Содержание
Как зарождается жизнь, или Что происходит в семени
Любой организм развивается только из одной клетки, но рождается многоклеточным. Как же сложно и закономерно делится эта клетка, что в итоге все живые организмы не похожи друг на друга! Что за закономерности приводят к правильному развитию животных, уже изучено достаточно подробно, а какова логика в случае растений?
Эмбриологию (учение о зародыше: εμβρυον — эмбрион, зародыш — плюс — λογια, от λογος — учение) теперь всё чаще называют биологией развития. Такое изменение неслучайно: все живые организмы развиваются из одной клетки, и благодаря современным методам мы можем проследить весь этот путь, то есть изучить не только зародыш, но и то, как он развился из зиготы — клетки, возникающей при оплодотворении яйцеклетки сперматозоидом. Скоординированная регуляция экспрессии тысяч генов из-за создающихся градиентов определенных веществ для животных описана довольно хорошо: например, о модели создания этих градиентов в процессе развития конечностей рассказывала моя коллега Дарья Сергеева [1, 2]. Я же расскажу вам про развитие растений.
Я же расскажу вам про развитие растений.
Орган размножения растений — цветок. Яркие лепестки венчика служат для привлечения опылителей, и, конечно, со школы все помнят, что есть пестики и тычинки (а зачем они — помнят уже немногие). В пестике — зародышевый мешок, в тычинке — пыльца. Пыльца, прорастая через пестик к зародышевому мешку, оплодотворяет яйцеклетку, и получается зигота, из которой развивается зародыш (рис. 1). Подобно птицам и рептилиям, эмбрион которых развивается под оболочками яйца, зародыш растения развивается под оболочками семени.
Рисунок 1. Закон зародышевого сходства Карла Бэра гласит, что ранние стадии эмбрионального развития животных похожи. А как насчет растений? Рисунок с сайта pixshark.com.
Первое деление зиготы растений уже говорит о том, что нас ждет что-то необычное, ведь оно несимметричное. Второе деление в верхней и нижней клетке происходит в разных плоскостях. В верхней клетке — вертикально, в нижней — горизонтально.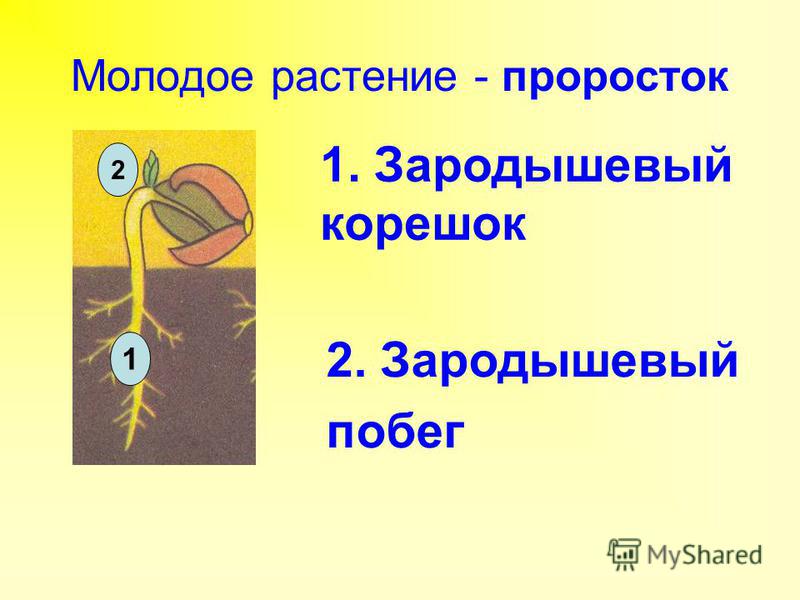 По прошествии еще нескольких делений зародыш превращается в «шарик на ножке» — глобулярная стадия. Эта ножка — суспензор — связывает зародыш с материнским растением, подобно пуповине млекопитающих, питая его. Между суспензором и глобулой есть одна чрезвычайно важная клетка — гипофиза, — из которой развивается апикальная меристема корня (рис. 2).
По прошествии еще нескольких делений зародыш превращается в «шарик на ножке» — глобулярная стадия. Эта ножка — суспензор — связывает зародыш с материнским растением, подобно пуповине млекопитающих, питая его. Между суспензором и глобулой есть одна чрезвычайно важная клетка — гипофиза, — из которой развивается апикальная меристема корня (рис. 2).
Рисунок 2. Стадии развития зародыша растений: октант, глобула, сердце, торпеда и зрелый зародыш. Разными цветами показаны области зародыша, из которых разовьются разные ткани. В этой статье речь пойдет главным образом о развитии апикальной меристемы побега (shoot apical meristem, SAM), показанной на рисунке красным цветом, и апикальной меристемы корня (root apical meristem, RAM), показанной фиолетовым. Рисунок с сайта www.mun.ca.
«Апикальная» значит «верхушечная» (от лат. apex — верхушка). Так случилось, что у растений две верхушки: одна растет вверх — побег, а другая вниз — корень.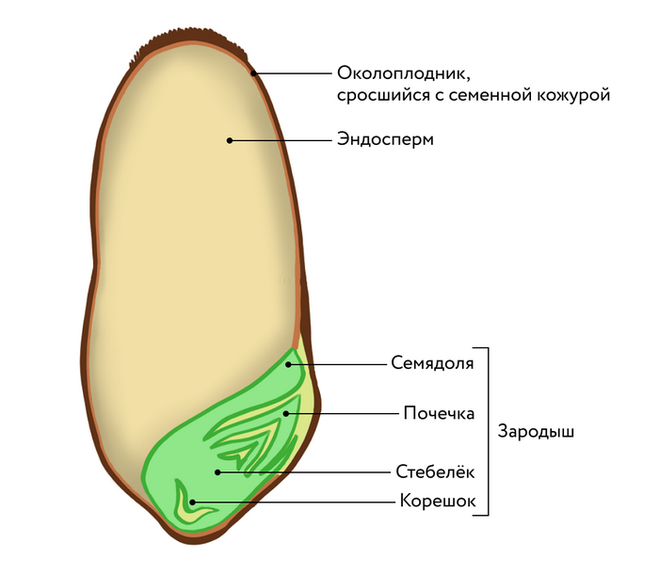 Поэтому меристема корня тоже называется апикальной (хотя правильнее было бы назвать ее базальной (от греч. basis — основа)). Меристема, или образовательный центр, — одна из самых важных частей растения: именно от меристематических клеток берут свое начало все остальные. Меристемы побега и корня имеют сходное строение: в них выделяют покоящийся центр, клетки которого редко делятся, но сигнализируют остальным клеткам меристемы оставаться недифференцированными, то есть не вытягиваться и не становиться клеткой сосуда, не накапливать хлорофилл, превращаясь в клетку листа, а сохраняться в эмбриональном виде.
Поэтому меристема корня тоже называется апикальной (хотя правильнее было бы назвать ее базальной (от греч. basis — основа)). Меристема, или образовательный центр, — одна из самых важных частей растения: именно от меристематических клеток берут свое начало все остальные. Меристемы побега и корня имеют сходное строение: в них выделяют покоящийся центр, клетки которого редко делятся, но сигнализируют остальным клеткам меристемы оставаться недифференцированными, то есть не вытягиваться и не становиться клеткой сосуда, не накапливать хлорофилл, превращаясь в клетку листа, а сохраняться в эмбриональном виде.
Как у всех живых организмов, функционирование клеток зависит от того, начнут ли работать определенные гены или нет, что в свою очередь определяется наличием транскрипционного фактора и его способностью запустить работу гена. Апикальную меристему побега регулирует транскрипционный фактор WUSCHEL (WUS), а апикальную меристему корня WUSCHEL-RELATED HOMEOBOX 5 (WOX5).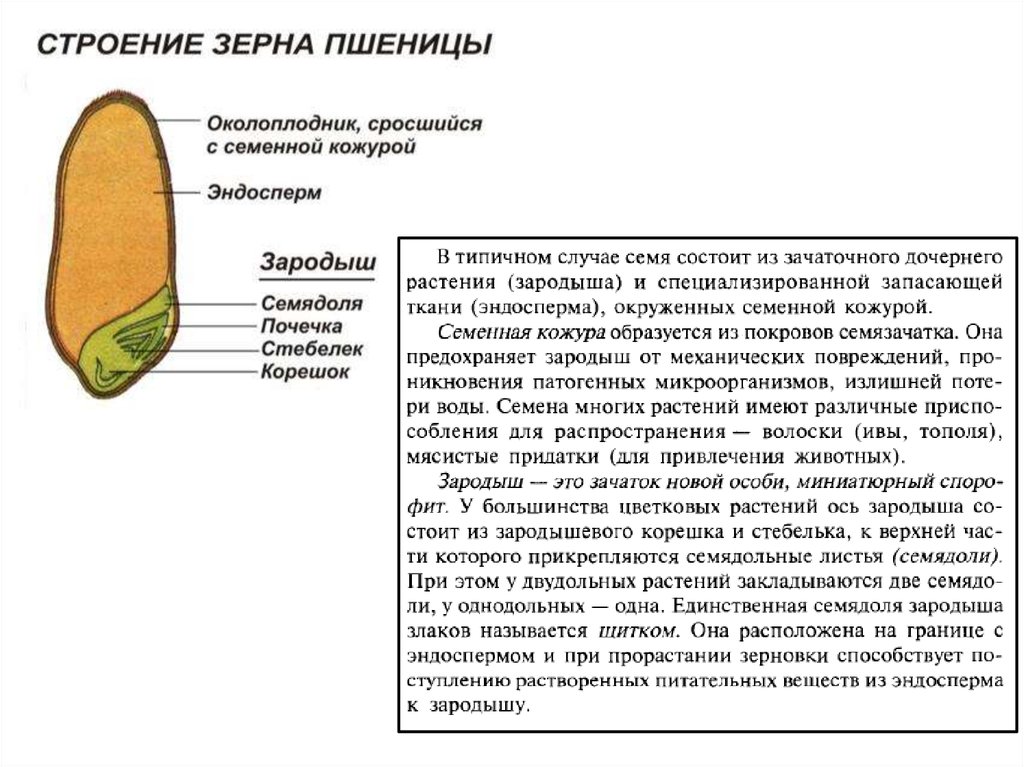
Рисунок 3. Экспрессионный каскад, запускающий формирование корня. Под воздействием ауксина (auxin) синтезируются транскрипционные факторы: MP (MONOPTEROS, или ARF5), NTT (NO TRANSMITTING TRACT), WOX5 (WUSCHEL-RELATED HOMEOBOX 5). В эксперименте экспрессируется и pDR5-GFP — репортерная система, состоящая из гена зеленого флуоресцентного белка и синтетического промотора DR5. Этот промотор содержит последовательности, «чувствующие» ауксин (у растений они находятся в промоторах генов, активируемых ауксином). Зеленый белок, соответственно, синтезируется только при контакте промотора с ауксином. Рисунок из [4].
