
Содержание
Этапы эволюции растений
Муниципальное образовательное учреждение
«Средняя общеобразовательная школа № 2»
Реферат по биологии
Этапы эволюции растений
Исполнитель:
Руководитель:
Алапаевск
2010
Введение
Предпосылок для появления наземных растений было, по-видимому, несколько:
Во-первых, независимый ход эволюции растительного мира подготовил появление новых, более совершенных форм.
Во-вторых, за счет фотосинтеза морских водорослей в атмосфере земли произошло увеличение количества кислорода; к началу силурийского периода оно достигло такой концентрации, при которой оказалась возможной жизнь на суше.
В-третьих, в начале палеозойской эры на обширных территориях земли происходили крупнейшие горообразовательные процессы, в результате которых возникли Скандинавские горы, горы Тянь-Шань, Саяны. Это вызвало обмеление многих морей и постепенное появление суши на месте бывших мелких водоемов. Если раньше водоросли , населявшие литоральную зону, только в отдельные кратковременные периоды жизни оказывались вне воды, то по мере обмеления морей они переходили к более длительному пребыванию на суше. Это, очевидно, сопровождалось массовой гибелью водорослей; выживали лишь те немногие растения, которые смогли противостоять новым условиям жизни. Я хочу рассказать о главных этапах эволюции растений.
ПОНЯТИЕ О БИОСФЕРЕ
Понятие биосфера вошло в науку до некоторой степени случайно. Около ста лет назад, в 1875 году, австрийский геолог Эдуард Зюсс, говоря о различных оболочках земного шара, впервые употребил этот термин в последней главе своей небольшой книжке о происхождении Альп. Однако эта концепция не сыграла заметной роли в развитии научной мысли до тех пор, пока в 1926 году не были опубликованы две лекции русского минералога Владимира Ивановича Вернадского. Концепция биосферы, которую мы принимаем сейчас, в основном опирается на идеи Вернадского, развитые им спустя 50 лет после работ Зюсса.
Около ста лет назад, в 1875 году, австрийский геолог Эдуард Зюсс, говоря о различных оболочках земного шара, впервые употребил этот термин в последней главе своей небольшой книжке о происхождении Альп. Однако эта концепция не сыграла заметной роли в развитии научной мысли до тех пор, пока в 1926 году не были опубликованы две лекции русского минералога Владимира Ивановича Вернадского. Концепция биосферы, которую мы принимаем сейчас, в основном опирается на идеи Вернадского, развитые им спустя 50 лет после работ Зюсса.
Сам Вернадский считал, что впервые к понятию биосферы подошел французский натуралист Жан Батист Ламарк, в чьих работах можно немало геохимических идей, пусть и архаично изложенных.
Биосферой называется та часть земного шара, в пределах которой
существует жизнь. Однако такое определение порождает ряд вопросов и требует уточнений. Пропуская через фильтр воздух, взятый на больших высотах, можно найти в нем споры бактерий и грибов. Но этот «аэропланктон», очевидно, не имеет активного метаболизма. Даже на поверхности Земли немало мест, слишком холодных, слишком жарких или слишком сухих, для того чтобы там могли существовать организмы с активным метаболизмом. Но и в таких местах всегда можно найти споры. Таким образом, оболочка Земли, называемая биосферой, имеет неправильную форму, т. к. она окружена некоей «парабиосферной» областью, в которой жизнь присутствует только в покоящемся состоянии. В настоящее время живой организм может, конечно, существовать далеко за пределами естественной биосферы, находясь в космическом корабле или скафандре. Такие искусственные местообитания можно рассматривать как участки биосферы, вырванные из нее и временно заброшенные в космос.
Но этот «аэропланктон», очевидно, не имеет активного метаболизма. Даже на поверхности Земли немало мест, слишком холодных, слишком жарких или слишком сухих, для того чтобы там могли существовать организмы с активным метаболизмом. Но и в таких местах всегда можно найти споры. Таким образом, оболочка Земли, называемая биосферой, имеет неправильную форму, т. к. она окружена некоей «парабиосферной» областью, в которой жизнь присутствует только в покоящемся состоянии. В настоящее время живой организм может, конечно, существовать далеко за пределами естественной биосферы, находясь в космическом корабле или скафандре. Такие искусственные местообитания можно рассматривать как участки биосферы, вырванные из нее и временно заброшенные в космос.
Что же характерно для биосферы как особой оболочки земного шара? Во-первых, это область, в которой имеется в значительных количествах жидкая вода.
Во-вторых, на нее падает мощный поток энергии от Солнца.
Наконец, в-третьих, в биосфере имеются поверхности раздела между веществами, находящимися в жидком, твердом и газообразном состояниях
Эволюция растительного мира.
Основные этапы эволюции растений.
В протерозойскую эру (около 1 млрд. лет назад) ствол древнейших эукариот разделился на несколько ветвей, от которых возникли растения, грибы и животные. Большинство растений этого периода свободно плавало в воде, часть из них прикреплялась ко дну.
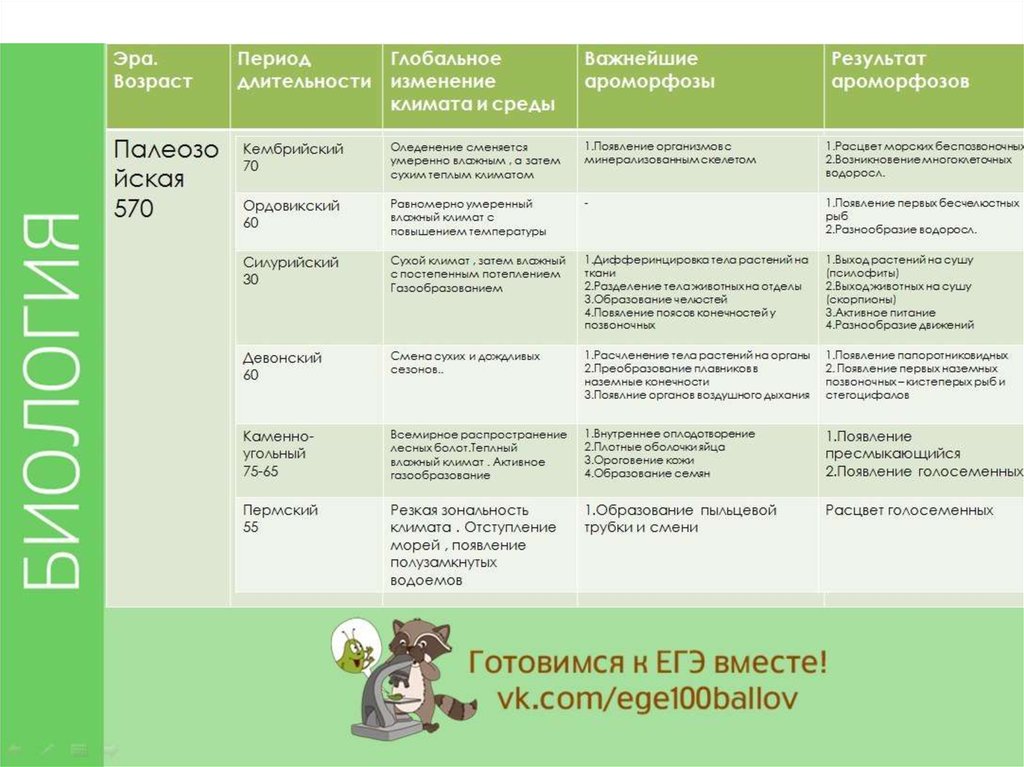
Табл. 4.2. Геохронологическая шкала Земли.
Эра | Период | Начало (млн. лет назад) | Эволюционные события |
Кайнозойская (новой жизни) | Четвертичный | 2,4 | Вымирание многих видов растений, упадок древесных форм, расцвет травянистых; растительный мир приобретает современный облик. |
Неогеновый | 2,5 | Преобладание покрытосеменных и хвойных, отступание лесов, увеличение площади степей. Видовой состав беспозвоночных приближается к современному. Расцвет плацентарных млекопитающих, сходных с современными. Появление человекообразных обезьян. | |
Палеогеновый | 66 | Расцвет диатомовых водорослей и основных групп покрытосеменных. Господство двустворчатых и брюхоногих моллюсков. Вымирание древнейших млекопитающих. Развитие сумчатых и примитивных плацентарных: насекомоядных, древних копытных, древних хищников. Начало развития антропоидов. | |
Мезозойская (средней жизни) | Меловой | 136 | В начале периода господство голосеменных и появление покрытосеменных, которые преобладают во второй половине периода. |
Юрский | 195 | Появление диатомовых водорослей. Господство папоротников и голосеменных. Расцвет головоногих и двустворчатых моллюсков. Расцвет пресмыкающихся: наземных, водоплавающих, летающих. Появление древних птиц, развитие древних млекопитающих. | |
Триасовый | 240 | Вымирание семенных папоротников. Развитие голосеменных. Вымирание многих животных, процветающих в палеозойскую эру. Вымирание стегоцефалов, развитие пресмыкающихся, появление древних млекопитающих. | |
Палеозойская (древней жизни) | Пермский
| 285 | Распространение первых групп голосеменных. Уменьшение количества видов хрящевых, кистеперых и двоякодышащих рыб. |
Каменноугольный | 345 | Расцвет плауновидных, хвощевидных, папоротниковидных, семенных папоротников; появление хвойных. Расцвет древних морских беспозвоночных. Появление первичнобескрылых и древнекрылых насекомых. Распространение акул, стегоцефалов. Появление и расцвет амфибий. Появление древних пресмыкающихся. | |
Девонский | 410 | Расцвет риниофитов, к началу позднего девона их вымирание. Появление современных типов сосудистых растении. Расцвет древних беспозвоночных, появление паукообразных. Расцвет панцирных, кистеперых и двоякодышащих рыб. В конце периода появление первых четвероногих — стегоцефалов (древних земноводных). | |
Силурийский | 435 | Возникновение современных групп водорослей и грибов. В конце периода достоверное появление первых наземных растений. | |
Ордовикский | 500 | Обилие морских водорослей. Предположительное появление первых наземных растений — риниофитов. Появление первых позвоночных— бесчелюстных. | |
Кембрийский | 570 | Жизнь сосредоточена в морях. Эволюция водорослей; развитие многоклеточных форм. Расцвет морских беспозвоночных с хитиново-фосфатной раковиной. | |
Протерозойская (ранней жизни) | Поздний протерозой | 1650 | Развитие водорослей, различных многоклеточных примитивных организмов, не имеющих скелетных образований. |
Ранний протерозой | 2600 | Развитие одноклеточных прокариоти-ческих и эукариотических фотосинте-зирующих организмов. Возникновение полового процесса. | |
Архей |
| 3500— 3800 | Возникновение жизни на Земле, появление первых клеток — начало биологической эволюции. |
Катархей |
| 3900 | Химическая эволюция, приведшая к возникновению биополимеров. |
 Развитие многих групп морских н пресноводных моллюсков, кораллов, иглокожих и др. Формирование ныне существующих сообществ, возникновение и эволюция человека.
