
Содержание
Почему кактусы не высыхают | Наука и жизнь
Что мы знаем о кактусах? Что у них есть колючки, что у них толстый стебель и что растут они в пустынях. И колючки, и толщина, и место обитания связаны между собой. Очевидно, растению, которое живёт в сухом и жарком климате, нужно научиться компенсировать недостаток воды, и одно из решений — просто запасать её внутри, для чего и необходим толстый, сочный стебель.
Фото Натальи Домриной.
Устьица открываются и закрываются, регулируя испарение влаги и дыхание растений.
Этапы CAM-фотосинтеза протекают в разное время суток. Ночью в растения через открытые устьица проникает CO2, который присоединяется к органическим кислотам, образовавшимся при расщеплении углеводов. В составе кислоты CO2 накапливается в специальном пузырьке-вакуоли до утра, а с появлением солнечного света он выходит из состава кислоты и отправляется в хлоропласт, где ферменты фотосинтетического цикла Кальвина синтезируют из него углеводы.
Коллекция кактусов в Национальном ботаническом саду им. Н. Н. Гришко (г. Киев, Украина). Фото Виталия Пирожкова.
Фото Натальи Домриной.
‹
›
Открыть в полном размере
Но в жару много воды теряется, и весь запас влаги может просто высохнуть. Вообще транспирация (так называется испарение воды растением) — необычайно важный процесс. Листья, испаряющие влагу, играют роль насоса: они создают всасывающую силу, которая заставляет воду с растворёнными веществами подниматься по корням и сосудам. Однако, если растению выпало жить в условиях постоянной жары и засухи, испарение лучше как-то затормозить. Для этого можно избавиться от листьев, тем самым уменьшив испаряющую поверхность. Кактусы так и сделали: их листья превратились в колючки, а фотосинтетическую функцию листьев взял на себя стебель. Можно усовершенствовать собственную «кожу»: снабдить клетки внешнего слоя (эпидермиса) волосками и толстой восковой кутикулой. Через восковой слой воде труднее пробиться наружу, волоски же ослабляют воздушные потоки непосредственно вблизи поверхности стебля, что тоже уменьшает влагопотерю.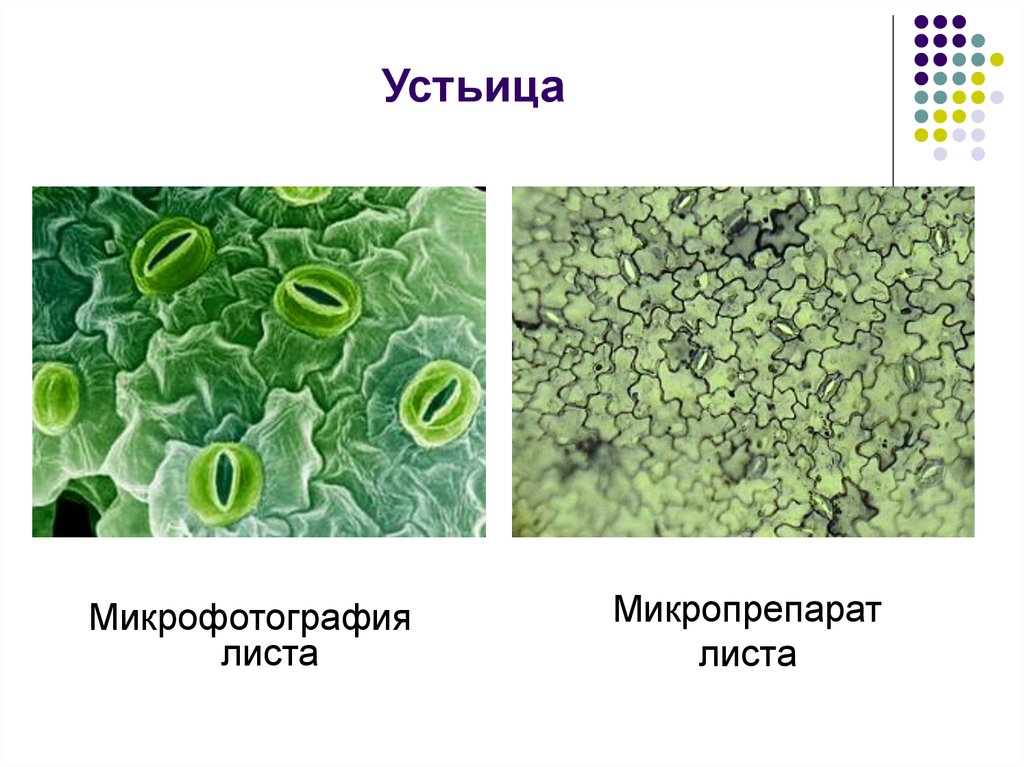 Но у кактусов есть ещё одна хитроумная уловка, которая связана с их способом фотосинтеза и которая тоже позволяет сохранять воду, невзирая на постоянную жару и засуху вокруг.
Но у кактусов есть ещё одна хитроумная уловка, которая связана с их способом фотосинтеза и которая тоже позволяет сохранять воду, невзирая на постоянную жару и засуху вокруг.
Фотосинтезом называют процесс образования органических веществ из углекислого газа и воды с использованием энергии солнечного света. Сначала энергия светового фотона с помощью сложных светособирающих молекул и молекулярных комплексов, в состав которых входит хлорофилл, сохраняется в специальных химических соединениях (именно на этом этапе нужна вода, из которой в качестве побочного продукта получается кислород), а потом с её помощью клетка синтезирует органические вещества. У фотосинтеза есть довольно интересные разновидности: например, некоторые бактерии способны осуществлять аноксигенный фотосинтез, при котором кислород не образуется. «Обычный» оксигенный фотосинтез присущ растениям, водорослям и цианобактериям.
Итак, растительной клетке, чтобы сделать молекулу глюкозы, нужны свет, вода и углекислый газ.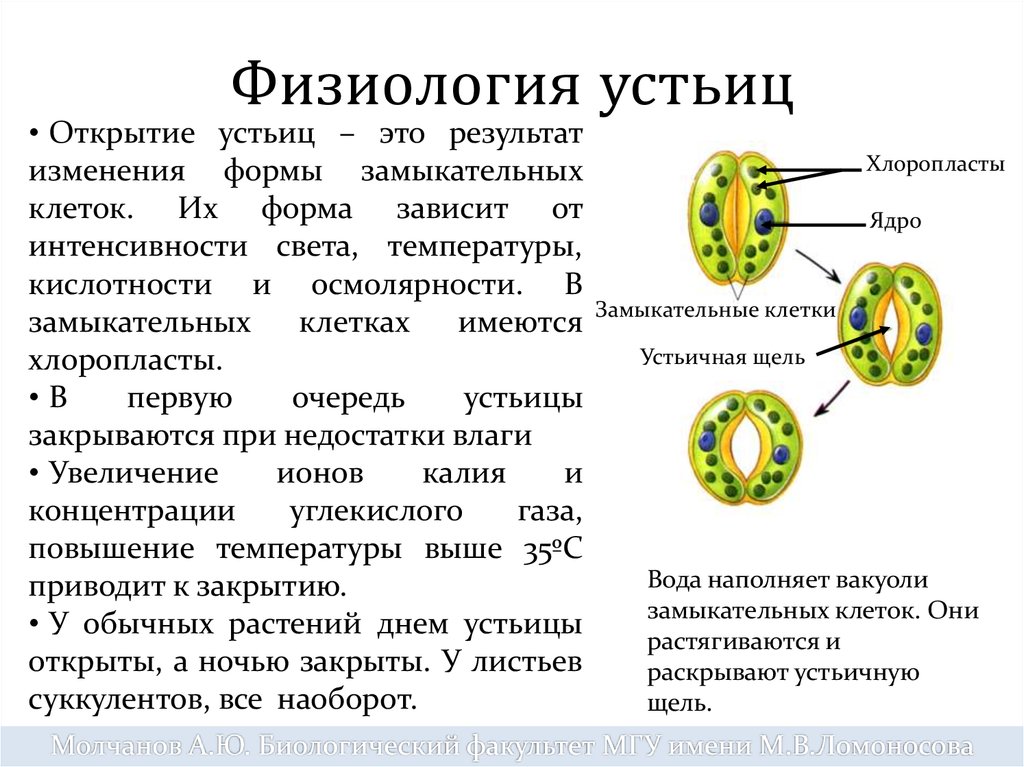 Вода поступает из-под земли через корни и систему сосудов, углекислый газ — из воздуха. Но у растения нет ни рта, ни лёгких, чтобы вдыхать СО2. Газообмен с окружающей средой осуществляется через устьица — особые поры в поверхности листьев и стеблей, окружённые замыкающими клетками.