А кто контролирует эти транскрипционные факторы? Почему клетки зародыша так странно и несимметрично делились на ранних этапах? И как из гипофизы развивается корень?
Всё определяется гормонами, главный из которых ауксин. Еще на начальном этапе его распределение «рассказало» клеткам, где низ.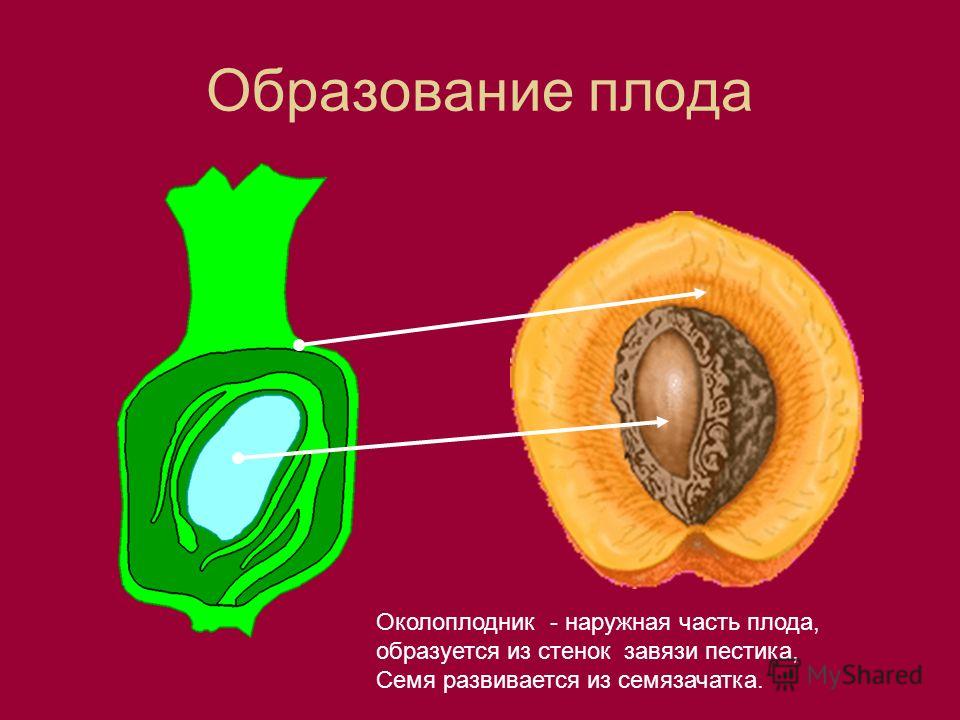 Давно известно, что переносчики ауксина (белки семейства PIN) показывают, куда гормон пойдет, а куда он пойдет — там корень и разовьется.
Давно известно, что переносчики ауксина (белки семейства PIN) показывают, куда гормон пойдет, а куда он пойдет — там корень и разовьется.
Итак, ауксин, попадая в гипофизу, начинает действовать через транскрипционный фактор MONOPTEROS (MP), активируя промоторы, содержащие ауксин-регулируемые cis-элементы (auxin-responsive cis-acting elements, AuxREs) [3], и запуская работу факторов транскрипции WOX5 и NTT (NO TRANSMITTING TRACT) (рис. 3). AuxREs-содержащие промоторы — например, DR5 — используются в репортерных системах для детекции ауксина, белок WOX5 также хорошо изучен, а вот механизмы работы NTT были приоткрыты авторами статьи [4] совсем недавно.
Оказалось, что мутация в этом гене не приводила к изменению фенотипа. Представьте, как странно — растению всё равно, работает ген или нет. Отсутствие мутантного фенотипа означает, что кодирование избыточно — есть еще гены «на подстраховке». Эти гены — WIP4 и WIP5 — кодируют белки из семейства WIP DOMAIN PROTEIN. Мутации по каждому в отдельности также не приводят к каким-либо драматическим последствиям. Только тройной мутант — мутант, у которого выключены все три гена (NTT, WIP4, WIP5), не развивал-таки корней. Коротко такой фенотип был назван nww (рис. 4).
Эти гены — WIP4 и WIP5 — кодируют белки из семейства WIP DOMAIN PROTEIN. Мутации по каждому в отдельности также не приводят к каким-либо драматическим последствиям. Только тройной мутант — мутант, у которого выключены все три гена (NTT, WIP4, WIP5), не развивал-таки корней. Коротко такой фенотип был назван nww (рис. 4).
Рисунок 4. Нормальное (WT) и мутантное (nww) растения Arabidopsis thaliana. а — Внешний вид. Чтобы помешать развитию корня, необходимо, чтобы мутировали три гена одновременно (NTT, WIP4, WIP5). б — Распределение ауксина в корне нормального и дефектного растения. О локализации гормона судят по зеленой флуоресценции от репортерной системы pDR5-GFP (описана под рисунком 3). Обратите внимание, как отличается нормальное распределение ауксина у WT-растения (слева) от «беспорядочного» у nww-растения (справа).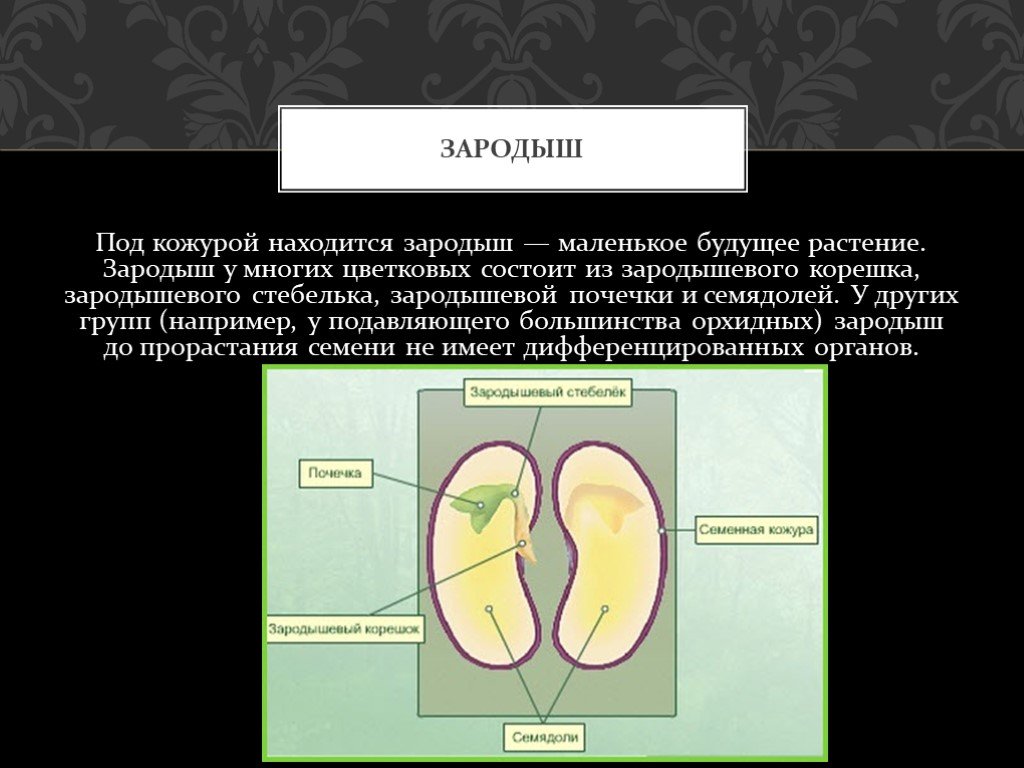 Рисунок из [4].
Рисунок из [4].
Интересно, что где ген NTT начнет работать, там и разовьется корень. Авторы «заставили» экспрессироваться NTT на месте семядольных листьев, и там развились слабенькие корешки (рис. 5).
