союзники папоротника вокруг.
Осмотрите живой заросток папоротника , выставленный под
рассечение
микроскоп. Обратите внимание на его характерную форму сердца. Некоторые заростки могут
имеют
крошечный новый папоротник, вырастающий из выемки сердцевины «арки», где
архегония
соток
расположен.
Изучите препараты проталлуса: Найдите архегония
(ты видишь яйцо?).
Осмотрите листовку с папоротником , выставленную под секционным
микроскоп.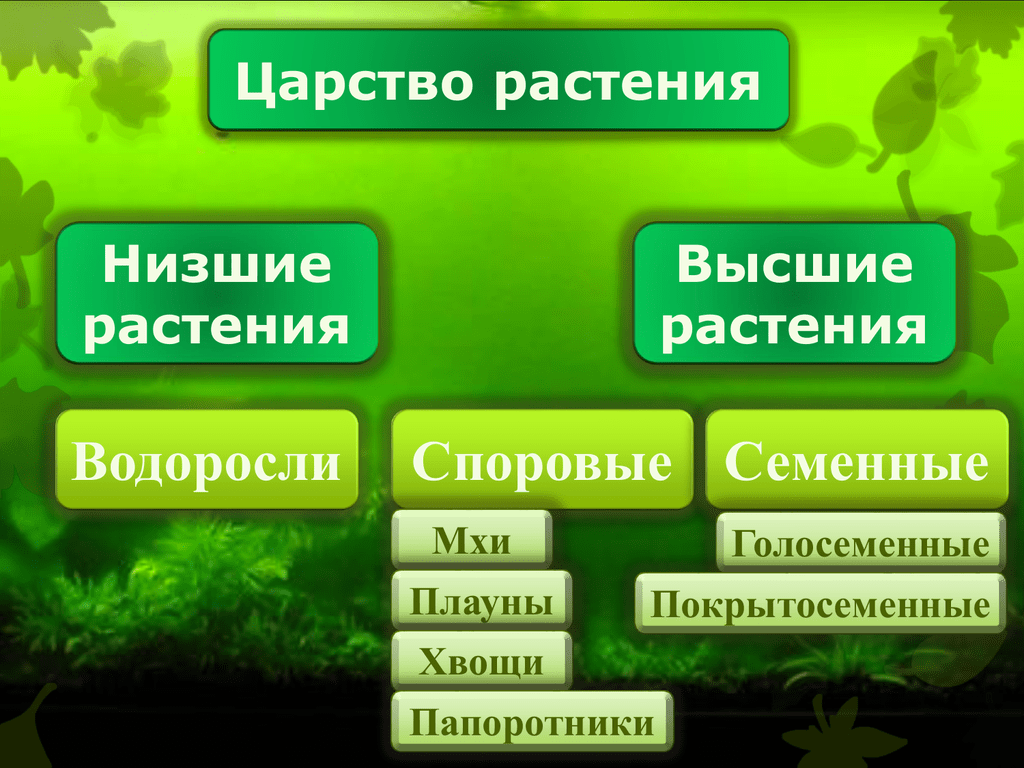
Обратите внимание на видную индусию и маленькую спорангию .
подглядывание
наружу из-под. Вы увидите несколько групп спорангиев, которые потеряли
их
индий. Если повезет, тепло микроскопа высушит
листовке, можно увидеть, как спорангии выбрасывают свои споры. Но нет
моргать
— это одно из самых быстрых и взрывоопасных явлений в природе. Используйте высокие
сила
наблюдать за споры .
Изучите слайды сори и индузии; использовать большую мощность
наблюдать за спорами .
Что нужно помнить
Знать жизненный цикл папоротника. Распознавать образцы и
слайды
различных этапов.
Экономическое, экологическое и эволюционное значение
Папоротники и их союзники в первую очередь ответственны за наше современное
депозиты
угля.
Головки некоторых видов папоротников съедобны.
Папоротники важны для цветоводов, садоводов и ландшафтных дизайнеров.
отрасли.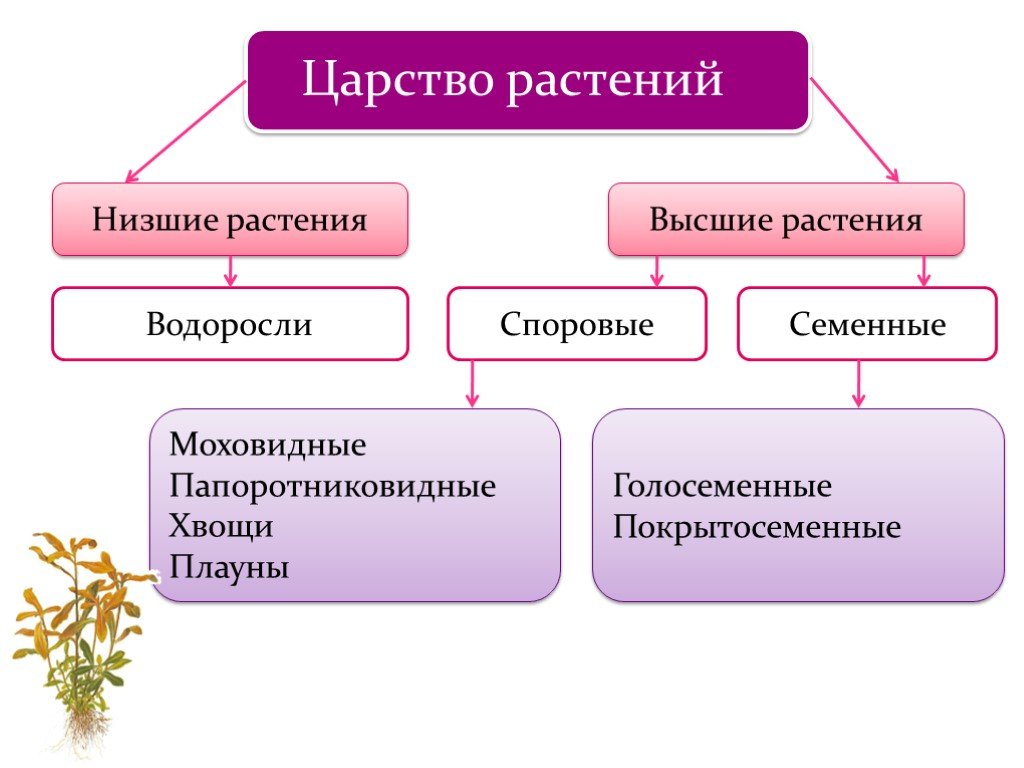
Подумайте об этом
Почему все эти растения ограничены влажными местообитаниями?
Какая группа простейших дала начало этим растениям? (Откуда нам знать?)
Почему эпидермис хвоща такой шероховатый? Что ему нужно
защита
из?
Все союзные папоротники в этой лаборатории когда-то возвышались на 50-100 футов и более.
Что случилось?
Ссылки для изучения
Таксономия мохообразных, изображения и многое другое по адресу:
http://bryophytes.plant.siu.edu/
Получите информацию о мохообразных в Ботаническом саду Миссури по адресу:
http://www.mobot.org/mobot/tropicos/most/
Полежите среди папоротников в Американском обществе папоротников, попробуйте их
длинный
список ссылок:
http://amerfernsoc.org
Основная информация о папоротниках и их союзниках, с изображениями, любезно предоставлено
Техас A&M:
http://www.csdl.tamu.edu/FLORA/fsb/fsbfern1.