Содержание
Двойное оплодотворение цветковых растений | Биология развития. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
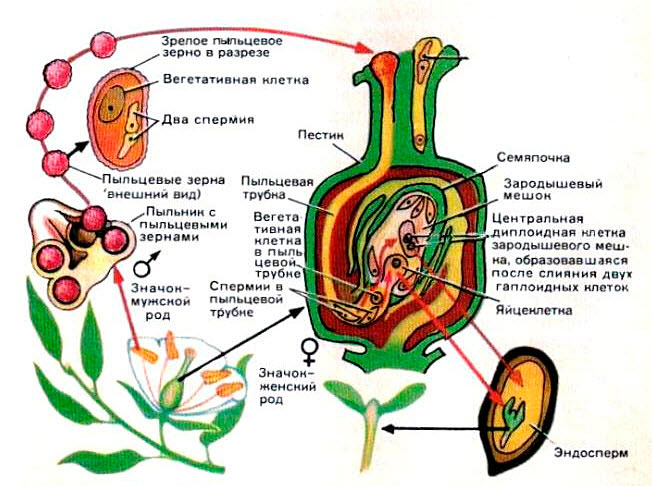
Двойное оплодотворение цветковых растений — это особый тип оплодотворения, который наблюдается только у цветковых (покрытосеменных) растений (рис 4). В зародышевом мешке, сформировавшемся в семязачатке завязи пестика цветка, развивается яйцеклетка. К моменту оплодотворения в зародышевом мешке помимо яйцеклетки образуется ещё двойное ядро (от слияния двух мелких клеток) и 5 других очень мелких (вспомогательных) клеток. Из проросшей на рыльце пестика пылинки по пыльцевой трубке в зародышевый мешок доставляются два спермия. Один из них сливается с яйцеклеткой, а другой спермий сливается с двойным ядром, находящимся в центральной части мешка. В итоге такого «двойного» оплодотворения из оплодотворенной яйцеклетки образуется зигота, дающая начало зародышу растения, а от слияния спермия с двойным ядром образуется особая питательная ткань, состоящая из клеток с триплоидным (3n) набором хромосом, — эндосперм, обеспечивающий питательными веществами зародыш семени.
Процесс двойного оплодотворения был открыт у цветковых растений на примере лилейных и детально изучен отечественным ботаником Сергеем Гавриловичем Навашиным в 1898 г.
Биологическое значение двойного оплодотворения у растений не совсем ясно. Однако несомненным является то, что в семенах очень быстро (опережая развитие зародыша) образуются питательные ткани с запасом высокоэнергетических питательных веществ только после оплодотворения. У цветковых растений при развитии семязачатков не тратится время на создание питательных веществ, как у голосеменных растений, поэтому развиваются они гораздо быстрее. Таким образом, благодаря двойному оплодотворению ускоряется процесс формирования и семяпочки, и яйцеклетки, и семени. Материал с сайта http://doklad-referat.ru
Загрузка…
Рис.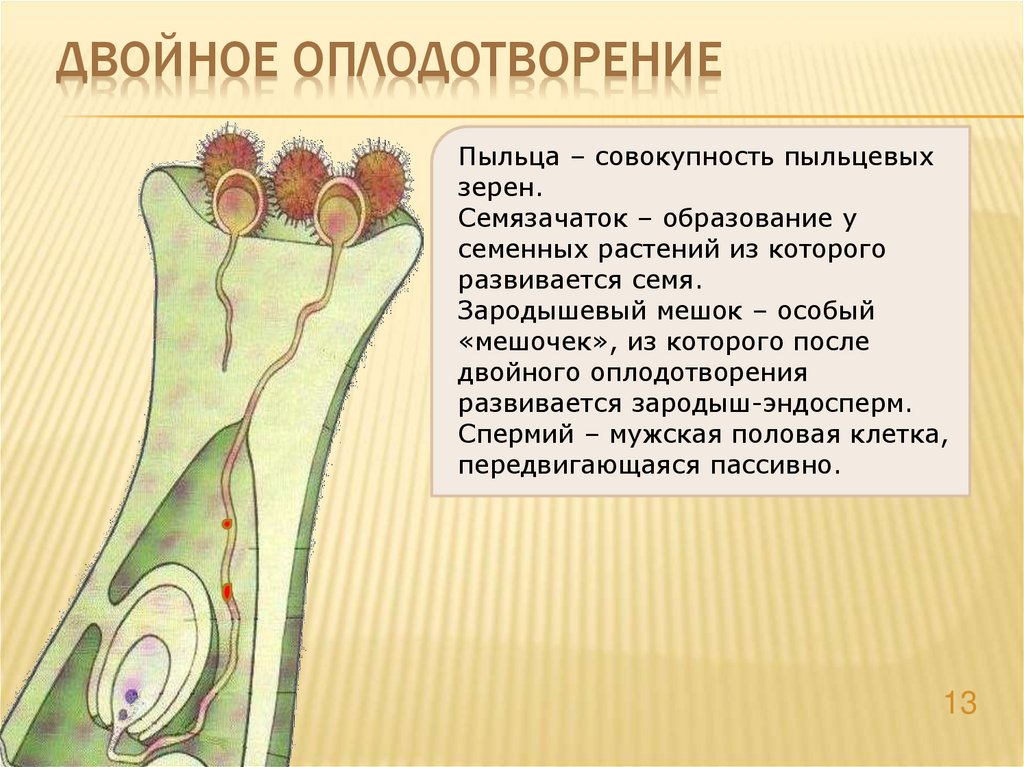 4. Двойное оплодотворение цветковых растений: 1 — яйцеклетка; 2 — завязь; 3 — центральное ядро; 4 — рыльце; 5 — пылинка; 6 — пыльник; 7 — семязачаток: 8 — зародышевый мешок; 9 — пыльцевая трубка; 10 — два спермия 4. Двойное оплодотворение цветковых растений: 1 — яйцеклетка; 2 — завязь; 3 — центральное ядро; 4 — рыльце; 5 — пылинка; 6 — пыльник; 7 — семязачаток: 8 — зародышевый мешок; 9 — пыльцевая трубка; 10 — два спермия |
На этой странице материал по темам:
Процесс у голосеменных как двойное оплодотворение цветковых
Конспект оплодотворение у цветковых растений
Двойное оплодотворение растений доклад
Двойное оплодотворение цветковых кратко
Оплодотворение у голосеменных
Вопросы по этому материалу:
Материал с сайта http://Doklad-Referat.ru
Загрузка…
Двойное оплодотворение покрытосеменных растений
Размножение
– это одно из обязательных свойств
любого живого организма.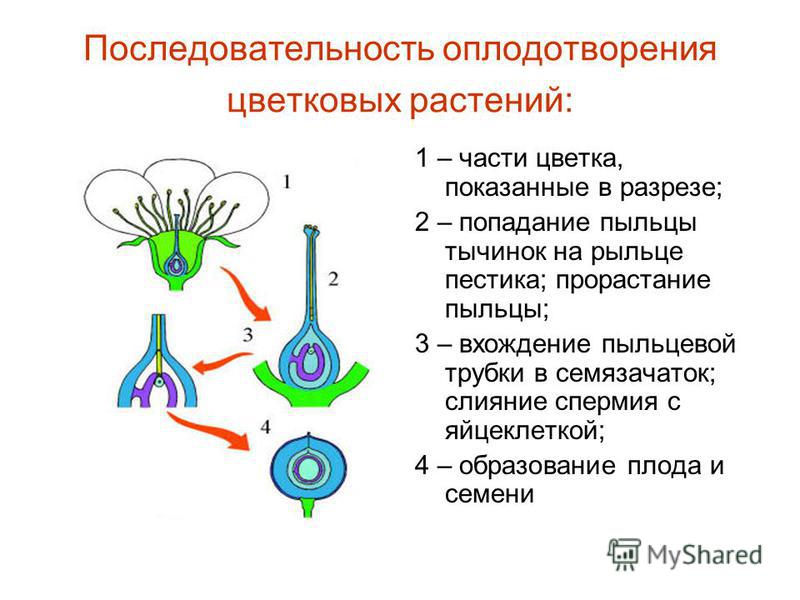 Оно заключается в увеличении числа особей.
Оно заключается в увеличении числа особей.
Различают
бесполое и половое размножение растений.
Бесполое размножение подразделяют на спорообразование и вегетативное.
Половое размножение происходит при
помощи особых половых клеток –гамет.
При
бесполом размножении быстро увеличивается численность вида, все потомки имеют
абсолютно такой же генотип, что и родительская особь. А также не происходит
увеличения генетического разнообразия, которое может оказаться очень полезным
при изменении условий существования вида.
По
этой причине большинство живых организмов на Земле размножаются половым
путём.
Сущность
полового размножения заключается в слиянии генетической информации родителей,
благодаря чему генетическое разнообразие в потомстве увеличивается.