Рисунок 5. Нормальный проросток A. thaliana и проросток с геном NTT, работающим в семядолях. Видно, что экспрессия этого гена обеспечивает закладку корней: у мутанта (справа) появились два корешка. Рисунок из [4].
А что же нового показали для апикальной меристемы побега?
Для белков семейств WUS/WOX была продемонстрирована связь с регуляторами семейства HAIRY MERISTEM (HAM). Авторы показали возможность взаимодействия этих белков с помощью дрожжевой двугибридной системы, выявили их согласованное действие на апикальную меристему побега и ко-регуляцию сходного набора генов, а также экспрессию в одних клетках (для этого к их мРНК «пришивали» флуоресцентные метки). Кроме того, фенотипы мутантов wox4 и ham1;2;3;4 были сходны: нет главных факторов, регулирующих меристему — белков семейства WUS/WOX, — а значит, и меристемы нет, то есть расти нечем.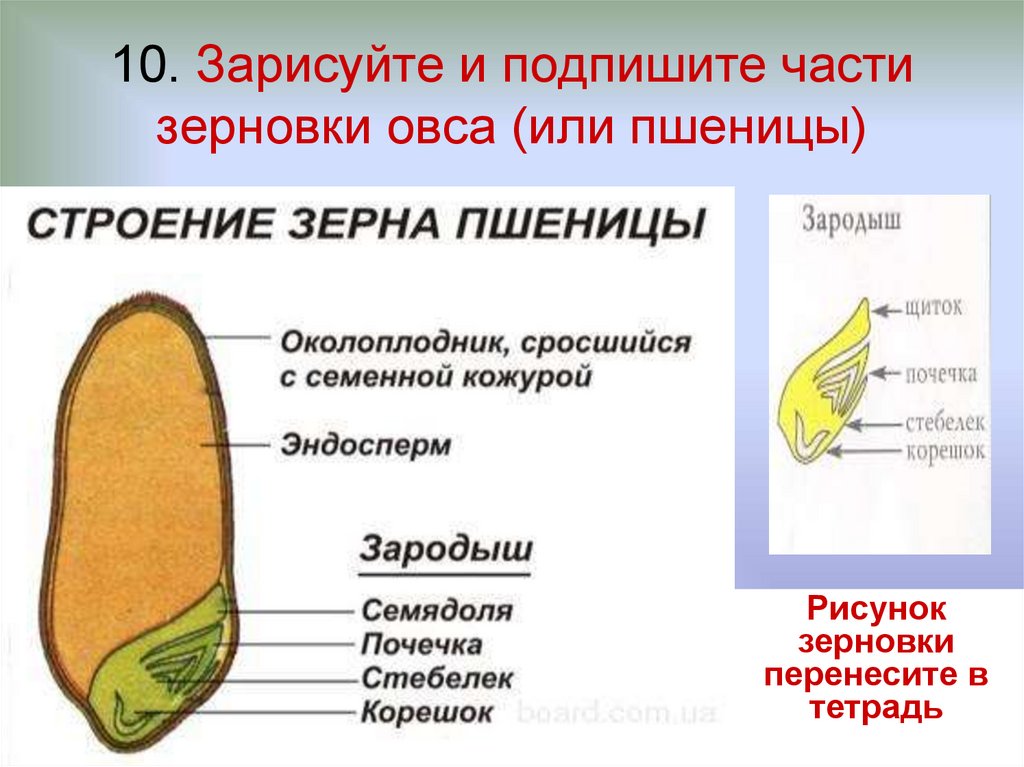 Действительно, даже спустя почти четыре недели после прорастания мутантные растения так и оставались внешне «свежепроклюнувшимися проростками» (рис. 6). Авторы предположили, что HAM — кофакторы белков WOX и тоже важны для становления апикальной меристемы как побега, так и корня [5]. В корне мутация ham1;2;3 влияет как минимум на закладку колумеллы (рис. 7).
Действительно, даже спустя почти четыре недели после прорастания мутантные растения так и оставались внешне «свежепроклюнувшимися проростками» (рис. 6). Авторы предположили, что HAM — кофакторы белков WOX и тоже важны для становления апикальной меристемы как побега, так и корня [5]. В корне мутация ham1;2;3 влияет как минимум на закладку колумеллы (рис. 7).
Рисунок 6. Нормальный побег (WT) и четверной мутант A. thaliana по генам HAM через 26 дней после прорастания. Так как апикальной меристемы нет, расти мутанту нечем. Из листьев у него будут только два семядольных, а проводящая система (сосуды ксилемы, транспортирующие воду, и клетки флоэмы, транспортирующие сахара) будет развита слабо. Рисунок из [5].
Рисунок 7. Гены HAM влияют и на апикальную меристему корня. Покоящийся центр меристемы показан на срезах звездочкой. Как видно, у растения дикого типа (слева) развиваются красивые, кубические меристематические клетки колумеллы (на них указывает стрелка), а у мутантного на том же месте — увеличенные клетки неправильной формы.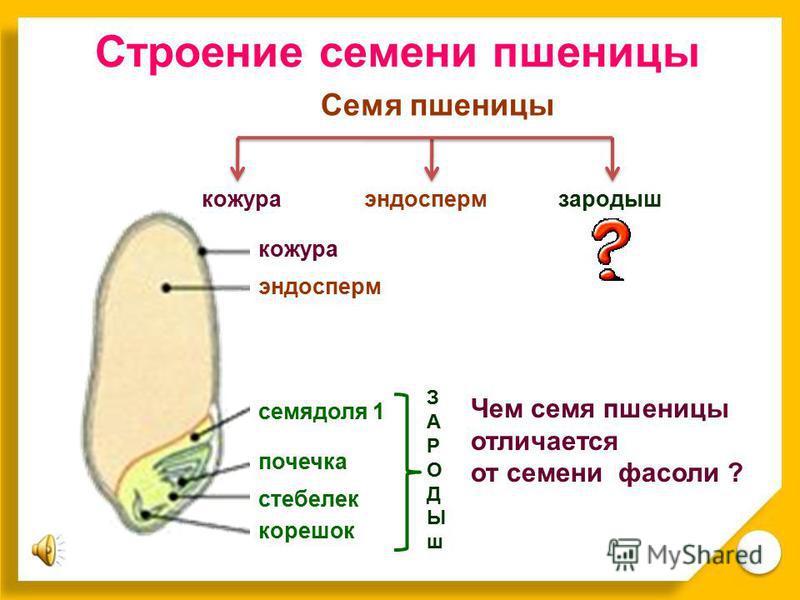 Этот эффект очень похож на мутацию wox5, но выражен серьезнее, что позволяет предположить, что HAM действуют и через WOX5, и через некий WOX5-независимый путь. Рисунок из [5].
Этот эффект очень похож на мутацию wox5, но выражен серьезнее, что позволяет предположить, что HAM действуют и через WOX5, и через некий WOX5-независимый путь. Рисунок из [5].
Таким образом, закладка меристем — критический период в формировании зародыша. Это понятно, ведь если не заложится меристема корня, то не будет и корня, растению нечем будет закрепляться в почве и всасывать воду; не заложится меристема побега — не быть листьям и стеблю со всей его сложной проводящей и опорной системой. Именно поэтому в эволюции сформировались сложные системы «подстраховок» белков друг другом, избыточность кодирования: только тройные (nww) и четверные (ham1;2;3;4) мутанты (то есть те, у которых выключены сразу несколько регуляторных генов) полностью теряли меристему. Исследования в биологии развития растений могут помочь селекционерам «манипулировать» корнем и побегом для повышения урожайности.
- Уточнен механизм, по которому развиваются конечности у куриного эмбриона;
- Дай пять: как математика управляет развитием пальцев;
- http://biomolecula.
 ru#;
ru#; - Crawford B.C.W., Sewell J., Golembeski G., Roshan C., Long J.A., Yanofsky M.F. (2015). Genetic control of distal stem cell fate within root and embryonic meristems. Science. 347 (6222), 655–659;
- Zhou Y., Liu X., Engstrom E.M., Nimchuk Z.L., Pruneda-Paz J.L., Tarr P.T. et al. (2015). Control of plant stem cell function by conserved interacting transcriptional regulators. Nature. 517 (7534), 377–380..
