Содержание
Двойное оплодотворение цветковых — что это, определение и ответ
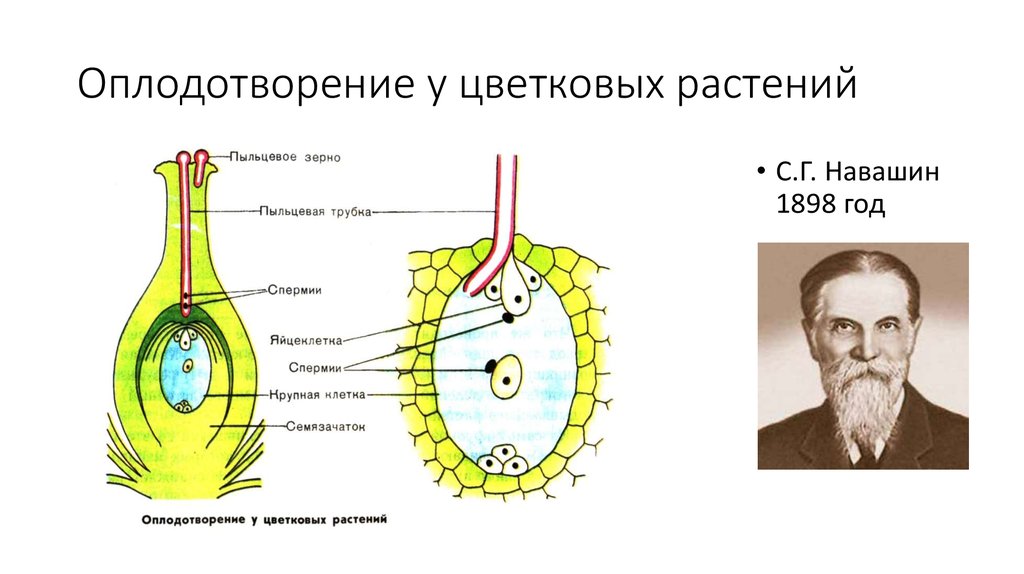
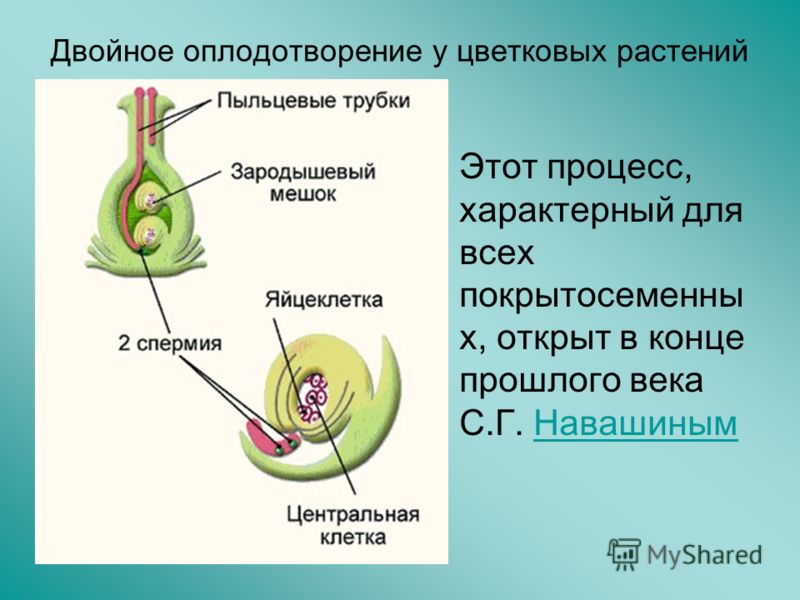
Двойное оплодотворение цветковых растений – особый тип оплодотворения, который наблюдается только у цветковых растений. Процесс двойного оплодотворения был открыт на примере лилейных и детально изучен отечественным ботаником Сергеем Гавриловичем Навашиным в 1898 г.
Оплодотворению у покрытосеменных предшествует микро — и мегаспорогенез.
Диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит – пыльцевое зерно. В пыльцевом зерне имеются две клетки: вегетативная и генеративная.
Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в итоге остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса, формируя зародышевый мешок. Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На одном полюсе находятся две синергиды и одна – яйцеклетка. На другом полюсе располагаются антиподы. Таким образом, зрелый зародышевый мешок содержит 7 клеток.
После первого деления два дочерних ядра расходятся к разным полюсам и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На одном полюсе находятся две синергиды и одна – яйцеклетка. На другом полюсе располагаются антиподы. Таким образом, зрелый зародышевый мешок содержит 7 клеток.
Двойное оплодотворение = первый спермий сливается с яйцеклеткой + второй спермий сливается с центральной клеткой.
Попав на рыльце пестика, пыльца прорастает. При прорастании пыльцы образуется пыльцевая трубка, растущая по столбику вниз к семяпочке. Кончик пыльцевой трубки выделяет ферменты, растворяющие клетки столбика, так что создается возможность для дальнейшего ее прорастания. Ядро трубки остается в кончике растущей пыльцевой трубки.
Генеративное ядро мигрирует в пыльцевую трубку и делится с образованием двух ядер – ядер спермиев.

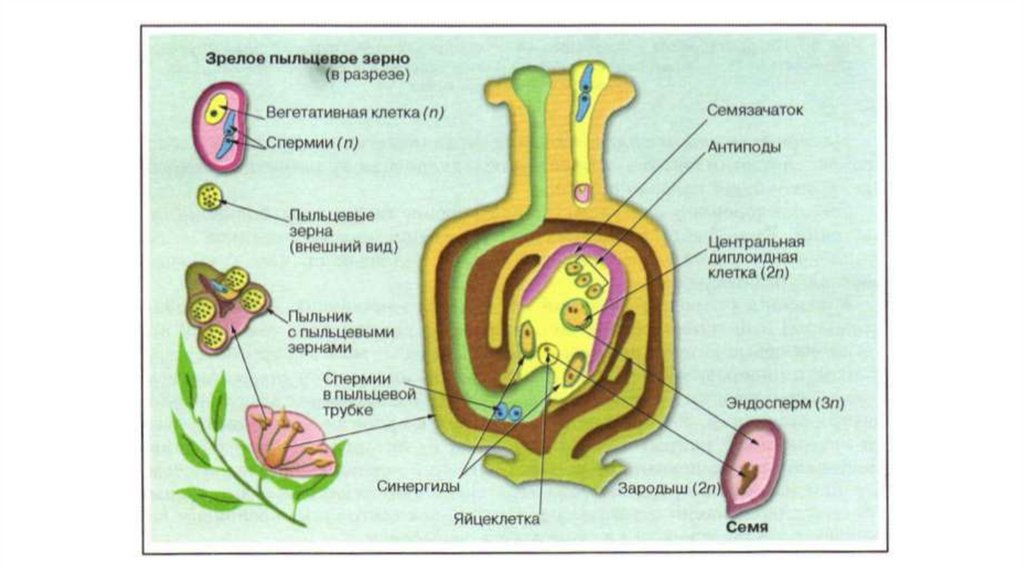
Зрелый мужской гаметофит состоит из пыльцевого зерна и пыльцевой трубки, ядра трубки и двух ядер спермиев, а также некоторого количества связанной с ними цитоплазмы.
Оба генеративных ядра проникают в макрогаметофит.
Одно из этих ядер перемещается к ядру яйцеклетки и сливается с ним; образующаяся в результате диплоидная зигота дает начало новому поколению спорофита.
Другое генеративное ядро перемещается к двум полярным ядрам, после чего все три ядра сливаются и образуют ядро эндосперма, содержащее тройной набор хромосом.
Внимание! Описанное явление двойного оплодотворения, приводящее к возникновению диплоидной зиготы и триплоидного эндосперма, специфично и характерно только для цветковых растений.
