
Содержание
Покрытосеменные. Двудольные и однодольные покрытосеменные
Биология. 6 класс. Костиков
Вы узнаете, что такое покрытосеменные, что такое двудольные и однодольные покрытосеменные.
Из предыдущей темы вы узнали об основных особенностях строения, жизнедеятельности и размножения цветковых растений. Все они называются цветковыми или покрытосеменными. Это значит, что их семезачатки глубоко скрыты в полости завязи, а пыльцу во время опыления улавливает рыльце пестика. Они имеют зародышевый мешок, им присуще двойное оплодотворение. Покрытосеменные распространены на всех континентах, даже в Антарктиде, произрастают во всех климатических зонах. Их насчитывается около 250 тыс. самых разнообразных видов. Эти растения заселяют наиболее засушливые пустыни и умеренно увлажненные территории, встречаются на болотах и в пресных водоёмах, а некоторые (например, морская трава) даже приспособились к жизни на морском дне, при этом морская вода переносит их пыльцевые зёрна.
Покрытосеменные — это семенные растения, у которых пыльцу улавливает рыльце пестика.
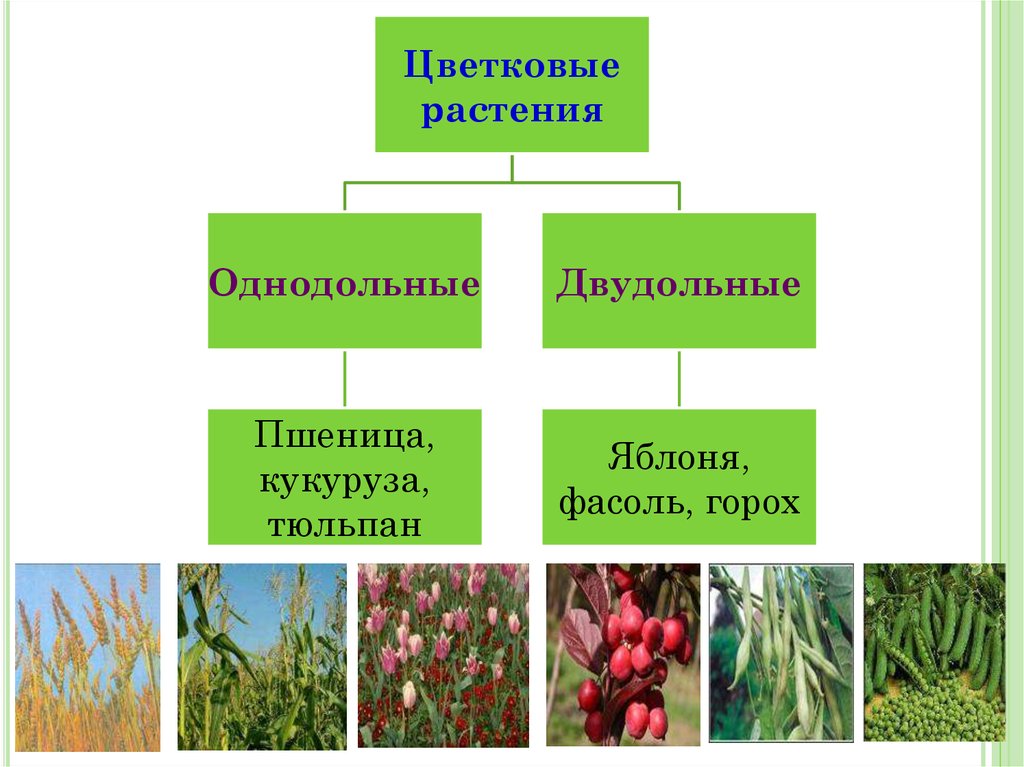
У цветковых растений имеются семена с зародышами, содержащие две или одну семядоли. С этим признаком связан целый ряд других особенностей. Поэтому покрытосеменные делятся на двудольные и однодольные.
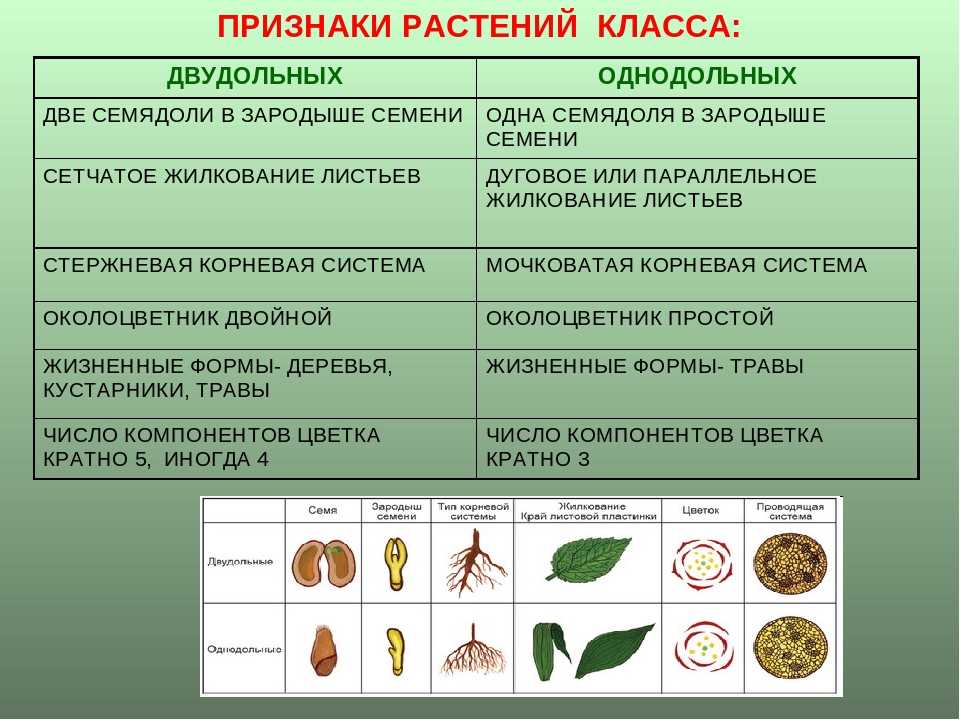
Корень. У большинства двудольных главный корень живёт продолжительное время, а поэтому их корневая система имеет один мощный корень, от которого отходят менее мощные боковые корни. Главный корень однодольных рано отмирает, поэтому их корневая система обычно состоит из многочисленных одинаково развитых корней.
Лист. У листьев двудольных обычно небольшое основание листа, часто есть прилистники, хорошо развит черенок и листовая пластинка с пальчатым или перистым жилкованием. Листья однодольных часто имеют основу в виде влагалища, окружающего стебель, в подавляющем большинстве без прилистников и черенка, с овальной или лентовидной листовой пластинкой, дугообразным или параллельным жилкованием.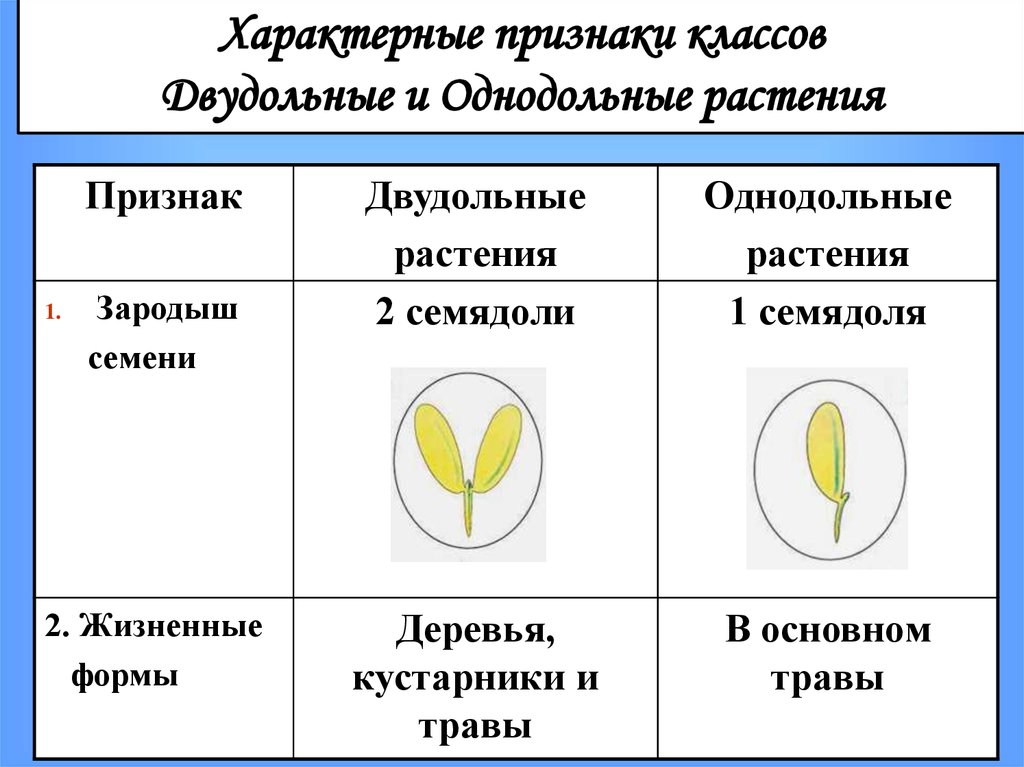
Стебель. На поперечном срезе стебля двудольных проводящие пучки расположены по кольцу, есть сердцевина. Для них типично образование камбия и мощное утолщение. Поэтому среди двудольных встречаются как травы, так и деревья. Проводящие пучки однодольных распределены почти по всему поперечному срезу стебля. Иногда в центре можно распознать сердцевину, но у некоторых однодольных, например злаков (бамбук, пшеница, рожь) на месте сердцевины возникает обширная воздушная полость. Камбий у однодольных не образуется. Это — преимущественно травы. Деревьев, принадлежащих к этой группе немного: это — пальмы, алоэ или драцены.