 ru#;
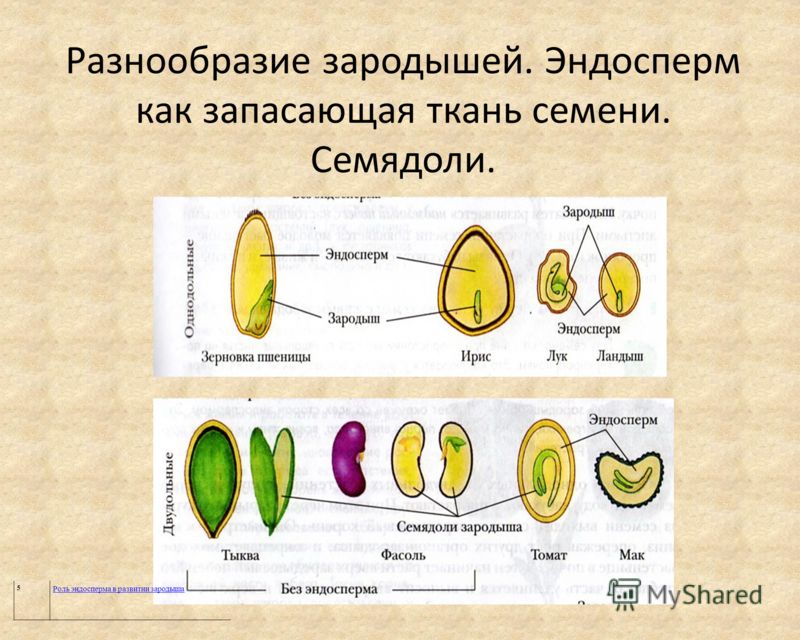
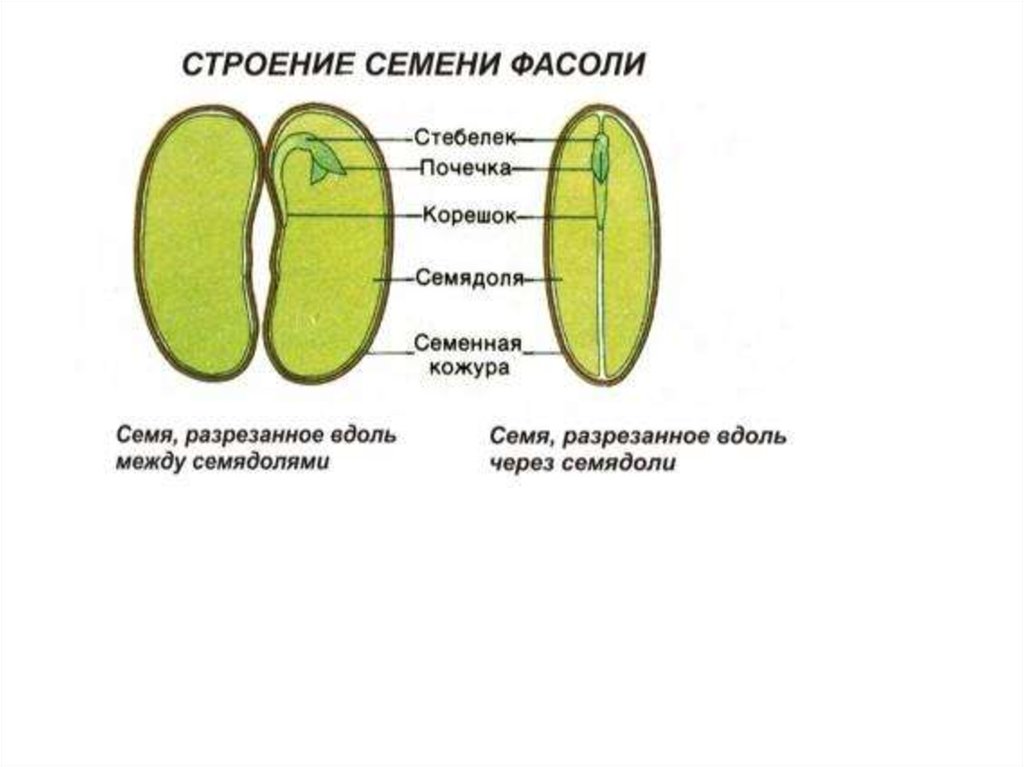
ru#;Семя
орган полового размножения и расселения растений (голосеменных растений и покрытосеменных растений).
Представляет собой оплодотворенную, созревшую семяпочку, состоящую из зародыша, запасающих тканей (эндосперма, перисперма) и семенной оболочки (спермодермы). Иногда развивается без оплодотворения.
Строение семени сосны: 1 — вид зародыша; 2 – продольный разрез семени; ск — семенная кожура; см — семядоли; з — зародыш; гк — гипокотиль; к — корешок; э – эндосперм. По степени морфологической дифференциации зародыши можно подразделить на 3 группы: хорошо дифференцированные, недифференцированные и рудиментарные. Дифференцированный зародыш имеет зачатки всех вегетативных органов будущего растения: зародышевый корень, стебель (гипокотиль), семядоли- первые листья. У растений с недифференцированными (недоразвитыми) зародышами их дальнейшее развитие и увеличение размеров происходит внутри семя, но после отделения его от материнского организма и попадания в благоприятную среду. Рудиментарные (редуцированные) зародыши, обычно представленные небольшой группой клеток, не дифференцируются и ко времени прорастания семян.
По степени морфологической дифференциации зародыши можно подразделить на 3 группы: хорошо дифференцированные, недифференцированные и рудиментарные. Дифференцированный зародыш имеет зачатки всех вегетативных органов будущего растения: зародышевый корень, стебель (гипокотиль), семядоли- первые листья. У растений с недифференцированными (недоразвитыми) зародышами их дальнейшее развитие и увеличение размеров происходит внутри семя, но после отделения его от материнского организма и попадания в благоприятную среду. Рудиментарные (редуцированные) зародыши, обычно представленные небольшой группой клеток, не дифференцируются и ко времени прорастания семян.
У двудольных растений обычно 2 семядоли. Зародыши некоторых растений (яблони, клена, люпина) имеют 3-4 семядоли (поликотилия), развивающиеся, вероятно, вследствие раннего расщепления их зачатков. Зародыш может быть и односемядольным (у некоторых зонтичных, лютиковых). У некоторых растений паразитов со слабо дифференцированными зародышами семени долей может вообще не быть.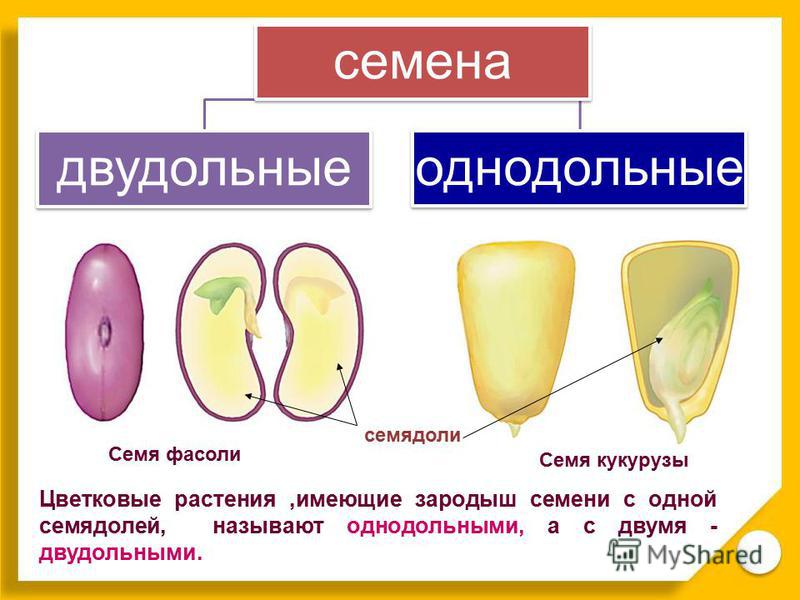 Чаще всего семядоли целые, реже — расчлененные (у липы). По форме зародыши двудольных могут быть прямыми, согнутыми, спиральнозакрученными, подковообразными и др. Для однодольных растений характерна одна семядоля.
Чаще всего семядоли целые, реже — расчлененные (у липы). По форме зародыши двудольных могут быть прямыми, согнутыми, спиральнозакрученными, подковообразными и др. Для однодольных растений характерна одна семядоля.
Форма, окраска и размеры семя сильно варьируют, в большинстве случаев они шаровидные, шаровидно удлиненные, цилиндрические. Очень крупные семена (до50 см в длину и массой до 20-25 кг) встречаются у некрых пальм. Пылевидные семена характерны для орхидных.
Форма, размеры и масса семени у растений одного вида чаще остаются постоянными, но иногда встречается разносемянность (гетероспермия) — образование семян, отличающихся по внешнему виду и по времени прорастания.
Очень разнообразны характер поверхности семени (гладкая, морщинистая, ребристая, бороздчатая и др.) и окраска. Семена некоторых растений имеют придатки в виде мясистых, обычно яркоокрашенных выростов, богатых разными веществами (жирными маслами, белками, крахмалом), привлекающими животных. Форма, размер, окраска и др.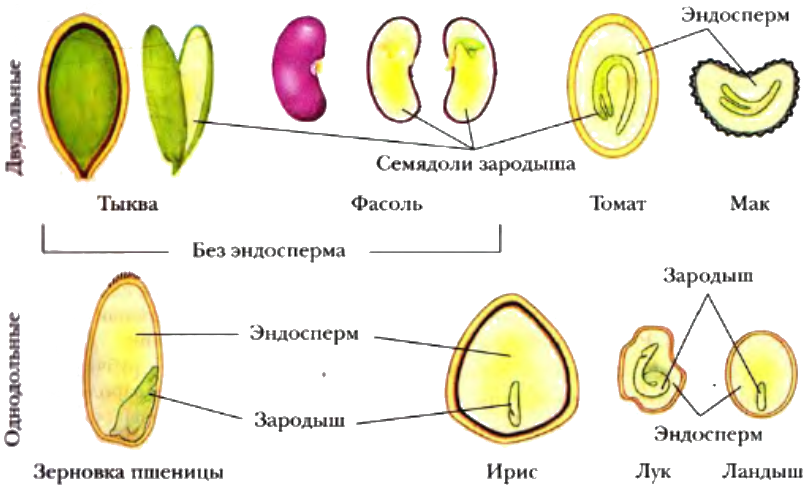
Особенности семян — диагностические признаки, широко используемые в систематике растений.