Момент оплодотворения
Развитие зародыша
После оплодотворения зигота многократно делится и формирует многоклеточный зародыш.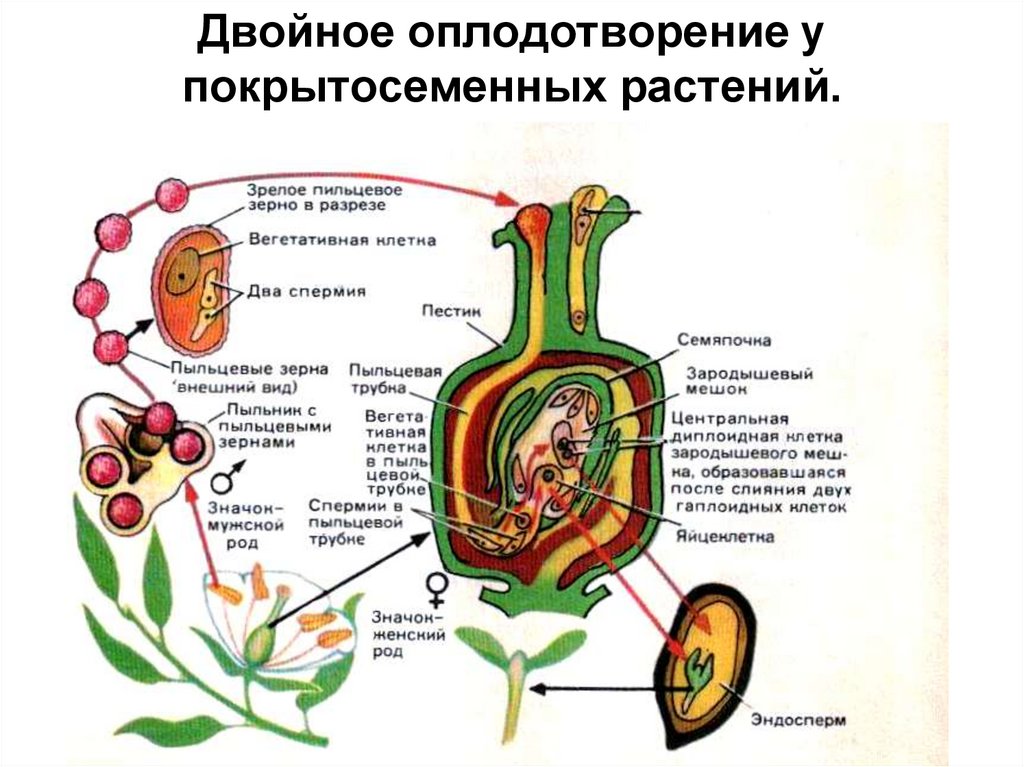 В результате делений ядра эндосперма образуются клетки эндосперма, наполненные питательными веществами. Эти клетки, окружающие зародыш, снабжают его питанием. После оплодотворения чашелистики, лепестки, тычинки, рыльце и столбик обычно завядают и опадают. Семяпочка вместе с содержащимся в ней зародышем превращается в семя; ее стенки утолщаются и превращаются в жесткие наружные покровы семени.
В результате делений ядра эндосперма образуются клетки эндосперма, наполненные питательными веществами. Эти клетки, окружающие зародыш, снабжают его питанием. После оплодотворения чашелистики, лепестки, тычинки, рыльце и столбик обычно завядают и опадают. Семяпочка вместе с содержащимся в ней зародышем превращается в семя; ее стенки утолщаются и превращаются в жесткие наружные покровы семени.
Семя состоит из зародыша и эндосперма с запасом питательных веществ, заключенных в прочную оболочку, которая возникла из стенки семяпочки.
Зародыш = корешок + стебелек + почечка + семядоля(-и)
Развитие зародыша
Оплодотворение и размножение у растений (биология, 10 класс)
3.9
Средняя оценка: 3.9
Всего получено оценок: 650.
3.9
Средняя оценка: 3.9
Всего получено оценок: 650.
Процесс слияния гамет (половых клеток) называется оплодотворением или копуляцией (сингамией).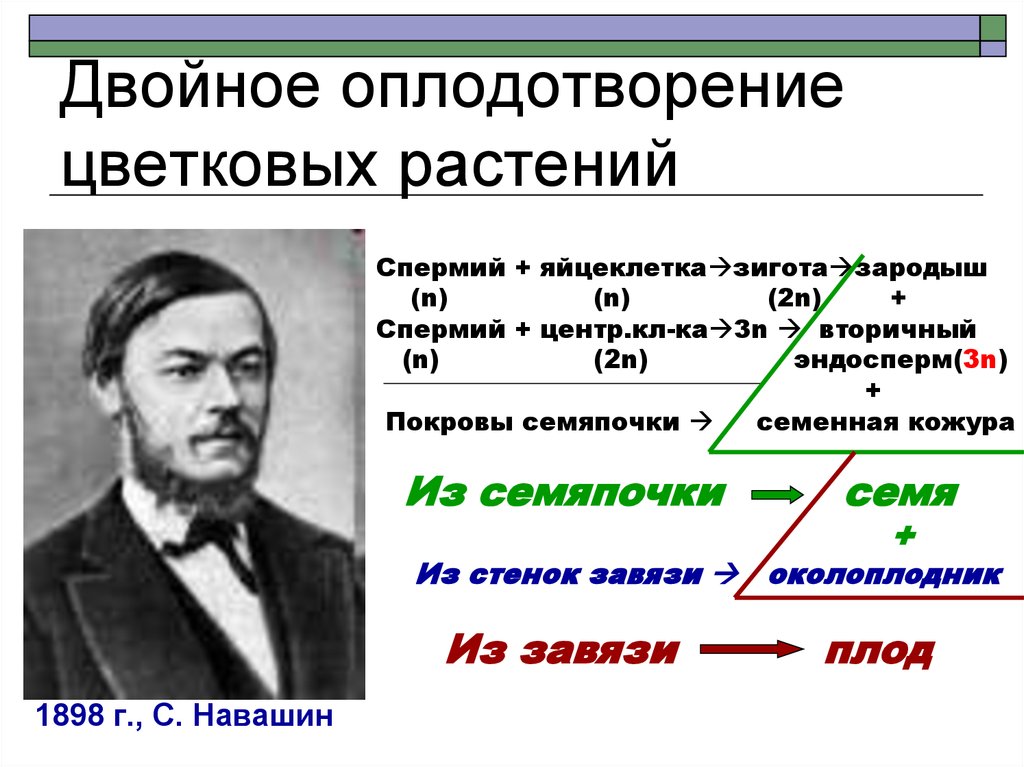 В результате такого слияния образуется зигота – первая клетка, из которой развивается новый организм. Оплодотворение происходит только при половом размножении.
В результате такого слияния образуется зигота – первая клетка, из которой развивается новый организм. Оплодотворение происходит только при половом размножении.
Виды
Оплодотворение характерно различным живым организмам – одноклеточным и многоклеточным растениям и животным. При слиянии двух гамет образуется организм с новым набором хромосом, обеспечивая тем самым генетическое разнообразие.
В зависимости от размера гамет различают три вида сингамии:
Рис. 1. Виды копуляции.
По способу оплодотворения выделяют два типа:
- наружное – происходит во внешней среде (обычно в воде), характерно кишечнополостным, морским червям, многим рыбам;
- внутреннее – перенос мужских гамет в женский организм, свойственный рептилиям, птицам, млекопитающим.