У
покрытосеменных растений половое размножение связано с цветком.
Важные
части цветка – это пестик и тычинка.
С их участием происходят сложные процессы полового размножения –опыление и оплодотворение.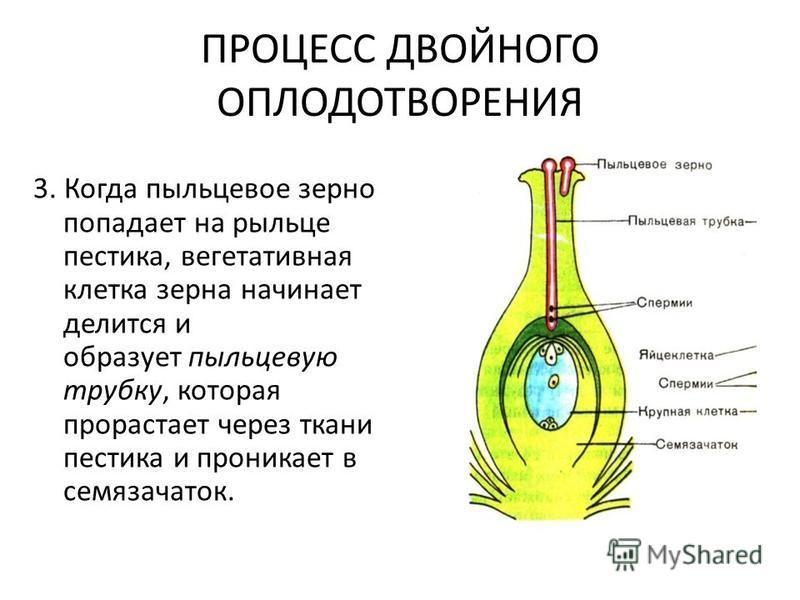
Но
сперва в будущем цветке начинают образовываться половые клетки.
В
жизненном цикле цветковых растений наблюдается смена поколений.
У
растений выделяют диплоидное поколение –
бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Гаметы
образуются в результате митоза, а споры –
в результате мейоза. И гаметы, и споры образуются в цветке, поэтому цветок
является органом и бесполого, и полового размножения.
Пыльцевое
зерно является спорой (микроспорой),
а не мужской гаметой, так как в нем самом развиваются мужские гаметы.
У
цветковых растений мужские гаметофиты
столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего
лишь из нескольких клеток.
Женский
гаметофит цветковых (зародышевый мешок) помещается внутри
семяпочки и состоит в наиболее распространённом случае из 7 клеток (содержит 8 – либо 7 после слияния двух ядер в
центральной клетке и образования вторичного ядра).
Посмотрим, как образуется мужской гаметофит.
Итак,
Микроспорогенез
В субэпидермальной ткани молодого
пыльника обособляется специальная спорогенная ткань,
называемая археспорием. Каждая первичная археспориальная
клетка после ряда делений становится материнской
клеткой пыльцы (микроспороцитом), которая проходит все
фазы мейоза.
В результате двух мейотических делений
возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и
называются клеточными тетрадами.
При созревании клеточные тетрады
распадаются на отдельные микроспоры с образованием внутренней (интина) и
наружной (экзина)
оболочек. Наружная оболочка, как правило, грубая, поверхность её либо гладкая,
либо шероховатая, приспособленная для переноса пыльцы и прилипания её к рыльцу
пестика.
Этим заканчивается микроспорогенез,
вслед за образованием одноядерной микроспоры начинается микрогаметогенез.
Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В
В
дальнейшем вегетативная клетка и её ядро не делятся. В ней накапливаются запасные питательные вещества, которые в
последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в
столбике пестика.
Генеративная клетка, содержащая
меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться ещё
в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В
результате образуются две мужские половые клетки, которые, в отличие от
сперматозоидов животных, называются спермиоклетками,
или спермиями.
Таким образом, из одной споры (микроспоры) с гаплоидным набором
хромосом в результате двух митотических делений образуются три ядра: два из них – спермии и одно – вегетативное. При
образовании пыльцевой трубки это вегетативное ядро
переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в
пыльцевой трубке были впервые подробно изучены российским и советским цитологом и эмбриологом растений Сергеем Гавриловичем Навашиным в
1910 г.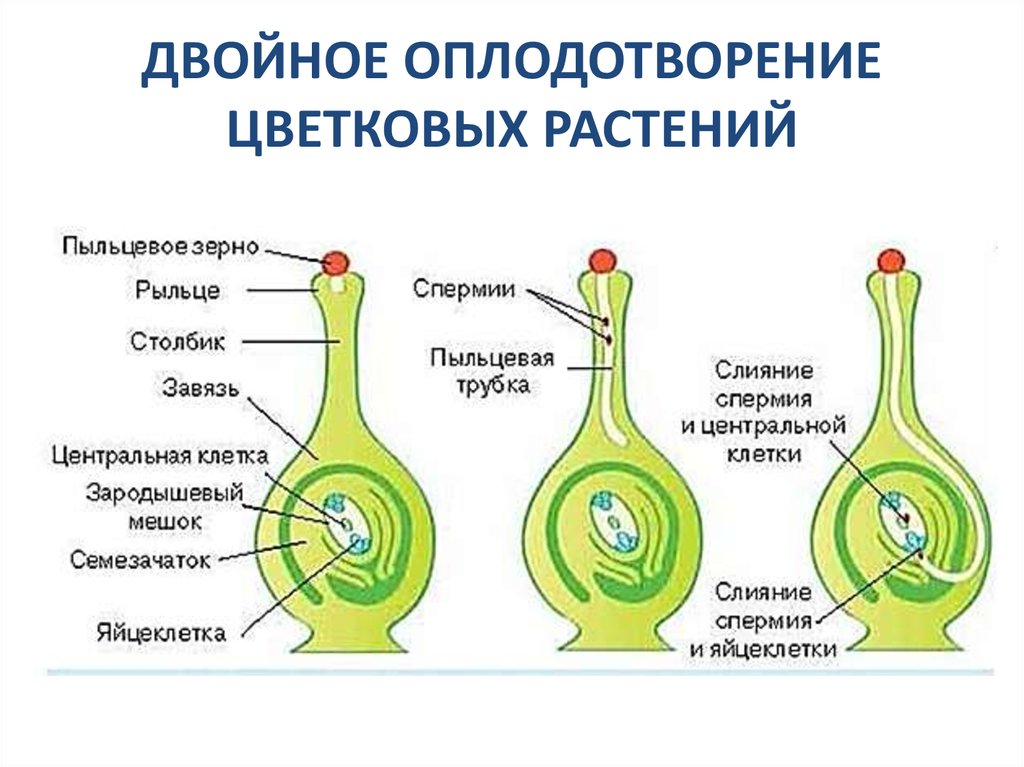 на лилейных растениях.
на лилейных растениях.
После образования гамет пыльник созревает, и пыльца высыпается.
Она несёт только генетическую информацию.
Посмотрим, как происходит мегаспорогенез
и мегагаметогенез цветковых.
У
покрытосеменных растений женский гаметофит –
это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Развитию
женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез.
В
субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка
археспория растёт, превращаясь в
материнскую клетку мегаспоры.
В
результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из
клеток тетрады по числу хромосом является гаплоидной.
Однако только одна из них продолжает развиваться, остальные три дегенерируют,
судьба этих клеток напоминает судьбу редукционных телец при созревании
яйцеклеток у животных.
На
следующем этапе осуществляется мегагаметогенез.
Оставшаяся функционировать мегаспора продолжает расти и затем её ядро
претерпевает ряд делений. При этом сама клетка не делится, а делится только
ядро.
У
разных систематических групп растений число делений ядра мегаспоры может
варьировать от одного до трёх. У большинства растений (70 % видов
покрытосеменных) этих делений, как правило, в результате возникает восемь
наследственно одинаковых ядер, вовремя этих делений ядра занимают
полярное положение, четыре из них оказываются лежащими ближе к микропиле
(место проникновения спермиев), а четыре других – в противоположном конце зародышевого мешка, называемого халазальным.
Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные
количества цитоплазмы.