У покрытосеменных растений семена находятся в плодах, где их число колеблется от одного (абрикос, вишня, слива) до многих тысяч (маки, орхидные). Лишь у немногих растений семя после отделения от материнского растения сразу прорастают, в большинстве случаев семя сначала переходит в состояние разного по длительности физиологического покоя. Очень глубокий продолжительный покой характерен для семени с толстой кутикулой. Такие семена долго сохраняют всхожесть.
Значение семени в жизни человека очень велико. Они широко используются в самых разных отраслях промышленности (пищевой, медицинской, кондитерской, парфюмерной и др.) и сельского хозяйства (в виде кормов для животных, посевного материала и т. д.).
Лит.: Лотова, Л. И. Морфология и анатомия высших растений : учеб. пособ. — М., 2001.
Что в семени? — 1 марта 2017 г. Джефф Шалау, агент по сельскому хозяйству и природным ресурсам Кооперативное расширение Университета Аризоны, графство Явапай, Каталоги семян уже поступили в продажу, и садовники планируют свои огороды и клумбы. Когда вы визуализируете будущие цветы и плоды, подумайте о сложности и элегантности самих семян. Семена — это зародыши растений, упакованные в небольшой контейнер с хранимой пищей. Мать-природа обеспечивает технологию и упаковку. Семеноводческие компании разводят растения, собирают и перерабатывают плоды, упаковывают их в конверт и продают нам. Семена – продукты полового размножения. Они содержат ДНК пыльцевого зерна (мужской родитель) и семязачатка (женский родитель). Мужские и женские родители могут быть разными растениями или одним и тем же растением в зависимости от вида. Как и во всех биологических системах, на эту тему существует множество вариаций. Понимание репродуктивных циклов различных растений имеет решающее значение для успешной селекции растений и сохранения семян. Как упоминалось выше, семя содержит зародыш нового растения с запасом пищи для зародыша до тех пор, пока он не сформирует достаточное количество корней и листьев для получения собственного питания. Эмбрион представляет собой миниатюрное растение в состоянии задержки развития. Пища зародыша, называемая эндоспермом, может находиться в листьях семени (также называемых семядолями) или вне листьев семени и поглощаться по мере прорастания семени. Источниками пищи для эндосперма могут быть углеводы, белки или жиры. Семена также имеют семенную оболочку (тесту), которая защищает их от болезней и насекомых. Семенная оболочка также предотвращает попадание воды в семена, что могло бы инициировать процесс прорастания раньше положенного времени. Процесс прорастания семян – это возобновление активного роста зародыша с тем, чтобы он мог стать растением. Прежде чем появятся какие-либо визуальные признаки роста, семя должно впитать воду через семенную оболочку и/или область семени, называемую микропиле. Прорастание семян происходит в три стадии. На первом этапе вода поглощается (называется впитыванием). Это реактивирует ферменты, присутствующие в семенах. Эти ферменты расщепляют запасные соединения в семенах, чтобы сделать их доступными для зародыша. Например, когда ячмень поглощает воду, активируются ферменты, превращающие крахмал в сахара. Некоторые трудолюбивые люди используют эти процессы в своих интересах для производства ячменного солода, который используется для приготовления пива. Второй этап — пищеварение и транслокация. На этой стадии ферменты, которые были синтезированы или активированы ранее, начинают расщеплять запасной материал внутри семени на простые соединения, которые перемещаются в зародыш. Третья стадия – возобновление роста: также называется прорастанием. Корешок часто является первой частью проростка, появляющейся из семени. Он разовьется в первичный корень, из которого развиваются корневые волоски и боковые корни. У большинства широколиственных растений есть два семенных листа (называемых семядолями), которые обычно отличаются по форме от настоящих листьев растения. Гипокотиль — это часть зародыша, которая становится стеблем. Эти растения часто называют двудольными растениями или двудольными. Растения, образующие одну семядолю, называются однодольными. Однодольные включают травы, лилии, орхидеи, пальмы, агавы и юкки. Описанные выше процессы упрощены и обобщены. Суть в том, что семена живые. Для дыхания требуется источник энергии (часто сахар или жир) и небольшое количество кислорода. Подпишитесь на Backyard Gardener в Твиттере, используйте ссылку на веб-сайте BYG. Если у вас есть другие вопросы по садоводству, позвоните в справочную службу Master Gardener в офисе Camp Verde по телефону 928-554-8992 или напишите нам по электронной почте [email protected] и обязательно укажите свое имя, адрес и номер телефона. Найдите прошлые колонки Backyard Gardener или оставьте отзыв на веб-сайте Backyard Gardener: http://cals.arizona.edu/yavapai/anr/hort/byg/. Дополнительные ресурсы Seed and Seedling Biology Размножение растений из семян |

 Кроме того, семена должны находиться в надлежащих условиях окружающей среды. Для большинства семян это наличие воды и кислорода и благоприятная температура почвы. Для других растений определенные условия освещения, жара, дым или другие факторы окружающей среды могут вызвать прорастание.
Кроме того, семена должны находиться в надлежащих условиях окружающей среды. Для большинства семян это наличие воды и кислорода и благоприятная температура почвы. Для других растений определенные условия освещения, жара, дым или другие факторы окружающей среды могут вызвать прорастание.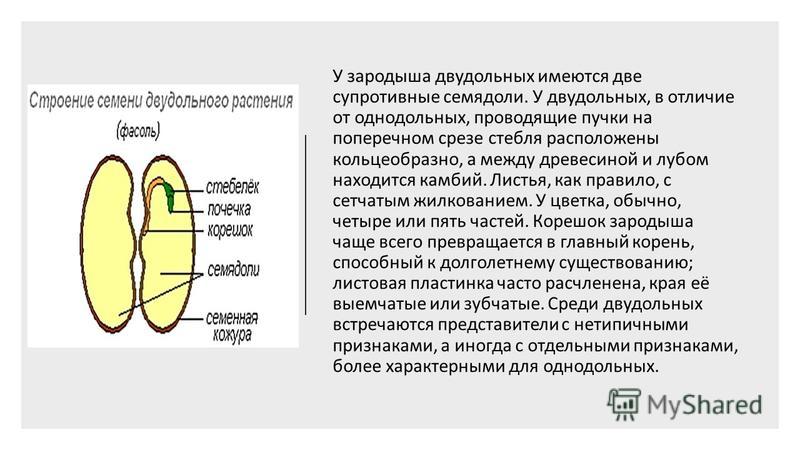 Эмбрион начинает расти по мере того, как клетки удлиняются и делятся. На заключительном этапе прорастающее семя продолжает претерпевать метаболические изменения, превращающие зародыш в проросток.
Эмбрион начинает расти по мере того, как клетки удлиняются и делятся. На заключительном этапе прорастающее семя продолжает претерпевать метаболические изменения, превращающие зародыш в проросток.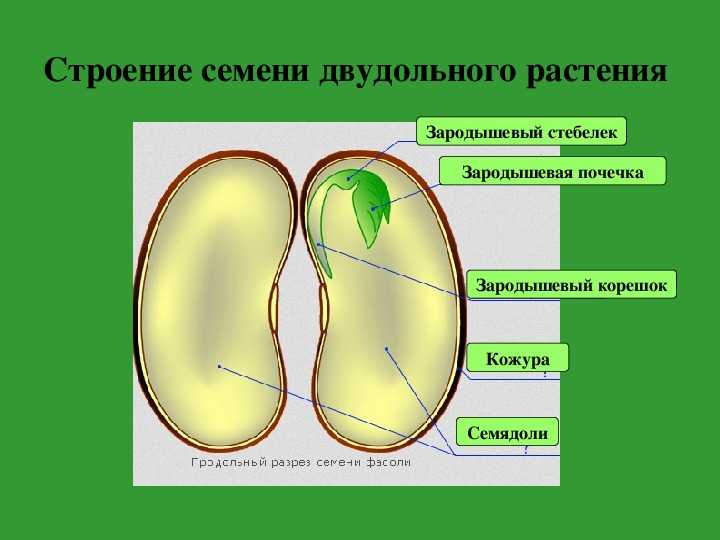 Жизнеспособность семян можно продлить в прохладных, сухих условиях хранения. Хотя есть много неподтвержденных сообщений о семенах, сохраняющих всхожесть в течение 1000 и более лет. Контролируемые эксперименты задокументировали жизнеспособность семян в течение 100 лет для нескольких исследованных видов. Лично я счастлив, когда семена, которые я купил в прошлом году, прорастают. Я также включил дополнительные ресурсы в онлайн-издание (см. URL-адрес ниже).