При наружном оплодотворении развитие зародышей происходит во внешней среде, при внутреннем – полностью или частично в утробе матери.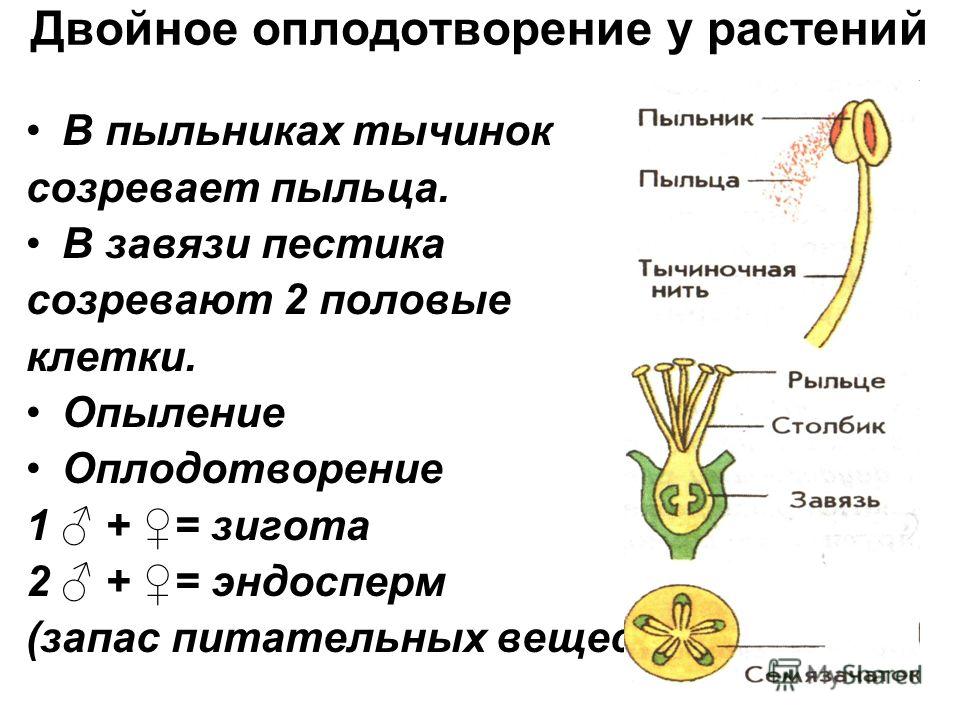 Например, для пресмыкающихся характерно внутренне оплодотворение, но наружное развитие зародышей (откладывание яиц).
Например, для пресмыкающихся характерно внутренне оплодотворение, но наружное развитие зародышей (откладывание яиц).
Внутреннее
При внутреннем оплодотворении подвижная мужская гамета (сперматозоид) достигает неподвижной женской гаметы (ооцита) в половых путях самки. Контакту клеток способствуют специальные вещества, выделяемые ооцитом. Акросома – органелла, находящаяся в передней части сперматозоида, – выделяет ферменты, разрушающие оболочку ооцита.
После попадания сперматозоида внутрь формируется плотная оболочка, закрывающая доступ другим сперматозоидам. В это время ооцит завершает второе деление мейоза в процессе оогенеза (развития женской клетки) и превращается в яйцеклетку.
Ядро мужской гаметы увеличивается до размера ядра яйцеклетки. Постепенно ядра сближаются и сливаются в единое ядро. Образуется зигота.
Рис. 2. Слияние сперматозоида и яйцеклетки.
Гаметы несут гаплоидный (одинарный) набор хромосом. После оплодотворения образуется клетка с диплоидным (двойным) набором хромосом.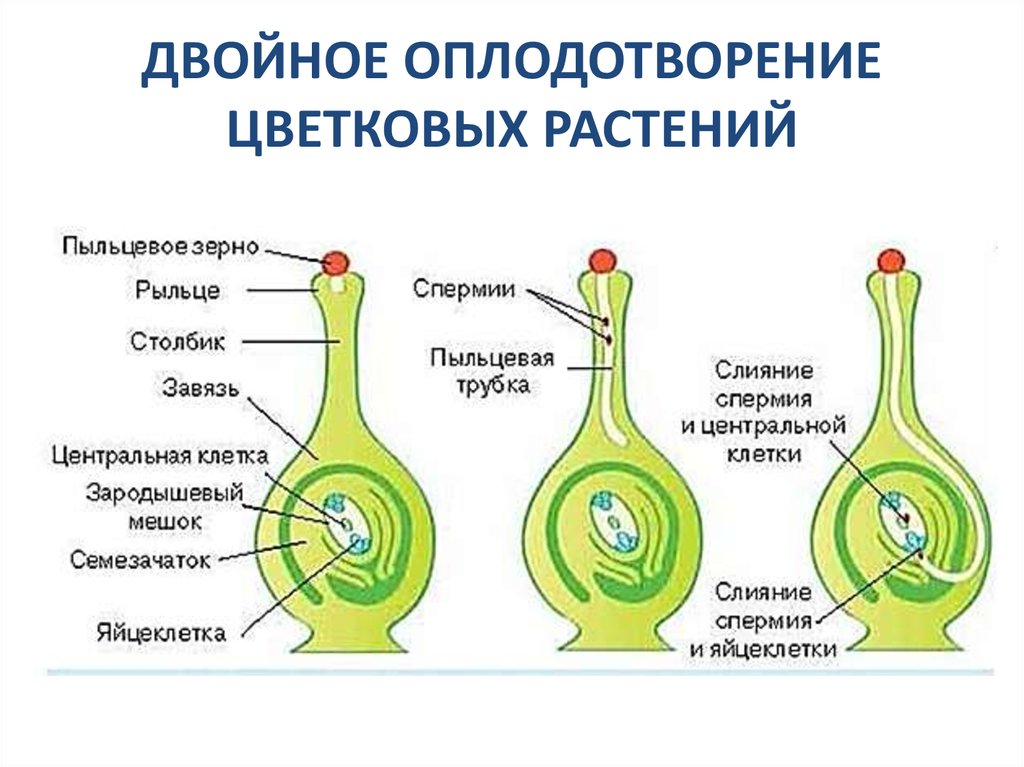
Двойное
Особый способ размножения и оплодотворения у растений носит название двойного оплодотворения. Это процесс, при котором одновременно образуются зигота и эндосперм. Двойное оплодотворение характерно только для цветковых (покрытосеменных) растений.
Процесс двойного оплодотворения состоит из четырёх этапов:
- формирование гамет – в пыльниках развиваются мужские спермии, в семязачатках завязи – яйцеклетки;
- образование дополнительных клеток – вместе с яйцеклеткой формируются центральная клетка, образующая центральное ядро, и пять маленьких клеток;
- попадание спермия в семязачаток – попавшие вместе с пыльцой на рыльце пестика два спермия проходят по пыльцевой трубке в семязачаток;
- оплодотворение – один спермий сливается с яйцеклеткой, образуя зиготу, второй – с центральным ядром, формируя эндосперм.
Рис. 3. Двойное оплодотворение.
Зигота при двойном оплодотворении имеет диплоидный набор хромосом.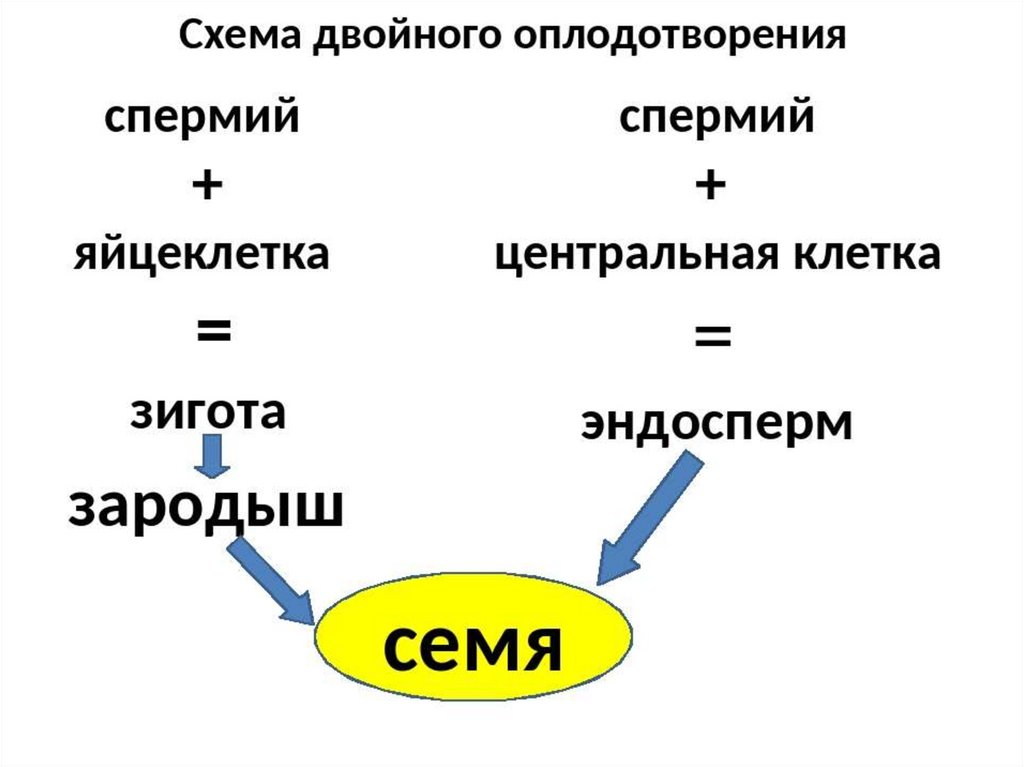 При слиянии спермия с центральным ядром образуется клетка с триплоидным набором хромосом.
