Растения и животные: параллельные миры
В Дарвиновском музее открылась выставка «Растения и животные:
параллельные миры». Тема выставки посвящена не только
биологическим аспектам, но и философским.
«Два мира: царство растений и царство животных. Флора и фауна —
такие разные и в то же время очень похожие местом происхождения и
единством генетического материала. Гармония меж двух миров
возникла, благодаря тому, что эволюция шла параллельными путями»,
— с приветственным словом обратилась к гостям музея
заведующая научно-просветительским отделом музея
Елена Прокофьевна Дубровская.
На языке музейных предметов авторскому коллективу, научным
сотрудникам и художникам удалось показать гармонию биологического
разнообразия.
«Наша выставка посвящена адаптации животных к условиям окружающей
среды. Мы проводим параллель между двумя царствами с целью
показать, что иногда они бывают удивительно похожи», — рассказала
куратор выставки и старший научный сотрудник музея Ирина
Александровна Елисейкина.
В экспозиции представлены «Мастера камуфляжа». «За миллионы лет
эволюции у растений и животных появились такие особенности
внешнего облика, которые помогают им стать «невидимками» или
походить на кого-то другого. Такое сходство организмов по окраске
и форме с другими защищенными видами или предметами окружающей
среды называется мимикрией».
Среди них животные, похожие на растения, — морской
конек-тряпичник, палочники, листохвостый мадагаскарский геккон,
клопы, богомолы, кузнечики и саранчовые. «Внешнее сходство
животных с растениями и предметами окружающей среды делает их
незаметными и позволяет им чувствовать себя в безопасности».
Растения тоже похожи на животных. Это различные виды офрисы:
оводоносная, шмелецветковая, насекомоносная и зеркальная. Цветки
некоторых орхидей выглядят и пахнут, как самки насекомых или
пауков. Самцы принимают цветки за самок и пытаются спариваться с
ними. Затем они перелетают с цветка на цветок и переносят пыльцу.
Происходит перекрестное опыление.
Следующая часть экспозиции — «Колючки и шипы у растений и
животных». Колючки и шипы растений отпугивают крупных животных, а
иглы животных — надежный способ обороны. Среди них растения:
синеголовник или эрингиум, мордовник и чертополох; и животные: еж
обыкновенный, ушастый еж, малый тенрек, толстоиглая крыса и
иглистая мышь.
«Осторожно — яд!» или ядовитые растения и животные. В этой части
экспозиции — представители «ядовитой флоры»: белена черная,
вороний глаз и вех ядовитый; и «ядовитой фауны»:
лягушка-древолаз, императорский скорпион и мангровая змея. «В
некоторых растениях содержатся неприятные на вкус ядовитые
вещества. Раз попробовав такое растение, животное больше не
захочет его есть, если останется в живых. Животные используют яд
не только для защиты, но и для нападения».
«Любители солененького» и экспонаты выставки — капский
буревестник, чернобровый альбатрос, фламинго Джеймса. Эти птицы
без вреда для здоровья пьют морскую воду. У буревестникообразных
У буревестникообразных
птиц выделяемая железами соленая жидкость выдувается мелкими
брызгами через носовые трубочки. У растений — франкении
порошистой, кермека бородавчатого, гребенщика изящного и авицении
морской — есть специальные железки на листьях или стеблях. С
помощью железок они выделяют насыщенный солями сок и тем самым
избавляются от излишков соли. Эти виды птиц и растений обитают в
местностях, где доступна только соленая вода: моря, океаны,
побережья и пустыни с засоленными почвами. Специальные
биомеханизмы позволяют удалять излишки соли.
«Наедине с пустыней» — животные и растения, обитающие в
засушливых регионах. В условиях дефицита воды животные и растения
вынуждены экономить воду.
Растения-суккуленты запасают влагу в толстых мясистых стеблях и
листьях. «Когда на них под углом светит солнце, ребра создают
тень на самом растении. В полдень освещены только верхушки живых
колонн или шаров. Это позволяет им не перегреваться».
Двугорбый верблюд хранит водные запасы не в горбах, а в крови.
Эритроциты животного впитывают воду, увеличивают свой объем в два
раза и отдают ее, когда верблюд испытывает жажду.
Хищная часть экспозиции — растения-мясоеды. О природе «растений с
характером» рассказал специалист по растениям-хищникам Сергей
Куницын:
«На выставке созданы два микроклимата для тропических и болотных
насекомоядных. В болотном климате — разные виды саррацений и
венериной мухоловки. В тропическом — росянки, жирянки и
непентесы. Микроклимат позволяет накормить растения, выпустив в
аквариум-травник мух. Самое главное условие для насекомоядных
растений — имитировать природную среду. Мы даже не догадываемся,
насколько много симбиозов в живой природе. Хищные растения
гармонируют со многими животными. Муравьи, птицы и лягушки
прочищают листья растений после того, как те не успели переварить
насекомых. Из непентесов обезьяны пьют воду».