Вода поступает из-под земли через корни и систему сосудов, углекислый газ — из воздуха. Но у растения нет ни рта, ни лёгких, чтобы вдыхать СО2. Газообмен с окружающей средой осуществляется через устьица — особые поры в поверхности листьев и стеблей, окружённые замыкающими клетками.
Устьица вносят довольно существенный вклад в испарение воды, и в жару их следовало бы держать всё время закрытыми. Но как тогда получать углекислый газ для фотосинтеза? Причём это не единственная проблема, связанная с фотосинтезом в жарком климате. Главнейший фотосинтетический фермент под названием «рибулозобисфосфаткарбоксилаза» (или РуБисКО), задача которого — присоединять углерод из углекислого газа к растущей молекуле сахара, при высокой температуре начинает работать в обратную сторону, то есть расщеплять полусинтезированный сахар. В таком случае клетке приходится возвращаться назад и заново повторять уже сделанную работу, естественно, с лишними тратами энергии. Поэтому эффективность фотосинтеза при повышении температуры сильно падает. Этого можно избежать, если поднять в листе концентрацию СО2 — тогда фермент при избытке углекислого сырья будет синтезировать углеводы. Но как это сделать?
Этого можно избежать, если поднять в листе концентрацию СО2 — тогда фермент при избытке углекислого сырья будет синтезировать углеводы. Но как это сделать?
Кактусы поступают так: они открывают устьица ночью и поглощают углекислый газ, но в производство глюкозы его не запускают — света-то нет. СО2 откладывается про запас в специальных мембранных пузырьках-вакуолях внутри клетки. Хранится он здесь не в чистом виде, а присоединённым к молекуле-посреднику, которая потом выдерживает ещё несколько превращений. В результате получается яблочная кислота. Но вот наступает день, и яблочная кислота отправляется из вакуоли в цитоплазму, где от неё отщепляется СО2, — теперь он может вступить в цикл фотосинтетических реакций, работающих от света. Растению уже не нужно открывать устьица, ведь можно использовать углекислый газ, запасённый за ночь, а значит, сильно сэкономить на испарении воды. Кроме того, соотношение СО2 и О2 за счёт запасов смещается в пользу первого, следовательно, фотосинтетические ферменты будут работать в сторону присоединения атомов углерода к растущей молекуле сахара, а не расщеплять её кислородом.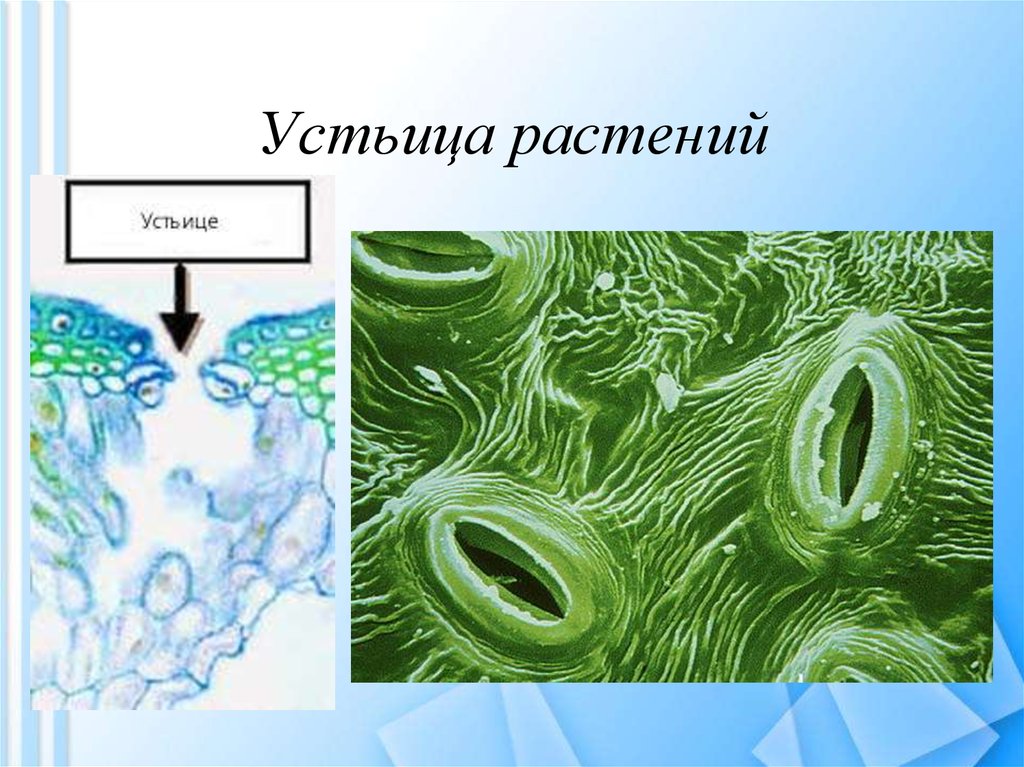
Такой тип фотосинтеза, когда фиксация СО2 и его использование в фотосинтетических реакциях разделены во времени, называется CAM-фотосинтезом. CAM расшифровывается как Crassu-laceae acid metabolism: здесь acid — кислота, в которую превращается запасённый углекислый газ, а Crassula-ceae, или толстянковые, — название семейства растений, у которых впервые обнаружили такой путь метаболизма. Но толстянки и кактусовые не единственные, кто его использует. CAM-фотосинтез нашли у ананаса и других представителей бромелиевых, у некоторых тыквенных, перечных, гераниевых и ряда других семейств, всего примерно у 9000 видов. Обычно это растения, которым приходится жить в жарком и сухом климате. Но не только: CAM-фотосинтез используют также и виды, живущие в воде, например полушники, стрелолисты и некоторые другие. Никакого противоречия тут нет: вод-ным растениям приходится решать ту же проблему, что и тем, которые вынуждены терпеть жару. Хотя в воде может быть довольно много растворённого СО2, диффундирует он в ней намного медленнее, чем в воздухе, так что рядом с растением, активно поглощающим углекислый газ, его будет хронически не хватать.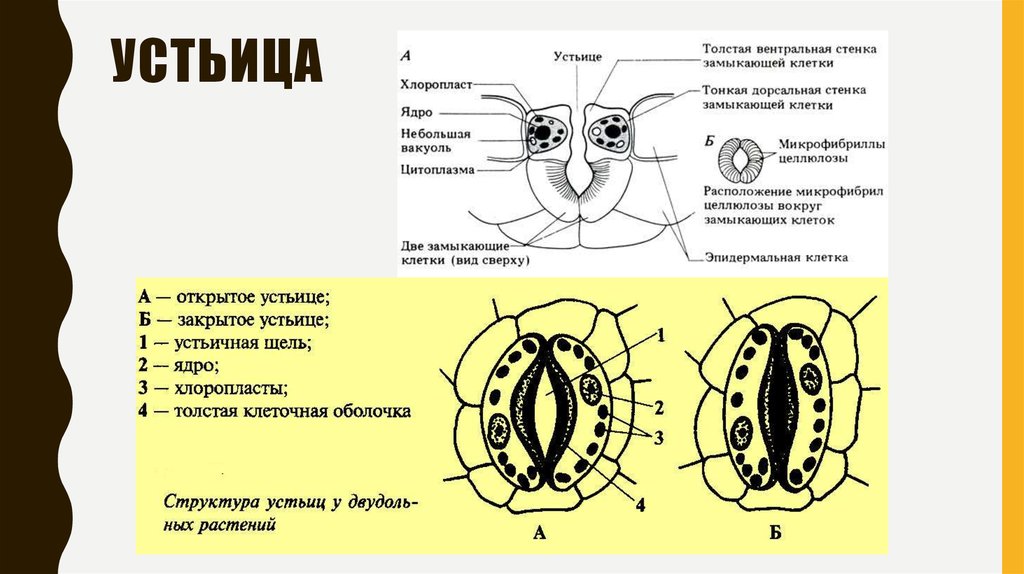 Выход — собирать СО2 не только днём, но и ночью, а поскольку ночью фотосинтезировать нельзя, то захваченный углекислый газ нужно запасать. А фотосинтез CAM-типа как раз и позволяет делать «углекислые» запасы.