При слиянии спермия с центральным ядром образуется клетка с триплоидным набором хромосом.
Что мы узнали?
Из урока биологии 10 класса узнали о способах и значении оплодотворения. В теме раскрыты понятия изогамии, анизогамия, оогамии, наружного и внутреннего оплодотворения, а также двойного оплодотворения. В процессе сингамии, копуляции или оплодотворения образуется новый, не похожий на родителей организм с уникальным генотипом.
Тест по теме
Доска почёта
Чтобы попасть сюда — пройдите тест.
Никита Метальников
10/10
Максим Шишигин
9/10
Оценка доклада
3.9
Средняя оценка: 3.9
Всего получено оценок: 650.
А какая ваша оценка?
Двойное оплодотворение
Двойное оплодотворение
После просмотра анимации перейдите к викторине ниже.
Введение
Просмотр/печать сценария
Заключение
Вопрос
1.
 1
1
35/qhnixosl9/ETxvNyn63IYJe+I5R0aAyOdoRwCc/0fUahWsxsMBhf4t1/MLBm9fDb8qfruaZ/6qvbfWx8RHPTfGhOPKoM06xSbb5kFHRuSFG5eaESomB0lkEMDPGbLRtXLa918i58Sq2mwU7dRPjSFmMHKgTNpkSGByb7/9ifWDDwPh4p2adIOwpk4gVtvneNBFaVQgYv5lYzZwXgcRCQpRhjx48kSoBQX+Y6TMy5p/NxAXWntuTHuAOCDFSC3MQecyG+XgmSf1EIp4b3z94Hvpmu8Nkl8lH+nC+2xmDgk2UjtM0vxassI+IUS9eJxHhd4t2gEwTkZBr+4v4Y94QSnlJsvtReaRUSAPg==
Правильно. См. ключевое понятие 37.1 Большинство покрытосеменных размножаются половым путем
Неверно. См. ключевое понятие 37.1 Большинство покрытосеменных размножаются половым путем
Вопрос
1,2
oCf+PrNA15WJABoXAdq1SQgLypqXRFgRkNMH/Z4oBFJE1W3yb0izP3E6JfvOD1uu4qwSFK7/AklcQeSxUG5Ycvga+0XDJSf1CLkvMldZOilBN4uqe1IOayYEkCUSJ8pQn9N65LJ8diPGMwPboPBhkQQI/6O1SBwxOjT/hq55n+Wdma3hCQuig09yLntqJw7SwBZndKGOvBEdfR1YuVy/sgprFh6ZQGeUp0As9rqFMUrYacZNp9v/sEPey0F3lBqshV7m/u8byo7IahHewq929zTzYZm5RYLyXAv0kwWVhSS6RY3UCx8s3DJN5155FWqeoACgvAvv/OvzOs6CydVfHzik3Pa+JUm7VwP/dzkSYmt35lmbENaI1OYyILEmMWvPzTYHRB7X77MKwG8iEOqIUco34rLrp0fNJiaRsnKT75UP57ckEEr2iNJVSIWcDSrNB5UVIGpESGhg01kgSQxxNpshWJmjCpirbAcqigwVarYLNbHLzBA3bQVIKaOqFqKDZcHzd1KdNkdKTkDtmJbhntj61an55cw+kOErmG7VZ84ebdbDUp8F0ojkz1fK99zk5ZlfRVXcgjC18YKpq
Правильно.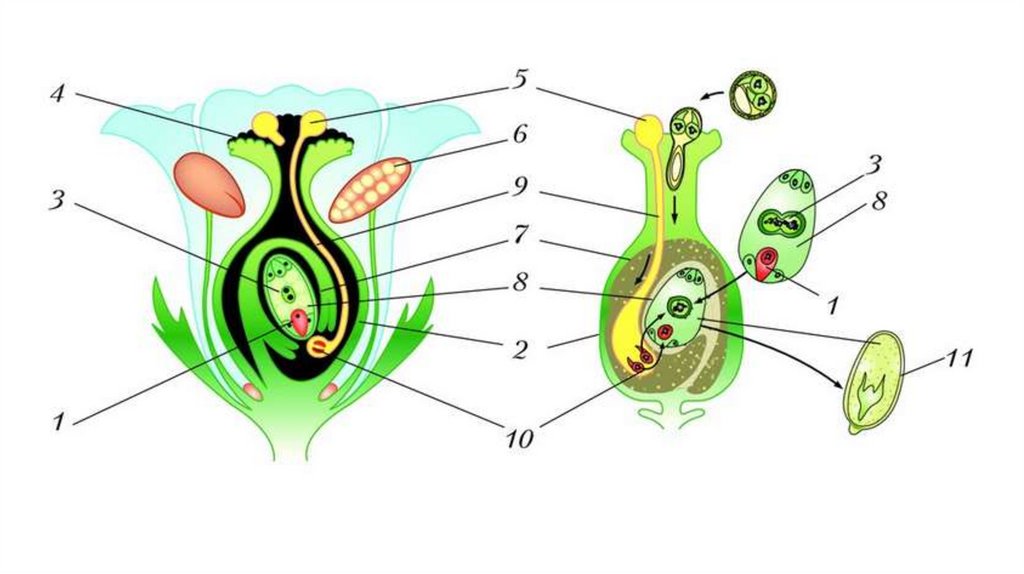 См. ключевую концепцию 37.1 Большинство покрытосеменных размножаются половым путем
См. ключевую концепцию 37.1 Большинство покрытосеменных размножаются половым путем
Неверно. См. ключевое понятие 37.1 Большинство покрытосеменных размножаются половым путем
Вопрос
1,3
KizudJX2Ov2z6RneFYTbHBpYERjOgyE800opOA5qR1uo0hd7Srlc8O39SPUUqPXejwFfJRUnKnL8YfV+Q5rIYEd1uGpEqxyYT3do+u0xx8NeM1rXftZ2rncnTgEpip5SCp7BA83xmfSVo4u96gGVH8CI+FqUN16mmKXWRD1aF9W3GubL17LvbsQFUAwDfrzkY4K4sgMzKr27Pyg8wFfZ1FCr71f+Cz6Ld9w7F2/zwPRH8bCkLWqVEbM2/r9TGQ5x0okjyshq5nQ=
Правильно. См. ключевую концепцию 37.1 Большинство покрытосеменных размножаются половым путем
Неверно. См. ключевое понятие 37.1 Большинство покрытосеменных размножаются половым путем
Вопрос
1,4
ASsyAtGEqSt1G+mVeJ7ntXW2nhlujqFWFmHlAPlxZ6EUULvYYA2QM2YJ/5oJefaaSjIMX/awGHCvkao9MRwWMQL1wSpDBT2Qd67bsZMcD9Wa7ONl+LKcatv+gSqqmv5o1v+e6AJVxYLdH5k553IgsCYEDgzX0IUXmVvarHwBFE39Xseko751OUlhuJRjeXBGXx0wpaVGgNcT4pRydLcOqSIRuzix0Uiqp8WsMJ1WTF5x6a1hUJ+2CKSRKQ6LnPvUv1zFDMOaxrLcDom4nOl72ePZXd8M9cWcEN0FKsdlV8jqhoxnso4pi9IOFkLfU116EA8G0ZvRoBceDXG3WPlQHdXv/AWIGPzHxh7ydCPaATddxpmb0JvtVIspPZbtde/uJqWmPC7Tm7mSkHxNrf7MdUYZSrXSpV9LwoyWpQ7sbFlXiA+5xyLFSAHJ+RNUcuyeRecfNPmsB0EeHv+NQYpuijp5/RQBUDKL1YcsN+wUJJVV+4GxYnrG5BGJVu3rNjxmUs9OHjm1/i6moMgaBTLtxPzBr+Gdniqc7gw1exMCIm0dLxlezeuqr4MAQRjxsKo0foAoyfxBWh3YYwTIrmVUDpprEEgoXEXYGBBD0g4cbbgEs20/BxQxMqBG6uMPWfbpcik48xjuxsz35V0o/b/9/iKc0DLiJKx06jNzswN+8aePH/icwNN0tZXWksMPmwxvsKuKz0DDU7MlP190PQXTQeB/T2zKdLt3jg7ya7KyafwIfao1Kp8KtA==
Правильно.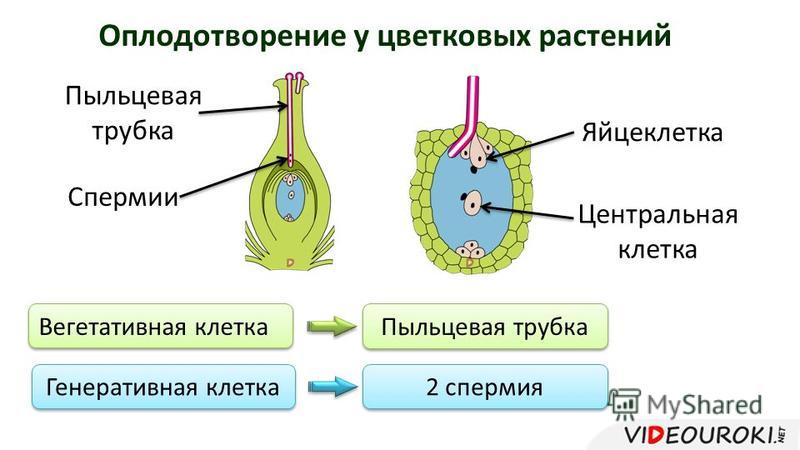 См. ключевое понятие 37.1 Большинство покрытосеменных размножаются половым путем
См. ключевое понятие 37.1 Большинство покрытосеменных размножаются половым путем
Неверно. См. ключевую концепцию 37.1 Большинство покрытосеменных размножаются половым путем
Результаты деятельности отправляются…