Насекомоядным растениям досталась непригодная среда обитания:
почва бедная, кислая, заболоченная или песчаная. В таких условиях
В таких условиях
у них образовались ловчие листья, которые приманивают насекомых
ловушками.
«У одних, как у венериной мухоловки и росянки ловушки подвижны —
они захлопываются или сворачиваются, удерживая добычу. У других —
гелиамфоры и непентеса — неподвижные ловушки: жертва,
привлеченная ароматом, прилипает к ее поверхности или тонет в
резервуаре. Переваривается насекомое с помощью ферментов
пищеварительных железок растений».
Экспозицию «Растения и животные: параллельные миры» дополнили
работы Ассоциации художников ботанического искусства. Закрытие
выставки пройдет 4 ноября.
параллельные миры Государственный Дарвиновский музей
Расположение: Выставочные залы музея / 0 этаж (цоколь) / Выставочный зал №1
На новой выставке в
Дарвиновском музее вы окажетесь на пересечении двух «параллельных миров» — царства
растений и царства животных. Разнообразные представители флоры и фауны, живые
хищные растения, иллюстрации из коллекционных книг XIX века, изящные акварели и
рисунки современных художников откроют тайну, почему некоторые животные и
растения так стремятся быть похожими друг на друга.
Растения и животные различаются между собой кардинально — на
клеточном уровне. Но необходимость приспосабливаться даже к самым экстремальным
условиям привела к тому, что жители этих двух миров порой очень похожи друг на друга:
строением, образом жизни и даже внешностью. Выставка расскажет, что общего у
«корабля пустыни» верблюда и алоэ, галапагосской игуаны и мангрового дерева,
лотоса и изумрудного колибри, шиншиллы и эдельвейса.
За миллионы лет эволюции растения и животные научились
мимикрировать — изменять свою внешность, чтобы стать «невидимками» или походить
на кого-то другого. Наивысшего мастерства в маскировке достигли насекомые,
подражающие окраской и формой тела веточкам, колючкам, листьям и цветам.
Попробуйте разглядеть членистоногих гуру камуфляжа на рисунках художников В. Евстафьева
и А. Ватагиной из коллекции Дарвиновского музея. Среди растений своей мимикрией
славятся тропические орхидеи — они научились выглядеть и пахнуть как насекомые,
чтобы привлекать к своим цветкам опылителей. У каждой орхидеи – свой секретный
У каждой орхидеи – свой секретный
приём, что убедительно показано в изящных акварелях, созданных мастерами
Сообщества художников ботанической живописи и иллюстрации (SABA).
Ещё одни мастера притворства — морские кораллы и губки,
внешне похожие на растения. На самом деле, это настоящие животные, которые
ведут прикреплённый образ жизни, а для опоры и защиты используют твёрдый
минеральный скелет. Даже морские лилии — это не подводные цветы, а примитивные
иглокожие. Подробно рассмотреть их совершенное строение позволят иллюстрации из
редкой книги «Красота форм в природе» (1904 г.) немецкого дарвиниста Эрнста
Геккеля, соединившего искусство и науку в единое целое.
К отряду хищных относят около 280 видов вооружённых до зубов
млекопитающих. А вы знаете, что в природе существует более 600 видов растений-хищников?
Ловушки, капканы, ловчие ямы и запахи-обманки — у жертвы практически нет шансов
выжить. На всём протяжении работы выставки вы сможете наблюдать за ростом и
кормлением живых хищных растений — венериных мухоловок, росянок, непентесов и
других плотоядных видов из частной коллекции.
Выставка традиционно сопровождается образовательной программой для детей, молодёжи и взрослых.
Актуальные мероприятия
Стирая грань между растениями и животными
Автор: Джеймс Лонг
показывать/скрывать слова, чтобы знать
Хлорофилл: пигмент, придающий растениям зеленый цвет и позволяющий им поглощать солнечный свет… подробнее
Цианобактерии: вид бактерий, которые получают энергию посредством фотосинтеза… подробнее
Макрофаг: иммунная клетка, которая поглощает чужеродный материал и мертвые клетки… подробнее
Органелла: « маленький орган «. Внутренний орган клетки… подробнее
Фотосинтез: набор цепных реакций, преобразующих световую энергию в химическую. Фотосинтез также производит богатые энергией углеводы, такие как крахмал. Фотосинтез происходит в хлоропластах растительной клетки… подробнее
Фотосинтез также производит богатые энергией углеводы, такие как крахмал. Фотосинтез происходит в хлоропластах растительной клетки… подробнее
О чем история?