Развитие многих групп морских н пресноводных моллюсков, кораллов, иглокожих и др. Формирование ныне существующих сообществ, возникновение и эволюция человека.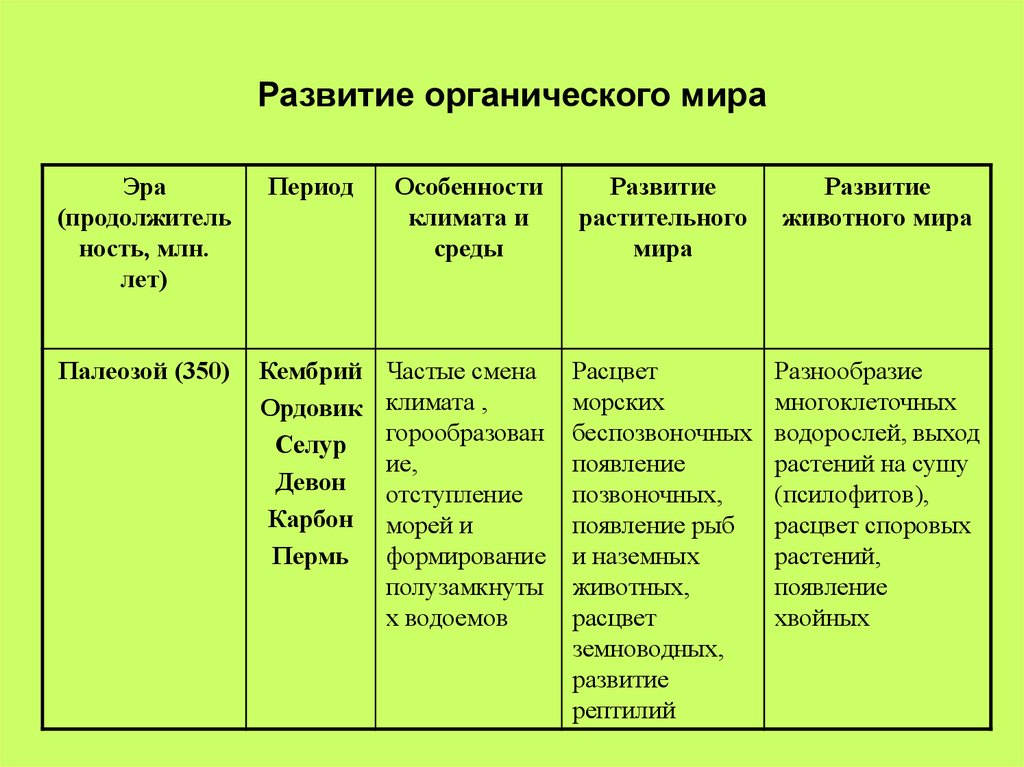 Развитие двустворчатых и брюхоногих моллюсков, других беспозвоночных. Развитие крупных рептилий в первой половине периода и их вымирание во второй половине периода. Развитие млекопитающих и птиц.
Развитие двустворчатых и брюхоногих моллюсков, других беспозвоночных. Развитие крупных рептилий в первой половине периода и их вымирание во второй половине периода. Развитие млекопитающих и птиц. Развитие стегоцефалов, пресмыкающихся, часть которых были предковыми по отношению к млекопитающим и птицам.
Развитие стегоцефалов, пресмыкающихся, часть которых были предковыми по отношению к млекопитающим и птицам. Появление наземных членистоногих —- скорпионов. Появление древних панцирных и хрящевых рыб.
Появление наземных членистоногих —- скорпионов. Появление древних панцирных и хрящевых рыб. Появление анаэробных автотрофных организмов, бактерий, цианобактерий.
Появление анаэробных автотрофных организмов, бактерий, цианобактерий.
Особенности эволюции растительного мира
1) Постепенный переход к преобладанию диплоидного поколения над
гаплоидным. У многих водорослей все клетки (кроме зиготы) гаплоидны, у
голосеменных и покрытосеменных почти полностью редуцируется гаметофит и
значительно удлиняется в жизненном цикле диплоидная фаза.
2) Независимое половое размножение от капельноводной среды. Мощное
развитие спорофита, переход от наружного оплодотворения к внутреннему,
возникновение двойного оплодотворения и обеспечение зародыша запасами
питательных веществ.
3) В связи с прикрепленным образом жизни на суше растение
расчленяется на корень, стебель и лист, развиваются сосудистая проводящая
система, опорные и защитные ткани.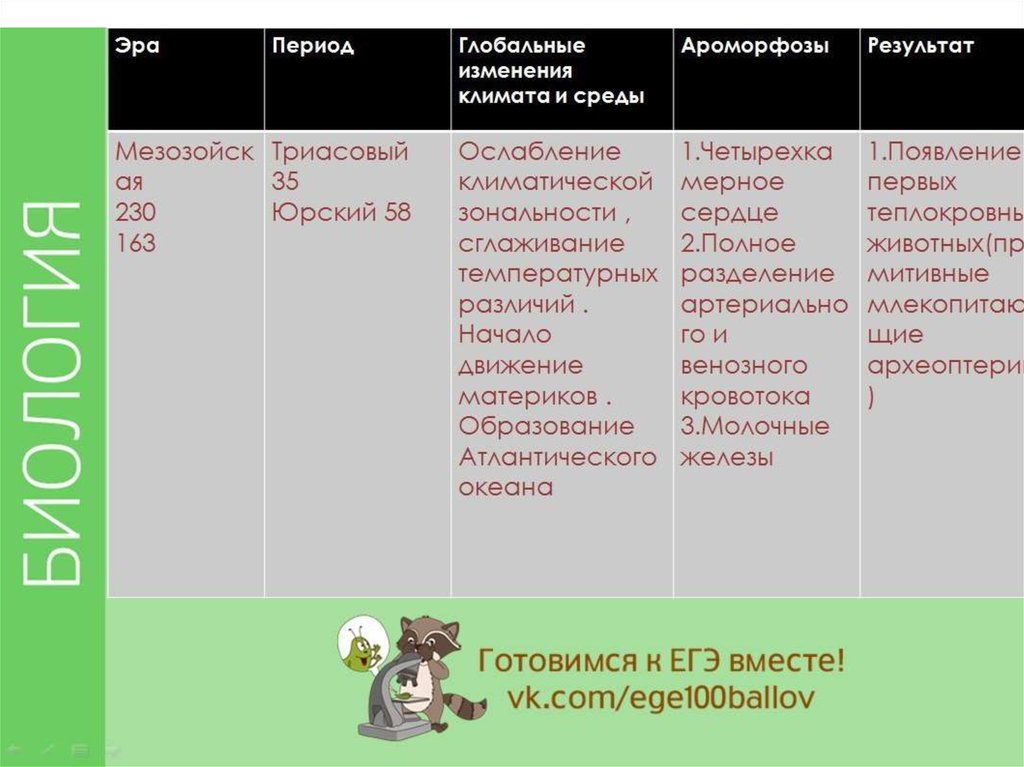
4) Совершенствование органов размножения и перекрестного опыления у
цветковых в сопряженной эволюции с насекомыми. Развитие зародышевого мешка для защиты растительного эмбриона от неблагоприятных влияний внешней среды. Возникновение разнообразных способов распространения семян и плодов физическими и биотическими факторами.
ОСОБЕННОСТИ ЭВОЛЮЦИИ ОРГАНИЧЕСКОГО МИРА , ВАЖНЫЕ ДЛЯ РАЗВИТИЯ БИОСФЕРЫ.
В каменноугольной флоре широко были распространены споровые
влаголюбивые растения (плуан, хвощи и папоротники). Затем началось
появление голосеменных растений, в том числе и хвойных.Эти растения
обладали значительными преимуществами по сравнению со своими предшественниками и поэтому не только шире их расселились по поверхности суши , но и в значительной мере вытеснили их из зон , которые те вначале безраздельно занимали. В связи с этим в мезозойскую эру господствующее положение на суши приобрели голосеменные растения.
Затем на смену им пришло господство покрытосеменных растений , еще
более высокоорганизованных. Первые покрытосеменные появились в середине мезозойской эры. В кайнозое они быстро завладели всей поверхностью Земли.
С точки зрения развития биосферы, большой интерес представляет
возможная направленность эволюции покрытосеменных. Травянистый тип
покрытосеменных растений произошел от древесного. Процесс развития
трав из древесных растений представляется следующим:
деревья -кустарники -полукустарники — многолетние травы — однолетние травы.
Травы являются наиболее приспособленными представителями растительности. Темпы эволюции травы был очень высок , и очень быстро после своего возникновения покрытосеменные растения расселились на громадных территориях.
Благодаря развитию биосферы Земля постепенно приобрела всевозможные цвета и оттенки и стала планетой-оазисом среди других планет солнечной системы. Живое вещество не только является главным и необходимым компонентом биосферы, определяющим ее развитие, но и наиболее изменчивой и наиболее быстро эволюционирующей составной частью земной коры.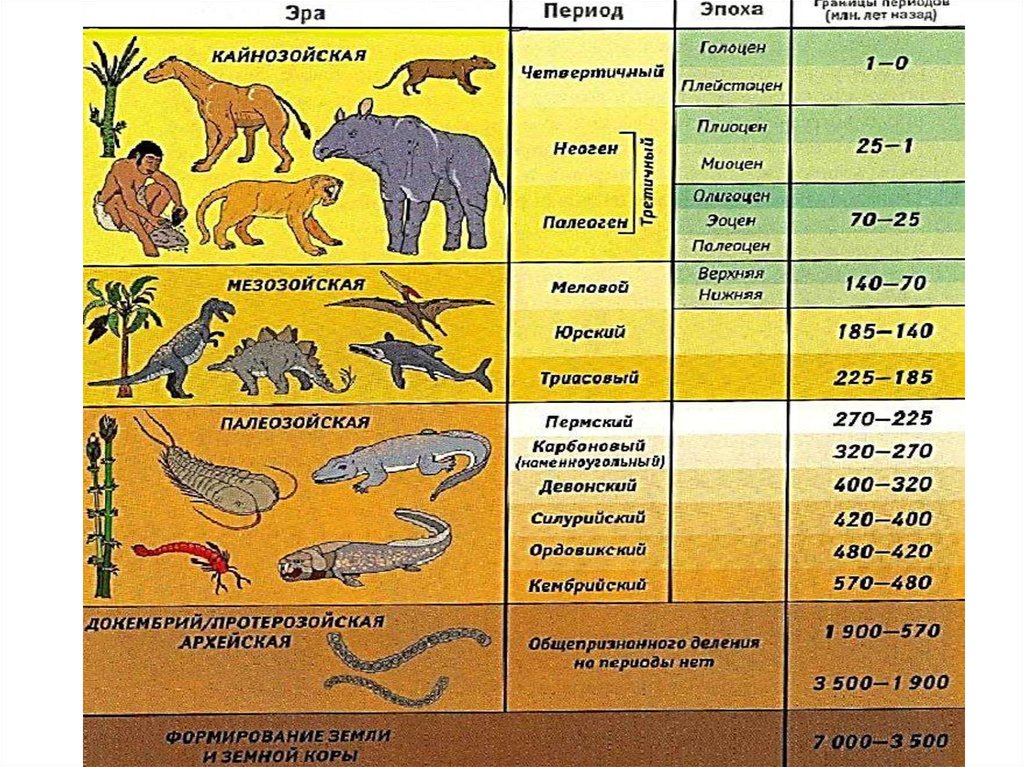
Разнообразные процессы, связанные с существованием и развитием
живого вещества, не только представляют собой главные факторы эволюции
всей биосферы и осадочного породообразования на протяжении длительной
геологической истории Земли, но и, очевидно, являются основной
причиной образования самой земной коры.
Заключение
Появление наземных, или высших, растений ознаменовало начало новой эры в жизни нашей планеты. Освоение растениями суши сопровождалось появлением новых, наземных, форм животных; сопряженная эволюция растений и животных привела к колоссальному разнообразию жизни на земле, изменила ее облик.
Эволюция растений (растительного мира) | Ботаника. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
Изучение ископаемых остатков, которыми занимается палеоботаника, позволяет установить этапы развития растительного мира на Земле. Возраст Земли, по современным представлениям — 4,5-4,6 млрд. лет. Геологическую историю Земли делят на эры: архейскую, протерозойскую, палеозойскую, мезозойскую, кайнозойскую и периоды.