Цветок. Для многих двудольных характерны цветки, у которых части размещены кругами, в каждом из которых по четыре или пять листиков околоцветника, тычинок или плодолистиков, из которых состоит пестик. А у типичного цветка однодольных круги трёхчленные. Цветки с большим и неопределённым числом частей встречаются как среди двудольных, так и среди однодольных.
Двудольных покрытосеменных насчитывается на Земле примерно 180 тыс. видов, а около 70 тыс. видов покрытосеменных имеют зародыш с одной семядолей и относятся к однодольным.
ВЫВОДЫ
- 1. Покрытосеменные — наиболее обширная группа растений, распространённая в самых разнообразных условиях произрастания.
- 2. Характерными признаками покрытосеменных есть цветок, плод, улавливание пыльцы рыльцем пестика, зародышевый мешок, двойное оплодотворение, ситовидные трубки.
- 3. Покрытосеменные растения по совокупности признаков делятся на двудольные и однодольные.
ТЕРМИНЫ И ПОНЯТИЯ, КОТОРЫЕ ВАЖНО ЗНАТЬ
Покрытосеменные, двудольные, однодольные.
КОНТРОЛЬНЫЕ ВОПРОСЫ
- 1. Каковы основные признаки покрытосеменных?
- 2. По каким признакам покрытосеменные делятся на двудольные и однодольные?
- 3. Сколько видов покрытосеменных, двудольных и однодольных существует в природе?
ЗАДАНИЯ
Сравните признаки двудольных и однодольных покрытосеменных, сведения об их особенностях занесите в таблицу (в тетради).
Признак | Двудольные | Однодольные |
Количество семядолей у зародыша семени | ||
Жизненные формы (деревья или травы) | ||
Внутреннее строение стебля | ||
Жилкование листа | ||
Тип корневой системы | ||
Количество частей цветка |
ГДЗ к учебнику можно найти тут.
Попередня
Сторінка
Наступна
Сторінка
Зміст
Цей контент створено завдяки Міністерству освіти і науки України
27.
 Систематика покрытосеменных. Однодольные и двудольные растения.
Систематика покрытосеменных. Однодольные и двудольные растения.
Классы
Двудольные и Однодольные делятся на
семейства. Растения каждого из семейств
имеют общие признаки. У цветковых
растений основными признаками являются
строение цветка и плода, тип соцветия,
а также особенности внешнего и внутреннего
строения вегетативных органов.
Класс
Двудольные включает
418 семейств, около 10 тыс. родов и свыше
190 тыс. видов, что составляет примерно
3/4 видов цветковых растений. К этому
классу принадлежат важнейшие
плодово-ягодные (яблоня, груша, вишня,
виноград, цитрусовые), кормовые (турнепс,
брюква, клевер, люцерна), прядильные
(лен, хлоп-чатник, конопля), масличные
(подсолнечник, клещевина), декоративные
(розы, хризантемы, астры, георгины) и др.
Все они принадлежат к одному или к разным
семействам и отличаются комплексом
признаков (табл. 9.2).
Класс
Однодольные образует
около 122 семейств, свыше 3 тыс. родов и
около 63 тыс. видов. Однодольные обеспечивают
видов. Однодольные обеспечивают
человека хлебом. К ним относятся хлебные
злаки (пшеница, рис, рожь, ячмень, кукуруза,
просо и др.), некоторые овоидные (лук,
чеснок, спаржа), декоративные (лилии,
тюльпаны, гиацинты, аспарагус, орхидеи)
растения.
Основную
массу многих сенокосов и пастбищ в
условиях Беларуси и России составляют
дикорастущие злаки — тимофеевка,
лисохвост, полевица, мятлик
и
др.
Семейство
бобовых представляют
около 12 000 видов деревьев, кустарников
и трав, среди которых много лиан. Листья
очередные с прилистниками, перисто- или
пальчатосложные, реже простые. Цветки
обоеполые, собраны в соцветия (головка
у клевера, кисть у люпина) или одиночные
(у гороха).
Чашечка
зубчатая, иногда двугубая. Венчик
пятилепестный «мотыльковый» (верхний
лепесток — парус, боковые — несла, два
нижних срослись в лодочку).
В
цветке 10 тычинок, из которых девять
срослись тычиночными нитями, а верхняя
(десятая) остается свободной. Пестик
Пестик
один. Плод — боб. Для бобовых характерно
взаимовыгодное сожительство двух разных
организмов симбиоз.На корнях бобовых
растений образуются клубеньки, в клетках
которых живут клубеньковые бактерии.
Они усваивают азот из воздуха, поэтому
все бобовые богаты белком. Среди бобовых
много продовольственных растений
(фасоль, горох, соя, арахис и др.), кормовых
(люцерна, клевер, эспарцет), декоративных
(люпин, душистый горошек, глициния, белая
акация и др.).
Представлено
главным образом травами, встречаются
полукустарники и кустарники.Для растений
семейства капустовых характерно
очередное листорасположение, цветки
правильные: 4 свободных чашелистика и
столько же лепестков. Чашелистики и
лепестки расположены крест-накрест,
отсюда и название — крестоцветные
Тычинок
6, из них 2 наружные» более короткие,
пестик состоит из 2-х плодолистиков.
Плод — стручок, стручочек, или односемянный
орешек. К семейству крестоцветные
относится хозяйственно важный вид
капусты — двулетнее растение,
представленное несколькими разновидностями
(кольраби, брюссельская, лиственная,
цветная, краснокочанная и др. ). Наибольшее
). Наибольшее
распространение получила капуста
кочанная. Ее возделывают во всех зонах.
К роду капусты относятся также брюква
и репа.
К вопросу о гомологии семядолей покрытосеменных
Бартон, М.К. и Р.С. Поэтиг . 1993. Формирование апикальной меристемы побега у Arabidopsis thaliana : анализ развития у дикого типа и у мутанта без побегов. Развитие 119: 823–831.
Google Scholar
Бхаратан, Г. и Э. Циммер . 1995. Ранние события ветвления однодольных — частичный анализ последовательности рибосомной ДНК 18s. Стр. 81–107 в P. J. Rudall et al. (ред.), Однодольные: систематика и эволюция. Том. 1. Королевские ботанические сады, Кью.
Google Scholar
Burger, W. 1977 [1978]. Piperales и однодольные: альтернативные гипотезы происхождения однодольных цветов. Бот. Откр. (Ланкастер) 43: 345–39.3.
Бот. Откр. (Ланкастер) 43: 345–39.3.
Артикул
Google Scholar
—. 1981. Возрождение ереси: однодольная теория происхождения покрытосеменных. Эвол. Теория 5: 189–225.
Google Scholar
—. 1996. Гомологичны ли тычинки и плодолистики? Стр. 111–117 в У. Д’Арси и Р. Китинг (ред.), Пыльник: форма, функция и фитогенез. Издательство Кембриджского университета, Нью-Йорк.
Google Scholar
Campbell, DH 1930. Филогения однодольных растений. Энн. Бот. 44: 311–331.
Google Scholar
Carlquist, S. 1981. Исследования Stylidiaceae: однодольные в семействе; номенклатурное изменение. Алисо 10: 35–38.