Утро пасмурное, и вы уже на полпути к финишу, ваши ноги уже болят от бега по тротуару, а ваша энергия быстро угасает. Внезапно облака рассеиваются, и солнце прорывается сквозь них и освещает вашу кожу. Вы чувствуете прилив новой энергии, которая заставляет вас закончить гонку.
Здесь мы видим круглые зеленые хлоропласты внутри растительных клеток. Внутри хлоропластов хлорофилл поглощает свет для использования в фотосинтезе для получения энергии. Хлорофилл также делает хлоропласты зелеными.
Солнечный свет может улучшить ваше самочувствие, особенно в холодный день, но он не может дать вам энергии… верно? Животные едят пищу для получения энергии, а растения используют солнечный свет для производства энергии, но не лучше ли было бы, если бы организмы могли использовать оба источника энергии? Если у вас закончилась еда, вы могли просто посидеть на солнце, чтобы накопить энергию, приберегая еду на черный день.
Синтетическая биология — это разработка и создание искусственных биологических продуктов (таких как клетки или белки). Синтетические биологи хотели выяснить, как они могут создать животную клетку, которая также имеет хлоропласты (редкое явление в природе). В статье PLOS ONE «На пути к синтетическому хлоропласту» ученые попытались создать в лаборатории организм, который использовал бы пищу и солнечный свет для производства энергии.
Требуется сборка
Проблема сложная; как бы вы поступили превращает в животную клетку, которая получает энергию как из пищи, так и из солнечного света? У нас может не быть инструкций к этой головоломке, но есть подсказки.
Когда вы истощены, вы можете есть и пить для энергии. Но растения, похоже, ничего не едят. Вместо этого они используют солнечный свет, воздух и воду для приготовления пищи. Мы получаем энергию иначе, чем растения, потому что растения и животные не используют одни и те же органеллы для этого процесса.
Ученые использовали рыбок данио (показанных здесь), хомяков и мышей, чтобы выяснить, смогут ли они заставить бактерии с хлоропластами жить внутри клеток животных.
Клетки животных используют митохондрии для преобразования пищи в энергию, а клетки растений используют как хлоропласты, так и митохондрии для получения энергии из света, воздуха и воды. Хотя мы видим некоторые примеры животных, у которых есть хлоропласты и митохондрии в некоторых клетках, например, у некоторых морских слизней, ученые хотели посмотреть, смогут ли они сделать животное, способное к фотосинтезу.
Как ученые нашли способ объединить обе органеллы в одной клетке? Узнав немного о том, как появились хлоропласты и митохондрии.
Откуда они взялись?
Вы когда-нибудь ели тарелку мороженого, когда ваша мама говорила вам: «Ты то, что ты ешь»? Ну, вы определенно не сделаны из мороженого, но «вы то, что вы едите», возможно, было правдой для некоторых клеток давным-давно.
Идея состоит в том, что в то время вокруг бродили большие клетки, питавшиеся меньшими клетками. Некоторые из этих маленьких клеток не могут быть должным образом разрушены и переварены большими клетками. Затем маленькие клетки могли осесть и жить внутри этих более крупных клеток. Эта идея называется эндосимбиозом, где эндо — значит внутри или внутри, а — симбиоз означает совместное проживание. Таким образом, эндосимбиоз означает, что две клетки живут вместе, одна внутри другой.
Некоторые из этих маленьких клеток не могут быть должным образом разрушены и переварены большими клетками. Затем маленькие клетки могли осесть и жить внутри этих более крупных клеток. Эта идея называется эндосимбиозом, где эндо — значит внутри или внутри, а — симбиоз означает совместное проживание. Таким образом, эндосимбиоз означает, что две клетки живут вместе, одна внутри другой.
Это модель хлоропласта. Хлоропласты когда-то были самостоятельными клетками. Они эволюционировали, чтобы жить внутри растительных клеток после того, как миллионы лет назад их поглотили более крупные клетки.
В этой ситуации обе клетки выиграли от присутствия другой. Большие клетки служили домами для маленьких клеток, а маленькие клетки снабжали большие клетки пищей или энергией. Считается, что эти маленькие клетки были предками того, что мы сейчас знаем как хлоропласты и митохондрии. Мы верим в это, потому что хлоропласты и митохондрии содержат свою собственную ДНК, уникальную от ДНК крупных клеток.