Возраст Земли, по современным представлениям — 4,5-4,6 млрд. лет. Геологическую историю Земли делят на эры: архейскую, протерозойскую, палеозойскую, мезозойскую, кайнозойскую и периоды.
Растительность на нашей планете непрерывно изменялась. От многих, когда-то широко распространённых групп остались отдельные представители. Например, от хвощевидных — только один род хвощ, от гинкговых — один вид, секвойи сохранились только в Калифорнии, а древовидные папоротники — только в Австралии и Новой Гвинее. Сравнительно редкими стали саговники, магнолии и многие другие виды. Появились новые виды, которые лучше приспособлены к современным климатическим условиям.
Архейская эра (древнейшая)
Архейская эра, или древнейшая, характеризуется зарождением жизни. Первыми фотосинтезирующими организмами были бактерии и сине-зелёные водоросли (цианобактерии). Существование фототрофных организмов в этот период (более трёх млрд. лет назад) подтверждается обнаружением в наиболее древних отложениях земной коры органических соединений в графитовых включениях, окисленного железа и известковых отложений (строматолитов), образованных, по мнению учёных, сине-зелёными водорослями и их предками.
лет назад) подтверждается обнаружением в наиболее древних отложениях земной коры органических соединений в графитовых включениях, окисленного железа и известковых отложений (строматолитов), образованных, по мнению учёных, сине-зелёными водорослями и их предками.
Протерозойская эра
В Протерозойской эре (2,6-0,6 млрд. лет назад) благодаря жизнедеятельности фототрофных прокариот начинает формироваться окислительная атмосфера. Параллельно с развитием прокариот появляются первые эукариотные организмы. В отложениях возрастом около 2 млрд. лет встречаются колонии одноклеточных и нитчатых форм зелёных и золотистых водорослей. В конце протерозойской эры появились многоклеточные эукариоты, в том числе и водоросли.
Палеозойская эра
Палеозойская эра продолжалась примерно 340-350 млн. лет. Её делят на 6 периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный и пермский.
Кембрийский период
В кембрийский период (80-90 млн. лет) оледенение в начале периода сменяется умеренно—влажным, а затем сухим климатом. Наступление моря на сушу в конце периода сменяется его отступлением. Жизнь развивается в водной среде. Фотосинтезирующие организмы представлены сине-зелёными, красными, зелёными и другими группами водорослей.
Ордовикский период
В ордовикском периоде (50-60 млн. лет) продолжается процесс эволюции водорослей. Они представлены разнообразными формами и группами. Благодаря их жизнедеятельности в атмосфере повышается содержание кислорода, создаются условия для образования озонового экрана. В конце ордовикского периода в связи с интенсивным горообразованием значительная часть территории освобождается от воды.
Силурийский период
В силурийский период (35 млн. лет) появились первые наземные высшие растения, которыми были, как предполагается, псилофиты, жившие 420 — 350 млн.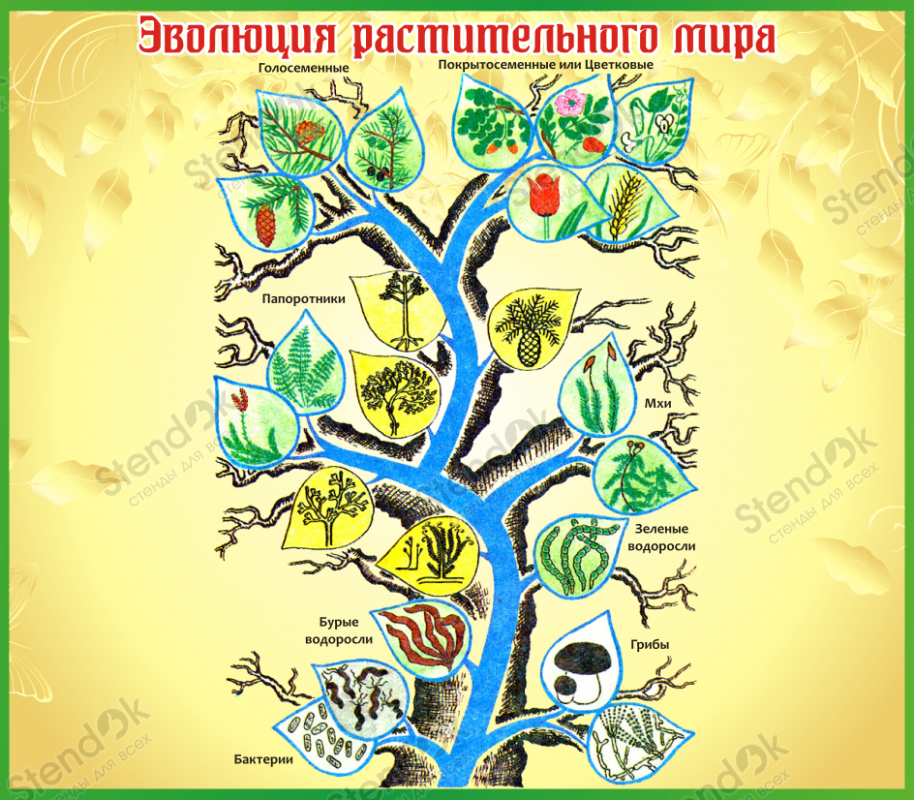 лет назад. Предками высших растений были зелёные водоросли. Выход на сушу потребовал приспособления к совершенно новым условиям и дал толчок к перестройке всей организации растения. Произошло разделение тела на органы, возникли ткани (проводящие, механические, покровные и др.).
лет назад. Предками высших растений были зелёные водоросли. Выход на сушу потребовал приспособления к совершенно новым условиям и дал толчок к перестройке всей организации растения. Произошло разделение тела на органы, возникли ткани (проводящие, механические, покровные и др.).
Девонский период
Девонский период (55 млн. лет) характеризуется оледенением современных Южной Америки и Южной Африки и освобождением от моря Сибири и Европейской части России. Происходит массовое расселение псилофитов, которые в конце этого периода вымирают. В девоне появляются древние плауны, хвощи и папоротники, представленные древовидными формами. В конце этого периода появились первые древние голосеменные растения — семенные папоротники.
Загрузка…
Каменноугольный период
Каменноугольный период (65 млн. лет) был эпохой господства в растительном мире древовидных форм споровых растений, которые образовывали леса. Тёплый влажный климат обусловил широкое распространение болот и мелких морей, в которых накапливались растительные остатки, погребались в наносах и постепенно обугливались, превращались в залежи каменного угля.
Тёплый влажный климат обусловил широкое распространение болот и мелких морей, в которых накапливались растительные остатки, погребались в наносах и постепенно обугливались, превращались в залежи каменного угля.
Наряду со споровыми растениями широко распространились семенные папоротники и появились кордаиты, гинкговые и первые хвойные деревья.
Пермский период
Пермский период (60 млн. лет) характеризуется резкой зональностью климата. Южное полушарие подвергалось оледенению. Происходило отступление морей и формирование полузамкнутых водоёмов. Исчезают леса карбона из-за вымирания древовидных хвощей, плаунов и папоротников. Хвойные распространяются в Северном полушарии.
Мезозойская эра
Мезозойская эра началась примерно 230 млн. лет назад и продолжалась 165 млн. лет. Её делят на три периода: триасовый, юрский и меловой.
Триасовый период
Триасовый период (30-40 млн. лет) отличался новыми условиями существования растений: изменилась конфигурация материков и морей. Климат стал более засушливым и солнечным. Во флоре господствуют голосеменные: саговники и хвойные, которые расселились по всему свету. Семенные папоротники в этот период вымирают.
лет) отличался новыми условиями существования растений: изменилась конфигурация материков и морей. Климат стал более засушливым и солнечным. Во флоре господствуют голосеменные: саговники и хвойные, которые расселились по всему свету. Семенные папоротники в этот период вымирают.
Юрский период
Юрский период (60 млн. лет) ознаменовался движением континентов и формированием Атлантического океана. Во флоре наблюдается дальнейший расцвет голосеменных: хвойных, саговников, гинкговых. Появились беннеттиты.
Меловой период
Меловой период (70 млн. лет) характеризуется обширных увеличением площади Мирового океана и новым поднятием суши. От Азии отделилась Австралия, образовался Берингов пролив. Во флоре значительную роль играют хвойные растения, образуя обширные леса. Беннеттиты к середине мелового периода вымирают. В конце периода появились покрытосеменные растения, которые быстро расселились по всей Земле.
Кайнозойская эра
Кайнозойская эра началась 67 млн. лет назад и продолжается в настоящее время. Её делят на три периода: палеогеновый, неогеновый и антропогеновый. Материал с сайта http://doklad-referat.ru
Палеогеновый период
Палеогеновый период (41-42 млн. лет) ознаменовался интенсивным горообразованием. Возникли горы: Кавказские, Памир, Гималаи, Анды и др. Происходило движение континентов и обособление морей Каспийского, Чёрного, Средиземного и Аральского. Во флоре господствовали покрытосеменные растения, среди которых преобладали деревья.
Неогеновый период
В неогеновый период (23 млн. лет) устанавливается тёплый климат, и покрытосеменные растения достигли наиболее пышного развития. Даже в Северном полушарии были широко распространены магнолии, платаны, бук, дикий виноград, секвойи, вечнозелёные дубы и др. В конце периода наступило похолодание. Возникли растительные зоны: тайга, тундра, лесостепь и степи.
В конце периода наступило похолодание. Возникли растительные зоны: тайга, тундра, лесостепь и степи.
Антропогеновый период
Антропогеновый период (1,5 — 2 млн. лет) характеризуется появлением человека. Климат неоднократно менялся с потепления на похолодание. Северное полушарие неоднократно подвергалось оледенению. Растительность резко изменила свой облик. Возникли холодоустойчивые виды, среди которых главное место заняли хвойные и лиственные деревья, кустарники и травы. В южной части Северного полушария леса сменились степью и пустыней. Теплолюбивые растения сохранились в Закавказье, Юго-Восточной Азии и на Западе Северной Америки. В послеледниковое время чётко обозначилась зональное распределение растительности. Растительный мир принимает современный облик.
На этой странице материал по темам:
Доклад каменноугольный период растения
Эволюция растительного мира реферат
Эволюция растений лекции
Вопросы по этому материалу:
Назовите эры и периоды, выделяемые в геологической истории земли.

Когда появились первые фотосинтезирующие организмы, и какими группами они были представлены?
В какой период истории Земли растительный мир был представлен в основном водорослями?
Когда появились первые высшие растения, и какой группой они были представлены?
В какой период появились древние плауновидные, хвощевидные, папоротниковидные, и какими жизненными формами они были представлены?
Когда наблюдалось господство высших споровых растений?
В какой период появились первые семенные растения, и какой группой они были представлены?
Когда наблюдался расцвет голосеменных растений? В какой период появились хвойные?
Какие группы голосеменных растений полностью вымерли, и когда это произошло?
В каком периоде появились первые покрытосеменные растения?
Как изменялся растительный мир в течение кайнозойской эры?
В какой период истории Земли растительный мир принял современный облик?

Plant Timeline Evolution: Строительство | StudySmarter
Наземные растения оказали огромное влияние на Землю, и без них мир, в котором мы живем, был бы совершенно неузнаваем. Изменения в физической географии и атмосфере Земли не произошли из ниоткуда — они произошли с появлением нескольких ключевых адаптаций наземных растений. Реконструкция хронологии истории эволюции наземных растений позволяет нам путешествовать во времени и отслеживать эти колоссальные изменения.