Жизнеспособность семян можно продлить в прохладных, сухих условиях хранения. Хотя есть много неподтвержденных сообщений о семенах, сохраняющих всхожесть в течение 1000 и более лет. Контролируемые эксперименты задокументировали жизнеспособность семян в течение 100 лет для нескольких исследованных видов. Лично я счастлив, когда семена, которые я купил в прошлом году, прорастают. Я также включил дополнительные ресурсы в онлайн-издание (см. URL-адрес ниже).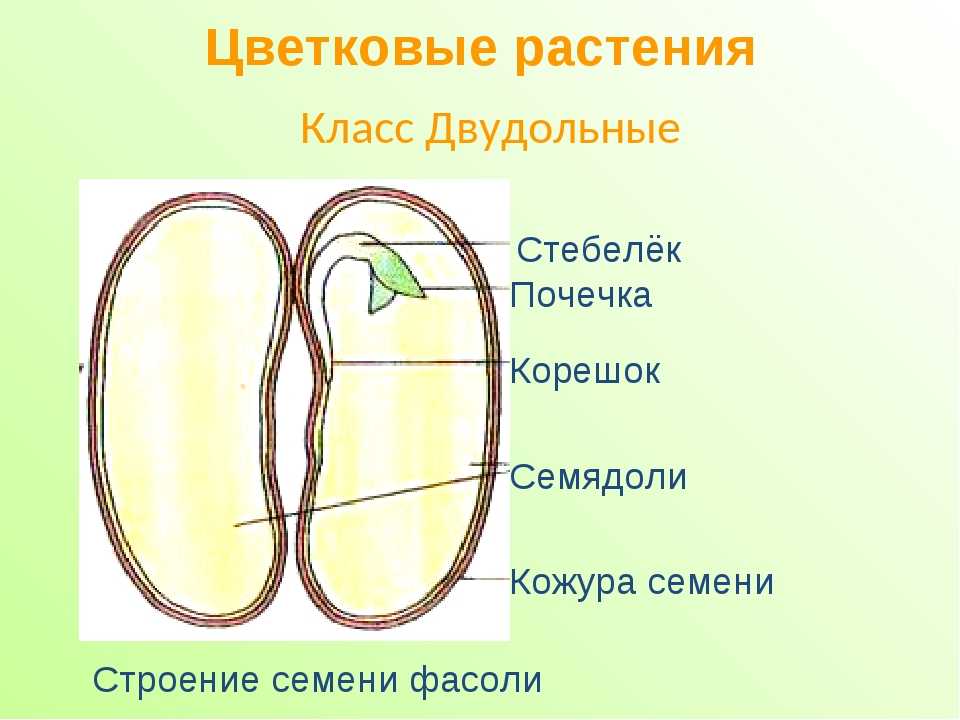 psu.edu/business/start-farming/vegetables/factsheets/seed-and-seedling-biology
psu.edu/business/start-farming/vegetables/factsheets/seed-and-seedling-biologyЭмбриональное развитие — биология развития
У растений термин эмбриогенез охватывает развитие с момента оплодотворения до наступления покоя. Основной план тела спорофита устанавливается во время эмбриогенеза; однако этот план повторяется и дорабатывается после выхода из состояния покоя. Основные проблемы эмбриогенеза:
- 1.
Установить основной план тела. Радиальное формирование паттерна создает три системы тканей, а осевое паттернирование устанавливает апикально-базальную (отросток-корень) ось.
- 2.
Для выделения меристематической ткани для постэмбрионального развития структуры тела (листья, корни, цветки и т.д.).
- 3.
Для создания доступного запаса пищи для прорастающего эмбриона до тех пор, пока он не станет автотрофным.

Эмбриогенез сходен у всех покрытосеменных с точки зрения установления основного плана тела (Steeves and Sussex 1989) (см. ). Однако существуют различия в развитии паттернов, в том числе различия в точности паттернов клеточного деления, степени развития эндосперма, развития семядолей и степени развития меристемы побегов (Исав 19).77; Джохри и др. 1992).
Рисунок 20.15
Эмбриогенез покрытосеменных. Показан репрезентативный двудольный; однодольное растение разовьет только одну семядолю. Хотя у покрытосеменных существуют основные модели эмбриогенеза, между видами существуют огромные морфологические различия.
Полярность устанавливается при первом клеточном делении после оплодотворения.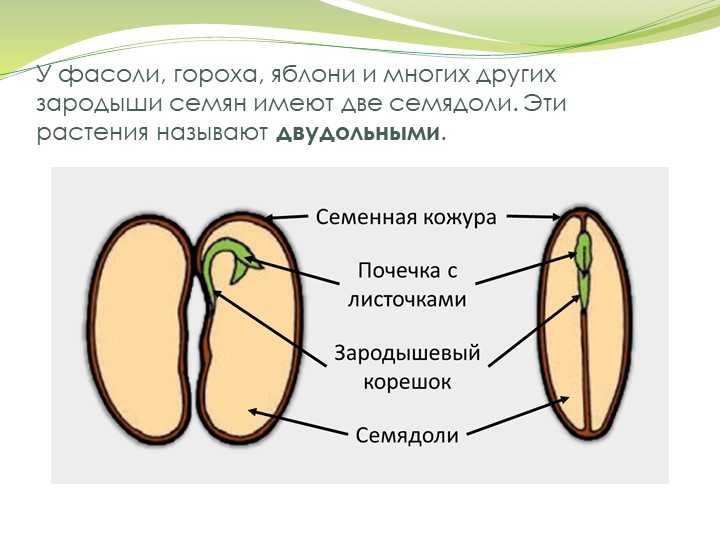 Установление полярности было исследовано с использованием бурых водорослей в качестве модельной системы (Belanger and Quatrano 2000). Зиготы этих растений не зависят от других тканей и поддаются манипуляциям. В результате первоначального клеточного деления образуется одна меньшая клетка, которая образует ризоид (гомолог корня) и закрепляет остальную часть растения, и одна большая клетка, дающая начало слоевищу (основному телу спорофита). Точка входа спермия фиксирует положение ризоидного конца апикально-базальной оси. Эта ось перпендикулярна плоскости первого клеточного деления. F-актин накапливается на ризоидном полюсе (Kropf et al. 19).99). Однако свет или гравитация могут отменить эту фиксацию оси и установить новое положение для клеточного деления (Alessa and Kropf 1999). Как только апикально-базальная ось установлена, секреторные везикулы направляются к ризоидному полюсу зиготы. Эти везикулы содержат материал для разрастания ризоидов с клеточной стенкой определенного макромолекулярного состава. Целенаправленная секреция может также помочь сориентировать первую плоскость клеточного деления. Сохранение судьбы ризоидов и талломов на ранних стадиях развития зависит от информации в клеточных стенках (Браунли и Бергер 19).95). Информация о клеточных стенках также важна для покрытосеменных растений (rev. Scheres and Benfey 1999).
Установление полярности было исследовано с использованием бурых водорослей в качестве модельной системы (Belanger and Quatrano 2000). Зиготы этих растений не зависят от других тканей и поддаются манипуляциям. В результате первоначального клеточного деления образуется одна меньшая клетка, которая образует ризоид (гомолог корня) и закрепляет остальную часть растения, и одна большая клетка, дающая начало слоевищу (основному телу спорофита). Точка входа спермия фиксирует положение ризоидного конца апикально-базальной оси. Эта ось перпендикулярна плоскости первого клеточного деления. F-актин накапливается на ризоидном полюсе (Kropf et al. 19).99). Однако свет или гравитация могут отменить эту фиксацию оси и установить новое положение для клеточного деления (Alessa and Kropf 1999). Как только апикально-базальная ось установлена, секреторные везикулы направляются к ризоидному полюсу зиготы. Эти везикулы содержат материал для разрастания ризоидов с клеточной стенкой определенного макромолекулярного состава. Целенаправленная секреция может также помочь сориентировать первую плоскость клеточного деления. Сохранение судьбы ризоидов и талломов на ранних стадиях развития зависит от информации в клеточных стенках (Браунли и Бергер 19).95). Информация о клеточных стенках также важна для покрытосеменных растений (rev. Scheres and Benfey 1999).
Рисунок 20.13
Формирование оси у бурой водоросли Pelvetia Compressa. (A) Пятно F-актина (оранжевое) сначала образуется в месте входа сперматозоида (синее пятно отмечает пронуклеус сперматозоида). (B) Позже в направлении стрелки засиял свет. Ось, индуцированная сперматозоидами (подробнее…)
Рисунок 20.14
Асимметричное деление клеток у бурых водорослей. Временная динамика от 8 до 25 часов после оплодотворения, показывающая клетки водорослей, окрашенные жизненным мембранным красителем для визуализации секреторных пузырьков, которые появляются первыми, и клеточной пластинки, которая начинает появляться примерно (подробнее.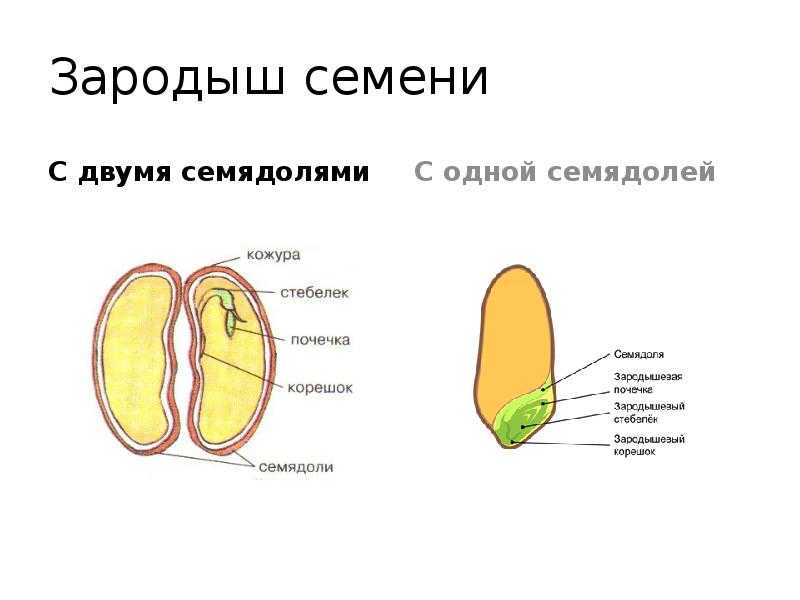 ..)
..)