В
дальнейшем от каждой из двух полярных четвёрки ядер к центру отходит по одному ядру,
которые сливаются, образуя вторичное
(центральное) ядро зародышевого мешка. Затем цитоплазма обособляется
вокруг ядер гаметофита, который из ядерной стадии развития переходит в
клеточную.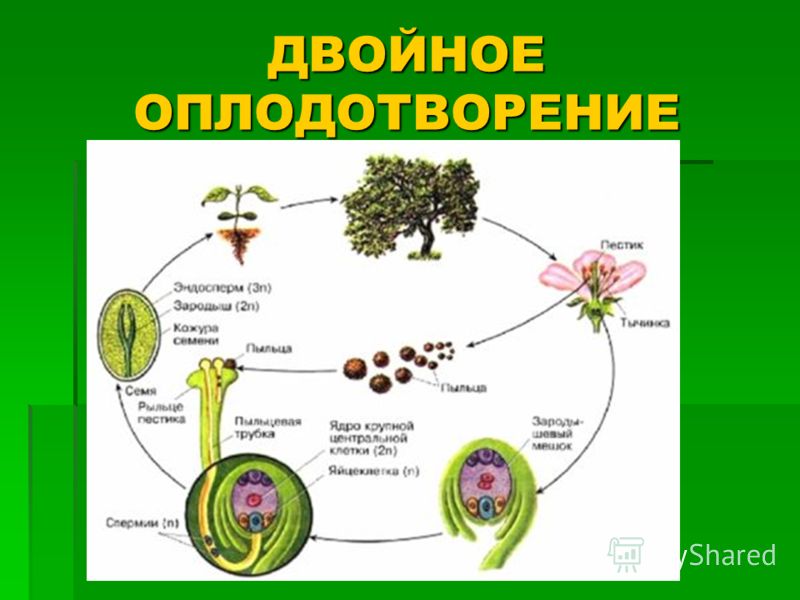 Три ядра, оставшиеся вблизи халазального
Три ядра, оставшиеся вблизи халазального
полюса, преобразуются в три клетки
(антиподы), которые питают гаметофит. Три ядра вблизи микропиле
отделяются клеточными перегородками, образуя отдельные клетки: крупную центральную яйцеклетку и
две боковые клетки синергиды.
Вся цитоплазма, расположенная между антиподами с одной стороны и клетками
яйцевого комплекса с другой (яйцеклетка и две синергиды), называется центральной клеткой. В ней находится
диплоидное вторичное (центральное) ядро.
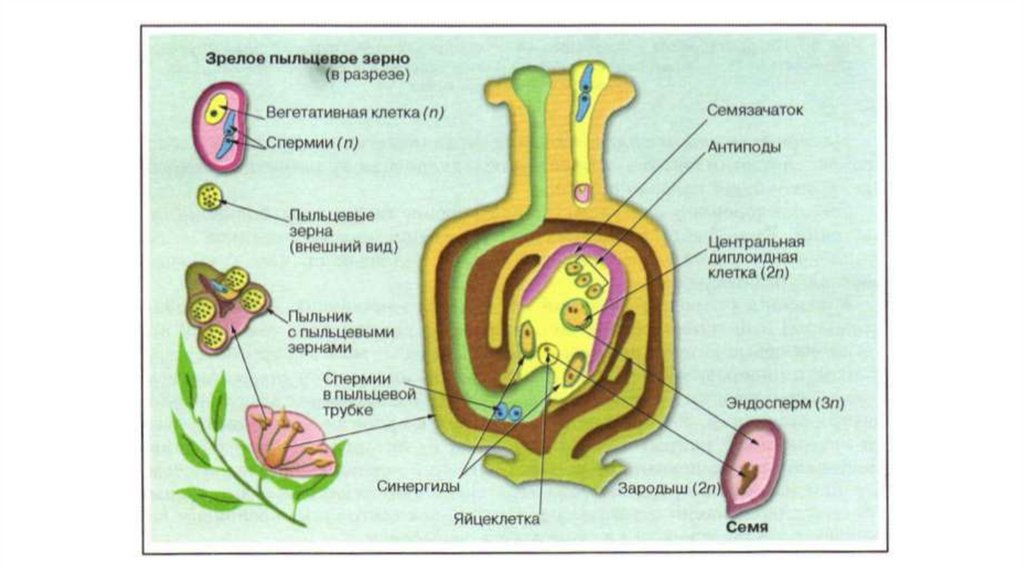
На
этом этапе женский гаметофит уже полностью сформирован и состоит из шести гаплоидных клеток (одной яйцеклетки, двух синергид и трёх
антипод) и одной диплоидной
(центральной). Его строение внешне напоминает мешочек, поэтому женский
гаметофит покрытосеменных называется зародышевым мешком.
Пылинка
попадает на рыльце пестика, и происходит опыление.
Опыление – это перенос пыльцевых зёрен на рыльце
пестика, у голосеменных пыльцевые зерна при опылении
попадают непосредственно на семязачаток.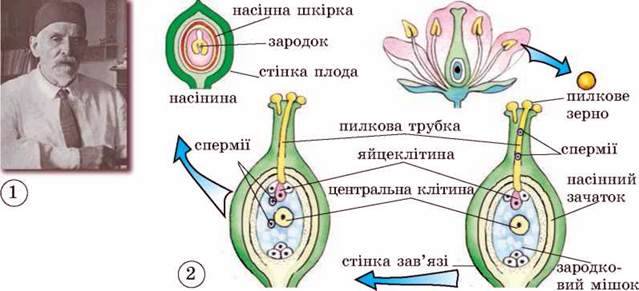
Имеется
два основных типа опыления: самоопыление (автогамия) (когда растение
опыляется собственной пыльцой) и перекрёстное опыление (аллогамия).
При
самоопылении исключён обмен генетической информацией,
поскольку пыльцевые зерна попадают на рыльце пестика либо с одной из тычинок
этого же цветка, либо с другого цветка, расположенного на том же растении. Это
приводит к появлению чистых линий гомозиготных популяций в пределах одного
вида, неспособных обмениваться мутировавшими генами, поэтому процессы
видообразования в этих популяциях идут самостоятельно.
Перекрёстное
опыление – это
перенос пыльцы одного растения на рыльце другого. Этот тип опыления встречается
более часто, чем самоопыление, между разными особями одного вида происходит
обмен аллелями, что приводит к увеличению доли гетерозиготных организмов.
Безусловно,
перекрёстное самоопыление имеет большие преимущества по сравнению с
самоопылением, поскольку возникшие мутации свободно распространяются в пределах
популяции.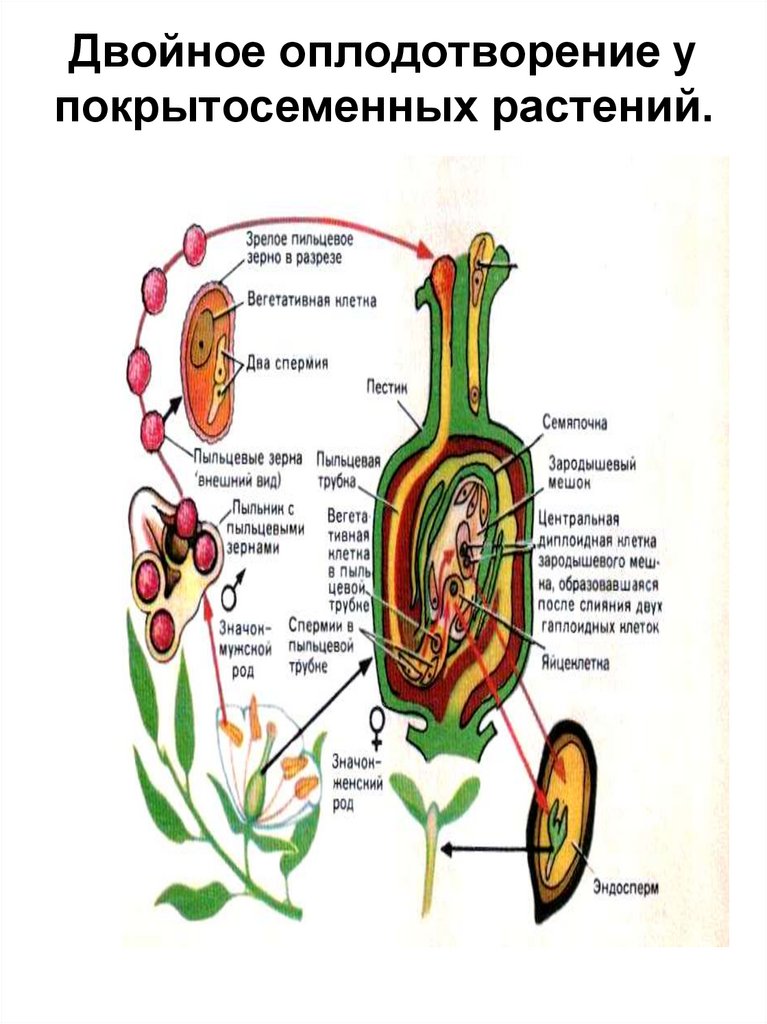
Способы
опыления у перекрёстноопыляемых
цветковых растений весьма разнообразны. Их можно разделить на две группы. Первая:
перенос пыльцы осуществляется главным образом насекомыми, а также некоторыми
позвоночными (птицами и летучими мышами). Соответственно, различают энтомофилию, орнитофилию и зоофилию. Растения, опыляемые животными, обычно имеют
яркоокрашенные крупные цветки. Мелкие цветки, как правило, собраны в соцветия,
что зрительно их увеличивает. Для привлечения опылителей служит нектар или
большое количество пыльцы, которую опылители охотно поедают.