Выход — собирать СО2 не только днём, но и ночью, а поскольку ночью фотосинтезировать нельзя, то захваченный углекислый газ нужно запасать. А фотосинтез CAM-типа как раз и позволяет делать «углекислые» запасы.
Напоследок снова вернёмся к жаровыносливым растениям. CAM-механизм позволяет максимально экономить воду, но, если оценивать количество готового продукта и энергию, потраченную на него, он менее эффективен, чем другие виды фотосинтеза. Так что некоторые CAM-виды используют его только по мере необходимости. Но кроме него существует ещё одна разновидность фотосинтеза, позволяющая днём держать бoльшую долю устьиц закрытыми. В этом случае фотосинтетические реакции происходят в глубинных клетках листа, окружающих жилки-сосуды. Клетки, лежащие ближе к поверхности, во-первых, с помощью энергии света производят топливо для синтеза углеводов, во-вторых, ловят углекислый газ и присоединяют его к молекуле-посреднику. Образовавшаяся в результате кислота и энергетические молекулы сразу отправляются в глубь листа, где СО2 отсоединяется от кислоты-переносчика и вступает в синтетический цикл.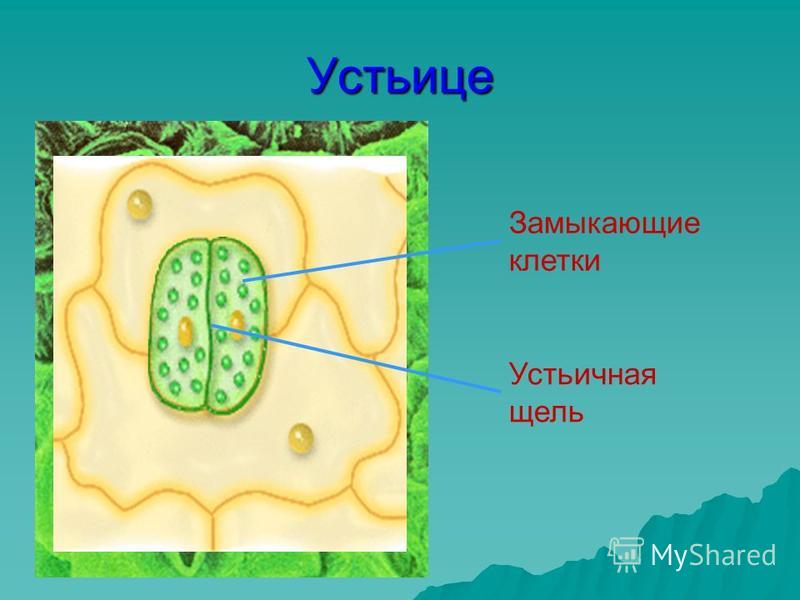 Такой путь называется С4-фотосинтезом, и он, как видим, похож на CAM, только здесь фиксация углекислого газа и его использование в синтезе разделены не во времени, между ночью и днём, а в пространстве, между разными клетками.
Такой путь называется С4-фотосинтезом, и он, как видим, похож на CAM, только здесь фиксация углекислого газа и его использование в синтезе разделены не во времени, между ночью и днём, а в пространстве, между разными клетками.
Смысл С4-фотосинтеза в том, чтобы переправить СО2 во внутренние ткани листа, где концентрация кислорода невелика. Мы помним, что при повышении температуры фермент РуБисКО всё сильнее начинает работать в обратную сторону, то есть расщеплять с помощью кислорода промежуточные продукты фотосинтеза. Но если кислорода мало, фермент будет работать в правильном синтетическом направлении. С другой стороны, С4-путь позволяет сократить испарения воды через устьица: в самое жаркое время дня растение может использовать накопленный углекислый газ, запасы которого образовались благодаря пространственному разделению разных блоков реакций; сами же устьица на время можно и закрыть. Экономия воды здесь не так велика, как при CAM-способе, но зато продуктивность фотосинтеза получается выше, так что неудивительно, что С4-схему используют около 7600 видов растений, среди которых множество злаков, в том числе кукуруза, сорго, просо и сахарный тростник.
Ночная жизнь растений
Что «делают» растения ночью? На этот вопрос так и хочется ответить: «Отдыхают». Ведь, казалось бы, вся «активная жизнь» растения происходит днем. В дневные часы цветы раскрываются и опыляются насекомыми, развертываются листья, молодые стебли растут и тянут свои верхушки к солнцу. Именно в течение светлого времени суток растения используют солнечную энергию для того, чтобы преобразовывать углекислый газ, поглощаемый ими из атмосферного воздуха, в сахар.