Основной план тела покрытосеменных, заложенный во время эмбриогенеза, также начинается с асимметричного деления клеток * , дающего начало терминальной клетке и базальной клетке (). Терминальная клетка дает начало собственно эмбриону . Базальная клетка формируется ближе всего к микропиле и дает начало суспензору . Гипофиз находится на границе между суспензором и собственно эмбрионом. У многих видов он дает начало некоторым корневым клеткам. (Суспензорные клетки делятся с образованием нитевидного или сферического органа, который дегенерирует позже в эмбриогенезе.) И у голосеменных, и у покрытосеменных суспензор ориентирует абсорбирующую поверхность зародыша в сторону источника пищи; у покрытосеменных он также, по-видимому, служит проводником питательных веществ для развивающегося эмбриона. Культивирование изолированных зародышей алой фасоли с суспензором и без него продемонстрировало потребность в суспензоре на стадии сердцевины у двудольных (Yeung and Sussex 19).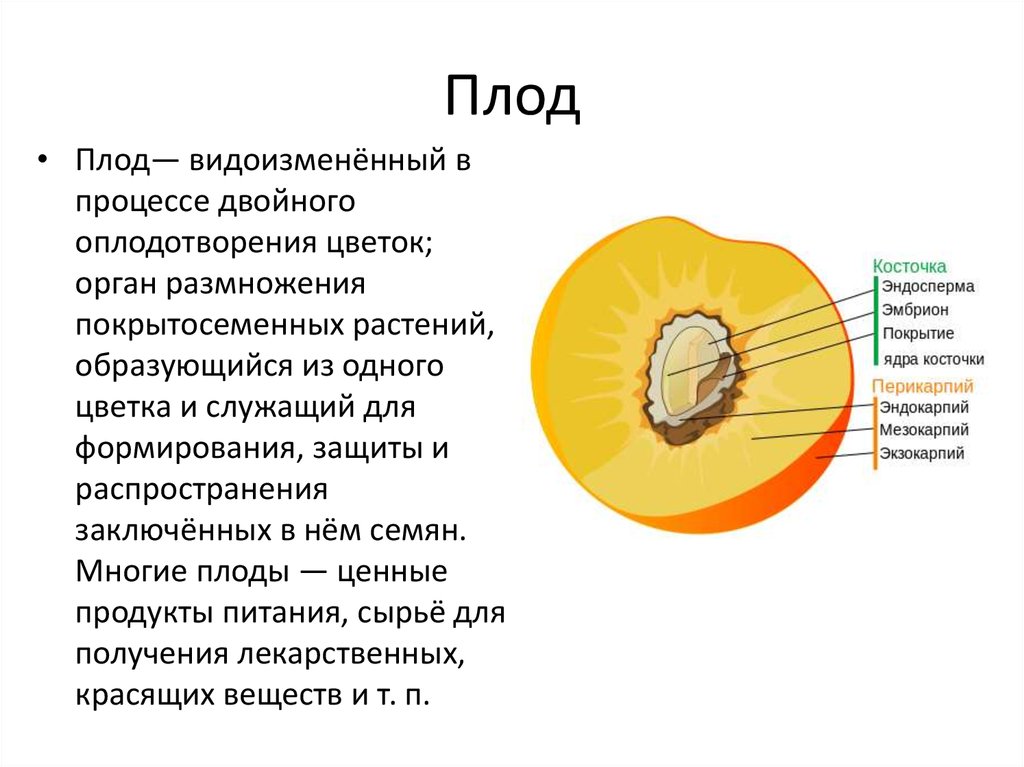 79). Эмбрионы, культивированные с суспензором, имеют в два раза больше шансов выжить, чем эмбрионы, культивированные без прикрепленного суспензора на этой стадии. Суспензор может быть источником гормонов. У алой побеговой фасоли молодые зародыши без суспензора могут выжить в культуре, если к ним добавляют гормон роста гибберелловую кислоту (Cionini et al., 1976).
79). Эмбрионы, культивированные с суспензором, имеют в два раза больше шансов выжить, чем эмбрионы, культивированные без прикрепленного суспензора на этой стадии. Суспензор может быть источником гормонов. У алой побеговой фасоли молодые зародыши без суспензора могут выжить в культуре, если к ним добавляют гормон роста гибберелловую кислоту (Cionini et al., 1976).
Рисунок 20.16
Роль суспензора в эмбриогенезе двудольных. Культивирование зародышей фасоли алой с суспензором и без него показало, что суспензор необходим на стадии сердцевидной формы, но не позже. (После Юнга и Сассекса 1979.)
Поскольку установление апикально-базальной полярности является одним из ключевых достижений эмбриогенеза, полезно рассмотреть, почему собственно суспензор и зародыш развивают уникальные морфологии. Здесь особенно полезным оказалось изучение эмбриональных мутантов кукурузы и Arabidopsis . Исследования суспензорных мутантов ( sus1,
sus2, и raspberry1 ) из Arabidopsis предоставили генетические доказательства того, что суспензор обладает способностью развивать эмбрионоподобные структуры (; Schwartz et al. 1994; Ядегари и др. 1994). У этих мутантов аномалии собственно эмбриона появляются до аномалий суспензора. † Более ранние эксперименты, в которых удаляли собственно эмбрион, также показали, что суспензоры могут развиваться подобно эмбрионам (Haccius 1963). Сигнал от эмбриона, свойственный суспензору, может быть важен для поддержания идентичности суспензора и блокирования развития суспензора как эмбриона. Молекулярный анализ этих и других генов позволяет понять механизмы коммуникации между суспензором и собственно эмбрионом.
1994; Ядегари и др. 1994). У этих мутантов аномалии собственно эмбриона появляются до аномалий суспензора. † Более ранние эксперименты, в которых удаляли собственно эмбрион, также показали, что суспензоры могут развиваться подобно эмбрионам (Haccius 1963). Сигнал от эмбриона, свойственный суспензору, может быть важен для поддержания идентичности суспензора и блокирования развития суспензора как эмбриона. Молекулярный анализ этих и других генов позволяет понять механизмы коммуникации между суспензором и собственно эмбрионом.
Рисунок 20.17
Ген SUS подавляет эмбриональное развитие в суспензоре. (A) Эмбрион дикого типа и суспензор. (B) мутант sus с суспензором, развивающимся как эмбрион (стрелка). (C) Модель, показывающая, как собственно эмбрион подавляет эмбриональное развитие в суспензоре (подробнее…)
Гены материнского эффекта играют ключевую роль в установлении эмбрионального паттерна у животных (см. главу 9). Роль экстразиготных генов в эмбриогенезе растений менее ясна, и вопрос осложняется как минимум тремя потенциальными источниками влияния: спорофитной тканью, гаметофитной тканью и полиплоидным эндоспермом. Все эти ткани тесно связаны с яйцеклеткой/зиготой (луч 19).98). На развитие эндосперма также могут влиять материнские гены. Гены спорофитного и гаметофитного материнского эффекта были идентифицированы у Arabidopsis, , и вполне вероятно, что геном эндосперма также влияет на зиготу. Первый идентифицированный ген материнского эффекта, SHORT INTEGUMENTS 1 ( SIN1 ), должен экспрессироваться в спорофите для нормального эмбрионального развития (Ray et al. 1996). Два фактора транскрипции (FBP7 и FBP11) необходимы в спорофите петунии для нормального развития эндосперма (Columbo et al. 19).97). Ген материнского эффекта женского гаметофита, MEDEA (в честь Медеи Еврипида, убившей своих собственных детей), имеет белковые домены, сходные с доменами гена материнского эффекта дрозофилы (Grossniklaus et al. 1998). Любопытно, что MEDEA относится к группе генов Polycomb (см. главу 9), продукты которых прямо или косвенно изменяют хроматин и влияют на транскрипцию.
Все эти ткани тесно связаны с яйцеклеткой/зиготой (луч 19).98). На развитие эндосперма также могут влиять материнские гены. Гены спорофитного и гаметофитного материнского эффекта были идентифицированы у Arabidopsis, , и вполне вероятно, что геном эндосперма также влияет на зиготу. Первый идентифицированный ген материнского эффекта, SHORT INTEGUMENTS 1 ( SIN1 ), должен экспрессироваться в спорофите для нормального эмбрионального развития (Ray et al. 1996). Два фактора транскрипции (FBP7 и FBP11) необходимы в спорофите петунии для нормального развития эндосперма (Columbo et al. 19).97). Ген материнского эффекта женского гаметофита, MEDEA (в честь Медеи Еврипида, убившей своих собственных детей), имеет белковые домены, сходные с доменами гена материнского эффекта дрозофилы (Grossniklaus et al. 1998). Любопытно, что MEDEA относится к группе генов Polycomb (см. главу 9), продукты которых прямо или косвенно изменяют хроматин и влияют на транскрипцию.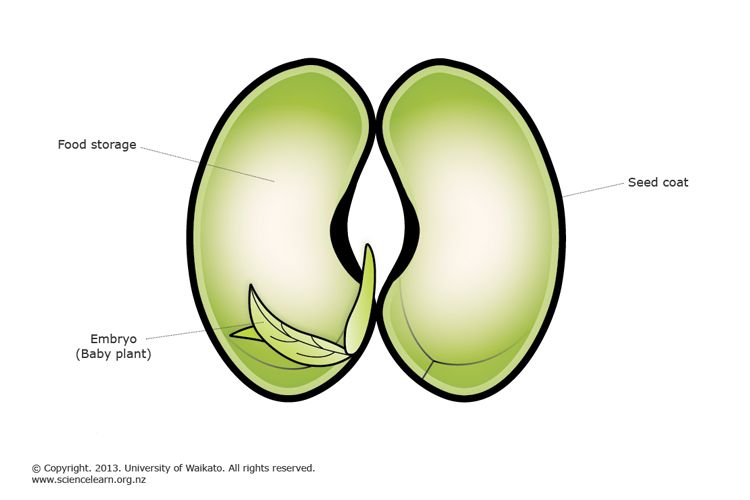 MEDEA воздействует на импринтированный ген (см. главу 5), который экспрессируется женским гаметофитом и аллелями, унаследованными от матери в зиготе, но не аллелями, унаследованными от отца (Vielle-Calzada et al. 19).99). Насколько значимы гены материнского эффекта в установлении плана тела спорофита, все еще остается без ответа.