Изменения в физической географии и атмосфере Земли не произошли из ниоткуда — они произошли с появлением нескольких ключевых адаптаций наземных растений. Реконструкция хронологии истории эволюции наземных растений позволяет нам путешествовать во времени и отслеживать эти колоссальные изменения.
Строительство линии развития завода
Современные виды можно проследить до их предков через филогенетических деревьев . Эти деревья объединяют ископаемые, химические, морфологические и геологические данные для реконструкции временной шкалы эволюции. Летопись окаменелостей — наш самый надежный источник при построении хронологии эволюции наземных растений. Информация, предоставленная этой записью, позволяет нам разместить появление и эволюцию различных признаков в геологической шкале времени. Это особенно важно, поскольку из этого мы можем увидеть новые шаги в эволюции наряду с предполагаемой средой, в которой они произошли. Безусловно, величайшим открытием, обнаруженным в летописи окаменелостей, является возрастающая скорость эволюции и сложность сообществ по мере того, как растения укореняются в среде обитания. недавно завоеванная земная среда.
недавно завоеванная земная среда.
Эволюция временной шкалы растений: эволюционная история растений
Из этих реконструкций мы можем увидеть четыре ключевых этапа эволюции временной шкалы эволюции растений. Это эволюция:
Эти ключевые шаги, вероятно, отражают сильное давление отбора и проблемы, с которыми растения сталкивались на протяжении всей своей эволюционной истории.
Проблемы, с которыми сталкивались наземные растения на протяжении всей своей эволюционной истории
Хотя выход на сушу не обошлось без трудностей, у успешных колонизирующих организмов, несомненно, было несколько преимуществ. Поскольку растения завоевали земной мир задолго до своих животных собратьев, ранним наземным растениям изначально была предоставлена роскошь отсутствия хищников. Другие благоприятные аспекты земной среды включают большее содержание CO2 и более широкий доступ к солнечным лучам. И то, и другое позволило ранним растениям больше фотосинтезировать и, следовательно, получить больше энергии для распространения, роста и размножения.
Хотя те же самые благоприятные аспекты земной среды представляли собой собственные факторы стресса и трудности, которые ранние виды растений должны были преодолеть, чтобы успешно превратиться в разнообразные растения, известные нам сегодня. Вода может фильтровать солнечный свет и уменьшать количество энергии, поглощаемой пигментами хлорофилла, но она также действует как барьер против вредного ультрафиолетового излучения. Барьер, которого нет на суше. Диффузия питательных веществ в воде может быть медленнее, но вода в водной среде обеспечивает плавучесть, поддержку и, что особенно важно, воду для ее обитателей. Все это первые наземные растения нуждались в дополнении новыми морфологическими приспособлениями. Риск высыхания на суше можно считать почти постоянным.
Основные факторы стресса, такие как высыхание, повреждение ультрафиолетовым излучением, отсутствие структурной поддержки и «плавательная» среда для обмена гаметами при размножении, подчеркивают важность ранней эволюции наземных растений. Как только первые наземные растения успешно прижились в исходной земной среде, они развили множество приспособлений, которые способствовали успеху как видов, так и популяций и их расселению в более широкой, более суровой наземной среде. Однако эти изменения произошли не сразу, и их можно проследить во времени по летописи окаменелостей.
Как только первые наземные растения успешно прижились в исходной земной среде, они развили множество приспособлений, которые способствовали успеху как видов, так и популяций и их расселению в более широкой, более суровой наземной среде. Однако эти изменения произошли не сразу, и их можно проследить во времени по летописи окаменелостей.
Хронология эволюции растений в течение геологических периодов
Растения прошли 4 ключевых этапа в своей эволюционной истории: переход на сушу , за которой последовала эволюция сосудистой ткани, семян и пыльцы, а также цветковых растений. Каждая из этих адаптаций возникла в разные геологические периоды и в последовательном порядке.
Древнейшие наземные растения (мохообразные и зеленые водоросли)
После того как предковая стрептофитная водоросль мигрировала на сушу, она прошла несколько ключевых этапов эволюции, прежде чем произвести разнообразие современных наземных растений. Ранние наземные растения на начальных этапах эволюции растений наиболее близки к мохообразным. Мохообразные , вероятно, возникли 450 миллионов лет назад во время ордовикского периода . Печеночники являются наиболее близкими существующими растениями к предковым зеленым водорослям. 1
Ранние наземные растения на начальных этапах эволюции растений наиболее близки к мохообразным. Мохообразные , вероятно, возникли 450 миллионов лет назад во время ордовикского периода . Печеночники являются наиболее близкими существующими растениями к предковым зеленым водорослям. 1
Летопись окаменелостей ранних наземных растений скудна, поскольку они были сделаны из мягкого органического материала. Изучая мохообразные и печеночники, мы можем больше узнать о ранних наземных растениях и предположить следующие ключевые характеристики:
Ранние наземные растения, вероятно, были небольшими. В воздухе им не хватало поддержки и способности переносить воду, поэтому они выживали близко к земле во влажной среде.
Ранние наземные растения не имели сосудов, то есть у них не было специализированной транспортной системы.
Ранние наземные растения использовали воду для полового размножения.
Единственным способом транспортировки мужских гамет или сперматозоидов было плавание по воде с помощью жгутика.
 Единственным способом транспортировки мужских гамет или сперматозоидов было плавание по воде с помощью жгутика.
Единственным способом транспортировки мужских гамет или сперматозоидов было плавание по воде с помощью жгутика.Эволюция сосудистых растений
Следующим этапом эволюции растений является развитие сосудистых систем . Сосудистая система растений похожа на наши вены и артерии. Они транспортируют воду и питательные вещества к различным частям растения. Сосудистые растения, вероятно, возникли в конце IX в.0007 Ордовикский период , в результате высокой конкуренции за солнечный свет. Сосудистые системы позволили растениям вырасти выше и получить доступ к большему количеству ресурсов, поскольку они больше не были ограничены диффузией и осмосом для транспортировки основных питательных веществ и воды. С эволюцией сосудистой ткани произошло развитие настоящих корней . Корни позволяли растениям получать больше воды и закрепляли растения для устойчивости по мере их роста. Ранние сосудистые растения предшествовали эволюции семян.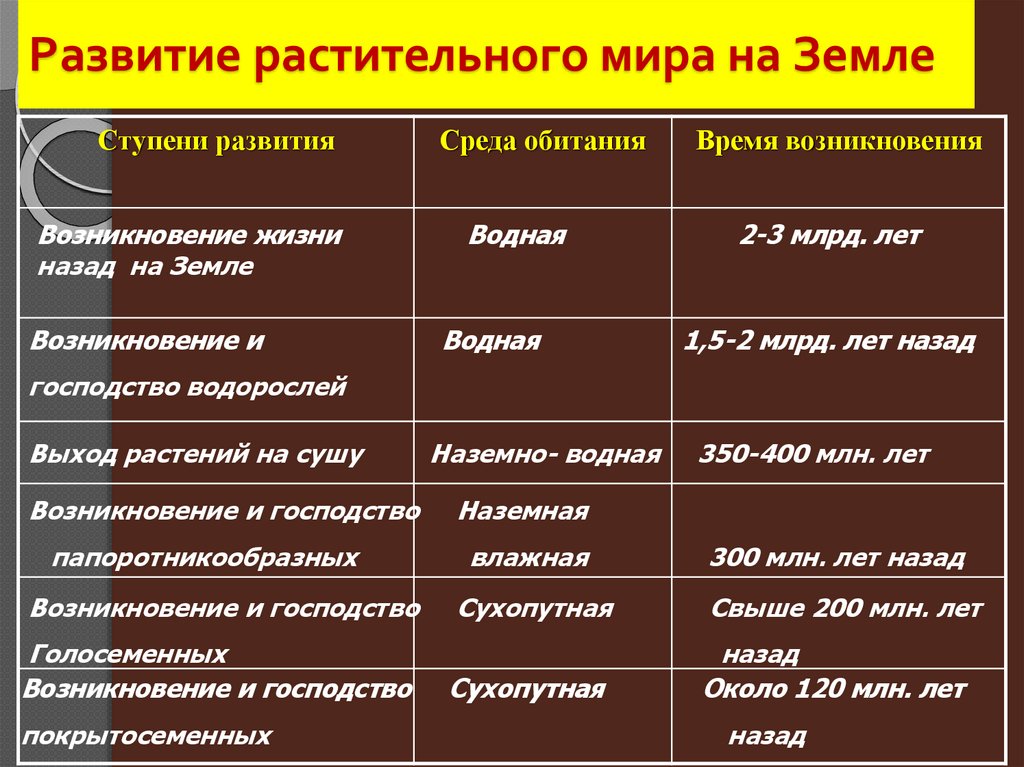 Как и несосудистые мохообразные, бессемянные сосудистые растения нуждаются в воде для полового размножения.
Как и несосудистые мохообразные, бессемянные сосудистые растения нуждаются в воде для полового размножения.
Папоротники и плауны являются современными потомками ранних сосудистых растений. Оба растения бессемянные, размножаются спорами.
Эволюция семян в биологии растений
До эволюции семян для полового размножения растений требовалась вода. Но с появлением семян игра полностью изменилась.
Бессемянные наземные растения производят споры, которые часто разносятся ветром. Это только половина истории репродукции. Все растения эволюционировали, чтобы размножаться посредством смена поколений . Это означает, что растения чередуются между двумя разными фазами организмов: гаметофитами и спорофитами. Гаметофиты размножаются с помощью половых клеток и митоза, спорофиты производят споры, которые позже прорастают в гаметофиты. Во время размножения гаметофитов у бессемянных наземных растений сперматозоиды оплодотворяют яйцеклетку, сперматозоиды приспособлены только для того, чтобы добраться до яйцеклетки, плавая по воде. Поэтому вода необходима для бессемянных удобрений.
Поэтому вода необходима для бессемянных удобрений.
С развитием семян и пыльцы эта потребность в воде для размножения исчезает. У посевных наземных растений мужские гаметы содержатся в пыльце, которая обеспечивает защитную оболочку от высыхания. Эти пыльцевые зерна перемещаются между людьми по ветру или прилипают к насекомым. Женские гаметы остаются прикрепленными к родительскому растению в семяпочках до тех пор, пока они не будут оплодотворены пыльцевыми зернами. Когда происходит оплодотворение, образуется зародыш, а семязачаток развивается в семя. Это семя образует защитную оболочку, в которой находится эмбрион вместе с некоторыми важными питательными веществами.
Семя содержит все питательные вещества, необходимые для выживания эмбриона. Позволить зародышу лежать в спящем состоянии, не высыхая, по мере рассеивания семени. Эволюция семян является полезной адаптацией в истории эволюции наземных растений, поскольку:
Семена устранили потребность в водной среде для размножения.
Семена содержат питательные вещества, которые позволяют развивающимся зародышам растений находиться в состоянии покоя до тех пор, пока условия не будут благоприятны для их роста и выживания.
Семена имеют защитную оболочку, которая увеличивает диапазон распространения новых растений, снижая их конкуренцию за ресурсы с родителями.
Появление голосеменных и покрытосеменных растений
Первыми семенными наземными растениями, которые эволюционировали, были растения с «голыми семенами» или голосеменные растения . Ранние голосеменные растения, впервые развившиеся в девонском или С каменноугольном периоде , производили специализированные мужские и женские споры в разных шишках. Много миллионов лет спустя, в меловой период 9Развились покрытосеменные 0007 , которые производят мужские и женские структуры внутри цветков. Несмотря на то, что покрытосеменные появились намного позже голосеменных, они считаются сестринскими ветвями. Последний общий предок всех голосеменных также считается последним общим предком всех голосеменных и покрытосеменных растений.