Однако растение не только синтезирует органические вещества – оно их и использует в процессе дыхания, снова окисляя до углекислого газа и поглощая при этом кислород. Но количество кислорода, необходимого растениям для дыхания, примерно в 30 раз меньше того, что выделяется ими в процессе фотосинтеза. Ночью, в темноте, фотосинтез не происходит, но и в это время растения потребляют так мало кислорода, что это нисколько не сказывается на нас с вами. Поэтому старая традиция выносить растения из комнаты больного на ночь совершенно не обоснованна.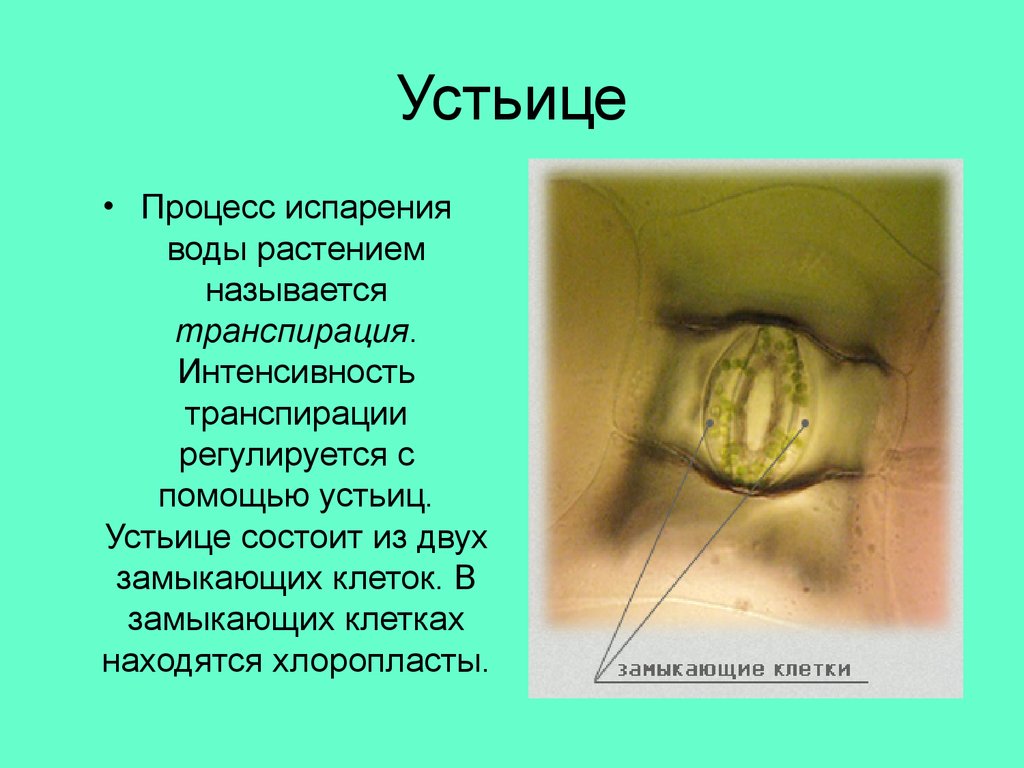
А еще есть ряд видов растений, которые потребляют углекислый газ именно ночью. Поскольку энергии солнечного света, необходимой для полного восстановления углерода, в это время нет, сахар, конечно, не образуется. Но поглощенная из воздуха углекислота сохраняется в составе яблочной или аспарагиновой кислот, которые потом, уже на свету, вновь разлагаются, высвобождая СО2. Именно эти молекулы углекислого газа включаются в цикл основных реакций фотосинтеза – так называемый цикл Кальвина. У большинства же растений этот цикл начинается с захвата молекулы СО2 непосредственно из воздуха. Такой «простой» способ носит название С3-пути фотосинтеза, а если углекислый газ предварительно запасается в яблочной кислоте – это С4-путь.
Казалось бы, зачем нужны дополнительные сложности? В первую очередь для того, чтобы экономить воду. Ведь поглощать углекислоту растение может только через открытые устьица, через которые происходит и испарение воды. И днем, в жару, воды через устьица теряется намного больше, чем ночью.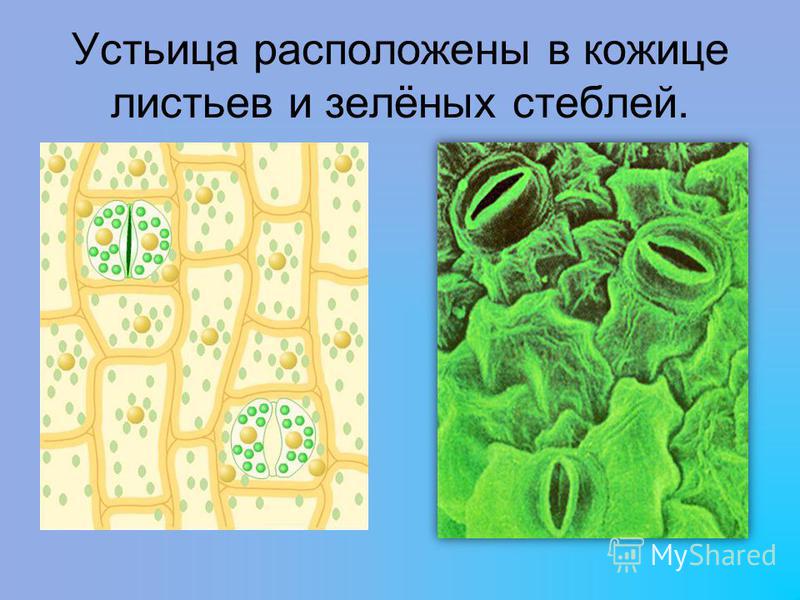 А у С4-растений устьица днем закрыты, и вода не испаряется. Газообмен же эти растения осуществляют в прохладные ночные часы. Кроме того, С4-путь в целом более эффективен, он позволяет синтезировать большее количество органических веществ в единицу времени. Но только в условиях хорошей освещенности и при достаточно высокой температуре воздуха.
А у С4-растений устьица днем закрыты, и вода не испаряется. Газообмен же эти растения осуществляют в прохладные ночные часы. Кроме того, С4-путь в целом более эффективен, он позволяет синтезировать большее количество органических веществ в единицу времени. Но только в условиях хорошей освещенности и при достаточно высокой температуре воздуха.
Так что С4-фотосинтез свойствен «южанам» – растениям из жарких областей. Он присущ большинству кактусов, некоторым другим суккулентам, ряду бромелиевых – например всем хорошо известному ананасу (Ananas comosus), сахарному тростнику и кукурузе.
Интересно, что еще в 1813 г., задолго до того, как стали известны биохимические реакции, лежащие в основе фотосинтеза, исследователь Бенджамин Хейн написал в Линнеевское научное общество о том, что листья ряда суккулентных растений имеют особенно острый вкус по утрам, а затем, к середине дня, их вкус становится более мягким.
Способность использовать связанный в органических кислотах СО2 обусловлена генетически, но реализация этой программы находится и под контролем внешней среды. При сильном дожде, когда угрозы высыхания нет, а освещенность невысока, С4-растения могут открывают свои устьица днем и переходить на обычный С3-путь.
При сильном дожде, когда угрозы высыхания нет, а освещенность невысока, С4-растения могут открывают свои устьица днем и переходить на обычный С3-путь.
А что еще может происходить с растениями по ночам?
Некоторые виды приспособились привлекать своих опылителей именно в ночные часы. Для этого они используют разные средства: и усиливающийся к ночи запах, и приятный и заметный для глаза ночных опылителей цвет – белый или желтовато-бежевый. На такие цветы летят ночные бабочки. Именно они опыляют цветы жасмина (Jasminum), гардении (Gardenia), лунных цветов (Ipomea alba), вечерницы, или ночной фиалки (Hesperis), любки двулистной (Platanthera bifolia), лилии кудреватой (Lilium martagon) и ряда других растений.