Эндосимбиоз считается редким явлением в природе, потому что большинство клеток имеют защиту от вторжения клеток. Но использование этой идеи помогло ученым найти инновационные способы заставить клетку с митохондриями принимать цианобактерии, особый тип бактерий, которые, вероятно, похожи на предков хлоропластов.
Вторжение зеленых инопланетян!
Тело подобно хорошо защищенному замку, куда иностранные захватчики не допускаются и погибают, если проникают внутрь. Итак, как вы можете поместить живые бактерии в организм и помочь ему выжить? Ученые придумали три разных способа проникновения цианобактерий.
Сначала ученые ввели цианобактерии непосредственно в середину эмбриона рыбки данио (или оплодотворенную яйцеклетку на одноклеточной стадии развития). Это позволило бактериям избежать внешней защиты эмбриональной клетки. Для двух других методов ученым нужно было модернизировать бактериальные клетки, чтобы иметь возможность избежать защиты животных клеток. Клетки, которые они использовали, были клетками яичника хомяка (женский репродуктивный орган) и иммунными клетками мыши, особенно макрофагами. Ученые дали цианобактериям специальные гены, которые помогли им избежать обнаружения этими клетками млекопитающих.
Ученые дали цианобактериям специальные гены, которые помогли им избежать обнаружения этими клетками млекопитающих.
Это рисунки трех методов, которые ученые использовали для внедрения цианобактерий в клетки животных. Щелкните для полной истории.
Затем модифицированным бактериям позволили либо самим проникнуть в клетки яичников, либо их проглотили мышиные макрофаги, поедающие бактерии. Ученые надеялись, что цианобактерии выживут и будут расти внутри клеток.
Растительные животные?
Хотя вы можете представить рыб, хомяков и мышей с зеленой кожей, ученые не смогли зайти так далеко. Цианобактерии смогли выжить и размножаться в клетках животных, но только в течение от пяти до 12 дней в некоторых клетках, в зависимости от метода.
Цианобактерии могли расти и размножаться в эмбрионах рыбок данио в течение двенадцати дней. После этого ученые прекратили эксперимент, потому что молекулы, окрашивающие кожу рыбы, не позволяли цианобактериям собирать солнечный свет.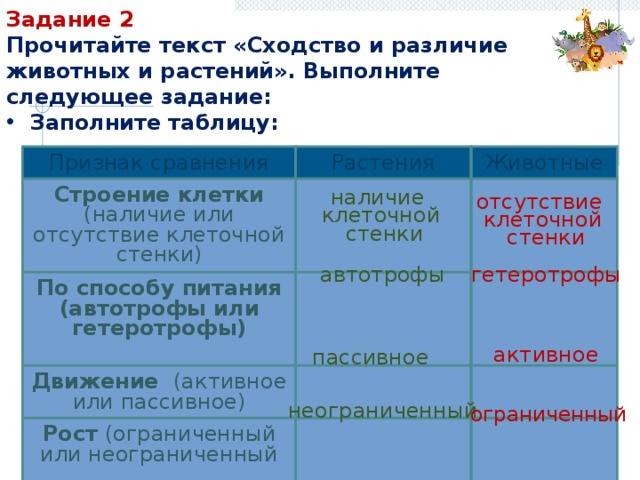
Попытки сохранить жизнь цианобактерий в клетках млекопитающих оказались менее успешными. В клетках хомяков модифицированные бактерии были помещены в клетку и выживали в течение, по крайней мере, короткого периода примерно в 5% клеток. Цианобактерии, которые были проглочены особыми поедающими бактерии клетками мышей, были способны реплицироваться (размножаться) в клетке. Однако они все равно начали умирать через 4 или 5 дней.
Даже у таких животных, как морские слизни, которые могут сохранять хлоропласты в своих собственных клетках, эти части клеток необходимо время от времени «пополнять». Они делают это, поедая водоросли или цианобактерии. Затем их пищеварительные клетки удерживают фотосинтетические части, а не разрушают их. Таким образом, краткосрочное выживание цианобактерий в клетках в этом эксперименте не так уж далеко от того, что происходит у фотосинтезирующих животных в дикой природе.