Вторая
группа: пыльца переносится абиотическими факторами – ветром и реже водой, в связи с чем различают анемофилию и
гидрофилию.
Оплодотворение.
После
попадания пыльцы на рыльце начинается прорастание пыльцевого зерна.
Экзина
мужского гаметофита прорывается в области борозды или поры прорастания, и
начинает формироваться пыльцевая трубка, которая растёт, проникает в рыльце,
через столбик движется по направлению к завязи, где находится семязачаток.
Трубка
растёт на кончике. В растущую пыльцевую трубку из пыльцевого зерна перемещается
ядро клетки-трубки, а также спермин.
Обычно
развитие пыльцевой трубки происходит при уже сформированных семязачатках в
завязи. Достигнув завязи, пыльцевая трубка через микропиле проникает внутрь одного из
находящихся там семязачатков.
Там
пыльцевая трубка направляется к яйцевому аппарату, проникает в одну из синергид и
освобождает спермин. Один из спермиев
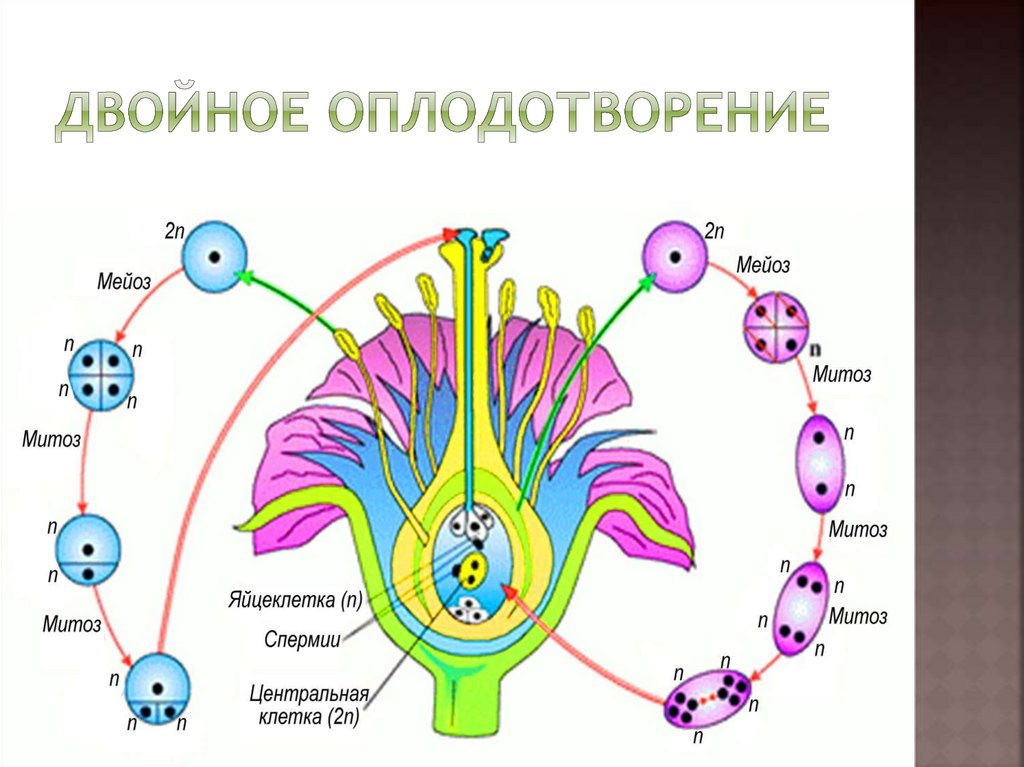
сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами
центральной клетки, образуя триплоидное ядро (с тройным набором хромосом). Зигота даёт начало зародышу, а из триплоидной
центральной клетки образуется ткань эндосперма.
Таким
образом оплодотворение, при котором одна мужская гамета сливается с
яйцеклеткой, а вторая ― с вторичным ядром, называется двойным
оплодотворением.
Механизм
оплодотворения цветковых –двойное
оплодотворение, открыто отечественным учёным Сергеем
Гавриловичем Навашиным в 1898 году.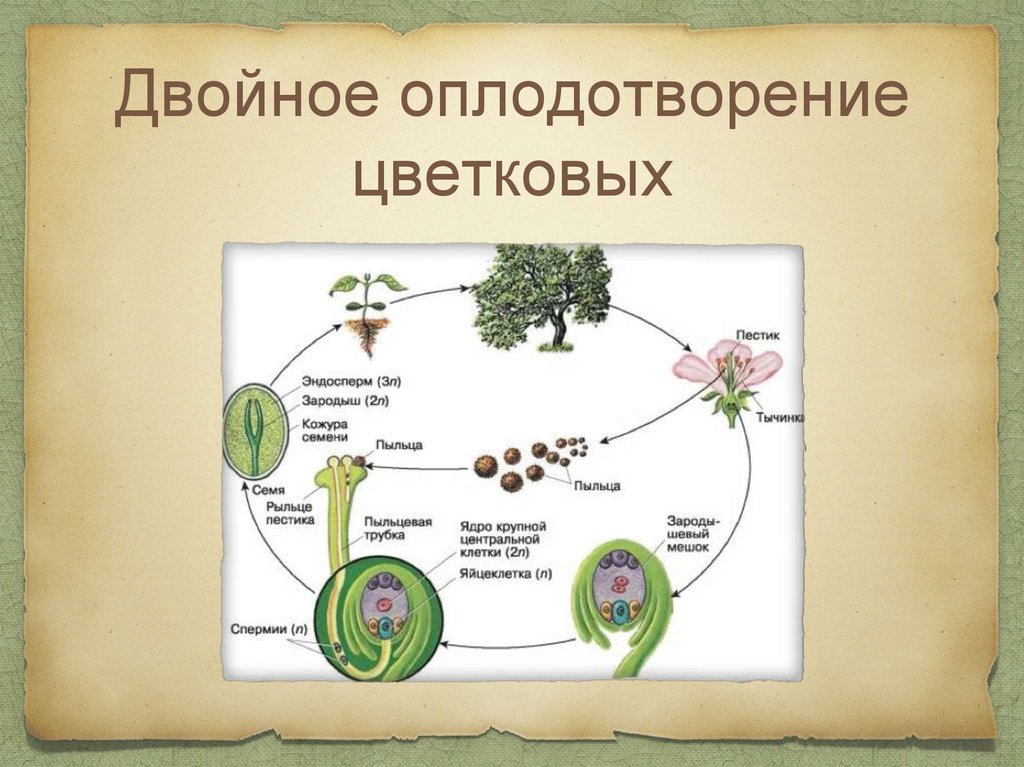
Таким образом, оплодотворённый семязачаток
развивается в семя, из зиготы возникает зародыш, из триплоидной центральной клетки – эндосперм, а из внешней части
семязачатка (интегументов) образуется семенная кожура, из стенок завязи цветка – стенки плода.
Развитие эндосперма.
Эндосперм
цветковых полностью отличается от первичного эндосперма голосеменных. У них
первичный эндосперм представляет собой гаплоидную вегетативную ткань женского
гаметофита, где накапливаются питательные вещества семени.
Эндосперм
у покрытосеменных происходит из триплоидной
центральной клетки, в образовании которой участвуют вторичное ядро женского
гаметофита и ядро спермия.
У
одних цветковых (например, злаков) эндосперм сильно разрастается
и занимает большую часть семени, оттесняя на периферию маленький зародыш.