А есть растения (их называют хироптерофильными), которые опыляются в ночные часы летучими мышами. Больше всего таких растений в тропиках Азии, Америки и Австралии, меньше в – Африке. Это бананы, агавы, боабабы, некоторые представители семейств миртовых, бобовых, бегониевых, геснериевых, синюховых.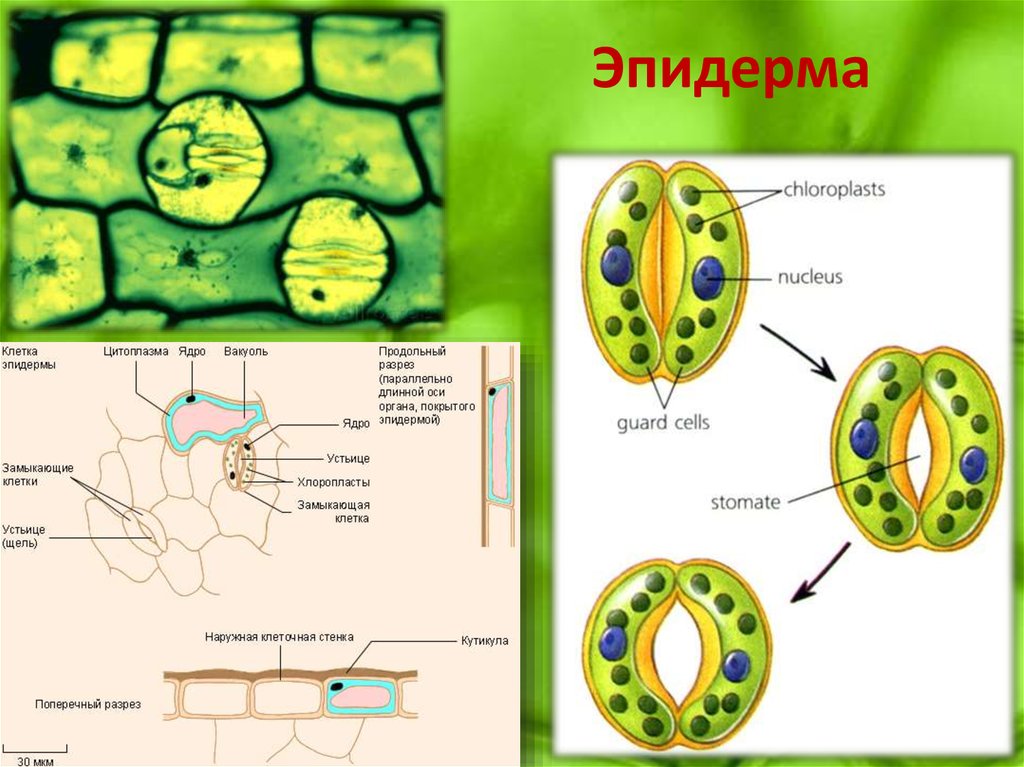
Цветки хироптерофильных растений раскрываются только с наступлением сумерек и не отличаются яркостью окраски – как правило, они зеленовато-желтые, коричневые или фиолетовые. Запах у таких цветков весьма специфический, часто неприятный для нас, но, наверное, привлекательный для летучих мышей. Кроме того, цветки хироптерофильных растений обычно крупные, с прочным околоцветником и снабжены «посадочными площадками» для своих опылителей. В качестве таких площадок могут выступать толстые цветоножки и цветоносы или безлистные участки ветвей, примыкающих к цветкам.
Некоторые хироптерофильные растения даже «разговаривают» со своими опылителями, привлекая их. Когда цветок лианы Mucuna holtonii, принадлежащей к семейству бобовых и произрастающей в тропических лесах Центральной Америки, становится готовым к опылению, один из его лепестков приобретает специфическую вогнутую форму. Этот вогнутый лепесток концентрирует и отражает сигнал, издаваемый летучими мышами, отправившимися на поиски корма, и таким образом сообщает им о своем местонахождении.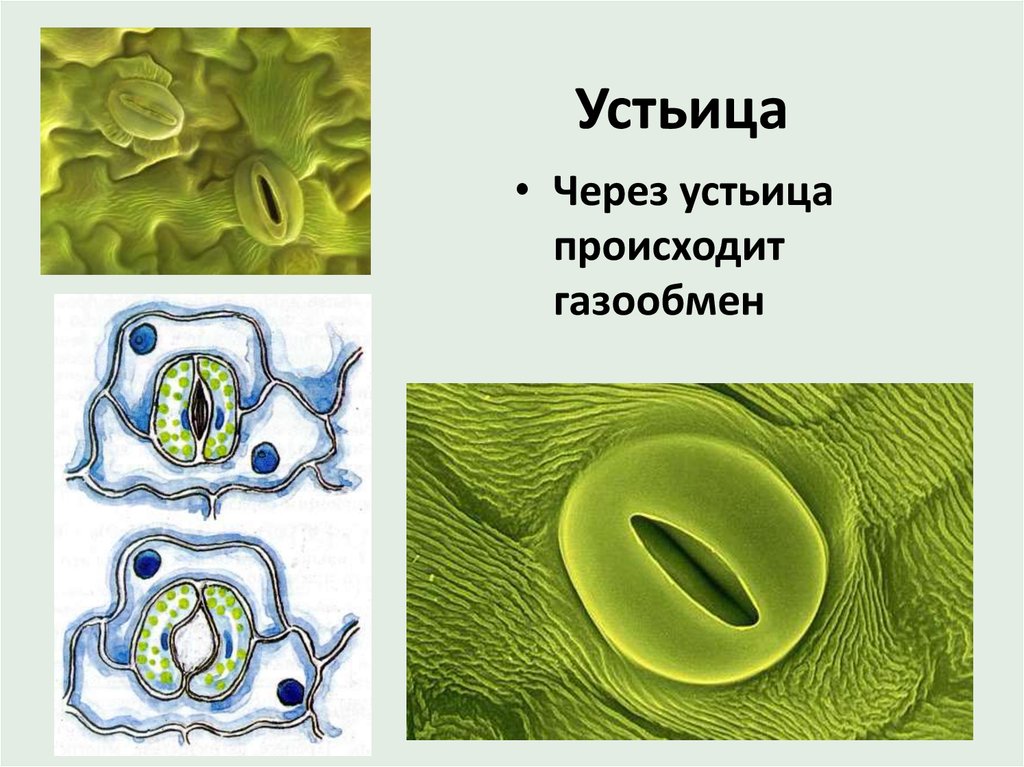
Но не только рукокрылые млекопитающие опыляют цветы. В тропиках известно более 40 видов зверьков из других отрядов, активно участвующих в опылении около 25 видов растений. У многих из этих растений, как и у тех, которые опыляются летучими мышами, цветки крупные и прочные, часто неприятно пахнущие и образующие большое количество пыльцы и нектара. Обычно число цветков на таких растениях или в их соцветиях невелико, цветки располагаются низко над землей и раскрываются только к ночи, чтобы обеспечить максимальное удобство ночным зверюшкам.