Последний общий предок всех голосеменных также считается последним общим предком всех голосеменных и покрытосеменных растений.
Голосеменные: Сосудистые растения, размножающиеся незащищенными семенами. Открытые семена не защищены завязью или плодом, как у покрытосеменных.
Покрытосеменные: Цветковые сосудистые растения, размножающиеся семенами с оболочкой. Как и у голосеменных, семена покрытосеменных возникают из семязачатков. Однако, в отличие от голосеменных, семязачатки покрытосеменных содержатся внутри завязей, которые после оплодотворения превращаются в плоды.
Эволюция семян и пыльцы положила начало миграции наземных растений во все земные ниши, что позволило растениям выжить на суше. Эволюция семенных растений сыграла большую роль в успехе царства растений, господствующего на земном шаре. Позволить голосеменным наземным растениям доминировать в земном ландшафте с момента их эволюционного появления до Триасовый и Юрский периоды . Только для того, чтобы быть замененным в качестве «короля» семенных растений 100 миллионов лет назад покрытосеменными растениями и их цветами в конце мезозойской эры .
Только для того, чтобы быть замененным в качестве «короля» семенных растений 100 миллионов лет назад покрытосеменными растениями и их цветами в конце мезозойской эры .
Хронология эволюции цветковых растений
Предки цветковых растений отделились от голосеменных 300 миллионов лет назад во время C каменноугольного периода . Однако с этого момента хронология эволюции цветковых растений становится загадкой.
Нет непрерывных ископаемых свидетельств эволюции цветов. Появление покрытосеменных трудно определить. Ископаемые свидетельства, по-видимому, указывают на то, что покрытосеменные отделились от голосеменных в триасовом периоде . Однако генетический анализ предполагает, что раскол произошел гораздо раньше в девонском периоде . Это подтверждается наличием окаменевшей пыльцы, похожей на пыльцу покрытосеменных.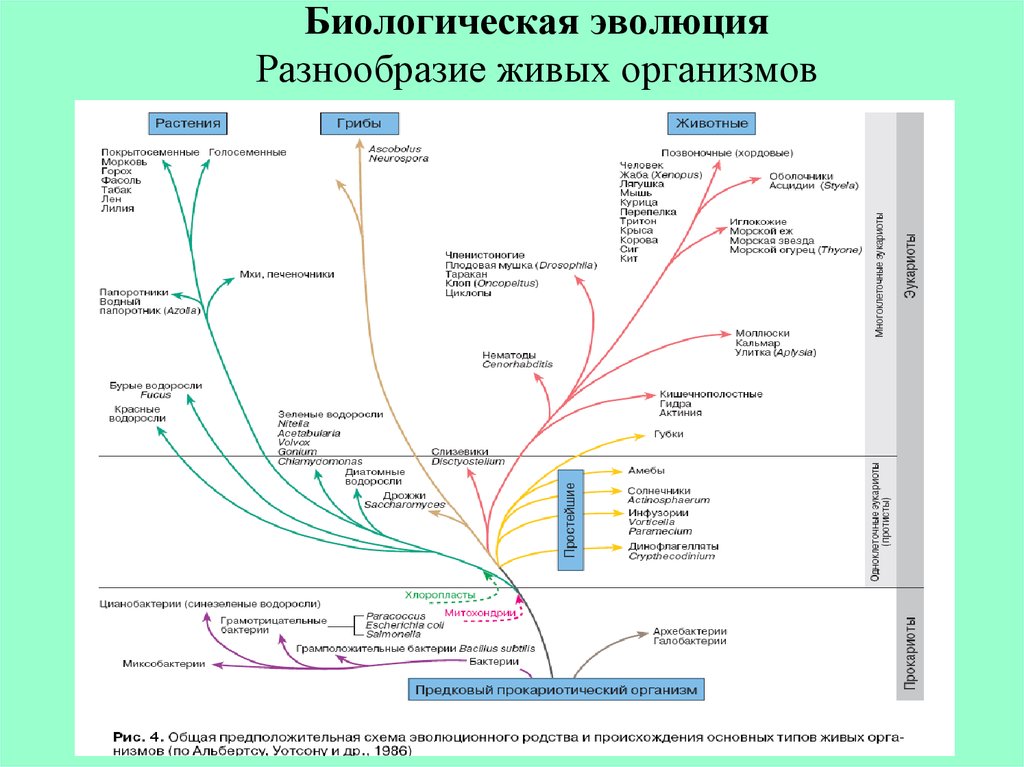
Первоначально появление цветущих семенных растений, казалось, происходило одновременно. Этот бум был настолько резким, что Чарльз Дарвин назвал этот конкретный шаг в эволюции «отвратительной тайной». Во многом это произошло из-за пробелов в летописи окаменелостей, и хотя коллекции выросли с тех пор, как хронология эволюции цветковых растений до сих пор окончательно не взломана.
В течение периода C retaceous покрытосеменные, или цветковые растения, широко распространились, заменив и вытеснив предыдущих обитателей папоротников. Считается, что в этот период покрытосеменные быстро сократили размер своего генома, что позволило им размножаться намного быстрее благодаря более быстрому делению клеток. Этот сдвиг, вероятно, является причиной того, что покрытосеменные в настоящее время составляют 90% всех растений.
На протяжении всей своей эволюционной истории у большинства покрытосеменных было симбиотические отношения с опылителем. Покрытосеменные растения и соответствующие им опылители являются прекрасным примером коэволюции . Покрытосеменные эволюционировали, чтобы иметь ярко окрашенные цветы разных размеров, форм и ароматов, содержащие сладкий нектар, чтобы привлекать опылителей и обслуживать их. Пыльца покрытосеменных растений прилипает к опылителям и уносится с ними, когда они посещают и, как мы надеемся, опыляют следующее растение. Опылители развили такие черты, как более длинные языки и клювы, чтобы лучше доставать нектар, которым они питаются. В то время как покрытосеменные растения продолжают делать этот нектар более сладким и привлекательным для друзей-опылителей.
Покрытосеменные растения и соответствующие им опылители являются прекрасным примером коэволюции . Покрытосеменные эволюционировали, чтобы иметь ярко окрашенные цветы разных размеров, форм и ароматов, содержащие сладкий нектар, чтобы привлекать опылителей и обслуживать их. Пыльца покрытосеменных растений прилипает к опылителям и уносится с ними, когда они посещают и, как мы надеемся, опыляют следующее растение. Опылители развили такие черты, как более длинные языки и клювы, чтобы лучше доставать нектар, которым они питаются. В то время как покрытосеменные растения продолжают делать этот нектар более сладким и привлекательным для друзей-опылителей.
Хронология развития растений — основные выводы
Эволюцию растений можно проследить по летописи окаменелостей, начиная с зарождения жизни ~3,7 миллиона лет назад.
Наземные растения столкнулись с большим количеством стрессоров, когда они впервые появились на суше.
Эти стрессоры вызвали сильное давление отбора и сформировали эволюцию наземных растений.В истории эволюции растений выделяют четыре ключевых последовательных этапа.
Хотя цветковые растения составляют 90% современной флоры, в летописи окаменелостей имеются значительные пробелы. Мало что известно о точном эволюционном пути цветковых растений после того, как они отделились от голосеменных растений в позднем девонском периоде.
Покрытосеменные растения и соответствующие им опылители являются прекрасным примером совместной эволюции.
 Эти стрессоры вызвали сильное давление отбора и сформировали эволюцию наземных растений.
Эти стрессоры вызвали сильное давление отбора и сформировали эволюцию наземных растений.Ссылки
- Lumen, Bryophytes, Boundless Biology
- C. Jill Harrison et al, Происхождение и ранняя эволюция побегов и листьев сосудистых растений, Philosophical Transactions of the Royal Society B, 2018.
- Т. Джонатан Дэвис и др., «Отвратительная тайна Дарвина: понимание супердерева покрытосеменных растений», Труды Национальной академии наук Соединенных Штатов Америки, 2004 г.

Ранняя жизнь растений | Биология для специальности II
Результаты обучения
- Описать хронологию эволюции растений и влияние наземных растений на другие живые существа
Царство Plantae включает большие и разнообразные группы организмов. Существует более 300 000 видов каталогизированных растений. Из них более 260 000 семенных растений. Мхи, папоротники, хвойные и цветковые растения — все это представители царства растений. Наземные растения возникли в составе Archaeplastida, включающего красные водоросли (Rhodophyta) и две группы зеленых водорослей, Chlorophyta и Charaphyta . Большинство биологов также считают по крайней мере некоторые зеленые водоросли растениями, хотя другие исключают все водоросли из царства растений. Причина этого разногласия связана с тем фактом, что только зеленые водоросли, хлорофиты и харофиты, имеют общие характеристики с наземными растениями (например, использование хлорофилла a и b плюс каротин в той же пропорции, что и растения). Эти характеристики отсутствуют у других видов водорослей.
Причина этого разногласия связана с тем фактом, что только зеленые водоросли, хлорофиты и харофиты, имеют общие характеристики с наземными растениями (например, использование хлорофилла a и b плюс каротин в той же пропорции, что и растения). Эти характеристики отсутствуют у других видов водорослей.
Водоросли и пути эволюции к фотосинтезу
Одни ученые считают растениями все водоросли, другие утверждают, что только зеленые водоросли принадлежат к царству Plantae. Третьи включают среди растений только Charophytes. Эти расхождения во мнениях связаны с разными эволюционными путями фотосинтеза, выбранными для разных типов водорослей. Хотя все водоросли являются фотосинтезирующими, то есть содержат ту или иную форму хлоропласта, они не все стали фотосинтезирующими одним и тем же путем.
Предки Archaeplastida стали фотосинтезирующими, установив эндосимбиотические отношения с зеленой фотосинтезирующей бактерией около 1,65 миллиарда лет назад. Эта линия водорослей превратилась в красные и зеленые водоросли и, в конечном итоге, в современные мхи, папоротники, голосеменные и покрытосеменные растения. Их эволюционная траектория была относительно прямой и монофилетической. Напротив, водоросли за пределами Archaeplastida, например бурые и золотистые водоросли страменопилов и т. д., — все стали фотосинтезирующими в результате вторичных или даже третичных эндосимбиотических событий; то есть они поглотили клетки, которые уже содержали эндосимбиотическую цианобактерию. Эти опоздавшие к фотосинтезу аналогичны Archaeplastida с точки зрения автотрофии, но они не расширились до такой степени, как Archaeplastida, и не колонизировали сушу.
Эта линия водорослей превратилась в красные и зеленые водоросли и, в конечном итоге, в современные мхи, папоротники, голосеменные и покрытосеменные растения. Их эволюционная траектория была относительно прямой и монофилетической. Напротив, водоросли за пределами Archaeplastida, например бурые и золотистые водоросли страменопилов и т. д., — все стали фотосинтезирующими в результате вторичных или даже третичных эндосимбиотических событий; то есть они поглотили клетки, которые уже содержали эндосимбиотическую цианобактерию. Эти опоздавшие к фотосинтезу аналогичны Archaeplastida с точки зрения автотрофии, но они не расширились до такой степени, как Archaeplastida, и не колонизировали сушу.
Ученые, отслеживающие исключительно прямые линии эволюции (то есть монофилию), считают растениями только харофиты. Общий предок Charophytes и наземных растений исключает других членов Archaeplastida. Харофиты также имеют общие черты с наземными растениями.
Перейдите к этой статье, чтобы получить более подробную информацию о Charophytes.