Гиногенез in vivo у кукурузы: новое понимание двойного оплодотворения растений
Гиногенез in vivo у кукурузы: новое понимание двойного оплодотворения растений | ANR
подать предложение
Нажимать
Присоединяйтесь к нам
CE20 — Биология животных, фотосинтез организмов и микроорганизмов
Как сделать детей «не похожими на папу»?
Отклонение от классического процесса двойного оплодотворения высших растений индуцирует у своеобразных линий кукурузы гаплоидные зародыши, содержащие только материнский геном. Этот проект направлен на использование этой уникальной линии кукурузы в качестве инструмента для лучшего понимания полового размножения растений.
Чтобы дать новое представление о механизме гаплоидной индукции кукурузы
Половое размножение является ключом к эволюционному успеху высших эукариот.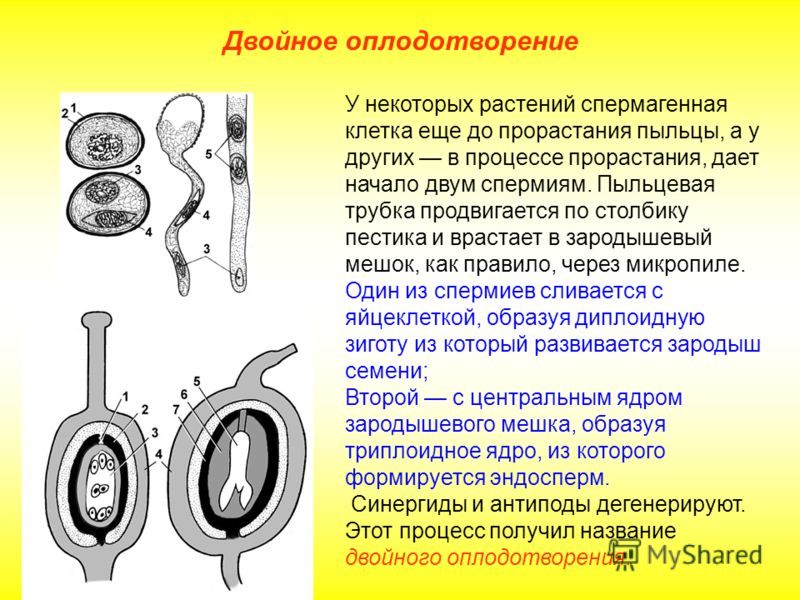 У всех цветковых растений половое размножение характеризуется уникальным биологическим процессом, называемым двойным оплодотворением. Двойное оплодотворение состоит из двух отдельных параллельных актов слияния между двумя сперматозоидами и двумя женскими гаметами, яйцеклеткой и центральной клеткой, в результате чего возникает зародыш и питательная ткань эндосперма соответственно. Линия кукурузы, называемая гаплоидной индукторной линией, отличается от этого классического процесса двойного оплодотворения: вместо нормальных диплоидных зародышей (2n) она индуцирует образование гаплоидных зародышей (1n), содержащих исключительно материнские хромосомы. Этот процесс известен как гиногенез in vivo. Производные этой гаплоидной индукторной линии кукурузы стали предпочтительным инструментом многих селекционных компаний, поскольку они могут производить совершенно гомозиготные растения всего за 2 поколения вместо 5-8 в классических схемах селекции.
У всех цветковых растений половое размножение характеризуется уникальным биологическим процессом, называемым двойным оплодотворением. Двойное оплодотворение состоит из двух отдельных параллельных актов слияния между двумя сперматозоидами и двумя женскими гаметами, яйцеклеткой и центральной клеткой, в результате чего возникает зародыш и питательная ткань эндосперма соответственно. Линия кукурузы, называемая гаплоидной индукторной линией, отличается от этого классического процесса двойного оплодотворения: вместо нормальных диплоидных зародышей (2n) она индуцирует образование гаплоидных зародышей (1n), содержащих исключительно материнские хромосомы. Этот процесс известен как гиногенез in vivo. Производные этой гаплоидной индукторной линии кукурузы стали предпочтительным инструментом многих селекционных компаний, поскольку они могут производить совершенно гомозиготные растения всего за 2 поколения вместо 5-8 в классических схемах селекции.
Спустя пятьдесят лет после открытия первой линии индуктора гаплоидов в нашей работе 2017 г.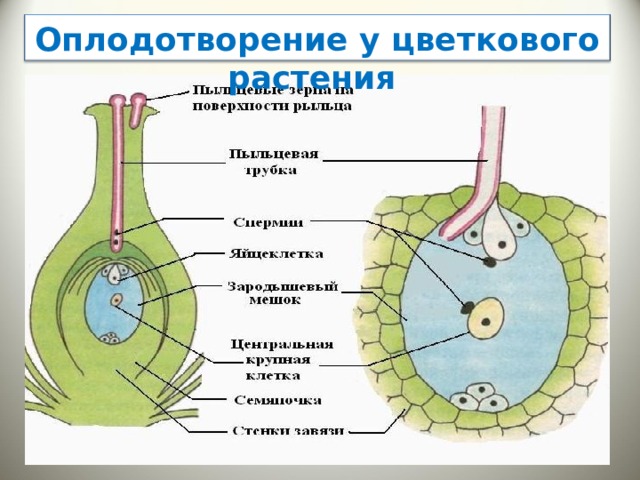 был выявлен основной ген, ответственный за индукцию гаплоидов (Gilles et al., 2017, EMBO Journal; Gilles et al., 2017, Current Biology; Martinant et al. и др., 2016 г., патент WO/2016/177887). Продукт гена кодирует фосфолипазу типа А2, которую мы назвали NOT LIKE DAD (NLD), поскольку мужской геном отсутствует у гаплоидных эмбрионов. Используя комбинацию генетики, геномики и клеточной биологии, мы продемонстрировали, что интактная локализованная в сперматозоидах фосфолипаза необходима для успешного полового размножения. Этот проект направлен на решение вопроса о том, почему белок NLD, связывающий мембрану сперматозоида, необходим для поддержания целостности отцовского генома у эмбрионов. Мы хотели бы понять, как целостность отцовского генома поддерживается во время нормального двойного оплодотворения, как биохимическая функция NLD связана с целостностью отцовского генома и, в более общем плане, как координируется двойное оплодотворение.