Н.Ю. Феоктистова, с сайта roman.by
P.S. Одно из наиболее удивительных опыляемых растений — амазонская лилия. О нем в следующей статье рубрики «Биопарк». Прочитайте, не пожалеете. Амазонская лилия еще никого не оставила равнодушным.
Поглощение и транспорт воды сосудистыми растениями
Агриос, Г. Н. Патология растений . Нью-Йорк,
Нью-Йорк: Academic Press, 1997.
Beerling, DJ & Franks, PJ Plant
наука: Скрытая стоимость транспирации. Природа
Природа
464, 495-496 (2010).
Brodersen, C. R. и др. . Динамика репарации эмболов в ксилеме: In vivo
визуализации с помощью компьютерной томографии высокого разрешения Физиология растений 154 , 1088-1095 (2010 г.).
Бродрибб, Т.Дж. и Холбрук, Н.М.
Водный стресс деформирует трахеиды, расположенные по периферии жилки листа тропического хвойного дерева.
Физиология растений 137 , 1139-1146 (2005)
Canadell, J. et al . Максимальная глубина укоренения типов растительности в мире
шкала. Экология 108, 583-595 (1996).
Чоат Б., Кобб А. Р. и Янсен С.
Структура и функция окаймленных карьеров: новые открытия и влияние на
гидравлическая функция всего завода. Новый
Фитолог 177, 608-626 (2008).
Чанг, Х. Х. и Крамер, П. Дж.
Поглощение воды и «Р через опробковевшие и неопробковевшие корни лоблолли
сосна. Canadian Journal of Forest Research 5,
229–235 (1975).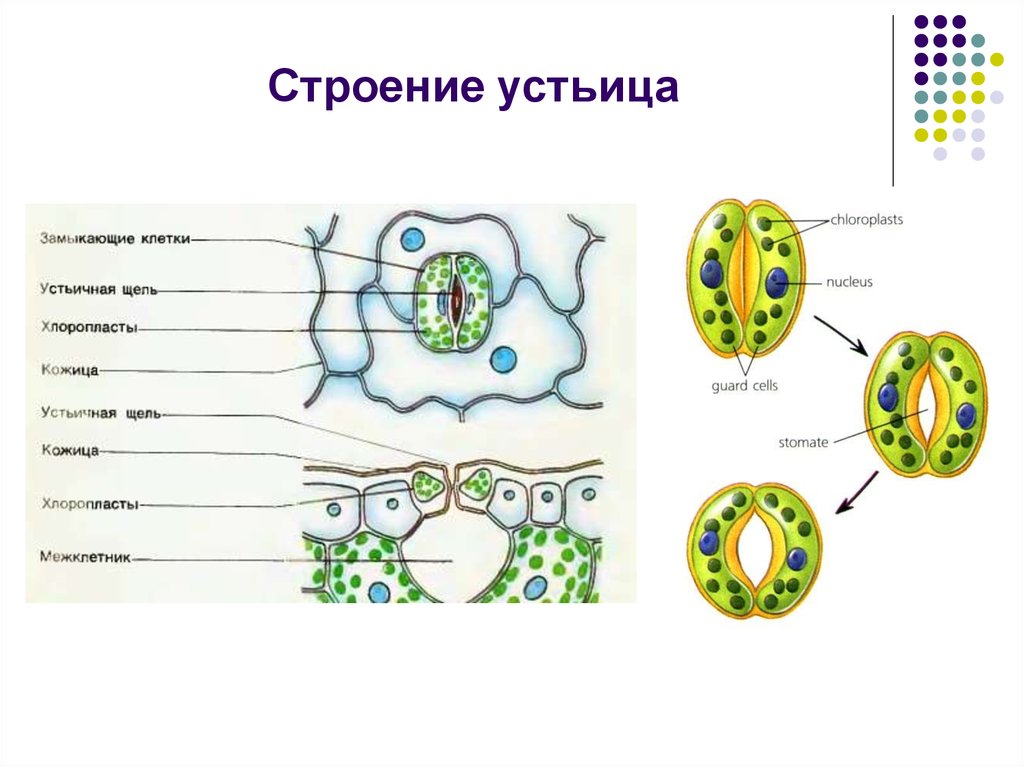
Ипен, Д. и др. . Гидротропность: реакция роста корней на воду. Trends in Plant Science 10, 44-50 (2005).
Hetherington, A.M. & Woodward, F.I.
Роль устьиц в восприятии и управлении изменениями окружающей среды. Природа 424, 901-908 (2003).
Holbrook, N.M. & Zwieniecki, M.A. Сосудистый транспорт в растениях . Сан-Диего, Калифорния:
Elsevier Academic Press, 2005.
Джавот, Х. и Морел, К. Роль
аквапорины в поглощении воды корнями. Анналы
ботаники 90, 1-13 (2002).
Kramer, P.J. & Boyer, J.S. Водные отношения растений и почв . Нью-Йорк, штат Нью-Йорк:
Academic Press, 1995.
Kramer, PJ & Bullock, HC
Сезонные изменения соотношения опробковевших и неопробковевших корней
деревьев по отношению к поглощению воды. Американский журнал ботаники 53,
200-204 (1966).
Макфолл, Дж. С.,
Джонсон, Г. А. и Крамер, П. Дж. Наблюдение за областью истощения воды
окружающие корни сосны лоболли с помощью магнитно-резонансной томографии. Производство
Производство
Национальной академии
наук Соединенных Штатов Америки 87 , 1203-1207 (1990).
Маккалли, М. Е. Корни в почве: раскопки
сложности корней и их ризосфер. Ежегодный обзор физиологии растений и молекулярной биологии растений 50, 695-718 (1999).
McDowell, N.G. и др. . Механизмы выживания и гибели растений при засухе:
Почему одни растения выживают, а другие погибают от засухи? Новый фитолог 178, 719-739 (2008).
Нардини, А., Ло Гулло, М. А. и Саллео,
S. Повторное заполнение эмболизированных каналов ксилемы: это вопрос разгрузки флоэмы? Растениеводство 180, 604-611 (2011).
Питтерманн, Дж. и др. . Ямки торус-марго помогают хвойным растениям конкурировать с покрытосеменными. Наука 310, 1924 (2005).
Сак, Л. и Холбрук, Н. М. Лиф
гидравлика. Ежегодный обзор завода
Биология 57, 361-381 (2006).
Сак, Л. и Тайри, М. Т. «Лист
Т. «Лист
гидравлика и ее влияние на структуру и функции растений», в Сосудистый транспорт в растениях , под редакцией Н.М.
Холбрук и М.А. Звенецкий. (Сан
Диего, Калифорния: Elsevier Academic
Пресс, 2005) 93-114.
Шенк, Х. Дж. и Джексон, Р. Б. Рутинг
глубина, боковое распространение корней и подземные / надземные аллометрии растений
в условиях ограниченного количества воды. Журнал
Экология 90, 480-494 (2002).
Сперри, Дж. С. и Тайри, М. Т.
Механизм ксилемной эмболии, вызванной водным стрессом. Физиология растений 88, 581-587
(1988).
Steudle, E. Сплоченность-напряжение
механизм и приобретение воды корнями растений. Ежегодный обзор физиологии растений и молекулярной биологии 52, 847-875 (2001).
Steudle, E. Транспорт воды в растениях.
Контроль окружающей среды в биологии 40, 29-37 (2002).