Адаптация растений к жизни на суше
Поскольку организмы приспособились к жизни на суше, им пришлось столкнуться с рядом проблем в земной среде. Вода была описана как «вещество жизни». Внутренняя часть клетки представляет собой густой суп: в этой среде растворяется и диффундирует большинство малых молекул, и происходит большинство химических реакций обмена веществ. Иссушение, или высыхание, представляет постоянную опасность для организма, подвергающегося воздействию воздуха. Даже когда части растения находятся близко к источнику воды, надземные части, скорее всего, высохнут. Вода также обеспечивает плавучесть организмам. На суше растениям необходимо развивать структурную поддержку в среде, которая не дает такой подъемной силы, как вода. Организм также подвергается бомбардировке мутагенным излучением, так как воздух не фильтрует ультрафиолетовые лучи солнечного света. Кроме того, мужские гаметы должны достичь женских гамет, используя новые стратегии, потому что плавание больше невозможно. Следовательно, и гаметы, и зиготы должны быть защищены от высыхания. Успешные наземные заводы разработали стратегии для решения всех этих проблем. Не все приспособления появились сразу. Некоторые виды никогда не уходили далеко от водной среды, в то время как другие продолжали завоевывать самые засушливые места на Земле.
Следовательно, и гаметы, и зиготы должны быть защищены от высыхания. Успешные наземные заводы разработали стратегии для решения всех этих проблем. Не все приспособления появились сразу. Некоторые виды никогда не уходили далеко от водной среды, в то время как другие продолжали завоевывать самые засушливые места на Земле.
Чтобы сбалансировать эти проблемы выживания, жизнь на суше предлагает несколько преимуществ. Во-первых, много солнечного света. Вода действует как фильтр, изменяя спектральное качество света, поглощаемого фотосинтетическим пигментом хлорофиллом. Во-вторых, углекислый газ более доступен в воздухе, чем в воде, поскольку он быстрее диффундирует в воздухе. В-третьих, наземные растения возникли раньше наземных животных; поэтому, пока суша не была заселена животными, никакие хищники не угрожали жизни растений. Эта ситуация изменилась, когда животные вышли из воды и питались обильными источниками питательных веществ укоренившейся флоры. В свою очередь, растения разработали стратегии отпугивания хищников: от шипов и шипов до ядовитых химикатов.
Ранние наземные растения, как и ранние наземные животные, жили недалеко от обильного источника воды и разработали стратегии выживания для борьбы с засухой. Одна из этих стратегий называется толерантностью. Многие мхи, например, могут высохнуть и превратиться в коричневый и ломкий мат, но как только дождь или наводнение сделает воду доступной, мхи впитают ее и вернут свой здоровый зеленый вид. Другая стратегия заключается в колонизации среды с высокой влажностью, где засухи случаются редко. Папоротники, которые считаются ранней линией растений, процветают во влажных и прохладных местах, таких как подлесок умеренных лесов. Позже растения ушли из влажной или водной среды, используя устойчивость к высыханию, а не толерантность. Эти растения, как и кактусы, сводят к минимуму потерю воды до такой степени, что могут выжить в чрезвычайно сухой среде.
Наиболее успешным адаптационным решением была разработка новых структур, которые давали растениям преимущество при колонизации новой и сухой среды. Успеху наземных растений способствуют четыре основных приспособления. Первая адаптация заключается в том, что в жизненном цикле всех наземных растений наблюдается чередование поколений: спорофита, в котором образуются споры, и гаметофита, производящего гаметы. Во-вторых, ткань апикальной меристемы в корнях и побегах. В-третьих, это эволюция восковой кутикулы, которая сопротивляется высыханию (отсутствует у некоторых мхов). Наконец, клеточные стенки с лигнином для поддержки структур над землей. Все эти адаптации способствуют успеху наземных растений, но их явно не хватает близкородственным зеленым водорослям — еще одна причина споров об их месте в царстве растений. Они также не все встречаются у мхов, которые можно рассматривать как представляющие собой промежуточную стадию адаптации к суше.
Успеху наземных растений способствуют четыре основных приспособления. Первая адаптация заключается в том, что в жизненном цикле всех наземных растений наблюдается чередование поколений: спорофита, в котором образуются споры, и гаметофита, производящего гаметы. Во-вторых, ткань апикальной меристемы в корнях и побегах. В-третьих, это эволюция восковой кутикулы, которая сопротивляется высыханию (отсутствует у некоторых мхов). Наконец, клеточные стенки с лигнином для поддержки структур над землей. Все эти адаптации способствуют успеху наземных растений, но их явно не хватает близкородственным зеленым водорослям — еще одна причина споров об их месте в царстве растений. Они также не все встречаются у мхов, которые можно рассматривать как представляющие собой промежуточную стадию адаптации к суше.
Смена поколений
Все организмы, размножающиеся половым путем, имеют в своем жизненном цикле как гаплоидные, так и диплоидные клетки. У организмов с гаплонтическими жизненными циклами доминирует гаплоидная стадия, а у организмов с диплоидным жизненным циклом доминирует диплоидная стадия. Доминантный в данном контексте означает как стадию, на которой организм проводит большую часть своего времени, так и стадию, на которой происходит наибольшее митотическое размножение клеток, — многоклеточную стадию. В гаплотических жизненных циклах единственной диплоидной клеткой является зигота, которая подвергается немедленному мейозу для восстановления гаплоидного состояния. В диплоидных жизненных циклах единственными гаплоидными клетками являются гаметы, которые объединяются, чтобы восстановить диплоидное состояние при первой же возможности. Люди, например, диплонтны.
Доминантный в данном контексте означает как стадию, на которой организм проводит большую часть своего времени, так и стадию, на которой происходит наибольшее митотическое размножение клеток, — многоклеточную стадию. В гаплотических жизненных циклах единственной диплоидной клеткой является зигота, которая подвергается немедленному мейозу для восстановления гаплоидного состояния. В диплоидных жизненных циклах единственными гаплоидными клетками являются гаметы, которые объединяются, чтобы восстановить диплоидное состояние при первой же возможности. Люди, например, диплонтны.
Чередование поколений описывает жизненный цикл, в котором организм имеет как гаплоидные, так и диплоидные многоклеточные стадии (рис. 1). Этот тип жизненного цикла, который встречается у всех растений, описывается как гаплодиплонтический .
Рисунок 1. Показано чередование поколений между гаметофитом 1 n и спорофитом 2 n .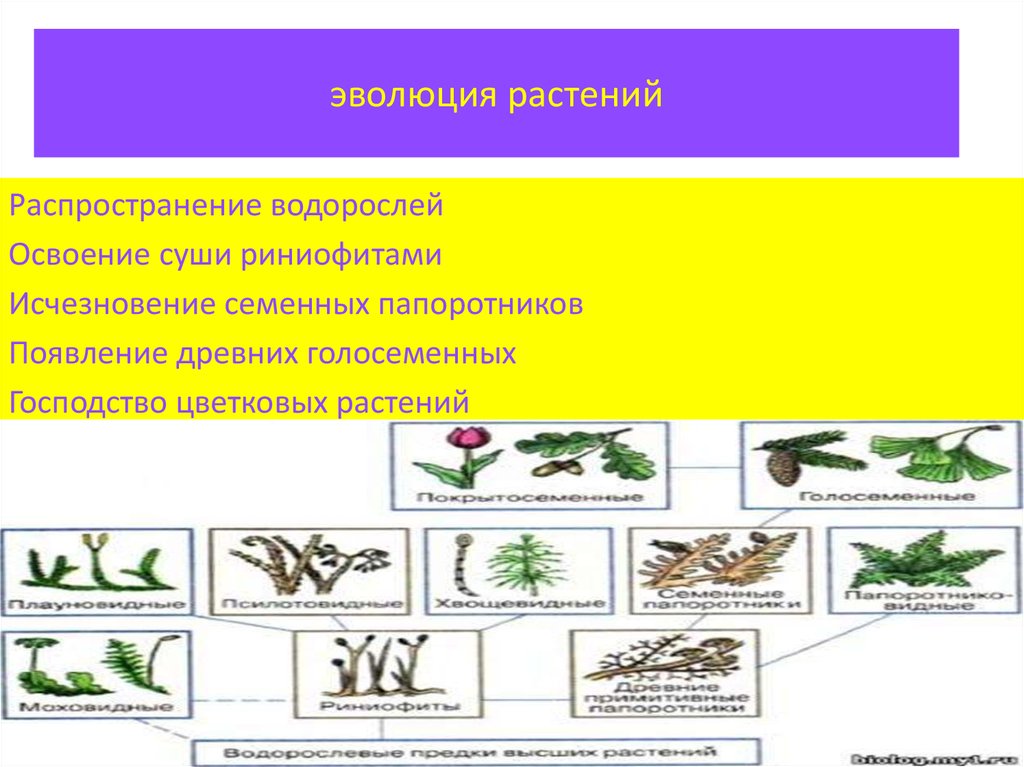 Митоз происходит как в гаметофитном, так и в спорофитном поколении. Диплоидные спорофиты производят гаплоидные споры путем мейоза, тогда как гаплоидные гаметофиты производят гаметы путем митоза. (кредит: Питер Коксхед)
Митоз происходит как в гаметофитном, так и в спорофитном поколении. Диплоидные спорофиты производят гаплоидные споры путем мейоза, тогда как гаплоидные гаметофиты производят гаметы путем митоза. (кредит: Питер Коксхед)
В чередовании поколений за многоклеточной гаплоидной формой, известной как гаметофит, следует в последовательности развития многоклеточная диплоидная форма, спорофит. Гаметофит дает начало гаметам (половым клеткам) путем митоза. Это может быть наиболее очевидная фаза жизненного цикла растения, как у мхов, или она может проявляться в микроскопической структуре, такой как пыльцевое зерно, в семенных растениях. Эволюция наземных растений отмечена возрастающим преобладанием поколения спорофитов. Стадия спорофита едва заметна у несосудистых растений (собирательный термин для растений, к которым относятся печеночники и мхи). У семенных растений фаза спорофита может быть высоким деревом, как у секвойи и сосны.
Защита зародыша является основным требованием для наземных растений. Уязвимый эмбрион должен быть защищен от высыхания и других опасностей окружающей среды. Как у бессемянных, так и у семенных растений женский гаметофит обеспечивает защиту и питательные вещества для зародыша, когда он развивается в новый спорофит. Эта отличительная черта наземных растений дала группе альтернативное название эмбриофитов .
Уязвимый эмбрион должен быть защищен от высыхания и других опасностей окружающей среды. Как у бессемянных, так и у семенных растений женский гаметофит обеспечивает защиту и питательные вещества для зародыша, когда он развивается в новый спорофит. Эта отличительная черта наземных растений дала группе альтернативное название эмбриофитов .
Спорангии в бессемянных растениях
Рисунок 2. На этой фотографии мха Esporangios bryum на концах длинных тонких стеблей растут спорообразующие мешочки, называемые спорангиями. (кредит: Хавьер Мартин)
Спорофит бессемянных растений является диплоидным и возникает в результате сингамии (слияния) двух гамет. Спорофит несет спорангии (единственные, спорангии). Термин «спорангий» буквально означает «сосуд для спор», так как это репродуктивный мешок, в котором образуются споры (рис. 2). Внутри многоклеточных спорангиев диплоидные спороциты , или материнские клетки, производят гаплоидные споры путем мейоза, в ходе которого число хромосом 2 n снижается до 1 n (обратите внимание, что у многих растений число хромосом усложняется полиплоидией). : например, твердая пшеница — тетраплоид, мягкая пшеница — гексаплоид, а некоторые папоротники — 1000-плоид). Позже споры высвобождаются из спорангиев и рассеиваются в окружающей среде. Когда гаплоидная спора прорастает в гостеприимной среде, она образует многоклеточный гаметофит путем митоза. Гаметофит поддерживает зиготу, образовавшуюся в результате слияния гамет, и образовавшегося молодого спорофита (вегетативная форма). Затем цикл начинается заново.