был выявлен основной ген, ответственный за индукцию гаплоидов (Gilles et al., 2017, EMBO Journal; Gilles et al., 2017, Current Biology; Martinant et al. и др., 2016 г., патент WO/2016/177887). Продукт гена кодирует фосфолипазу типа А2, которую мы назвали NOT LIKE DAD (NLD), поскольку мужской геном отсутствует у гаплоидных эмбрионов. Используя комбинацию генетики, геномики и клеточной биологии, мы продемонстрировали, что интактная локализованная в сперматозоидах фосфолипаза необходима для успешного полового размножения. Этот проект направлен на решение вопроса о том, почему белок NLD, связывающий мембрану сперматозоида, необходим для поддержания целостности отцовского генома у эмбрионов. Мы хотели бы понять, как целостность отцовского генома поддерживается во время нормального двойного оплодотворения, как биохимическая функция NLD связана с целостностью отцовского генома и, в более общем плане, как координируется двойное оплодотворение.
В этом проекте мы предлагаем ряд скоординированных мероприятий для изучения и разъяснения этих увлекательных аспектов воспроизводства растений.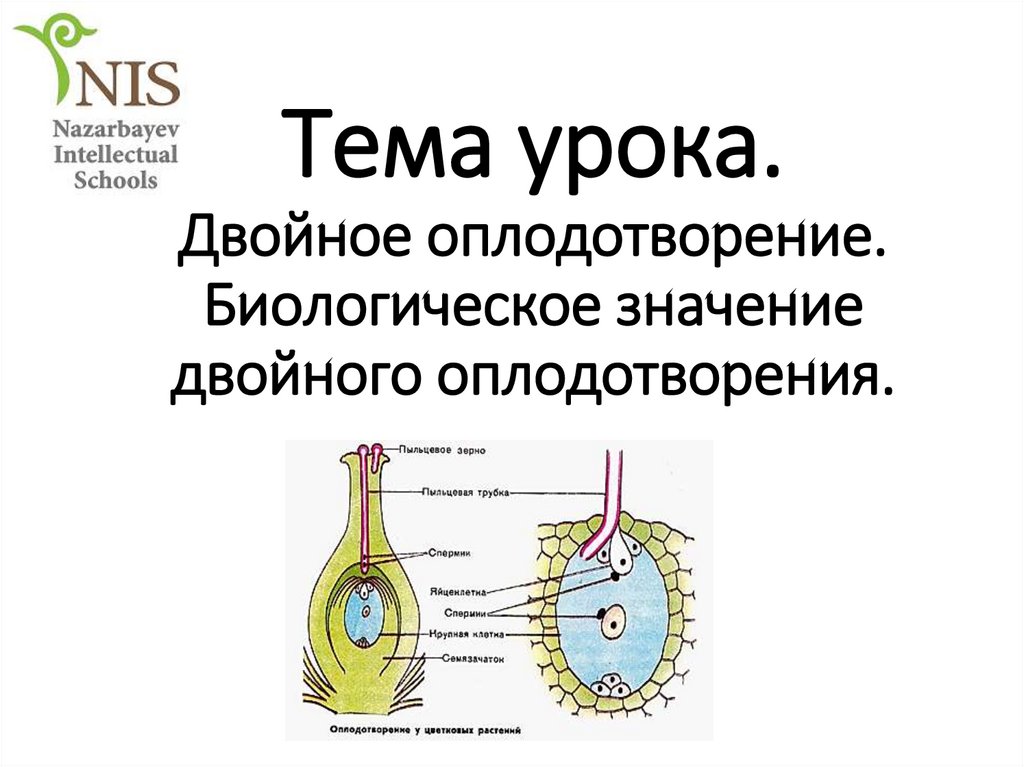 Визуализация двойного оплодотворения с маркерными линиями ядерной и плазматической мембраны будет использоваться для исследования дефектов оплодотворения в мутантных линиях nld. Мы оценим время, степень и специфичность фрагментации мужского генома в сперматозоидах, гиногенетических эмбрионах и эндоспермах. Биохимическая характеристика активности фосфолипазы NLD in vitro в сочетании со сравнительным анализом липидного состава пыльцы in vivo прольет свет на субстраты/продукты NLD и прояснит их биологическую функцию. Наконец, частично на основе существующего интерактома и частично на генетических подходах будут идентифицированы и функционально охарактеризованы новые молекулярные игроки, связанные с NLD.
Визуализация двойного оплодотворения с маркерными линиями ядерной и плазматической мембраны будет использоваться для исследования дефектов оплодотворения в мутантных линиях nld. Мы оценим время, степень и специфичность фрагментации мужского генома в сперматозоидах, гиногенетических эмбрионах и эндоспермах. Биохимическая характеристика активности фосфолипазы NLD in vitro в сочетании со сравнительным анализом липидного состава пыльцы in vivo прольет свет на субстраты/продукты NLD и прояснит их биологическую функцию. Наконец, частично на основе существующего интерактома и частично на генетических подходах будут идентифицированы и функционально охарактеризованы новые молекулярные игроки, связанные с NLD.
Первым основным результатом этого проекта является новая субклеточная локализация NLD за пределами сперматозоидов на уникальной мембране. Действительно, вопреки предыдущим сообщениям, мы продемонстрировали, что NLD локализуется не в цитозоле и плазматической мембране сперматозоидов, а на специфической мембране, полученной из PM вегетативной клетки пыльцы, которая окружает две клетки спермы: эндо-PM.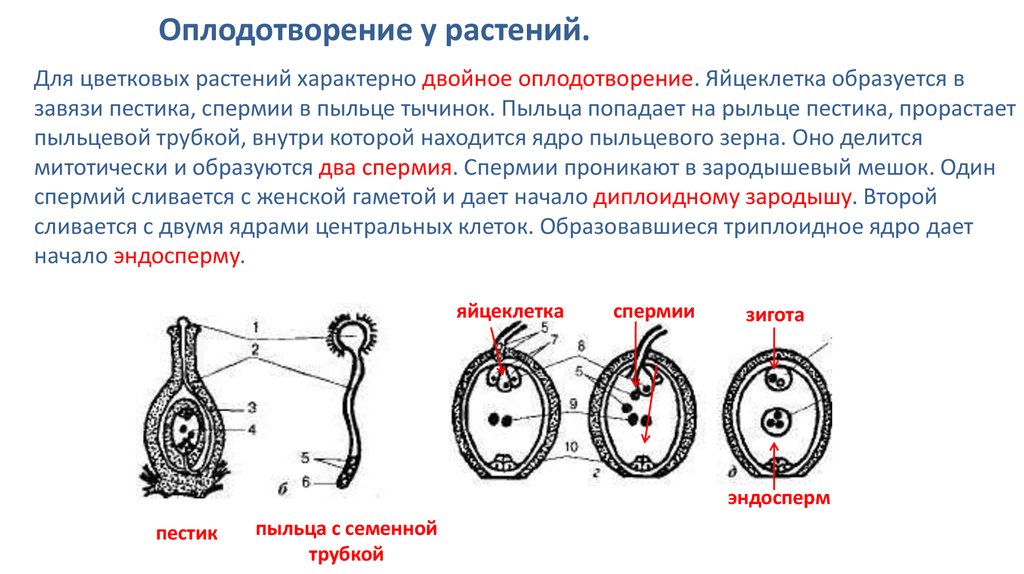 Мы дополнительно описываем перенос NLD и продемонстрировали, что липидное заякоривание и электростатические взаимодействия нацеливают NLD на эндо-PM пыльцы. Использование липидных биосенсоров позволяет нам расшифровать липидную сигнатуру этой эндо-ПМ пыльцы (https://doi.org/10.1083/jcb.202010077).