У
других (к пример у бобовых) весь эндосперм поглощается
зародышем и используется его семядолями, которые становятся самыми большими
структурами зрелого семени.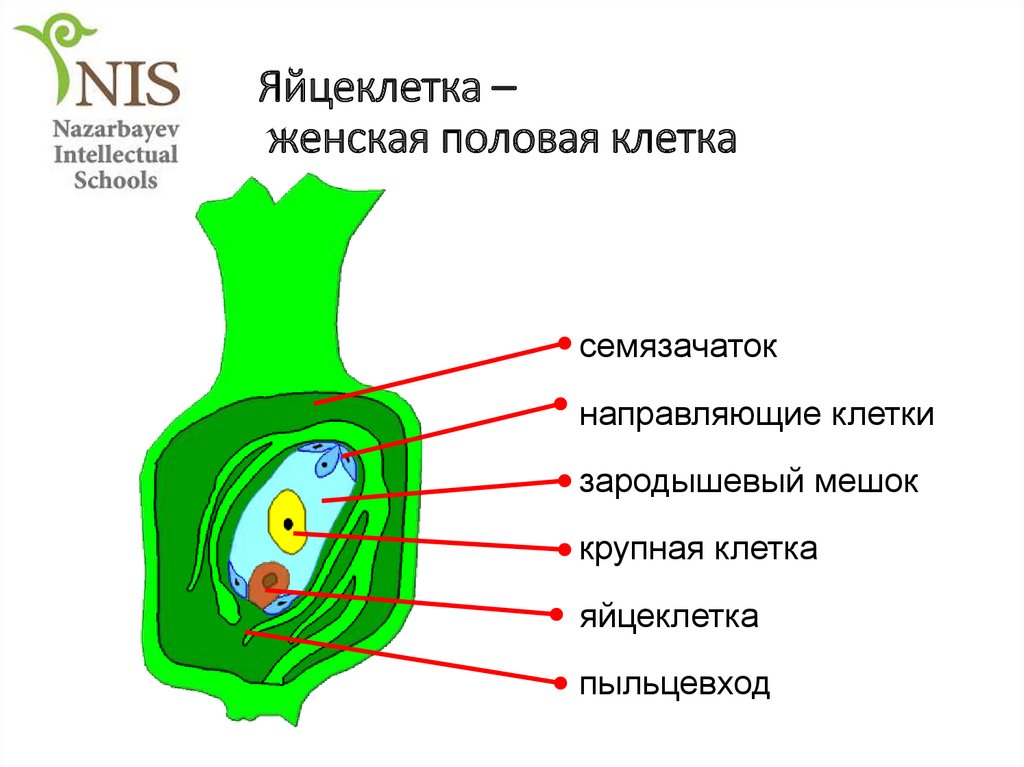
Двойное оплодотворение | Биология для специальностей II
Результаты обучения
- Дайте определение двойному оплодотворению
После того, как пыльца отложится на рыльце пестика, она должна прорасти и пройти через столбик, чтобы достичь семязачатка. Микроспоры, или пыльца, содержат две клетки: клетку пыльцевой трубки и генеративную клетку. Клетка пыльцевой трубки вырастает в пыльцевую трубку, по которой проходит генеративная клетка. Для прорастания пыльцевой трубки требуются вода, кислород и определенные химические сигналы. Проходя через столбик и достигая зародышевого мешка, рост пыльцевой трубки поддерживается тканями столбика. Тем временем, если генеративная клетка еще не разделилась на две клетки, она теперь делится, образуя два спермия. Пыльцевая трубка направляется химическими веществами, выделяемыми синергидами, присутствующими в зародышевом мешке, и входит в семязачаточный мешок через микропиле. Из двух сперматозоидов один сперматозоид оплодотворяет яйцеклетку, образуя диплоидную зиготу; другой сперматозоид сливается с двумя полярными ядрами, образуя триплоидную клетку, которая развивается в эндосперм .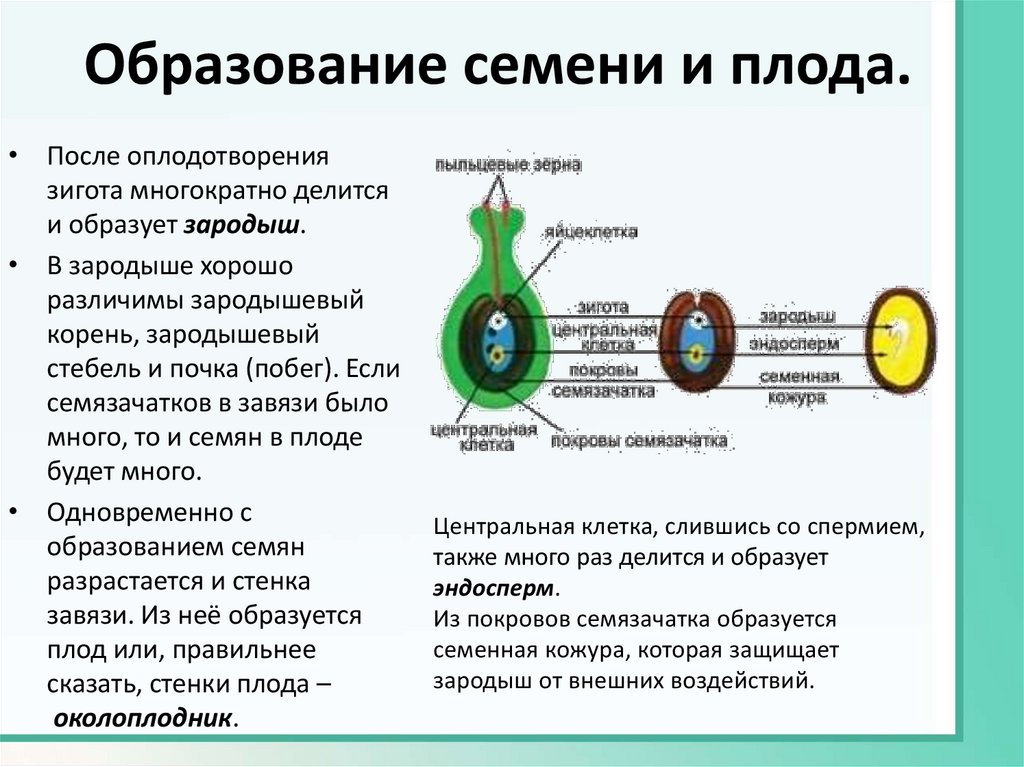 Вместе эти два события оплодотворения у покрытосеменных известны как двойное оплодотворение (рис. 1). После завершения оплодотворения никакие другие сперматозоиды не могут проникнуть внутрь. Оплодотворенная семяпочка образует семя, тогда как ткани завязи становятся плодом, обычно обволакивающим семя.
Вместе эти два события оплодотворения у покрытосеменных известны как двойное оплодотворение (рис. 1). После завершения оплодотворения никакие другие сперматозоиды не могут проникнуть внутрь. Оплодотворенная семяпочка образует семя, тогда как ткани завязи становятся плодом, обычно обволакивающим семя.
Рисунок 1. У покрытосеменных один сперматозоид оплодотворяет яйцеклетку с образованием зиготы 2 n , а другой сперматозоид оплодотворяет центральную клетку с образованием эндосперма 3 n . Это называется двойным оплодотворением.
После оплодотворения зигота делится на две клетки: верхнюю, или терминальную, и нижнюю, или базальную. Деление базальной клетки дает начало суспензору , который в конечном итоге соединяется с материнской тканью. Суспензор обеспечивает транспортировку питательных веществ от материнского растения к растущему зародышу. Терминальная клетка также делится, давая начало проэмбриону шаровидной формы (рис. 2а). У двудольных (эвдикотовых) развивающийся зародыш имеет форму сердца из-за наличия двух рудиментарных семядолей (рис.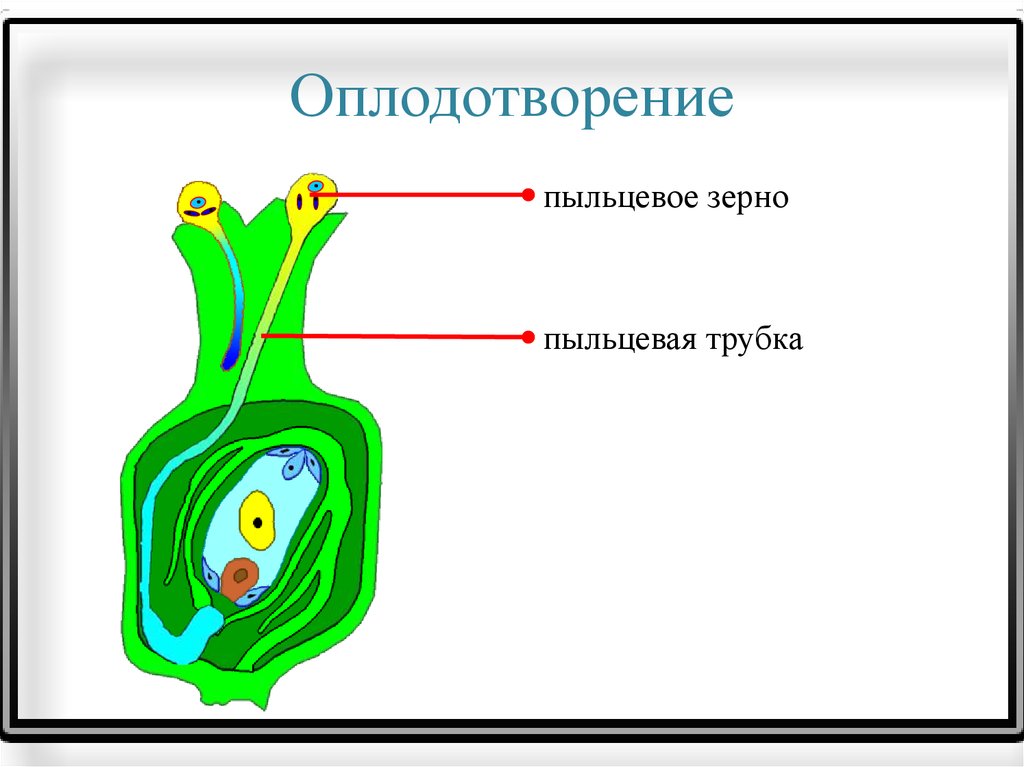 2б). У неэндоспермных двудольных, таких как Capsella bursa , сначала развивается эндосперм, но затем он переваривается, и запасы пищи перемещаются в две семядоли. По мере увеличения зародыша и семядолей им не хватает места внутри развивающегося семени, и они вынуждены изгибаться (рис. 2в). В конечном итоге зародыш и семядоли заполняют семя (рис. 2d), и семя готово к распространению. Эмбриональное развитие через некоторое время приостанавливается, и рост возобновляется только при прорастании семени. Развивающийся саженец будет полагаться на запасы пищи, хранящиеся в семядолях, до тех пор, пока первый набор листьев не начнет фотосинтез.