: например, твердая пшеница — тетраплоид, мягкая пшеница — гексаплоид, а некоторые папоротники — 1000-плоид). Позже споры высвобождаются из спорангиев и рассеиваются в окружающей среде. Когда гаплоидная спора прорастает в гостеприимной среде, она образует многоклеточный гаметофит путем митоза. Гаметофит поддерживает зиготу, образовавшуюся в результате слияния гамет, и образовавшегося молодого спорофита (вегетативная форма). Затем цикл начинается заново.
Растения, образующие споры только одного типа, называются гомоспоровыми, а образующийся в результате гаметофит производит как мужские, так и женские гаметы, обычно у одной и той же особи. Несосудистые растения гомоспоровые, а гаметофит является доминирующим поколением в жизненном цикле. Растения, образующие два типа спор, называются разноспоровыми. Мужские споры называются микроспорами из-за их меньшего размера и развиваются в мужской гаметофит; сравнительно более крупные мегаспоры развиваются в женский гаметофит. Некоторые бессемянные сосудистые растения и все семенные растения являются гетероспористыми, а спорофит является доминирующим поколением.
Споры бессемянных растений окружены толстыми клеточными стенками, содержащими прочный полимер, известный как спорополленин. Как следует из названия, он также содержится в стенках пыльцевых зерен. Это сложное вещество характеризуется длинными цепочками органических молекул, связанных с жирными кислотами и каротиноидами: отсюда и желтый цвет большинства пыльцы. Спорополленин необычайно устойчив к химическому и биологическому разложению. У семенных растений, у которых пыльца является мужским гаметофитом, устойчивость спорополленина объясняет существование хорошо сохранившихся окаменелостей пыльцы. Спорополленин когда-то считался нововведением наземных растений; однако харофит Coleochaetes также образует споры, содержащие спорополленин .
Гаметангии бессемянных растений
Гаметангии (единственное число, гаметангии) представляют собой структуры, наблюдаемые на многоклеточных гаплоидных гаметофитах. В гаметангиях клетки-предшественники дают начало гаметам путем митоза. Мужской гаметангий ( антеридий ) выделяет сперму. Многие бессемянные растения производят сперму, оснащенную жгутиками, которые позволяют им плавать во влажной среде до архегония : женский гаметангий. Зародыш развивается внутри архегония как спорофит. Гаметангии заметны у бессемянных растений, но очень редко встречаются у семенных растений.
Мужской гаметангий ( антеридий ) выделяет сперму. Многие бессемянные растения производят сперму, оснащенную жгутиками, которые позволяют им плавать во влажной среде до архегония : женский гаметангий. Зародыш развивается внутри архегония как спорофит. Гаметангии заметны у бессемянных растений, но очень редко встречаются у семенных растений.
Апикальные меристемы
Рисунок 3. Добавление новых клеток в корень происходит в апикальной меристеме. Последующее увеличение этих клеток вызывает рост и удлинение органа. Корневой чехлик защищает хрупкую апикальную меристему, поскольку кончик корня проталкивается через почву за счет удлинения клеток.
Побеги и корни растений увеличиваются в длину за счет быстрого деления клеток в ткани, называемой апикальной меристемой, которая представляет собой небольшую зону клеток, расположенную на кончике побега или кончике корня (рис. 3). Апикальная меристема состоит из недифференцированных клеток, которые продолжают размножаться на протяжении всей жизни растения. Меристематические клетки дают начало всем специализированным тканям организма. Удлинение побегов и корней позволяет растению получить доступ к дополнительному пространству и ресурсам: свету в случае побега и воде и минеральным веществам в случае корней. Отдельная меристема, называемая боковой меристемой, производит клетки, увеличивающие диаметр стволов деревьев.
Меристематические клетки дают начало всем специализированным тканям организма. Удлинение побегов и корней позволяет растению получить доступ к дополнительному пространству и ресурсам: свету в случае побега и воде и минеральным веществам в случае корней. Отдельная меристема, называемая боковой меристемой, производит клетки, увеличивающие диаметр стволов деревьев.
Дополнительная адаптация наземных растений
По мере того, как растения адаптировались к засушливой земле и становились независимыми от постоянного присутствия воды во влажных местах обитания, у них появлялись новые органы и структуры. Ранние наземные растения не вырастали выше нескольких дюймов от земли, конкурируя за свет на этих низких циновках. Развивая побег и становясь выше, отдельные растения улавливали больше света. Поскольку воздух обеспечивает значительно меньшую поддержку, чем вода, наземные растения содержат более жесткие молекулы в своих стеблях (а позже и в стволах деревьев). У небольших растений, таких как одноклеточные водоросли, достаточно простой диффузии для распределения воды и питательных веществ по всему организму.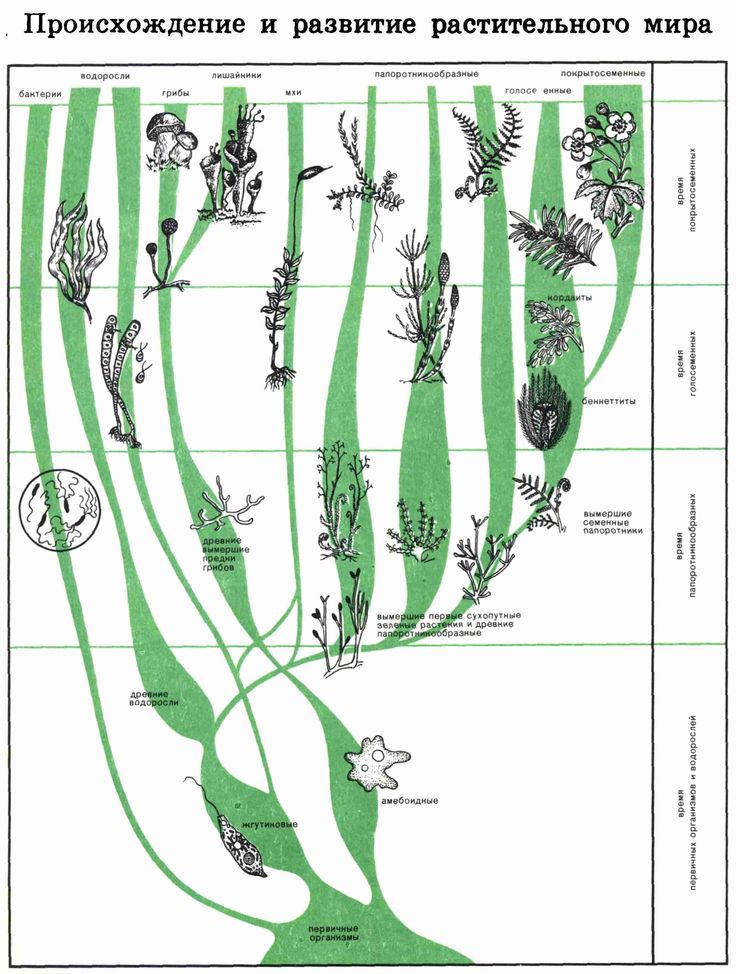 Однако для развития более крупных форм у растений предпосылкой была эволюция сосудистой ткани для распределения воды и растворенных веществ. Сосудистая система состоит из тканей ксилемы и флоэмы. Ксилема проводит воду и минеральные вещества, поглощенные из почвы, к побегам, а флоэма переносит пищевые продукты, полученные в результате фотосинтеза, по всему растению. Корневая система развилась, чтобы поглощать воду и минералы из почвы и закреплять в почве все более высокие побеги.
Однако для развития более крупных форм у растений предпосылкой была эволюция сосудистой ткани для распределения воды и растворенных веществ. Сосудистая система состоит из тканей ксилемы и флоэмы. Ксилема проводит воду и минеральные вещества, поглощенные из почвы, к побегам, а флоэма переносит пищевые продукты, полученные в результате фотосинтеза, по всему растению. Корневая система развилась, чтобы поглощать воду и минералы из почвы и закреплять в почве все более высокие побеги.
У наземных растений восковой водостойкий покров, называемый кутикулой, защищает листья и стебли от высыхания. Однако кутикула также предотвращает поступление углекислого газа, необходимого для синтеза углеводов посредством фотосинтеза. Чтобы преодолеть это, у растений появились устьица или поры, которые открываются и закрываются, чтобы регулировать поток газов и водяного пара, когда они перемещаются из влажной среды в более сухую среду обитания.
Вода фильтрует ультрафиолетовое излучение типа B (UVB), которое вредно для всех организмов, особенно для тех, которые должны поглощать свет, чтобы выжить. Эта фильтрация не происходит для наземных растений. Это создало дополнительную проблему для колонизации суши, которая была решена эволюцией путей биосинтеза для синтеза защитных флавоноидов и других соединений: пигментов, которые поглощают УФ-свет и защищают надземные части растений от фотодинамического повреждения.
Эта фильтрация не происходит для наземных растений. Это создало дополнительную проблему для колонизации суши, которая была решена эволюцией путей биосинтеза для синтеза защитных флавоноидов и других соединений: пигментов, которые поглощают УФ-свет и защищают надземные части растений от фотодинамического повреждения.
Растения не могут избежать поедания животными. Вместо этого они синтезируют большое количество ядовитых вторичных метаболитов: сложные органические молекулы, такие как алкалоиды, чей ядовитый запах и неприятный вкус отпугивают животных. Эти токсичные соединения также могут вызывать тяжелые заболевания и даже смерть, что препятствует хищничеству. Люди веками использовали многие из этих соединений в качестве лекарств, лекарств или специй. Напротив, поскольку растения развивались вместе с животными, развитие сладких и питательных метаболитов привлекало животных к оказанию ценной помощи в распространении пыльцевых зерен, фруктов или семян. Таким образом растения привлекали животных себе в помощники на протяжении сотен миллионов лет.
Эволюция наземных растений
Обсуждение эволюции наземных растений невозможно без краткого обзора хронологии геологических эпох. Ранняя эра, известная как палеозой, делится на шесть периодов. Он начинается с кембрийского периода, за которым следуют ордовик, силур, девон, каменноугольный период и пермь. Важнейшим событием ордовика, произошедшим более 500 миллионов лет назад, была колонизация земли предками современных наземных растений. Окаменелые клетки, кутикулы и споры ранних наземных растений датируются ордовикским периодом раннего палеозоя. Древнейшие из известных сосудистых растений обнаружены в отложениях девона. Одним из богатейших источников информации являются кремни Райни, месторождение осадочных пород, обнаруженное в Райни, Шотландия (рис. 4), где были идентифицированы встроенные окаменелости некоторых из самых ранних сосудистых растений.
Рисунок 4. Кремни Райни содержат окаменелый материал сосудистых растений. Область внутри круга содержит луковичные подземные стебли, называемые клубнелуковицами, и корневидные структуры, называемые ризоидами. (кредит b: модификация работы Питера Коксхеда на основе оригинального изображения «Smith609»/Wikimedia Commons; данные масштабной линейки от Мэтта Рассела) до сих пор живут. Вымершие сосудистые растения, классифицируемые как зостерофиллы и тримерофиты, скорее всего, не имели настоящих листьев и корней и образовывали невысокие растительные маты, размеры которых сходны с современными мхами, хотя некоторые триметофиты могли достигать метра в высоту. Более поздний род Cooksonia , которая процветала в силурийском периоде, была тщательно изучена на хорошо сохранившихся экземплярах. На отпечатках Cooksonia видны тонкие ветвящиеся стебли, оканчивающиеся чем-то вроде спорангиев. Из извлеченных образцов невозможно точно установить, имел ли Cooksonia сосудистые ткани. Окаменелости указывают на то, что к концу девонского периода ландшафт заселили папоротники, хвощи и семенные растения, давшие начало деревьям и лесам.