Takahashi, H. Гидротропизм и его взаимодействие с гравитропизмом в
корнеплоды.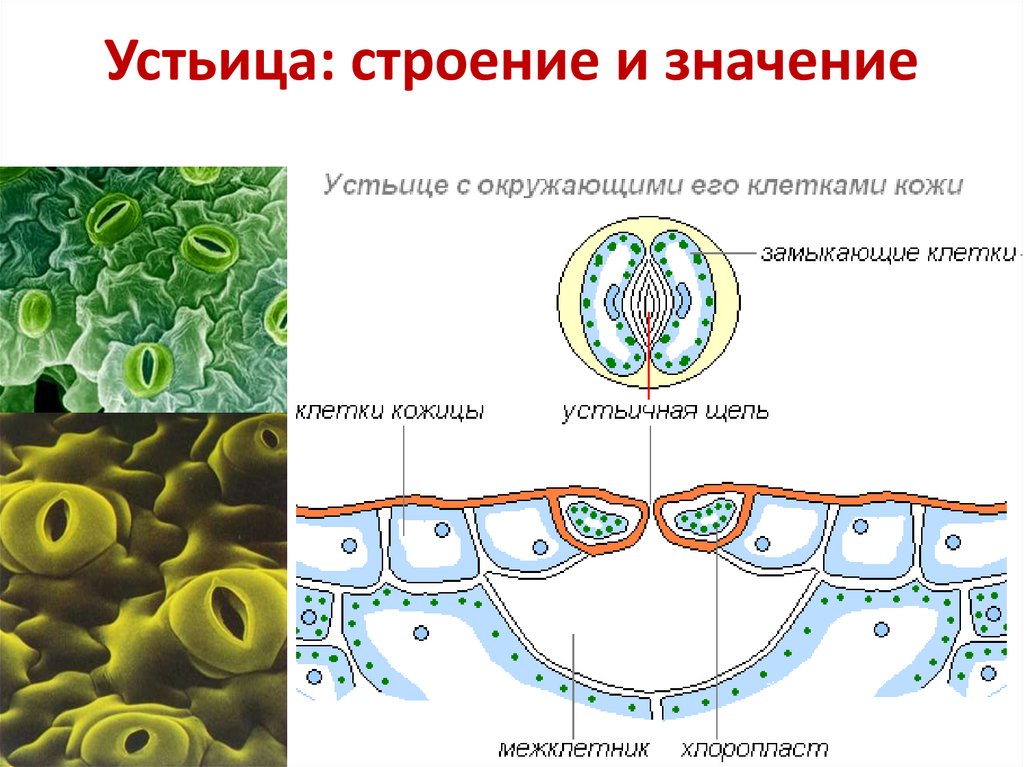 Почва для растений 165 , 301-308 (1994).
Почва для растений 165 , 301-308 (1994).
Тайри, М. Т. и Эверс, Ф. В.
гидравлическая архитектура деревьев и других древесных растений. Новый фитолог 119, 345-360
(1991).
Тайри, М. Т. и Сперри, Дж. С.
Уязвимость ксилемы к кавитации и эмболии. Ежегодный обзор физиологии растений и молекулярной биологии 40, 19-38 (1989).
Тайри, М. Т. и Циммерман, М. Х. Структура ксилемы и восхождение сока . 2-й
изд. Нью-Йорк, штат Нью-Йорк: Springer-Verlag, 2002.
Тайри, М. Т. и Эверс, Ф.
гидравлическая архитектура деревьев и других древесных растений. Новый фитолог 119, 345-360
(1991).
Уилер, Т. Д. и Струк, А. Д.
транспирация воды при отрицательном давлении в синтетическом дереве. Природа 455, 208-212 (2008).
Wullschleger, S.D., Meinzer, F.C. &
Вертесси, Р.А. Обзор исследований использования воды деревьями всего растения. Физиология деревьев 18, 499-512 (1998).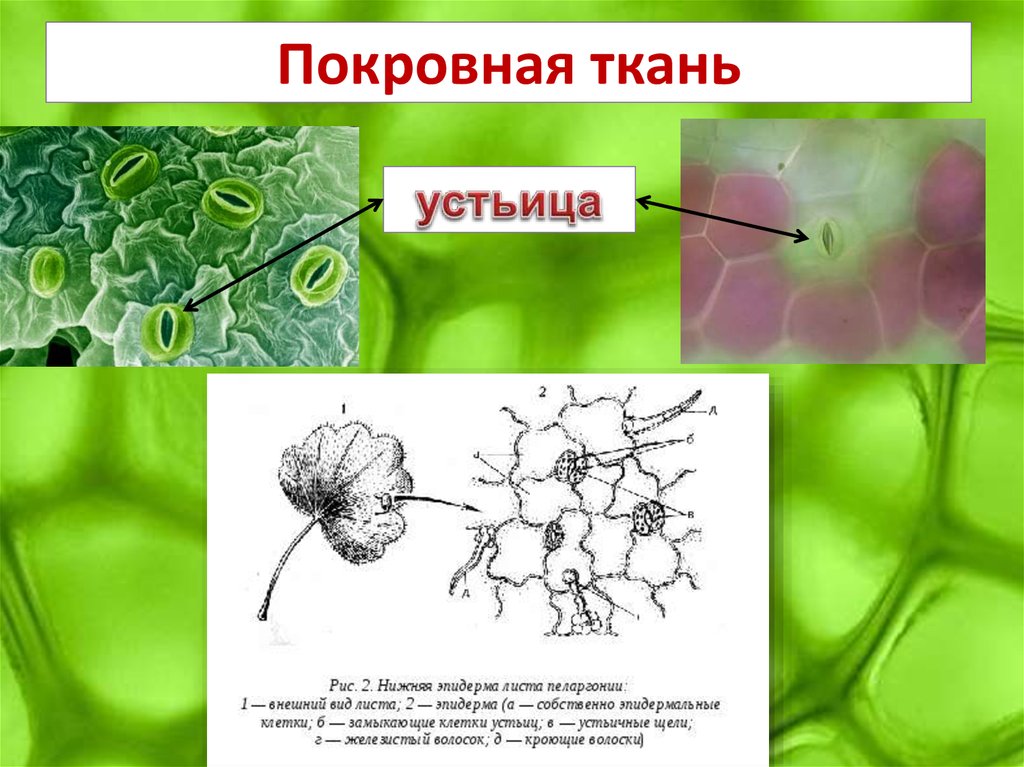
Циммерман, MH Xylem Structure and the Ascent of Sap . 1-е изд. Берлин, Германия:
Springer-Verlag, 1983.
Стэнфордские исследователи изучают, как растения приспосабливаются к климату
Сколько ртов нужно растению, чтобы выжить? Ответ меняется в зависимости от климата, и некоторые решения принимаются задолго до того, как прорастет новый лист.
Исследователи из Стэнфорда обнаружили, что образование микроскопических пор, называемых устьицами (от греческого слова stoma, означающего рот), контролируется специфическим сигнальным путем, который блокирует активность одного белка, необходимого для развития устьиц. Выводы описаны в статье, опубликованной 14 ноября в журнале 9.0003 Наука .