Мы дополнительно описываем перенос NLD и продемонстрировали, что липидное заякоривание и электростатические взаимодействия нацеливают NLD на эндо-PM пыльцы. Использование липидных биосенсоров позволяет нам расшифровать липидную сигнатуру этой эндо-ПМ пыльцы (https://doi.org/10.1083/jcb.202010077).
На данном этапе важным шагом проекта является выделение и характеристика новых генов, участвующих в гаплоидной индукции. Были отобраны гены-кандидаты, и CRISPR, и инсерционные мутанты находятся в стадии разработки и будут проанализированы в течение оставшегося времени проекта.
1. Жиль Л.М., Калхау А., Ла Падула В., Жакье НМА, Лионнет С., Мартинан Ж.-П., Роговски П., Видиз Т. (2021)
Липидное заякоривание и электростатические взаимодействия нацеливают NOT-LIKE-DAD на эндоплазматическую мембрану пыльцы. Журнал клеточной биологии, doi.org/10.1083/jcb.202010077.
2. Жакье НМА, Жиль Л.М., Пиотт Д.Е., Мартинан Дж.П., Роговский П.М., Видиз Т. (2020).
Разгадка репродукции растений путем гаплоидной индукции для инноваций в селекции растений.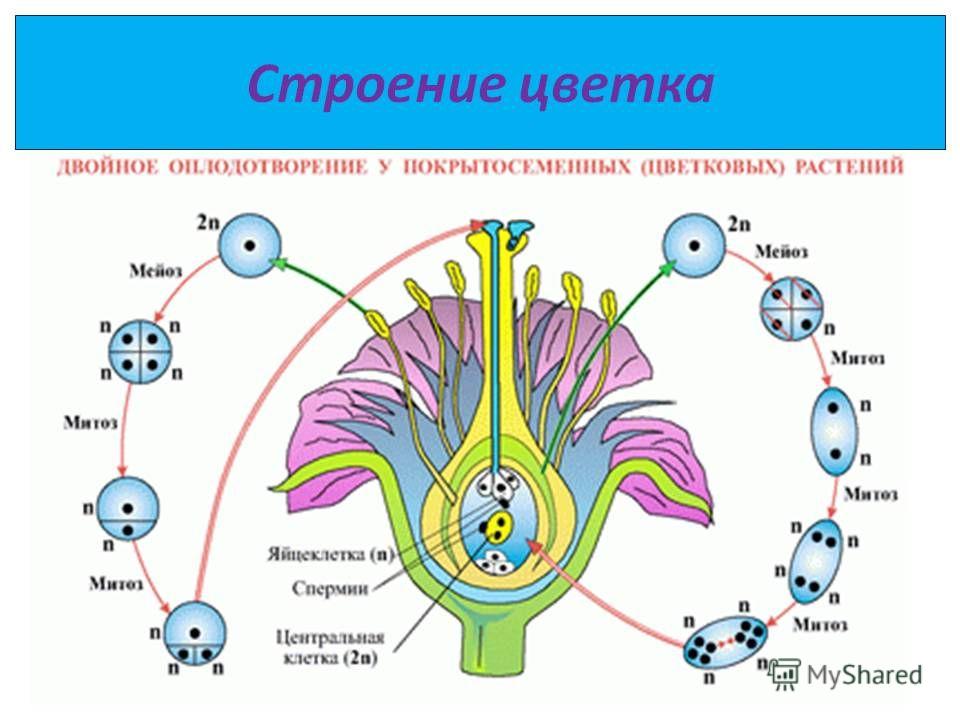 Природные растения, 6(6):610-619. doi.org/10.1038/s41477-020-0664-9
Природные растения, 6(6):610-619. doi.org/10.1038/s41477-020-0664-9
3. Сеги-Симарро Дж. М., Жакье НМА и Видиз Т. (2021 г.)
Обзор технологий удвоения гаплоидов in vitro и in vivo. Методы молекулярной биологии (Springer), doi.org/10.1007/978-1-0716-1315-3_1
4. Жакье НМА, Жиль Л.М., Мартинан Дж.П., Роговский П.М. и Видиз Т. (2021)
Кукуруза в линиях индукции гаплоидных плантаций, краеугольный камень технологии двойных гаплоидов. Methods in Molecular Biology (Springer), doi.org/10.1007/978-1-0716-1335-1_2
Половое размножение является ключом к эволюционному успеху высших эукариот. У всех цветковых растений половое размножение характеризуется уникальным биологическим процессом, называемым двойным оплодотворением. Двойное оплодотворение состоит из двух отдельных параллельных актов слияния между двумя сперматозоидами и двумя женскими гаметами, яйцеклеткой и центральной клеткой, в результате чего возникает зародыш и питательная ткань эндосперма соответственно.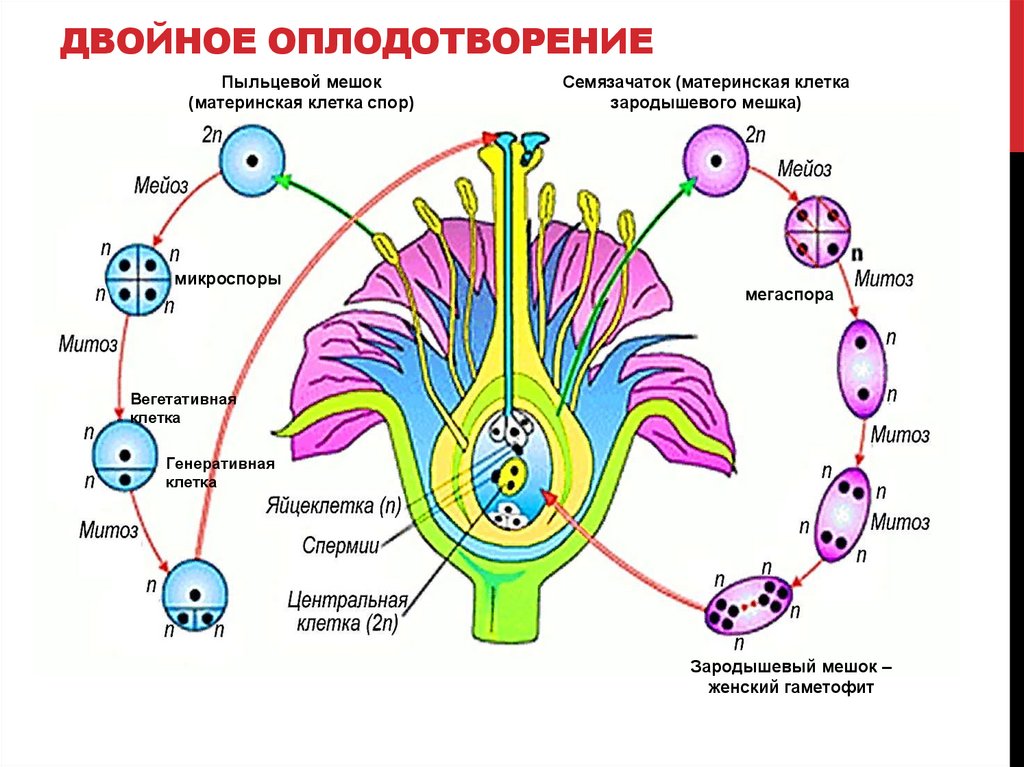 Линия кукурузы, называемая гаплоидной индукторной линией, отличается от этого классического процесса двойного оплодотворения: вместо нормальных диплоидных зародышей (2n) она индуцирует образование гаплоидных зародышей (1n), содержащих исключительно материнские хромосомы. Этот процесс известен как гиногенез in vivo. Производные этой гаплоидной индукторной линии кукурузы стали предпочтительным инструментом многих селекционных компаний, поскольку они могут производить совершенно гомозиготные растения всего за 2 поколения вместо 5-8 в классических схемах селекции.
Линия кукурузы, называемая гаплоидной индукторной линией, отличается от этого классического процесса двойного оплодотворения: вместо нормальных диплоидных зародышей (2n) она индуцирует образование гаплоидных зародышей (1n), содержащих исключительно материнские хромосомы. Этот процесс известен как гиногенез in vivo. Производные этой гаплоидной индукторной линии кукурузы стали предпочтительным инструментом многих селекционных компаний, поскольку они могут производить совершенно гомозиготные растения всего за 2 поколения вместо 5-8 в классических схемах селекции.