(кредит b: модификация работы Питера Коксхеда на основе оригинального изображения «Smith609»/Wikimedia Commons; данные масштабной линейки от Мэтта Рассела) до сих пор живут. Вымершие сосудистые растения, классифицируемые как зостерофиллы и тримерофиты, скорее всего, не имели настоящих листьев и корней и образовывали невысокие растительные маты, размеры которых сходны с современными мхами, хотя некоторые триметофиты могли достигать метра в высоту. Более поздний род Cooksonia , которая процветала в силурийском периоде, была тщательно изучена на хорошо сохранившихся экземплярах. На отпечатках Cooksonia видны тонкие ветвящиеся стебли, оканчивающиеся чем-то вроде спорангиев. Из извлеченных образцов невозможно точно установить, имел ли Cooksonia сосудистые ткани. Окаменелости указывают на то, что к концу девонского периода ландшафт заселили папоротники, хвощи и семенные растения, давшие начало деревьям и лесам.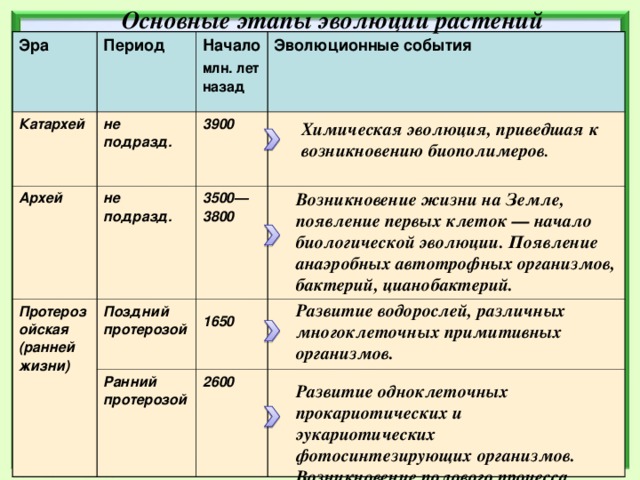 Эта пышная растительность помогла обогатить атмосферу кислородом, облегчив колонизацию суши дышащими воздухом животными. Растения также установили ранние симбиотические отношения с грибами, создав микоризу: отношения, при которых грибковая сеть нитей увеличивает эффективность корневой системы растений, а растения обеспечивают грибы побочными продуктами фотосинтеза.
Эта пышная растительность помогла обогатить атмосферу кислородом, облегчив колонизацию суши дышащими воздухом животными. Растения также установили ранние симбиотические отношения с грибами, создав микоризу: отношения, при которых грибковая сеть нитей увеличивает эффективность корневой системы растений, а растения обеспечивают грибы побочными продуктами фотосинтеза.
Палеоботаник
Каким образом организмы приобрели признаки, позволяющие им колонизировать новые среды, и как сформировалась современная экосистема, — это фундаментальные вопросы эволюции. Палеоботаника (наука о вымерших растениях) решает эти вопросы посредством анализа окаменелых образцов, извлеченных из полевых исследований, воссоздавая морфологию давно исчезнувших организмов. Палеоботаники прослеживают эволюцию растений, следя за изменениями в морфологии растений: проливая свет на связь между существующими растениями, идентифицируя общих предков, обладающих одинаковыми чертами. Эта область направлена на поиск переходных видов, которые преодолевают пробелы на пути к развитию современных организмов. Окаменелости образуются, когда организмы попадают в отложения или среду, где сохраняются их формы. Палеоботаники собирают ископаемые образцы в полевых условиях и помещают их в контекст геологических отложений и других окаменелых организмов, окружающих их. Эта деятельность требует большой осторожности, чтобы сохранить целостность хрупких окаменелостей и слоев горных пород, в которых они обнаружены.
Окаменелости образуются, когда организмы попадают в отложения или среду, где сохраняются их формы. Палеоботаники собирают ископаемые образцы в полевых условиях и помещают их в контекст геологических отложений и других окаменелых организмов, окружающих их. Эта деятельность требует большой осторожности, чтобы сохранить целостность хрупких окаменелостей и слоев горных пород, в которых они обнаружены.
Одним из самых захватывающих недавних достижений в палеоботанике является использование аналитической химии и молекулярной биологии для изучения окаменелостей. Для сохранения молекулярных структур необходима среда, свободная от кислорода, так как от его присутствия зависит окисление и деградация материала под действием микроорганизмов. Одним из примеров использования аналитической химии и молекулярной биологии является идентификация олеанана, соединения, отпугивающего вредителей. До этого момента олеанан был присущ только цветковым растениям; однако теперь он был извлечен из отложений, датируемых пермским периодом, намного раньше, чем современные даты появления первых цветковых растений. Палеоботаники также могут изучать ископаемую ДНК, которая может дать большой объем информации, анализируя и сравнивая последовательности ДНК вымерших растений с последовательностями ДНК живых и родственных организмов. Благодаря этому анализу можно построить эволюционные отношения для линий растений.
Палеоботаники также могут изучать ископаемую ДНК, которая может дать большой объем информации, анализируя и сравнивая последовательности ДНК вымерших растений с последовательностями ДНК живых и родственных организмов. Благодаря этому анализу можно построить эволюционные отношения для линий растений.
Некоторые палеоботаники скептически относятся к выводам, сделанным на основе анализа молекулярных окаменелостей. Например, интересующие химические материалы быстро разлагаются при воздействии воздуха во время их первоначального выделения, а также при дальнейших манипуляциях. Всегда существует высокий риск загрязнения образцов посторонними материалами, в основном микроорганизмами. Тем не менее, по мере совершенствования технологий анализ ДНК окаменелых растений предоставит бесценную информацию об эволюции растений и их адаптации к постоянно меняющейся среде.
Основные подразделения наземных растений
Зеленые водоросли и наземные растения сгруппированы в подтип, называемый Streptophytina, и поэтому называются Streptophytes. В дальнейшем наземные растения подразделяются на две основные группы в зависимости от отсутствия или наличия сосудистой ткани, как показано на рис. 5. Растения, у которых отсутствует сосудистая ткань, образованная из специализированных клеток для транспортировки воды и питательных веществ, относятся к относятся к несосудистым растениям . Печеночники, мхи и роголистники — это бессемянные несосудистые растения, которые, вероятно, появились на ранней стадии эволюции наземных растений. Сосудистые растения развили сеть клеток, проводящих воду и растворенные вещества. Первые сосудистые растения появились в конце ордовика и, вероятно, были сходны с плауновидными, к которым относятся плауны (не путать с мхами) и птерофиты (папоротники, хвощи, метелки). Ликофиты и птерофиты относят к бессемянным сосудистым растениям, потому что они не дают семян. Семенные растения, или сперматофиты, составляют самую большую группу всех существующих растений и, следовательно, доминируют в ландшафте. К семенным растениям относятся голосеменные, прежде всего хвойные (голосеменные), дающие «голые семена», и самые успешные из всех растений — цветковые (покрытосеменные).
В дальнейшем наземные растения подразделяются на две основные группы в зависимости от отсутствия или наличия сосудистой ткани, как показано на рис. 5. Растения, у которых отсутствует сосудистая ткань, образованная из специализированных клеток для транспортировки воды и питательных веществ, относятся к относятся к несосудистым растениям . Печеночники, мхи и роголистники — это бессемянные несосудистые растения, которые, вероятно, появились на ранней стадии эволюции наземных растений. Сосудистые растения развили сеть клеток, проводящих воду и растворенные вещества. Первые сосудистые растения появились в конце ордовика и, вероятно, были сходны с плауновидными, к которым относятся плауны (не путать с мхами) и птерофиты (папоротники, хвощи, метелки). Ликофиты и птерофиты относят к бессемянным сосудистым растениям, потому что они не дают семян. Семенные растения, или сперматофиты, составляют самую большую группу всех существующих растений и, следовательно, доминируют в ландшафте. К семенным растениям относятся голосеменные, прежде всего хвойные (голосеменные), дающие «голые семена», и самые успешные из всех растений — цветковые (покрытосеменные).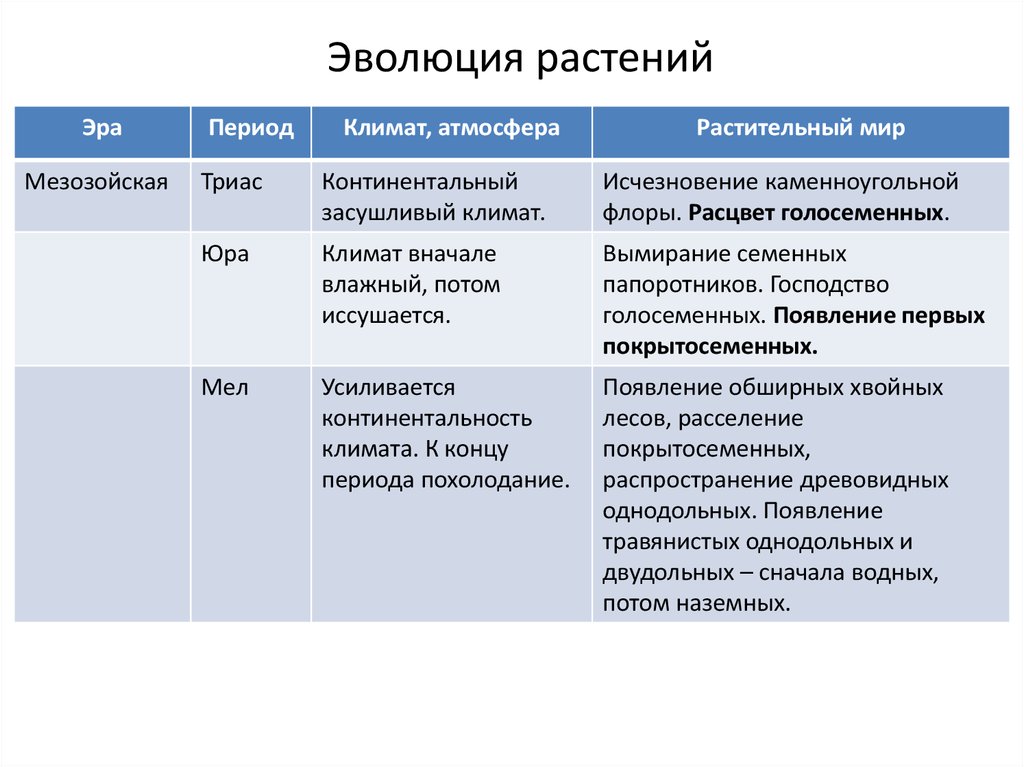 Покрытосеменные растения защищают свои семена внутри камер в центре цветка; стенки камеры позже развиваются в плод.
Покрытосеменные растения защищают свои семена внутри камер в центре цветка; стенки камеры позже развиваются в плод.
Рисунок 5. В этой таблице показаны основные подразделения зеленых растений.
Практический вопрос
Какое из следующих утверждений о подразделениях растений неверно?
- Ликофиты и птерофиты представляют собой бессемянные сосудистые растения.
- Все сосудистые растения дают семена.
- Все несосудистые эмбриофиты являются мохообразными.
- Семенные растения включают покрытосеменные и голосеменные растения.
Показать ответ
Вкратце: ранняя жизнь растений
Наземные растения приобрели черты, позволившие колонизировать сушу и выживать вне воды. Все наземные растения имеют следующие характеристики: чередование поколений, при этом гаплоидное растение называется гаметофитом, а диплоидное растение называется спорофитом; защита зародыша, образование гаплоидных спор в спорангии, образование гамет в гаметангии и апикальная меристема.