Google Scholar
Chase, M. W., D.E. Soltis, R.G. Olmstead, D. Morgan, D.H. Les, B.D. Mishler, M.R. Duvall, R.A. Price, H.G. Hills, Y.-L. Куи, К. А. Крон, Дж. Х. Реттиг, Э. Конти, Дж. Д. Палмер, Дж. Р. Манхарт, К. Дж. Сытсма, Х. Дж. Майклс, В. Дж. Кресс, К. Г. Кароль, В. Д. Кларк, М. Хедрен, Б. С. Гаут, Р. К. Янсен, Х.-Дж. Ким, С. Ф. Вимпи, Дж. Ф. Смит, Г. Р. Фурнье, С. Х. Штраус, Q.-Y. Xiang, G.M. Plunkctt, P.S. Soltis, S. Swensen, S.E. Williams, P.A. Gadek, C.J. Quinn, L.E. Eguiarte, E. Golenberg, G.H. Learn , Jr., SW Graham, SCH Barrett, S. Dayanandan и VA Albert . 1993. Филогенез семенных растений: анализ нуклеотидных последовательностей пластидного гена rbc L. Ann. Миссури Бот. Гард. 89: 528–580.
W., D.E. Soltis, R.G. Olmstead, D. Morgan, D.H. Les, B.D. Mishler, M.R. Duvall, R.A. Price, H.G. Hills, Y.-L. Куи, К. А. Крон, Дж. Х. Реттиг, Э. Конти, Дж. Д. Палмер, Дж. Р. Манхарт, К. Дж. Сытсма, Х. Дж. Майклс, В. Дж. Кресс, К. Г. Кароль, В. Д. Кларк, М. Хедрен, Б. С. Гаут, Р. К. Янсен, Х.-Дж. Ким, С. Ф. Вимпи, Дж. Ф. Смит, Г. Р. Фурнье, С. Х. Штраус, Q.-Y. Xiang, G.M. Plunkctt, P.S. Soltis, S. Swensen, S.E. Williams, P.A. Gadek, C.J. Quinn, L.E. Eguiarte, E. Golenberg, G.H. Learn , Jr., SW Graham, SCH Barrett, S. Dayanandan и VA Albert . 1993. Филогенез семенных растений: анализ нуклеотидных последовательностей пластидного гена rbc L. Ann. Миссури Бот. Гард. 89: 528–580.
Артикул
Google Scholar
Чейз, М. В., Д. В. Стивенсон, П. Уилкинс и П. Дж. Рудалл . 1995. Систематика однодольных растений: комбинированный анализ. Стр. 685–730 в PJ Rudall et al. (ред.), Однодольные: систематика и эволюция, Том 2. Королевские ботанические сады, Кью.
(ред.), Однодольные: систематика и эволюция, Том 2. Королевские ботанические сады, Кью.
Google Scholar
Christiansen, ML 1986. Карта судьбы верхушки организующего побега в Gossypium . амер. Дж. Бот. 73: 947–958.
Артикул
Google Scholar
Cook, MT 1906. Эмбриология некоторых кубинских Nymphaeaceae. Бот. Газ. 42:376–392.
Артикул
Google Scholar
Coulter, JM и WJG Land . 1914. Происхождение однодольных. Бот. Газ. 57: 509–519.
Артикул
Google Scholar
Кронквист, А. 1981. Комплексная система классификации цветковых растений. Издательство Колумбийского университета, Нью-Йорк.
Google Scholar
Дальгрен Р. и Ф. Н. Расмуссен . 1983. Эволюция однодольных: признаки и филогенетическая оценка. Эвол. биол. 16: 255–395.
Google Scholar
Davis, J.I. 1995. Филогенетическая структура однодольных, как следует из вариаций сайта рестрикции ДНК хлоропластов, и сравнение мер поддержки клады. Сист. Бот. 10: 503–527.
Артикул
Google Scholar
Delendick, T.K. 1990. Обзор лиственных флавоноидов Aceraceae. Мем. Нью-йоркский бот. Гард. 54: 1–29.
Google Scholar
Донохью, М. Дж. и М. Дж. Сандерсон . 1994. Комплексность и гомология растений. Стр. 394–421 в Б. К. Холл (ред.), Гомология: иерархическая основа сравнительной биологии. Академик Пресс, Нью-Йорк.
К. Холл (ред.), Гомология: иерархическая основа сравнительной биологии. Академик Пресс, Нью-Йорк.
Google Scholar
Дубе, В.П., Д.К. Авастхи и В.П. Сингхал . 1981. Сравнительные анатомические наблюдения над двудольными и трехдольными сеянцами Raphanus sativus L. (Brassicaceae). Акта Бот. Индика 9: 134–137.
Google Scholar
Дюваль, М. Р., М. Т. Клегг, М. В. Чейз, В. Д. Кларк, В. Дж. Кресс, Х. Г. Хиллз . LE Eguiarte, JF Smith, BS Gaut, EA Zimmer & GH Learn Jr. 1993. Филогенетические гипотезы для однодольных, построенные на основе данных rbcL. Энн. Миссури Бот. Гард. 80: 607–619.
Артикул
Google Scholar
Eames, AJ 1961. Морфология покрытосеменных растений. Макгроу-Хилл, Нью-Йорк.
Макгроу-Хилл, Нью-Йорк.
Google Scholar
Гиффорд, Э. М. и А. С. Фостер . 1988. Морфология и эволюция сосудистых растений. Эд. 3. У. Х. Фримен, Нью-Йорк.
Google Scholar
Гольдберг, Р. Б., Г. де Пайва и Р. Ядегари . 1994. Эмбриогенез растений: от зиготы до семени. Наука 266: 605–614.
Артикул
пабмед
КАС
Google Scholar
Гупта, М. Л. и В. К. Джайн . 1980. Сравнительная анатомия нормальных и трехдольных проростков Brassica oleracea 9.0009 вар. на душу населения . Курс. науч. 49: 277–278.
Google Scholar
Гуттенберг, Х. и Р. Мюллер-Шредер . 1958. Untersuchungen Ober die Entwicklung des Embryos und der Keimpflanze von Nuphar luteum Smith. Планта 51: 481–510.
1958. Untersuchungen Ober die Entwicklung des Embryos und der Keimpflanze von Nuphar luteum Smith. Планта 51: 481–510.
Артикул
Google Scholar
Haccius, B. 1954. Embryologische und histogenetische Studien an «monokotylen Dikotylen». I. Claytonia virginica . Эстерр. Бот. Т. 106: 373–389.
Артикул
Google Scholar
— & К. К. Лакшаманан . 1967. Экспериментальные исследования однодольных двудольных: индуцированные фенилборной кислотой «двудольные» зародыши у Cyclamen persicum . Фитоморфология 17: 488–494.
КАС
Google Scholar
— & В. Дж. Филип . 1979. Развитие эмбриона у Cocos nucifera L.: важный вклад в общее понимание эмбриогенеза пальмы. пл. Сист. Эвол. 132: 91–106.
Артикул
Google Scholar
Хейнс, Р.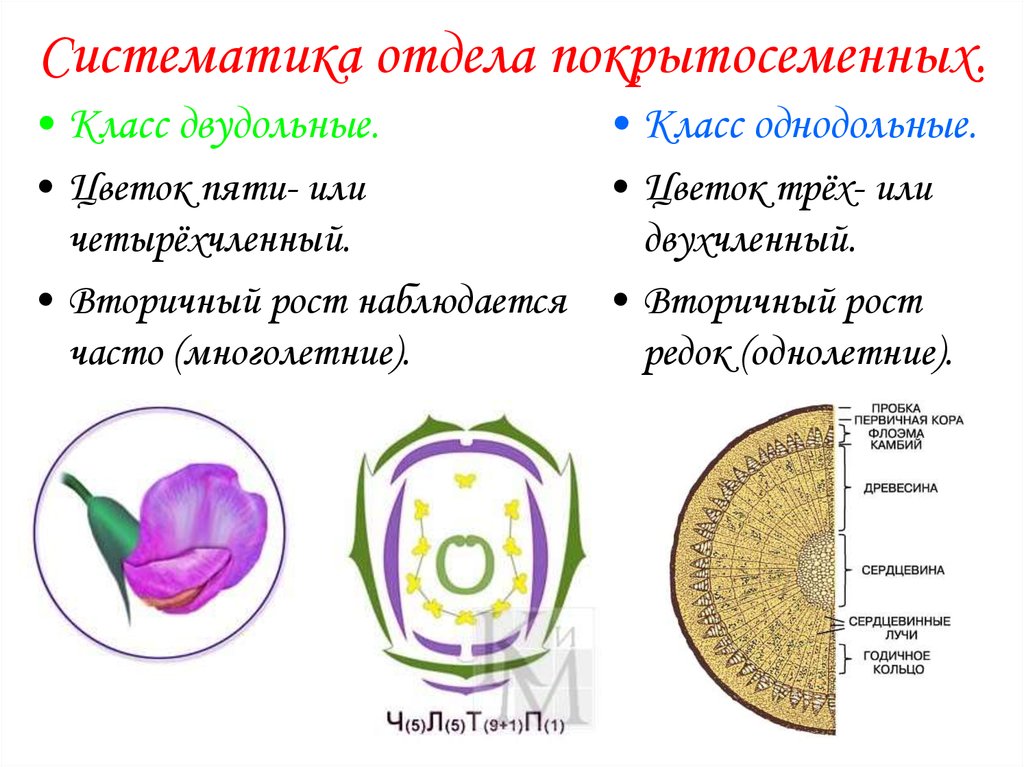 В. и К. А. Лай . 1975. Проростки кувшинчатых. Дж. Линн. соц., бот. 70: 255–265.