MEDEA воздействует на импринтированный ген (см. главу 5), который экспрессируется женским гаметофитом и аллелями, унаследованными от матери в зиготе, но не аллелями, унаследованными от отца (Vielle-Calzada et al. 19).99). Насколько значимы гены материнского эффекта в установлении плана тела спорофита, все еще остается без ответа.
Радиальные и осевые паттерны развиваются по мере продолжения клеточного деления и дифференцировки (см. также Bowman 1994 для подробных световых микрофотографий эмбриогенеза Arabidopsis ). Клетки собственно зародыша делятся в поперечной и продольной плоскостях, образуя зародыш глобулярной стадии с несколькими ярусами клеток. Внешне эта стадия имеет некоторое сходство с дроблением у животных, но ядерно-цитоплазматическое отношение не обязательно увеличивается. Формирующаяся форма зародыша зависит от регуляции плоскостей клеточного деления и расширения, так как клетки не способны двигаться и изменять форму зародыша. Плоскости клеточного деления во внешнем слое клеток становятся ограниченными, и этот слой, называемый протодерма , становится отчетливой. Радиальный паттерн появляется на глобулярной стадии, когда инициируются три тканевые системы (дермальная, основная и сосудистая) растения. Кожная ткань (эпидермис) будет формироваться из протодермы и вносить вклад в внешние защитные слои растения. Основная ткань (кора и сердцевина) образуется из основной меристемы, лежащей под протодермой. Прокамбий , образующийся в ядре зародыша, даст начало сосудистая ткань (ксилема и флоэма), которая будет поддерживать и транспортировать. Дифференцировка каждой тканевой системы по крайней мере частично независима. Например, у мутанта keule Arabidopsis, кожная система дефектна, в то время как внутренние тканевые системы развиваются нормально (Mayer et al. 1991).
Радиальный паттерн появляется на глобулярной стадии, когда инициируются три тканевые системы (дермальная, основная и сосудистая) растения. Кожная ткань (эпидермис) будет формироваться из протодермы и вносить вклад в внешние защитные слои растения. Основная ткань (кора и сердцевина) образуется из основной меристемы, лежащей под протодермой. Прокамбий , образующийся в ядре зародыша, даст начало сосудистая ткань (ксилема и флоэма), которая будет поддерживать и транспортировать. Дифференцировка каждой тканевой системы по крайней мере частично независима. Например, у мутанта keule Arabidopsis, кожная система дефектна, в то время как внутренние тканевые системы развиваются нормально (Mayer et al. 1991).
Рисунок 20.18
Радиальное и осевое формирование. (A) Радиальное формирование паттерна у покрытосеменных начинается на глобулярной стадии и приводит к установлению трех тканевых систем. (B) Осевой паттерн (ось отростка-корня) устанавливается стадией сердца.
Шаровидная форма зародыша теряется, когда начинают формироваться семядолей («первые листья»). Двудольные имеют две семядоли, которые при формировании зародыша придают ему форму сердца. Осевой план тела очевиден на этой сердечной стадии развития. Гормоны (в частности, ауксины) могут опосредовать переход от радиальной к билатеральной симметрии (Liu et al. 1993). У однодольных, таких как кукуруза, появляется только одна семядоля.
У многих растений семядоли помогают питать растение, становясь фотосинтезирующими после прорастания (хотя у некоторых видов семядоли никогда не появляются из земли). В некоторых случаях, например у гороха, пищевые запасы в эндосперме расходуются еще до прорастания, и источником питания для прорастающего проростка служат семядоли. ‡ Даже при наличии стойкого эндосперма (как у кукурузы) семядоли хранят пищевые запасы, такие как крахмал, липиды и белки. У многих однодольных семядоля вырастает в большой орган, прижатый к эндосперму, и способствует переносу питательных веществ к проростку.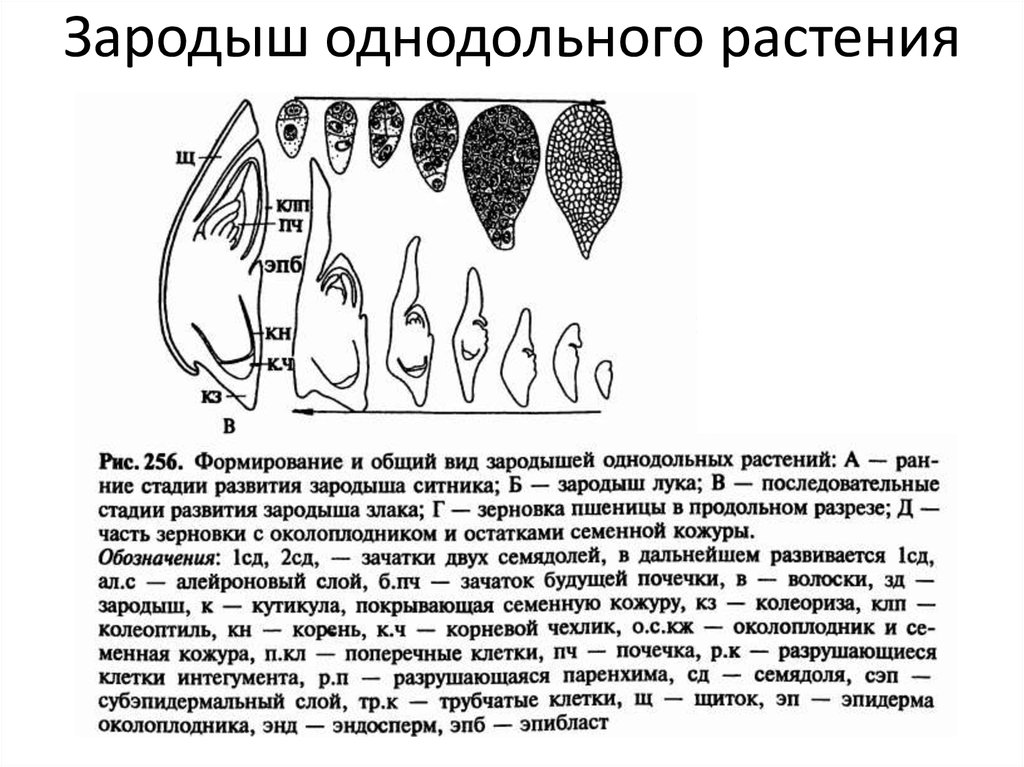 Вертикальные семядоли могут придавать зародышу торпедообразную форму. У некоторых растений семядоли вырастают настолько длинными, что им приходится изгибаться, чтобы поместиться в пределах семенной кожуры. Тогда эмбрион выглядит как трость. К этому моменту подвеска дегенерирует.
Вертикальные семядоли могут придавать зародышу торпедообразную форму. У некоторых растений семядоли вырастают настолько длинными, что им приходится изгибаться, чтобы поместиться в пределах семенной кожуры. Тогда эмбрион выглядит как трость. К этому моменту подвеска дегенерирует.
Апикальная меристема побега и апикальная меристема корня представляют собой кластеры стволовых клеток, которые сохраняются в постэмбриональном растении и дают начало большей части тела спорофита. У некоторых видов корневая меристема частично происходит из гипофиза. Все остальные части тела спорофита происходят от собственно зародыша. Генетические данные указывают на то, что образование побегов и корневых меристем регулируется независимо. Эту независимость демонстрируют dek23 мутант кукурузы и без побегов ( STM ) мутант Arabidopsis, , оба из которых образуют корневую меристему, но не могут инициировать меристему побега (Clark and Sheridan 1986; Barton and Poethig 1993). Ген STM , который был клонирован, экспрессируется на поздней глобулярной стадии, до образования семядолей. Также были идентифицированы гены, которые специфически влияют на развитие корневой оси во время эмбриогенеза. Мутации Ген HOBBIT в Arabidopsis (Willemsen et al. 1998), например, влияют на производные гипофиза и устраняют функцию корневой меристемы.
Верхушечная меристема побега будет инициировать листья после прорастания и, в конечном итоге, перехода к репродуктивному развитию. У арабидопсиса , семядоли образуются из общей зародышевой ткани, а не из меристемы побега (Barton and Poethig 1993). У многих покрытосеменных растений во время эмбриогенеза закладывается несколько листьев. В случае 9Клональный анализ 0096 Arabidopsis, указывает на наличие листьев у зрелого зародыша, даже если они морфологически недостаточно развиты (Irish and Sussex 1992). Клональный анализ показал, что семядоли и два первых настоящих листа хлопчатника происходят из эмбриональной ткани, а не из организованной меристемы (Christianson 1986).