Через пятьдесят лет после открытия первой линии индуктора гаплоидов в нашей недавней работе был идентифицирован основной ген, ответственный за индукцию гаплоидов (Gilles et al., 2017, EMBO Journal; Gilles et al., 2017, Current Biology; Martinant et al. , 2016 г., патент WO/2016/177887). Продукт гена кодирует фосфолипазу типа А2, которую мы назвали NOT LIKE DAD (NLD), поскольку мужской геном отсутствует у гаплоидных эмбрионов.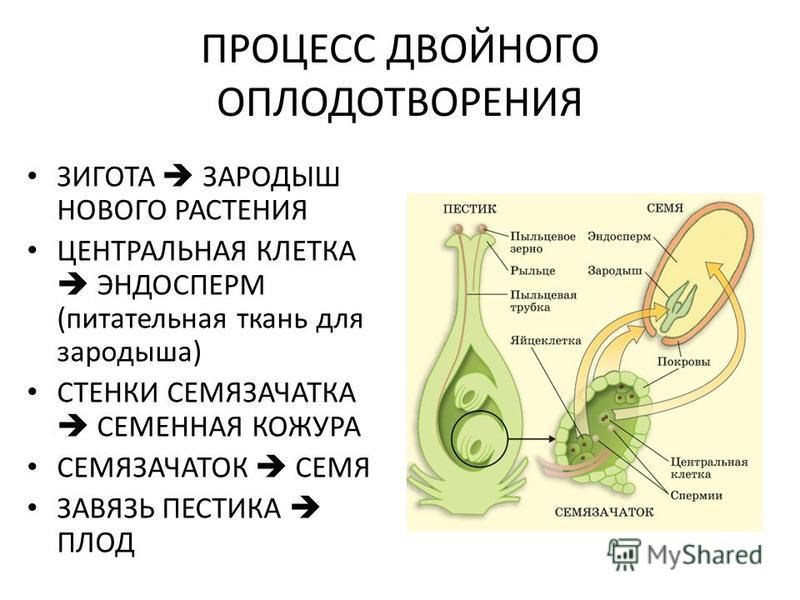 Используя комбинацию генетики, геномики и клеточной биологии, мы продемонстрировали, что интактная локализованная в сперматозоидах фосфолипаза необходима для успешного полового размножения. Этот проект направлен на решение вопроса о том, почему белок NLD, связывающий мембрану сперматозоида, необходим для поддержания целостности отцовского генома у эмбрионов. Мы хотели бы понять, как целостность отцовского генома поддерживается во время нормального двойного оплодотворения, как биохимическая функция NLD связана с целостностью отцовского генома и, в более общем плане, как координируется двойное оплодотворение.
Используя комбинацию генетики, геномики и клеточной биологии, мы продемонстрировали, что интактная локализованная в сперматозоидах фосфолипаза необходима для успешного полового размножения. Этот проект направлен на решение вопроса о том, почему белок NLD, связывающий мембрану сперматозоида, необходим для поддержания целостности отцовского генома у эмбрионов. Мы хотели бы понять, как целостность отцовского генома поддерживается во время нормального двойного оплодотворения, как биохимическая функция NLD связана с целостностью отцовского генома и, в более общем плане, как координируется двойное оплодотворение.
Опираясь на недавние результаты, мы предлагаем ряд скоординированных действий, которые позволят изучить и пролить свет на эти увлекательные аспекты воспроизводства растений. Визуализация двойного оплодотворения в режиме реального времени с маркерными линиями ядерной и плазматической мембран, а также анализы оплодотворения in vitro будут использоваться для исследования дефектов оплодотворения в мутантных линиях nld.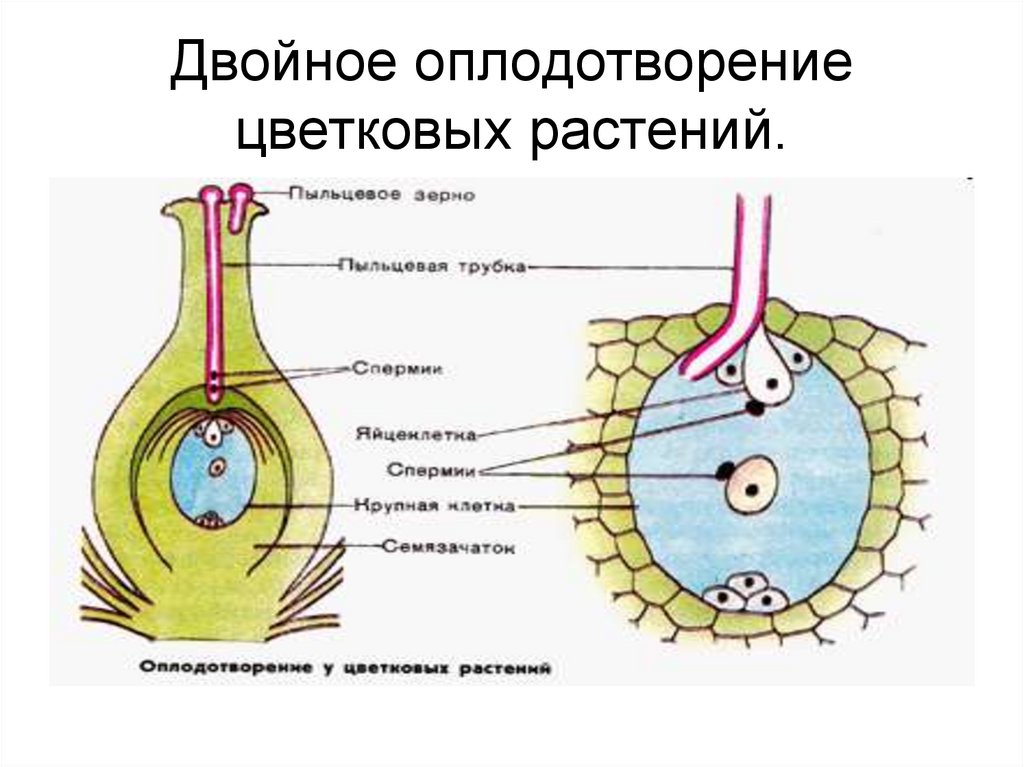 Мы оценим время, степень и специфичность фрагментации мужского генома в сперматозоидах, гиногенетических эмбрионах и эндоспермах. Биохимическая характеристика активности фосфолипазы NLD in vitro в сочетании со сравнительным анализом липидного состава пыльцы in vivo прольет свет на субстраты/продукты NLD и прояснит их биологическую функцию. Наконец, частично на основе существующего интерактома и частично на генетических подходах будут идентифицированы и функционально охарактеризованы новые молекулярные игроки, связанные с NLD. Вместе эти независимые и дополняющие друг друга эксперименты позволят по-новому взглянуть на механизм гаплоидной индукции кукурузы и в более общем плане ответить на фундаментальные вопросы о том, как координируется двойное оплодотворение растений.
Мы оценим время, степень и специфичность фрагментации мужского генома в сперматозоидах, гиногенетических эмбрионах и эндоспермах. Биохимическая характеристика активности фосфолипазы NLD in vitro в сочетании со сравнительным анализом липидного состава пыльцы in vivo прольет свет на субстраты/продукты NLD и прояснит их биологическую функцию. Наконец, частично на основе существующего интерактома и частично на генетических подходах будут идентифицированы и функционально охарактеризованы новые молекулярные игроки, связанные с NLD. Вместе эти независимые и дополняющие друг друга эксперименты позволят по-новому взглянуть на механизм гаплоидной индукции кукурузы и в более общем плане ответить на фундаментальные вопросы о том, как координируется двойное оплодотворение растений.
Monsieur Thomas WIDIEZ (ВОСПРОИЗВЕДЕНИЕ И РАЗВИТИЕ РАСТЕНИЙ)
Автором этого резюме является координатор проекта, который несет ответственность за содержание этого резюме. ANR не несет никакой ответственности за его содержание.