В. и К. А. Лай . 1975. Проростки кувшинчатых. Дж. Линн. соц., бот. 70: 255–265.
Артикул
Google Scholar
——. 1979. Однодольные сеянцы: обзор доказательств, подтверждающих происхождение от слияния. Дж. Линн. соц., бот. 78: 123–140.
Артикул
Google Scholar
Haszpruner, G. 1992. Типы гомологии и их значение для эволюционной биологии и филогенетики. Дж. Эвол. биол. 5:13–24.
Артикул
Google Scholar
Жак-Феликс, Х. 1982a. Les Monocotylédones n’ont pas de cotylédon. Бык. Мус. История Нац. (Париж), 4е сер., 4, секция. Б, Адансония 1–2: 3–40.
Google Scholar
—. 1982б. Embryologie vegétal — les idees meconnues Анри Байона о структуре эмбриона однодольных. Комп. Ренд. хебд. Сеансы акад. науч. сер. III: 255–257.
Комп. Ренд. хебд. Сеансы акад. науч. сер. III: 255–257.
Google Scholar
—. 1988. Les Liliopsida (ex Monocotyledones) n’ont pas de cotylédon. II. La préfeuille de la plantule: ses rapports avec celles des axes feuillés. Бык. Мус. История Нац. (Париж), 4е сер. 10, секция. Б, Адансония 3:275–333.
Google Scholar
Juguet, M. 1973. Expression précoce de la однодольные и др. место на месте вегетативного тигра среди однодольных, avec quelques remarques sur les types de symétrie des Monocotylédones et de Dicotylédones. Бык. соц. Бот. Франция, мем. 23: 337–354.
Google Scholar
Kaplan, DR 1984. Концепция гомологии и ее центральная роль в выяснении систематических взаимоотношений растений. Стр. 51–70 в Т. Дункан и Т. Ф. Стюесси (ред.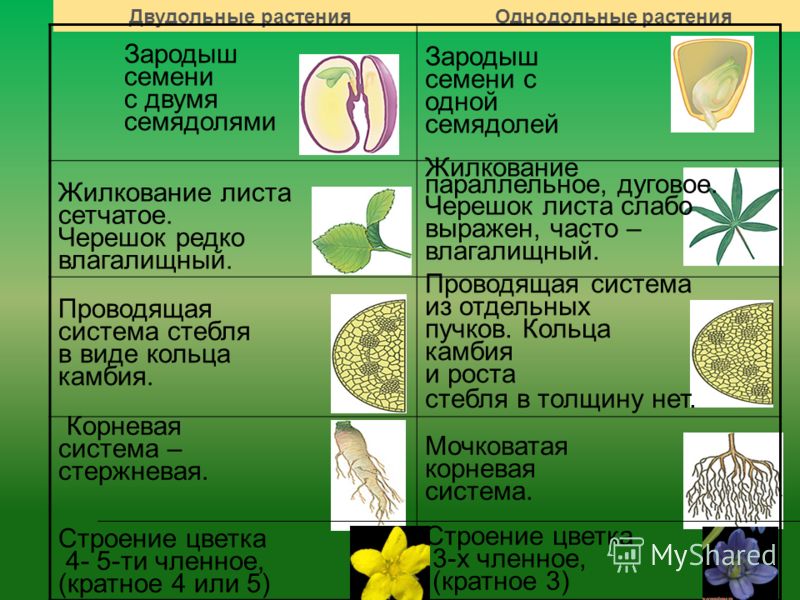 ), Кладистика: перспективы реконструкции эволюционной истории. Издательство Колумбийского университета, Нью-Йорк.
), Кладистика: перспективы реконструкции эволюционной истории. Издательство Колумбийского университета, Нью-Йорк.
Google Scholar
Kraus, F. 1988. Эмпирическая оценка использования критерия поляризации онтогенеза в филогенетическом выводе. Сист. Зоол. 37: 106–141.
Артикул
Google Scholar
Кудраишов Л.В. 1964. Происхождение однодольных растений (на примере Helobiae). Бот. Журн. (Москва и Ленинград) 49: 473–486.
Google Scholar
Lakshmanan, KK 1978. Исследования по развитию Commelina benghalensis III. Семядоли. Фитоморфология 28: 253–261.
Google Scholar
Лаукс, Т. и Г. Юргенс . 1994. Создание плана кузова Arabidopsis 9.Эмбрион 0009. Акта Бот. Нерл. 43: 247–260.
1994. Создание плана кузова Arabidopsis 9.Эмбрион 0009. Акта Бот. Нерл. 43: 247–260.
Google Scholar
Lawton, JRS и JT Lawton . 1967. Морфология покоящегося зародыша и молодых проростков пяти видов Dioscorea из Нигерии Proc. Линн. соц. Лондон 178: 153–159.
Google Scholar
Лес, Д. Х. и Э. Л. Шнайдер . 1995. Nymphaeales, Alismatidae и теория происхождения водных однодольных растений. Стр. 23–42 в P. J. Rudall et al. (ред.), Однодольные: систематика и эволюция. Том. 1. Королевские ботанические сады, Кью.
Google Scholar
Локонте, Х. и Д. В. Стивенсон . 1990. Кладистика семенных растений. Бриттония 42: 197–211.
Артикул
Google Scholar
——. 1991. Кладистика Magnoliidae. Кластика 7: 267–296.
1991. Кладистика Magnoliidae. Кластика 7: 267–296.
Артикул
Google Scholar
Lyon, HL 1901. Наблюдения за эмбриогенезом Nelumbo . Миннесота Бот. Стад. 2: 643–656.
Google Scholar
Meinke, D.W. 1995. Молекулярная генетика эмбриогенеза растений. Годовой оборот Pl. Физиол. молек. биол. 46: 369–394.
Артикул
КАС
Google Scholar
Местре, Ж.-Ч. и М. Гедес . 1983. Природа и филогенетическое значение семядолей. Бык. соц. Бот. Франция, факт. Бот. 130, 3/4: 7–22.
Google Scholar
Мозли, М.Ф., Э.Л. Шнайдер и П.С. Уильямсон . 1993. Филогенетические интерпретации отдельных цветочных сосудистых признаков у Nymphaeaceae sensu lato.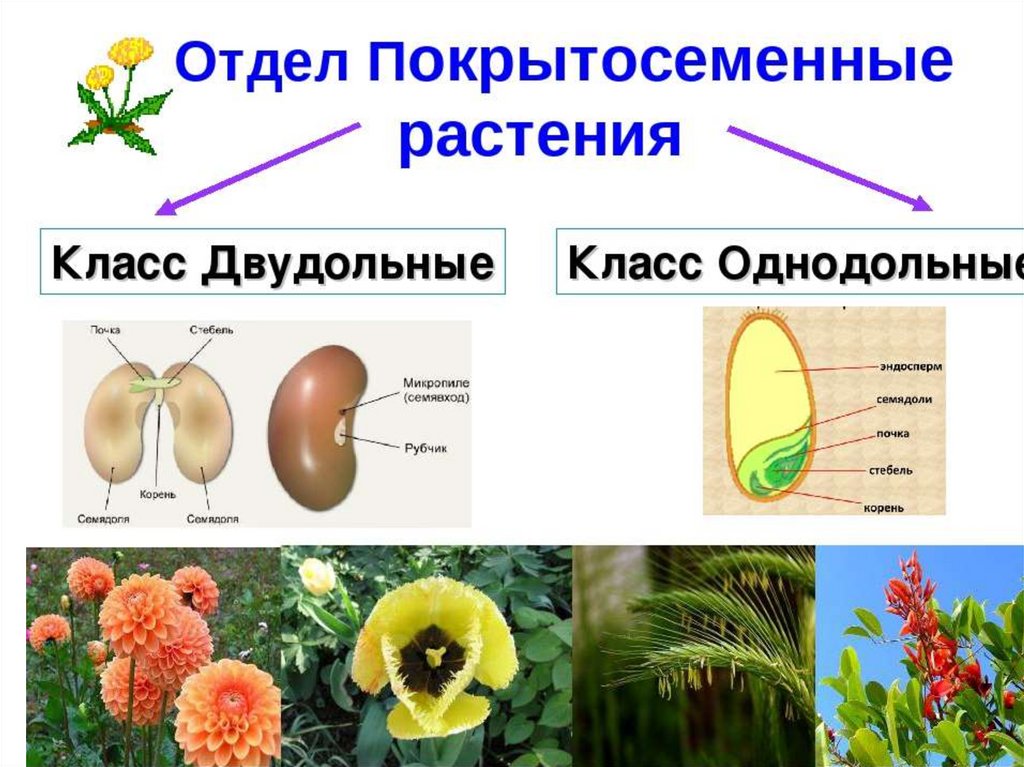 Аква. Бот 44: 325–342.