2б). У неэндоспермных двудольных, таких как Capsella bursa , сначала развивается эндосперм, но затем он переваривается, и запасы пищи перемещаются в две семядоли. По мере увеличения зародыша и семядолей им не хватает места внутри развивающегося семени, и они вынуждены изгибаться (рис. 2в). В конечном итоге зародыш и семядоли заполняют семя (рис. 2d), и семя готово к распространению. Эмбриональное развитие через некоторое время приостанавливается, и рост возобновляется только при прорастании семени. Развивающийся саженец будет полагаться на запасы пищи, хранящиеся в семядолях, до тех пор, пока первый набор листьев не начнет фотосинтез.
Рисунок 2. Показаны стадии развития эмбриона в семязачатке пастушьей сумки (Capsella bursa). После оплодотворения зигота делится на верхнюю терминальную и нижнюю базальную клетки. (а) На первой стадии развития терминальная клетка делится, образуя глобулярный проэмбрион. Базальная клетка также делится, давая начало суспензору.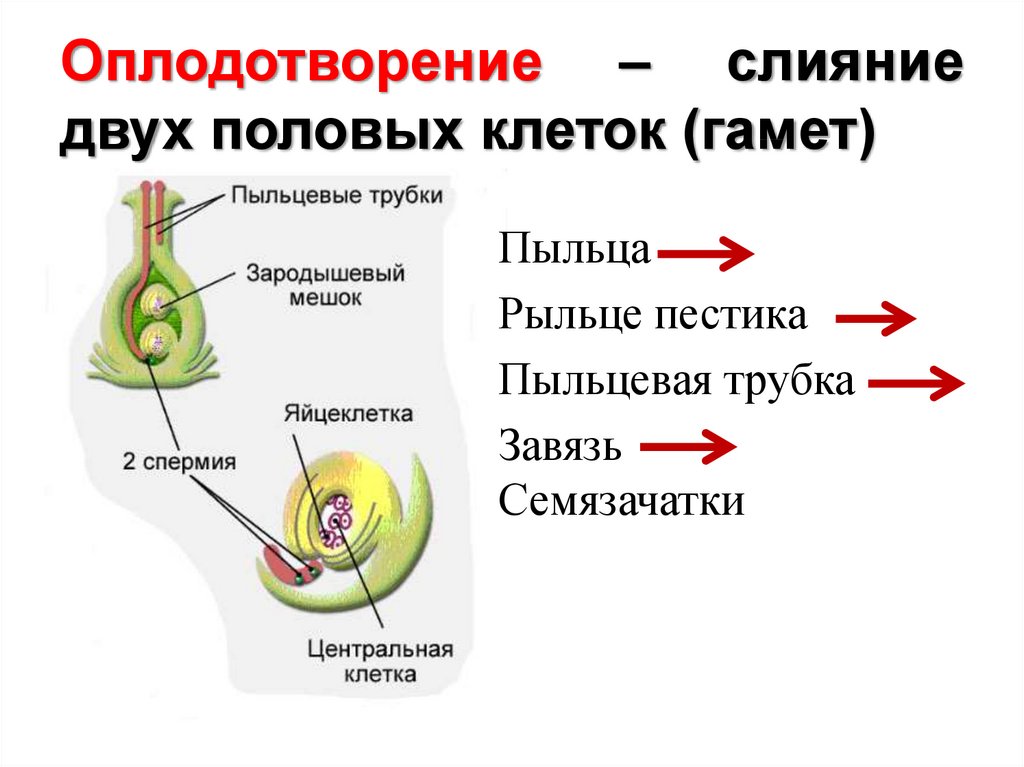 (б) На второй стадии развивающийся зародыш имеет форму сердца из-за наличия семядолей. (c) На третьей стадии растущий эмбрион выбегает из места и начинает изгибаться. (d) В конце концов, он полностью заполняет семя. (кредит: модификация работы Роберта Р. Уайза; данные масштабной линейки от Мэтта Рассела)
(б) На второй стадии развивающийся зародыш имеет форму сердца из-за наличия семядолей. (c) На третьей стадии растущий эмбрион выбегает из места и начинает изгибаться. (d) В конце концов, он полностью заполняет семя. (кредит: модификация работы Роберта Р. Уайза; данные масштабной линейки от Мэтта Рассела)
Попробуйте
У вас есть идеи по улучшению этого контента? Мы будем признательны за ваш вклад. Улучшить эту страницуПодробнее клетки-предшественники зародыша и эндосперма соответственно. Благодаря видоспецифическим процессам направления и притяжения пыльцевых трубок неподвижные сперматозоиды переносятся пыльцевой трубкой к семязачатку, в котором находится женский гаметофит. Два спермия объединяются с яйцеклеткой и центральной клеткой после разрыва пыльцевой трубки внутри женского гаметофита, вызывая образование семян. Эндосперм образован оплодотворенной центральной клеткой, а зародыш и суспензор — оплодотворенной яйцеклеткой — зиготой. Последний компонент соединяет зародыш со спорофитными материнскими тканями развивающегося семени.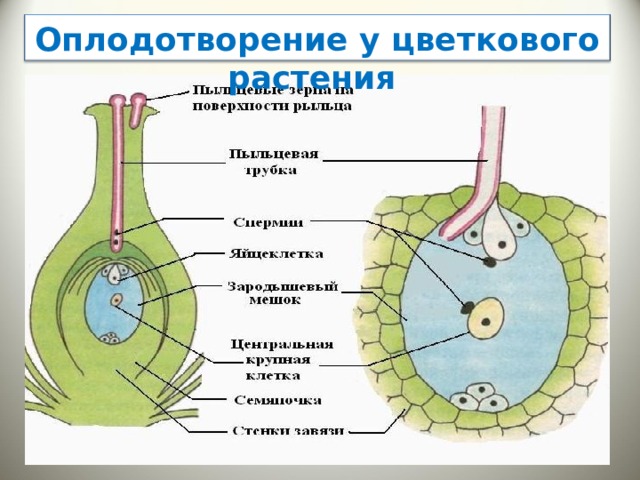 Чтобы обеспечить транспорт жизнеспособных сперматозоидов и производство как функциональной зиготы, так и эндосперма, основные механизмы двойного оплодотворения строго регулируются. Современное состояние знаний о направленном росте пыльцевых трубок и его связи с синергидными клетками, приводящими к разрыву пыльцевых трубок, взаимодействии четырех гамет, приводящем к слиянию клеток, и механизмах, с помощью которых цветковые растения предотвращают проникновение множественных сперматозоидов (полиспермия) для максимизации репродуктивной функции. Успех будет обсуждаться в этом обзоре.
Чтобы обеспечить транспорт жизнеспособных сперматозоидов и производство как функциональной зиготы, так и эндосперма, основные механизмы двойного оплодотворения строго регулируются. Современное состояние знаний о направленном росте пыльцевых трубок и его связи с синергидными клетками, приводящими к разрыву пыльцевых трубок, взаимодействии четырех гамет, приводящем к слиянию клеток, и механизмах, с помощью которых цветковые растения предотвращают проникновение множественных сперматозоидов (полиспермия) для максимизации репродуктивной функции. Успех будет обсуждаться в этом обзоре.
Двойное оплодотворение Определение
Размножение растений обычно достигается путем оплодотворения или, точнее, двойного оплодотворения. Двойное оплодотворение — это сложный механизм оплодотворения, который состоит в слиянии одного женского гаметофита (мегагаметофита, также называемого зародышевым мешком) с двумя мужскими гаметами (сперматозоидами).
Что такое двойное оплодотворение?