Устьица есть почти на каждом наземном растении на Земле. Их многочисленные функции включают выделение влаги и кислорода в окружающую среду, обеспечение внутреннего кондиционирования воздуха для растения и обеспечение проникновения углекислого газа в лист, где он превращается в сахар во время фотосинтеза.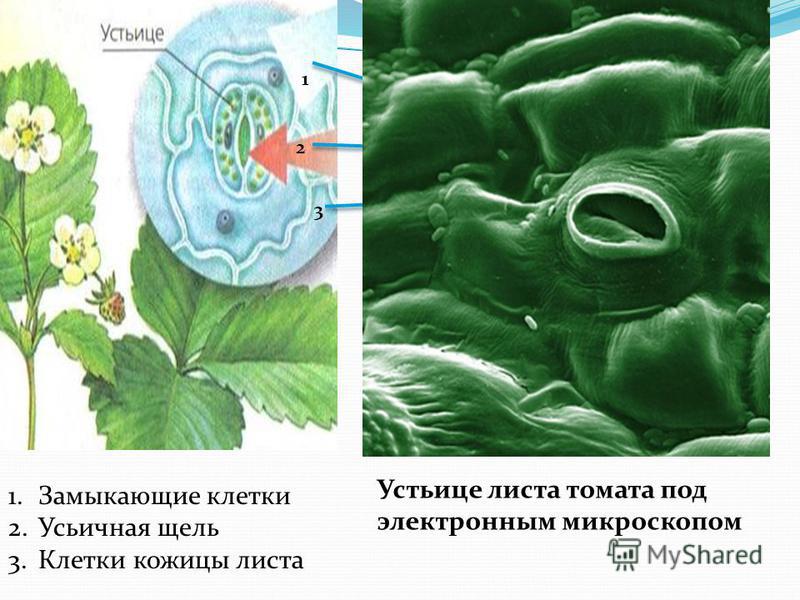 Устьица необходимы для выживания растений и, поглощая углерод из атмосферы, играют важную роль в поддержании здоровья планеты.
Устьица необходимы для выживания растений и, поглощая углерод из атмосферы, играют важную роль в поддержании здоровья планеты.
Использование Arabidopsis thaliana , быстрорастущее цветущее растение, используемое для генетических исследований и исследований развития, Доминик Бергманн, доцент биологии, и соавторы статьи Грегори Лэмпард, постдокторский научный сотрудник, и Кора Макалистер, аспирант, обнаружили уникальная структурная область на белке с 10 сайтами, которые могут быть модифицированы хорошо известным сигнальным путем, контролируемым окружающей средой, чтобы определять количество устьиц, которые образует растение.
«Ученые говорят, что окружающая среда влияет на развитие растений, но никто не может указать белок, ответственный за эту реакцию», — сказал Бергманн. «Теперь мы знаем основную мишень внутри клетки и то, как она регулируется».
Знание того, как работает этот процесс, может быть использовано для модификации сельскохозяйственных культур, чтобы максимизировать их продуктивность в изменяющихся климатических условиях. Бергманн сказал, что первоначально растения могут получить выгоду в результате увеличения поступления углерода в атмосферу из-за глобального потепления, но они также отреагируют на эти условия, производя меньше устьиц. Результат? Потеря охлаждения через устьица может привести к массовым неурожаям из-за повышения температуры, связанного с глобальным потеплением.
Бергманн сказал, что первоначально растения могут получить выгоду в результате увеличения поступления углерода в атмосферу из-за глобального потепления, но они также отреагируют на эти условия, производя меньше устьиц. Результат? Потеря охлаждения через устьица может привести к массовым неурожаям из-за повышения температуры, связанного с глобальным потеплением.
«Есть обстоятельства, когда вы можете захотеть отключить сигналы, получаемые растениями от окружающей среды, чтобы они могли выжить», — сказал Бергманн.
Белок, который исследователи назвали SPEECHLESS, инициирует первый из трехэтапных процессов клеточного деления, который приводит к образованию устьиц у растений. Несмотря на структурное сходство с SPEECHLESS, два других белка, участвующих в последующих стадиях, не содержат той же контрольной области, которая регулируется сигнальным путем. Это обеспечивает уникальный механизм сигнального пути для контроля активности SPEECHLESS в наборе клеток, подобных стволовым клеткам, и, следовательно, окончательного развития устьиц.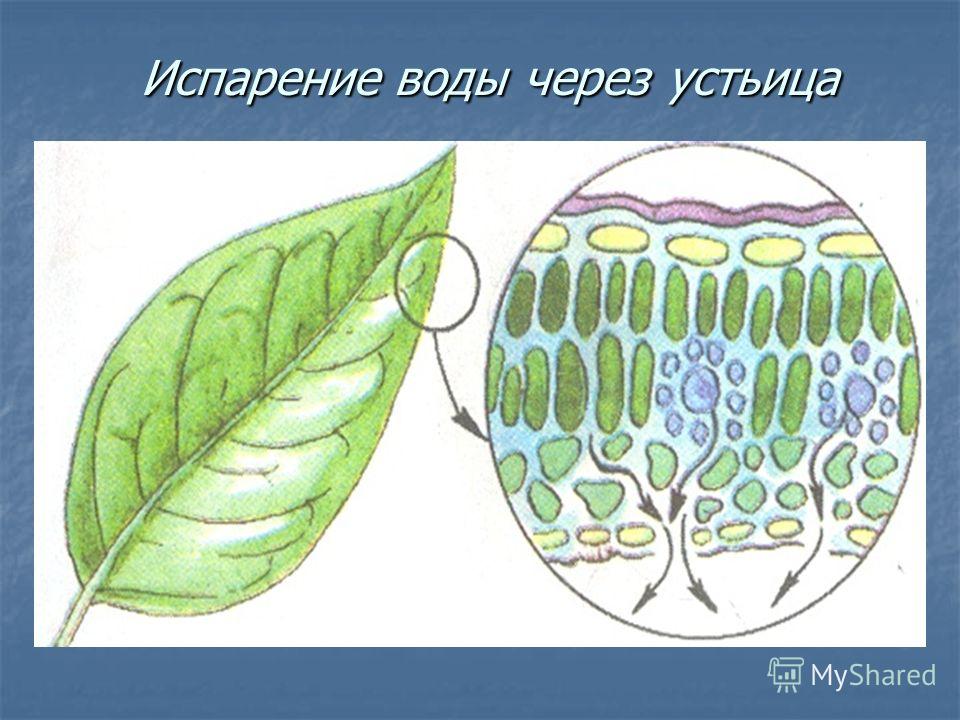
«Если бы я разрабатывал лист, то именно эту часть я бы тщательно контролировал, — сказал Бергманн. «Похоже, это то, что сделали растения».
Определенные компромиссы существуют для растений, имеющих слишком много или слишком мало устьиц. Чтобы помочь определить количество устьиц, которые должен сформировать только что проросший лист, растение принимает во внимание ключевые факторы окружающего климата — уровень углекислого газа, температуру и влажность.
Для восприятия этих факторов растение использует тот же сигнальный путь, что и для управления БЕЗГЛАВНОЙ активностью. В исследовании определяется критический узел, который связывает то, как растение может ощущать условия окружающей среды, с тем, как эта информация передается путям развития устьиц. Таким образом, развитие устьиц можно изменить «на лету», чтобы растение лучше справлялось с условиями окружающей среды.
Например, в листе меньше пор, когда углекислого газа в атмосфере много, и больше, когда его мало.