Аква. Бот 44: 325–342.
Артикул
Google Scholar
Натеш, С. и М. А. Рау . 1984. Эмбрион. Стр. 377–443 в Б. М. Джон (ред.), Эмбриология покрытосеменных растений. Springer Verlag, Берлин.
Google Scholar
Нельсон Г. 1994. Гомология и систематика. Стр. 101–149 в Б. К. Холл (ред.), Гомология: иерархическая основа сравнительной биологии. Академик Пресс, Нью-Йорк.
Google Scholar
Nickrcnt, DL и DE Soltis . 1995. Сравнение фитогенеза покрытосеменных ядерных последовательностей 18S рДНК и rbc L. Энн. Миссури Бот. Гард. 82: 208–234.
Артикул
Google Scholar
Nitzschke, J.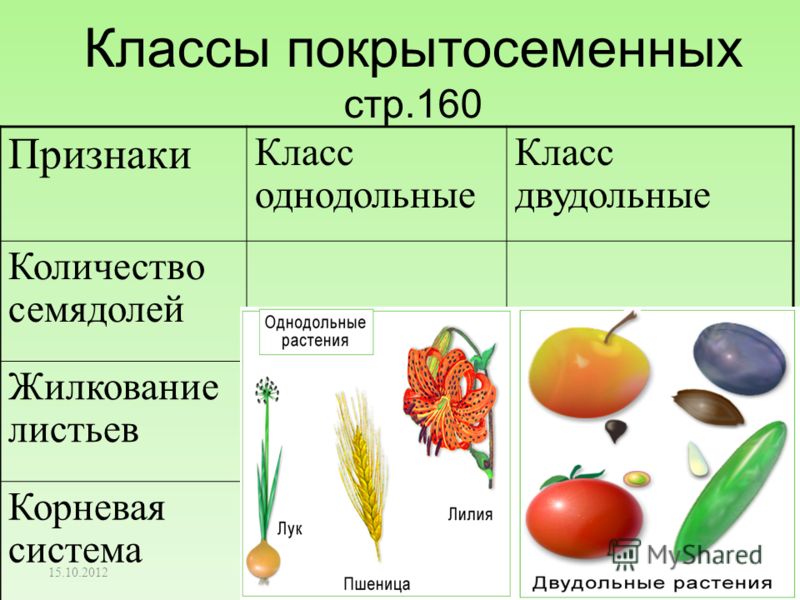 1914. Beiträg zur Phylogenie der Monokotyln, gegrundet auf der Embryosackentwicklung apokarper Nymphaeaceen und Helobien. Бейтр. биол. Пфланцен 12: 223–268.
1914. Beiträg zur Phylogenie der Monokotyln, gegrundet auf der Embryosackentwicklung apokarper Nymphaeaceen und Helobien. Бейтр. биол. Пфланцен 12: 223–268.
Google Scholar
Palser, B. 1975. Основы филогении покрытосеменных растений: эмбриология. Энн. Миссури Бот. Гард. 62: 621–646.
Артикул
Google Scholar
Patterson, C. 1982. Морфологические признаки и гомология. Стр. 21–74 в К. А. Джойси и А. Э. Фрайдей (ред.), Проблемы филогенетической реконструкции. Ассоциация систематики, специальный том. 21. Академик Пресс, Лондон.
Google Scholar
Пиллаи, А. и С. К. Гоял . 1983. Анатомия развития некоторых масличных растений. IV. Нормальные и трехдольные сеянцы Sesamum indicum L.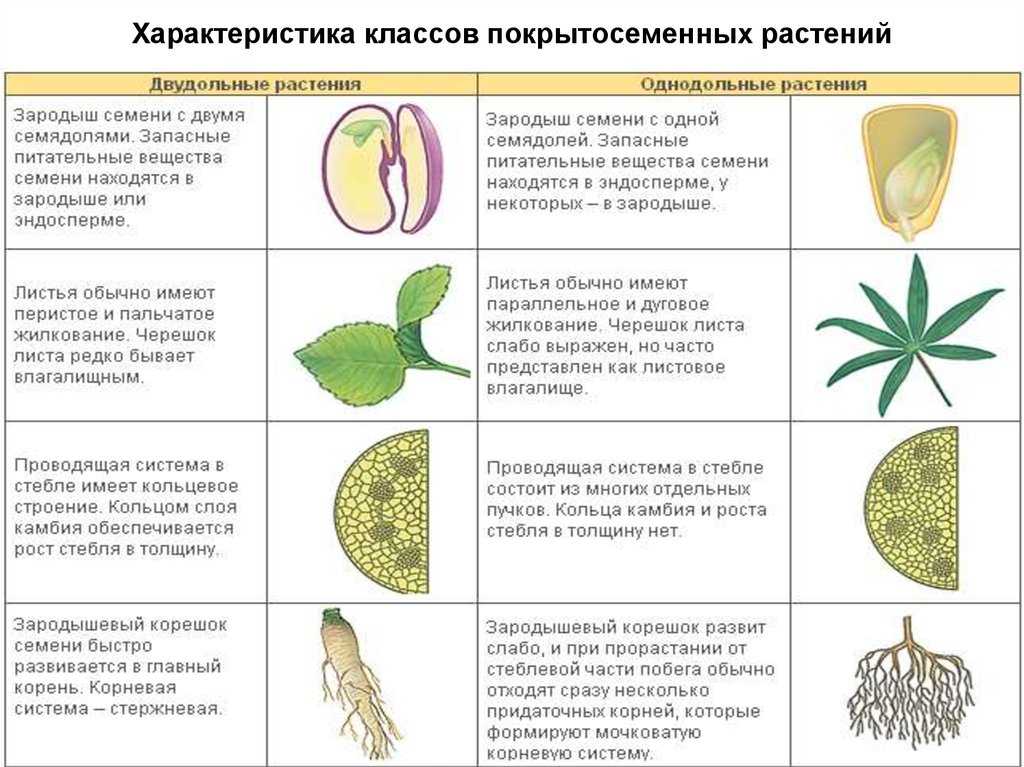 Feddes Repert. 94: 87–90.
Feddes Repert. 94: 87–90.
Google Scholar
Raff, R. 1996. Форма жизни: гены, развитие и эволюция форм животных. Издательство Чикагского университета, Чикаго.
Google Scholar
Raju, MVS 1990. Соцветия и семена дикого овса — анатомия, развитие и морфология. Канадский центр равнин, Реджайна.
Google Scholar
— & Э. М. В. Намбудири . 1995. Зародыш эуспорангиатных сосудистых растений и возможная древность зародыша однодольных. Фитоморфология 45:191–206.
Google Scholar
Rangaswamy, NS 1967. Морфогенез прорастания семян у покрытосеменных растений. Фитоморфология 17: 477–487.
Google Scholar
Riedl, R. 1978. Порядок в живых организмах. RPS Jefferies, пер. Джон Уайли, Нью-Йорк. [Первоначально опубликовано как Die Ordnung des Lebendigen. 1975. Пол Пари, Гамбург.]
1978. Порядок в живых организмах. RPS Jefferies, пер. Джон Уайли, Нью-Йорк. [Первоначально опубликовано как Die Ordnung des Lebendigen. 1975. Пол Пари, Гамбург.]
Google Scholar
Rieppel, O. 1990. Онтогенез — путь вперед для систематики, путь назад для филогенеза. биол. Дж. Линн. соц. 39:177–191.
Артикул
Google Scholar
—. 1993. Концептуальные отношения онтогенеза, филогенеза и классификации: тактический подход. Эвол. биол. 27:1–32.
Google Scholar
Ryberg, M. 1959. Морфологическое исследование Corydalis nobilis, C. cava, C. solida и некоторых родственных видов с особым упором на их подземные органы. Акта Хорти Берг. 19: 15–119.