Двойное оплодотворение – это процесс, развившийся у цветковых растений (или покрытосеменных растений).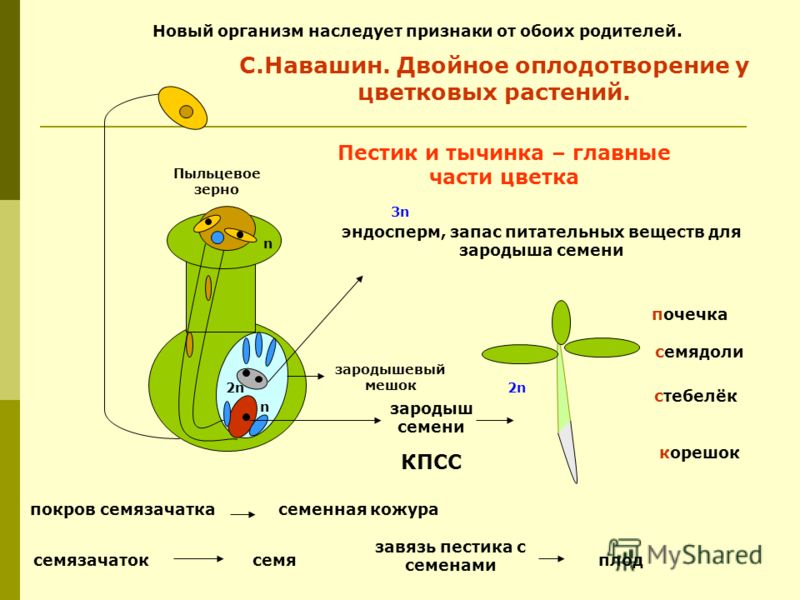 В общих чертах, процесс состоит из пары сперматозоидов, где яйцеклетка оплодотворяется одним сперматозоидом с образованием зиготы, а другой сперматозоид сливается с двумя полярными ядрами или вторичными ядрами, в результате чего образуется эндосперм.
В общих чертах, процесс состоит из пары сперматозоидов, где яйцеклетка оплодотворяется одним сперматозоидом с образованием зиготы, а другой сперматозоид сливается с двумя полярными ядрами или вторичными ядрами, в результате чего образуется эндосперм.
Репродуктивные структуры покрытосеменных
Покрытосеменные — это растения, производящие флору и конечный результат, и они являются наиболее распространенными видами растительной жизни на Земле. растения являются репродуктивными структурами покрытосеменных растений. Они состоят из 4 основных компонентов:
Плодолистик: женская репродуктивная структура, производящая яйца.
Тычинка: Мужская репродуктивная форма, производящая сперматозоиды.
Лепестки: окружают плодолистик и тычинку и регулярно ярко окрашены.

Чашелистики: занимают лепестки цветка до того, как он раскроется.
В ходе реплики цветкового растения образуются две системы: зигота и эндосперм. Зигота разовьется и превратится в зародыш или дочернее растение. Эндосперм является запасом пищи зародыша растения. Этот процесс образования зиготы и эндосперма называется двойным оплодотворением, и он очень точен для покрытосеменных.
Опыление
Чтобы произошло двойное оплодотворение, необходимо опыление. Когда пыльцевое зерно, произведенное тычинкой цветка, оседает на плодолистика другого цветка, это называется опылением. Пыльцевое зерно должно специально осесть на липкой платформе или рыльце плодолистика. Пыльцевое зерно представляет собой защитную структуру, содержащую сперму. У него вырастает пыльцевая трубка, когда она попадает на рыльце пестика. Эта структура будет развиваться от стиля плодолистика до его завязи.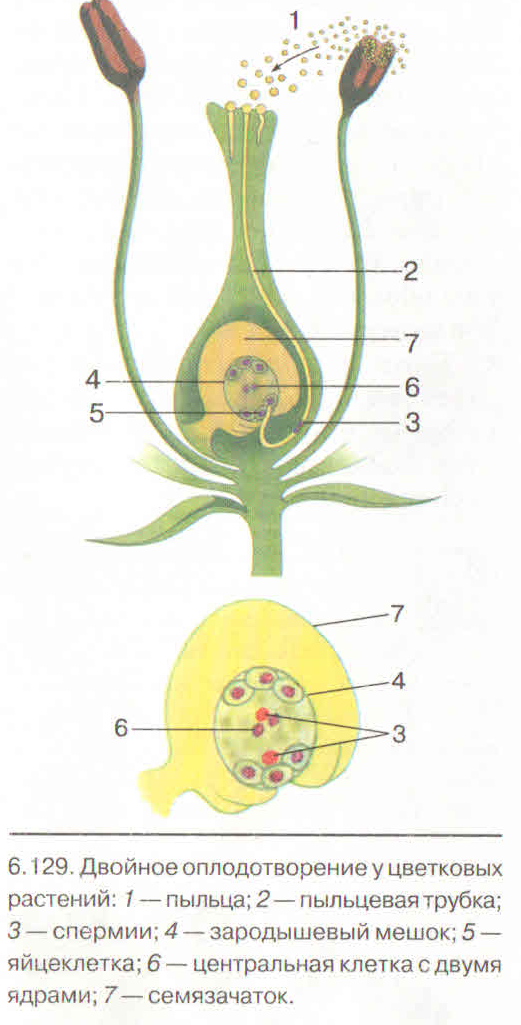 В яичнике произойдет двойное оплодотворение.
В яичнике произойдет двойное оплодотворение.
Процесс двойного оплодотворения
Пыльца должна развиваться и эволюционировать по всему столбику, чтобы пробиться в семязачаток после того, как она отложится на рыльце пестика. Пыльца или микроспоры состоят из двух клеток: генеративной клетки и клетки пыльцевой трубки.
Клетка пыльцевой трубки развивается в пыльцевую трубку, через которую проходит развивающаяся клетка. Эволюция пыльцевой трубки требует кислорода, воды и определенных химических веществ. Когда пыльцевая трубка проходит через столбик, чтобы пройти через зародышевый мешок, ткани столбика поддерживают его дальнейшее развитие.
На протяжении всей процедуры, если генеративная клетка еще не разделилась на две клетки, она диссоциирует с образованием двух сперматозоидов. Синергиды, существующие в зародышевом мешке, выделяют определенные химические вещества, которые обеспечивают путь для пыльцевой трубки. Далее пыльцевая трубка проходит через микропиле в семязачаточный мешок.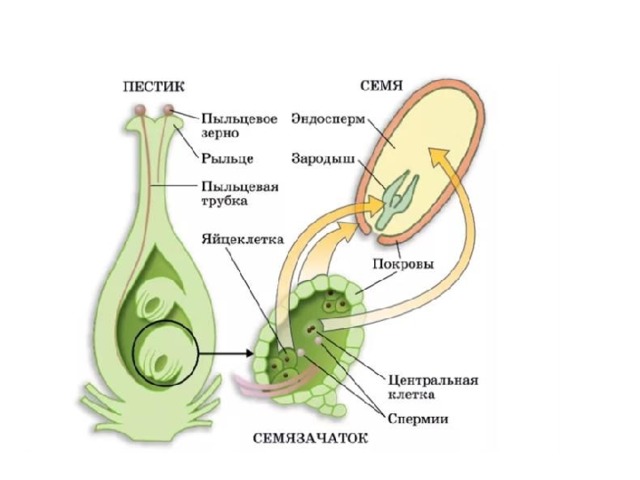
Любой из двух сперматозоидов оплодотворяет яйцеклетку с образованием диплоидной зиготы, а альтернативный сперматозоид связан с двумя другими вторичными ядрами (полярными ядрами) для развития триплоидной клетки, которая развивается в эндосперм.
Эти два отдельных метода оплодотворения у покрытосеменных в сочетании друг с другом называются двойным оплодотворением. После завершения механизма оплодотворения для других сперматозоидов не остается места. Оплодотворенная яйцеклетка производит семя, в то время как ткани завязи развиваются в плоды, в которых обычно есть семена.