Google Scholar
Sargent, E.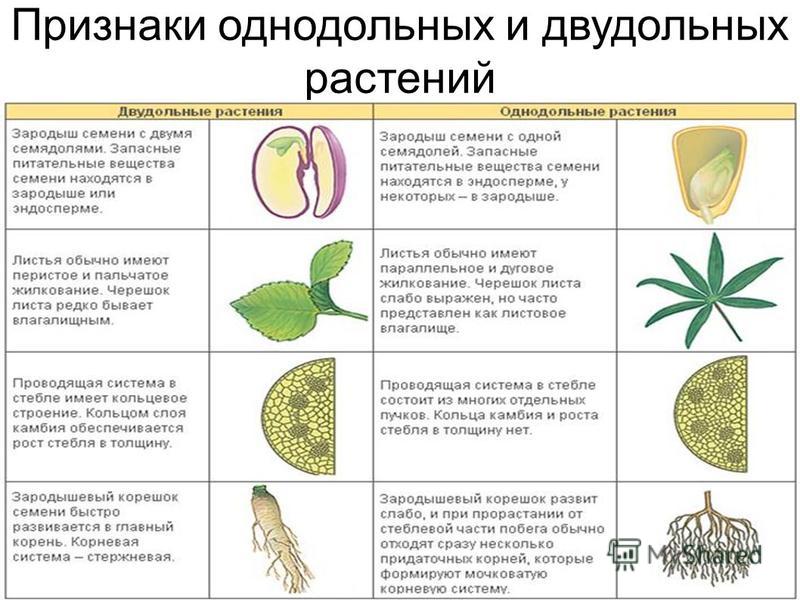 1903. Теория происхождения однодольных, основанная на строении их проростков. Энн. Бот. 17: 1–92.
1903. Теория происхождения однодольных, основанная на строении их проростков. Энн. Бот. 17: 1–92.
Google Scholar
Sattler, R. 1994. Гомология, гомеозис и морфология процессов у растений. Стр. 423–475 в BK Hall (ред.), Homology. Иерархическая основа сравнительной биологии. Академик Пресс, Нью-Йорк.
Google Scholar
— & Р. Рутисхаузер . 1997. Фундаментальное значение морфологии и морфогенеза для исследований растений. Энн. Бот. 80: 571–582.
Артикул
Google Scholar
Schaffner, JH 1904. Некоторые морфологические особенности Nymphaeaceae и Helobiae. Натуралист из Огайо 4: 83–92.
Google Scholar
—. 1934. Филогенетическая систематика растений.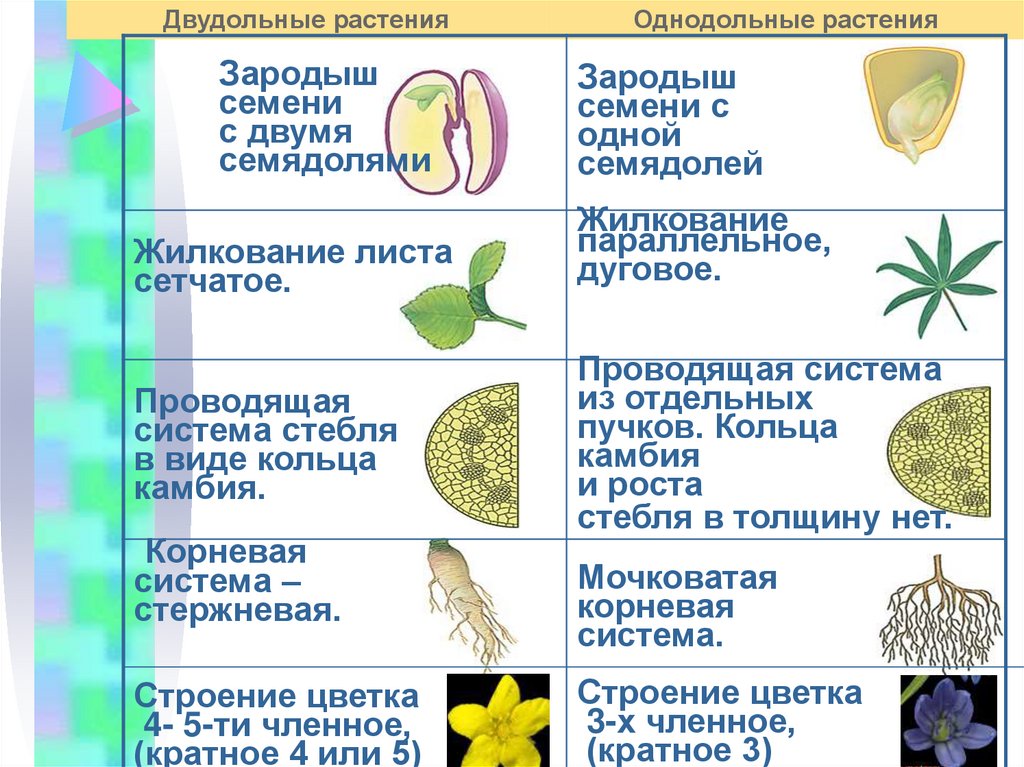 кв. преподобный биол. 9: 129–160.
кв. преподобный биол. 9: 129–160.
Артикул
Google Scholar
Schneider, EL 1978. Морфологические исследования Nymphaeaceae. IX. Семена Barclaya longifolia Wall. Бот. Газ. 139: 223–230.
Артикул
Google Scholar
Шубин Н.Х. 1994. Филогенез развития и происхождение гомологии. Стр. 201–225 в Л. Гранде и О. Риппель (ред.), Интерпретация иерархии природы. От систематических закономерностей к теориям эволюционного процесса. Академик Пресс, Нью-Йорк.
Google Scholar
Сингх, Х. 1978. Эмбриология голосеменных растений. Handbuch der Pflanzenanatomie. Том. 10, пт. 2: 1–302. Гебрюдер Борнтрегер, Берлин.
Google Scholar
Smith, DL 1964.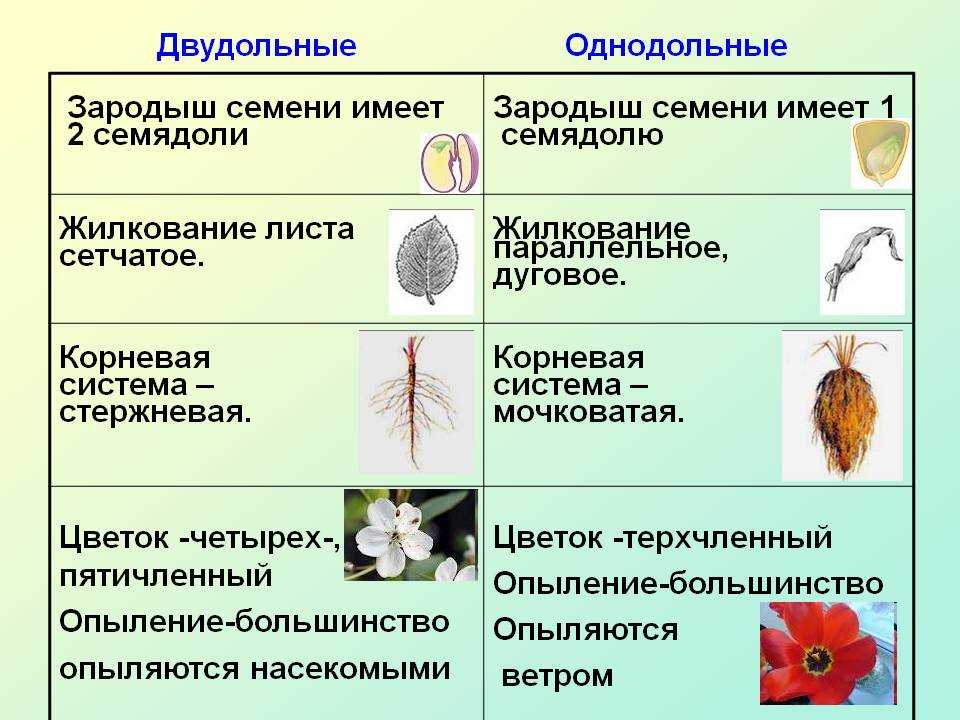 Эволюция семязачатка. биол. Преподобный Кембридж Филос. соц. 39: 137–159.
Эволюция семязачатка. биол. Преподобный Кембридж Филос. соц. 39: 137–159.
Артикул
Google Scholar
Solms-Laubach, H. 1878. Über monocotyle Embryonen mit Scheitelbürtigen Vegetationspunkt. Бот. Zeitung (Берлин) 36: 65–74, 81–93.
Google Scholar
Стеббинс, Г. Л. 1974. Цветковые растения: эволюция выше видового уровня. Издательство Гарвардского университета, Кембридж.
Google Scholar
Stevens, PF 1984. Гомология и филогения; Морфология и систематика. Сист. Бот. 9: 395–409.
Артикул
Google Scholar
Стивенсон, Д. В. и Х. Локонте . 1995. Кладистический анализ семейств однодольных.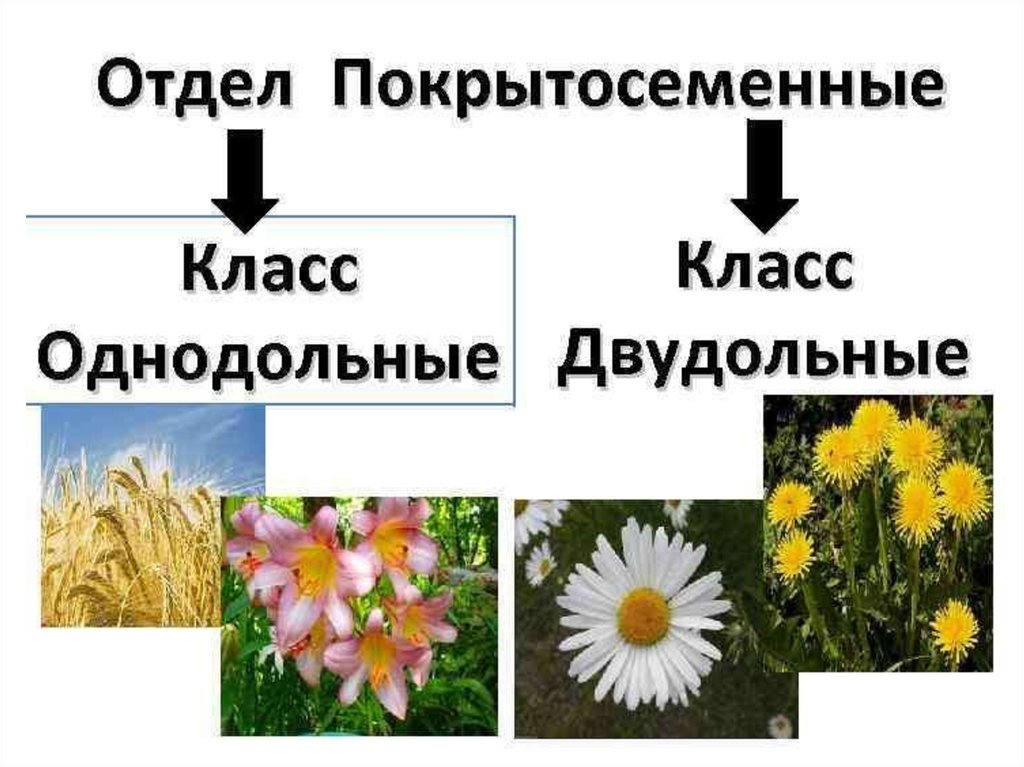 Стр. 543–576 в P. J. Rudall et al. (ред.), Однодольные: систематика и эволюция. Том. 2. Королевские ботанические сады, Кью.
Стр. 543–576 в P. J. Rudall et al. (ред.), Однодольные: систематика и эволюция. Том. 2. Королевские ботанические сады, Кью.
Google Scholar
Swamy, BGL 1949. Дальнейший вклад в морфологию Degeneriaceae. Дж. Арнольд Арбор. 30: 10–38.
Google Scholar
Сыцма, К. Дж. и Д. А. Баум . 1996. Молекулярная фитогенез и разнообразие покрытосеменных растений. Стр. 314–340 в Д. В. Тейлор и Л. Дж. Хики (ред.), Происхождение, эволюция и филогения цветковых растений. Чепмен и Холл, Нью-Йорк.
Глава
Google Scholar
Тахтаджан, А. 1969. Цветковые растения, происхождение и распространение. Оливер и Бойд, Эдинбург.
Google Scholar
—. 1991. Эволюционные тенденции цветковых растений. Издательство Колумбийского университета, Нью-Йорк.
1991. Эволюционные тенденции цветковых растений. Издательство Колумбийского университета, Нью-Йорк.
Google Scholar
Тиллих, Х.-Й. 1992. Bauprinzipin und Evolutionslinien bei monocotylen Keimpflanzen. Бот. Ярб. Сист. 114:91–132.
Google Scholar
—. 1995. Саженцы и систематика однодольных растений. Стр. 303–352 в P. J. Rudall et al. (ред.), Однодольные: систематика и эволюция. Том. 1. Королевские ботанические сады, Кью.
Google Scholar
Титова Г.Е. и Батыгина Т.Б. . 1996. Является ли зародыш кувшинковых растений (Nymphaeales s.l.) двудольным? Фитоморфология 46: 171–190.
Google Scholar
Валлад, Дж., Ф. Буньон и З. Ибаннайн . 1993. Морфологическая интерпретация эмбрионов среди зародышей с применением злаков (Poaceae). Канада. Дж. Бот. 71: 256–272.
Ибаннайн . 1993. Морфологическая интерпретация эмбрионов среди зародышей с применением злаков (Poaceae). Канада. Дж. Бот. 71: 256–272.
Артикул
Google Scholar
Вагнер, Г. П. 1989a. Происхождение морфологических признаков и биологические основы гомологии. Эволюция 43: 1157–1171.
Артикул
Google Scholar
—. 1989б. Концепция биологической гомологии. Энн. Преподобный Экол. Сист. 20: 51–69.
Артикул
Google Scholar
Williams, DM, RW Scotland и S. Blackmore . 1990. Существует ли в систематике прямой онтогенетический критерий? биол. Дж. Линн. соц. 39: 99–108.
Артикул
Google Scholar
Wilson, C.L. 1979. Idiospermum australiense (Idiospermaceae) — аспекты вегетативной анатомии. амер. Дж. Бот. 66: 280–289.
Idiospermum australiense (Idiospermaceae) — аспекты вегетативной анатомии. амер. Дж. Бот. 66: 280–289.
Артикул
Google Scholar
Wimsatt, WC и SC Schank . 1988. Два ограничения эволюции сложных адаптаций и средства их предотвращения. Стр. 231–273 в М. Х. Нитецкий (ред.), Эволюционный прогресс. Издательство Чикагского университета, Чикаго.
Google Scholar
Yamashita, T. 1976. Über die Embryound Wurtzelentwicklung bei Aponogeton madagascariensis (Mirbel) van Bruggen. Дж. Фак. науч. ун-т Токио, секта. 3, Бот. 12: 37–63.
Google Scholar
Однодольные против двудольных – Heatherkellyblog – путешествуйте со мной и удивляйтесь тому, что видите!
Я и раньше задавался этим вопросом, но так и не ответил на этот вопрос к моему собственному удовлетворению – почему в это время года (март) так много растений, которые мы видим в цвету, являются однодольными или однодольными для краткости? Подумайте, например, о подснежниках, нарциссах, ирисах, тюльпанах и крокусах.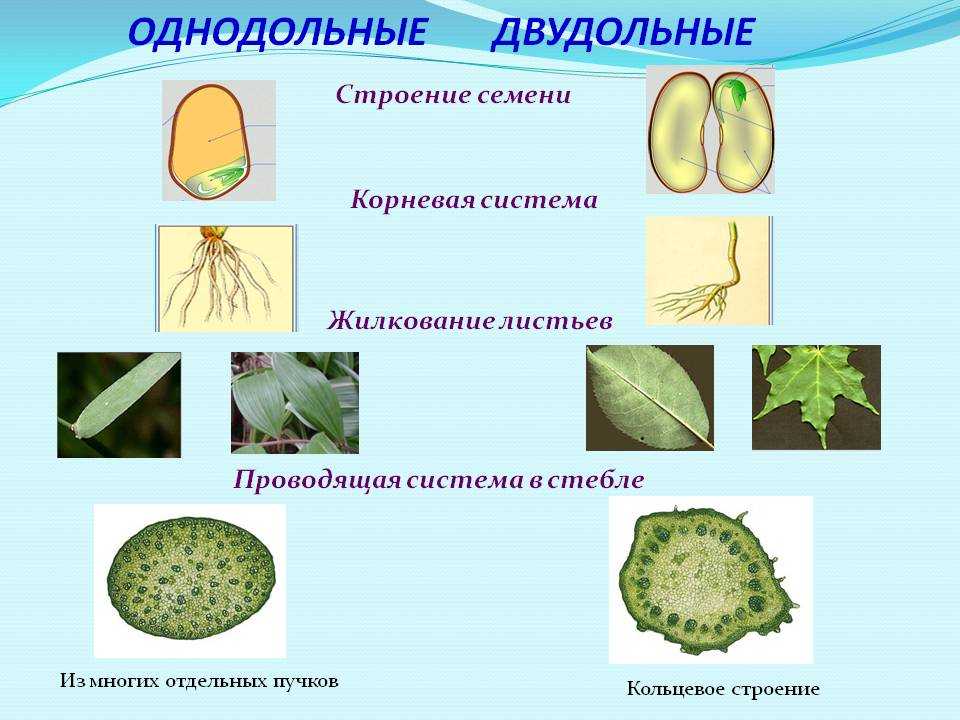
ЦИФРОВАЯ КАМЕРА OLYMPUS
То же самое было на прошлой Пасхе, когда мы были в Узбекистане, где растут некоторые из наших весенних садовых цветов – ирисы, лилии, тюльпаны и Вифлеемская звезда.