После процесса оплодотворения начинается развитие эмбриона. Зигота делится на две клетки, а именно верхнюю клетку или терминальную клетку и нижнюю клетку или базальную клетку. Диссоциация базальной клетки приводит к суспензору, который в конечном итоге создает связь с материнской тканью. Суспензор предлагает путь для передачи пищи развивающемуся зародышу от родительского растения.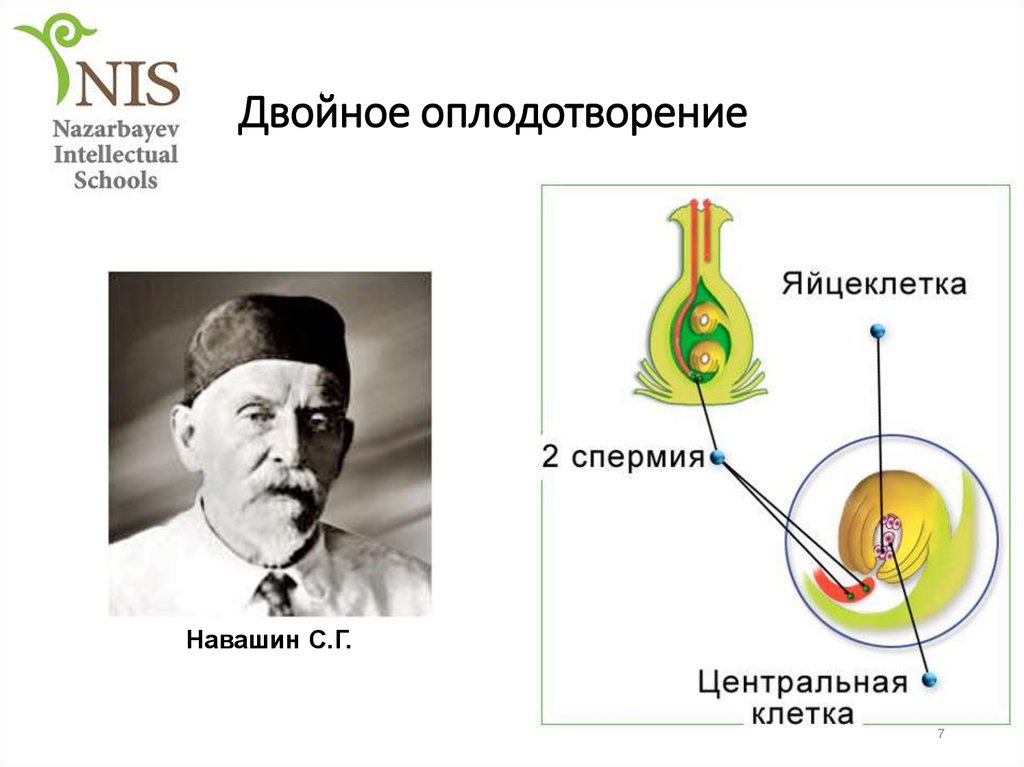
Терминальная клетка также диссоциирует, в результате чего образуется проэмбрион в форме земли. В случае эвдикотов или двудольных форма растущего зародыша похожа на сердце из-за наличия двух рудиментарных семядолей. В случае неэндоспермных двудольных, включая Capsella bursa, эндосперм растет на начальной стадии, но затем переваривается. В этом случае запасы питательных веществ переносятся в две семядоли.
Семядоли и зародыши скапливаются внутри растущего семени и вынуждены скручиваться по мере увеличения размера. В конце концов, семядоли и зародыш полностью занимают семя, в течение которого семя становится доступным для рассеивания. Развитие зародыша останавливается через определенный промежуток времени и возобновляется только тогда, когда семя прорастает. Эволюция проростка зависит от запасов питательных веществ, хранящихся в семядолях, если только начальная пара листьев не начнет фотосинтез.
Двойное оплодотворение у покрытосеменных
Покрытосеменные относятся к цветоносной флоре и представляют собой максимально разнообразные институты наземной растительной жизни.
Цветки образуют репродуктивную часть покрытосеменных с отдельными мужскими и женскими репродуктивными органами. каждая несет гаметы – сперматозоиды и яйцеклетки соответственно.Опыление помогает пыльцевым зернам достичь стигмы через стиль. 2 спермия входят в яйцеклетку-синергид. Это переходит к оплодотворению.
В результате оплодотворения образуются две структуры, в частности, зигота и эндосперм, по этой причине название «двойное оплодотворение».
Двойное оплодотворение представляет собой сложный процесс, при котором одна из сперматозоидов сливается с яйцеклеткой, а другая сливается с двумя полярными ядрами, что приводит к образованию диплоидной (2n) зиготы и триплоидного (3n) ядра эндосперма номер один (PEN) соответственно.
Учитывая, что эндосперм образуется в результате слияния трех гаплоидных ядер, такое слияние называется тройным.
В конечном итоге ядро эндосперма номер один превращается в подвижный эндосперм номер один (pc), а затем в эндосперм.Зигота станет эмбрионом после нескольких клеточных делений.
Эмбрион начинает расти, становится тесно и начинает изгибаться.
На заключительном этапе зародыш полностью заполняет семя.
 Цветки образуют репродуктивную часть покрытосеменных с отдельными мужскими и женскими репродуктивными органами. каждая несет гаметы – сперматозоиды и яйцеклетки соответственно.
Цветки образуют репродуктивную часть покрытосеменных с отдельными мужскими и женскими репродуктивными органами. каждая несет гаметы – сперматозоиды и яйцеклетки соответственно.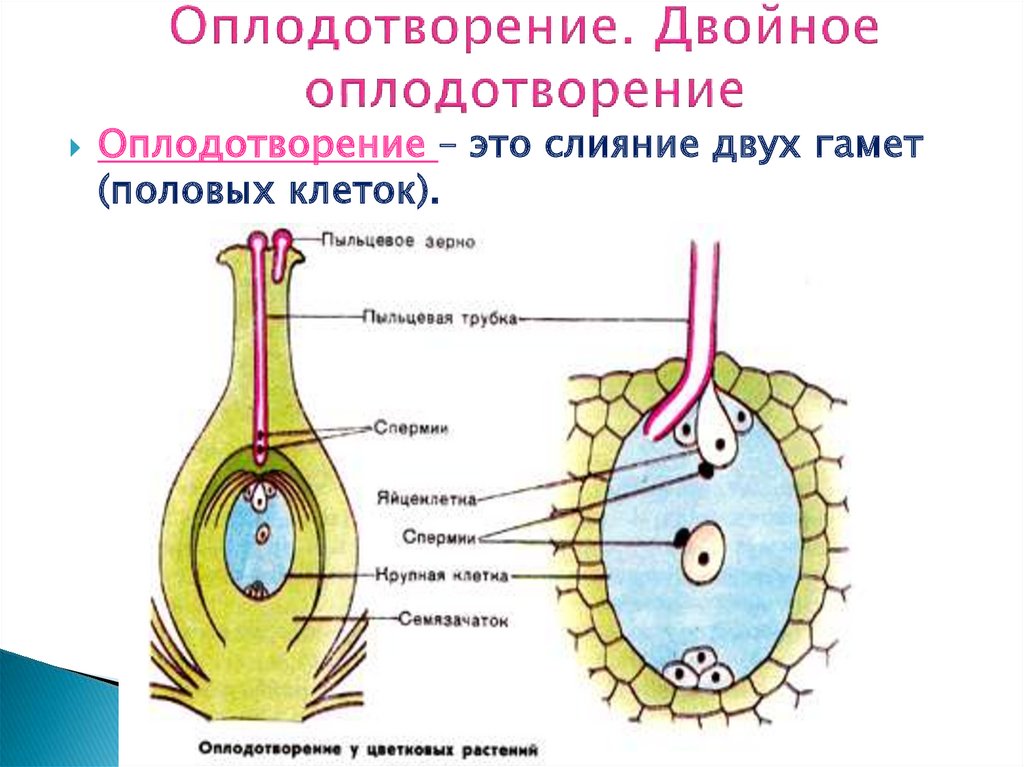 В конечном итоге ядро эндосперма номер один превращается в подвижный эндосперм номер один (pc), а затем в эндосперм.
В конечном итоге ядро эндосперма номер один превращается в подвижный эндосперм номер один (pc), а затем в эндосперм.Развитие зародыша покрытосеменных растений
В тот момент, когда завершается процесс оплодотворения, начинается развитие зародыша, и никакие другие сперматозоиды не могут пройти через яичник. Оплодотворенная яйцеклетка вырастает в семя, а ткани завязи развиваются в виде мясистого плода, окружающего семя.
(Изображение будет загружено в ближайшее время)
После процесса оплодотворения зигота разделяется на верхнюю концевую клетку и нижнюю базальную клетку. Последний превращается в подвеску, которая помогает транспортировать пищу к развивающемуся эмбриону. Верхняя терминальная клетка превращается в проэмбрион.
Последний превращается в подвеску, которая помогает транспортировать пищу к развивающемуся эмбриону. Верхняя терминальная клетка превращается в проэмбрион.
Фазы развития зародыша покрытосеменных растений
В развитии зародыша участвуют следующие фазы:
В начальной фазе развития терминальная клетка диссоциирует, образуя проэмбрион шаровидной формы. Кроме того, базальная клетка также распадается на суспензор.
Развивающийся зародыш приобретает сердцевидную форму благодаря наличию семядолей.
Развивающийся эмбрион становится перегруженным и начинает приспосабливаться к различным формам.
Зародыш занимает все пространство семени.
Значение двойного оплодотворения
Значение двойного оплодотворения у покрытосеменных можно уточнить в следующих пунктах:
Так как существует только два слияния; образуются только два продукта.
Вторичный продукт оплодотворения — триплоидное ядро первичного эндосперма (PEN) — превращается в питательную ткань, известную как эндосперм, которая отвечает за обеспечение питания развивающегося эмбриона.
Диплоидное состояние в жизненном цикле сохраняется у продукта процесса синтеза. Диплоидная зигота превращается в зародыш, из которого, как следствие, вырастает новое растение.
Процесс двойного оплодотворения заключается в использовании обеих мужских гамет, образованных пыльцевым зерном.
Таким образом, шансы полиэмбриологии увеличиваются, что также приводит к увеличению шансов на выживание нового растения.
Двойное оплодотворение стимулирует растения, отвечающие за развитие завязи в плоды и завязи в семена.