Olympus Digital Camera
Olympus Digital Camera
Olympus Digital Camera
Olympus Digital Camera
Слева справа, сверху до дна: Iris Tubergeniana, Ornithogalum Sp., Tulipa Bifloriformis и Gageaea Lutea,
,
,
,
,
,
,
,
,
,
,
,
,
,
,
, 9000 3,
, 9000 3, для Практики. систематики растений традиционно делили цветковые растения ( покрытосеменных ) на две группы — однодольные и двудольные — на основании числа зародышевых листьев ( семядолей ) внутри их семян. Однодольные, как следует из названия, имеют один семенной лист, а двудольные — два. Однако с 1990-х годов мы смогли использовать унаследованные различия в последовательностях ДНК организмов в качестве своего рода молекулярных часов для получения информации об их эволюционных взаимоотношениях. Если предположить, что скорость, с которой происходят изменения в генетическом коде организмов, остается неизменной в течение длительных периодов времени (тысячелетий), то чем дольше она длится с тех пор, как два вида отделились друг от друга, тем больше различий будет в последовательности изменений. оснований, составляющих их генетический код. Мы можем использовать эту информацию для создания молекулярного филогенетического дерева, которое может показать новые и удивительные эволюционные отношения.
Если предположить, что скорость, с которой происходят изменения в генетическом коде организмов, остается неизменной в течение длительных периодов времени (тысячелетий), то чем дольше она длится с тех пор, как два вида отделились друг от друга, тем больше различий будет в последовательности изменений. оснований, составляющих их генетический код. Мы можем использовать эту информацию для создания молекулярного филогенетического дерева, которое может показать новые и удивительные эволюционные отношения.
Такое исследование показало нам, что двудольные на самом деле не являются единой группой, состоящей из всех потомков общего предка, но в разное время отделились от этой родовой линии. Некоторые, такие как Magnolia и ее родственники и так называемые базальных покрытосеменных растений (включая водяные лилии, Nymphaea ), разошлись раньше однодольных, которые ранее считались самой старой линией.
ЦИФРОВАЯ КАМЕРА OLYMPUS
ЦИФРОВАЯ КАМЕРА OLYMPUS
Magnolia grandiflora (слева) и Nymphaea tetragona (справа)
Кладограмма, показывающая положение однодольных среди покрытосеменных растений по состоянию на APG IV (2016)
называемые двудольными, теперь попадают в категорию eudicot , хотя разделение пяти групп внутри основных покрытосеменных на диаграмме выше произошло за относительно короткий промежуток времени (около 4 миллионов лет) по сравнению с эволюцией покрытосеменных в целом. Однако термин «двудольные» все еще иногда используется для обозначения всего, что не является однодольным, и это то, что я собираюсь здесь сделать.
Однако термин «двудольные» все еще иногда используется для обозначения всего, что не является однодольным, и это то, что я собираюсь здесь сделать.
Однодольные, напротив, представляют собой монофилетическую линию, включающую всех потомков их общего предка. У этой группы есть ряд отличительных морфологических особенностей, кроме единственного семенного листа, что позволяет их довольно легко обнаружить; их листья обычно длинные и тонкие, с жилками, идущими параллельно краям, а не, например, в сети разветвлений. Части цветка состоят из трех или кратных им частей, а не из четырех и пяти, более типичных для двудольных.
Ирис желтый флаг, Iris pseudoacorus
У однодольных также отсутствуют боковые меристемы , поэтому они не могут производить вторичный рост, то есть боковой рост, сопровождающий увеличение высоты двудольных деревьев и других крупных многолетних растений – подробнее об этом значение этого грядет…
Орхидеи — крупнейшее семейство однодольных, но травы наиболее важны с экономической точки зрения, включая все наши основные зерновые культуры, сахарный тростник и бамбук.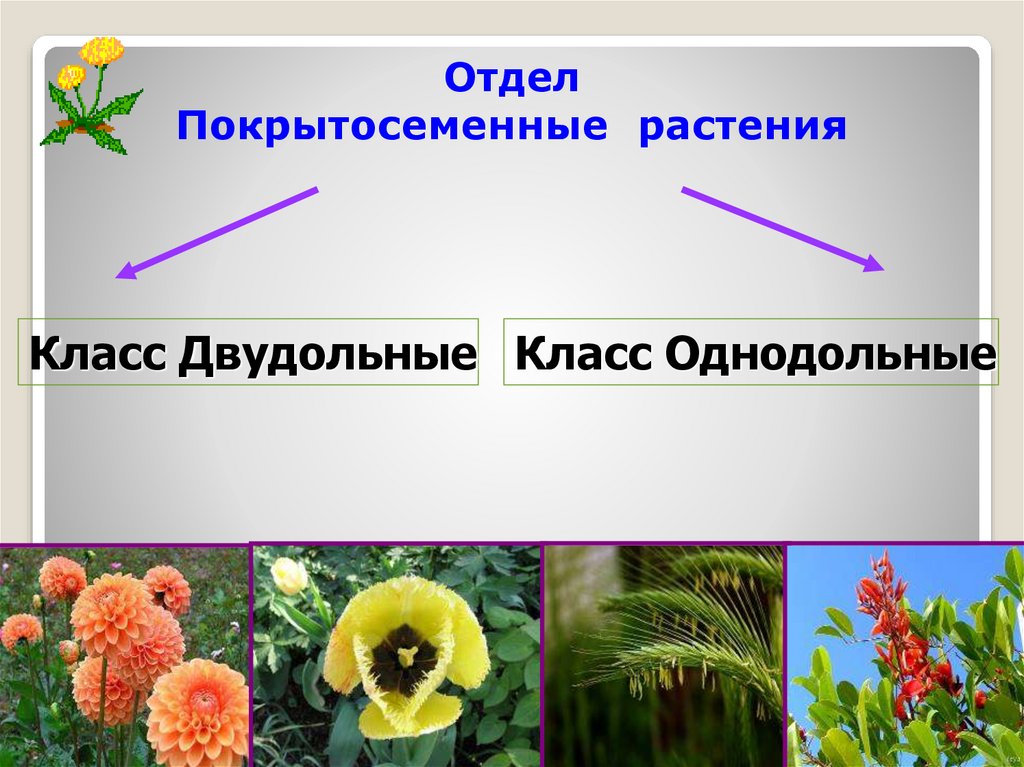 Различные пальмы, бананы и бананы, лук-порей, лук и чеснок — все это однодольные растения, как и такие специи, как имбирь, куркума и кардамон.
Различные пальмы, бананы и бананы, лук-порей, лук и чеснок — все это однодольные растения, как и такие специи, как имбирь, куркума и кардамон.
ЦИФРОВАЯ КАМЕРА OLYMPUS
ЦИФРОВАЯ КАМЕРА OLYMPUS
Слева направо: имбирь (Zingiber spectabile), орхидея вересковая пятнистая (Dactylorhiza maculata), голубая вересковая трава (Sesleria caerulea)
900 дерево здесь — все раннецветущие однодольные, о которых я думал, — это, по-видимому, случайно, многолетние виды, которые полагаются на подземные запасающие органы. В основном они размножаются вегетативно из луковиц или клубнелуковиц (обеих видоизмененных частей подземного стебля), а не из семян. Не в этом ли залог их раннего цветения? В конце концов, другой древний главный продукт британских лесов, актиния, не является однодольным растением, а распространяется в основном через подземные корневища.
Древесные анемоны (двудольные – Anemone nemorosa) перед местными колокольчиками (однодольные – Hyacinthoides non-scripta), Дарем
Существует множество причин, по которым раннецветущие растения могут больше полагаться на вегетативное распространение чем при семенном производстве – в начале года меньше насекомых для опыления; растение получает меньше энергии от солнца, потому что интенсивность света ограничена; температура ниже, поэтому все реакции, необходимые для роста, протекают медленнее. Хотя мы выращиваем многие из этих видов в наших парках и садах для весеннего цвета, их естественная среда обитания часто находится в лесистой местности, где необходимо быстрое завершение цикла роста, прежде чем деревья наверху полностью заблокируют их свет. Я много думал об этом в 2016 году, когда следил за развитием платана и его нижнего яруса в течение года.
Хотя мы выращиваем многие из этих видов в наших парках и садах для весеннего цвета, их естественная среда обитания часто находится в лесистой местности, где необходимо быстрое завершение цикла роста, прежде чем деревья наверху полностью заблокируют их свет. Я много думал об этом в 2016 году, когда следил за развитием платана и его нижнего яруса в течение года.
Однако вегетативное размножение и однодольные растения не так уж не связаны между собой, как может показаться. На рост корней у однодольных влияет то же отсутствие боковых меристем, что и на ствол — вы никогда не увидите однодольных с гигантской первичной структурой стержневого корня наших самых больших деревьев. Вместо этого корневые системы однодольных вынуждены рано и неоднократно ветвиться, чтобы образовать широкую сеть. Некоторые из этих корней на самом деле являются придаточными корнями, происходящими из побега. Легко увидеть, как некоторые из этих нижних побегов превращаются в корневища, которые часто имеют дополнительную функцию хранения — луковицы и клубнелуковицы являются их специализированными версиями.