Этого краткосрочного прогресса достаточно, чтобы дать ученым надежду.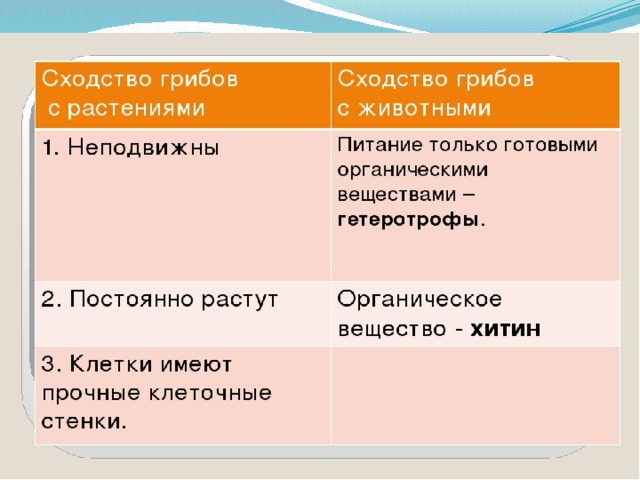 Может быть, проведя гораздо больше исследований, мы сможем однажды осуществить мечту о питании с помощью хлоропластов в нашей коже.
Может быть, проведя гораздо больше исследований, мы сможем однажды осуществить мечту о питании с помощью хлоропластов в нашей коже.
Дополнительные изображения с Викисклада. Danio rerio (рыбка данио) от Монте Вестерфилд через Национальный институт общей медицины.
Сравнение пластичности растений и животных
1. Борхес Р.М. Различаются ли растения и животные по фенотипической пластичности? Дж Биоски. 2005; 30:41–50. [PubMed] [Google Scholar]
2. Grime JP. Доказательства существования трех основных стратегий у растений и их актуальность для экологической и эволюционной теории. амер нат. 1977;111:1169–1194. [Google Scholar]
3. Grime JP, Mackey JML. Роль пластичности в захвате ресурсов растениями. Эвол Экол. 2002; 16: 299–307. [Google Scholar]
4. Брэдшоу А.Д. Эволюционное значение фенотипической пластичности растений. Ад Генет. 1965; 13: 115–155. [Google Scholar]
5. Кэмпбелл Б.Д., Грайм Дж.П., Макки Дж.М.Л. Компромисс между масштабом и точностью в поиске ресурсов.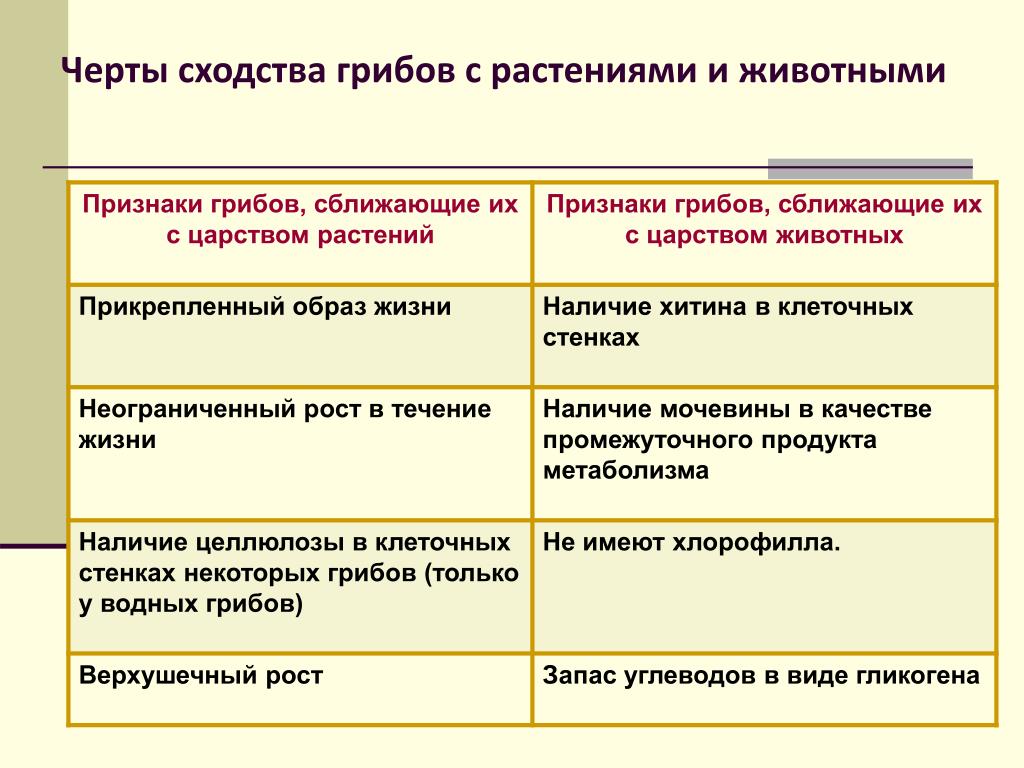 Экология. 1991; 87: 532–538. [PubMed] [Google Scholar]
Экология. 1991; 87: 532–538. [PubMed] [Google Scholar]
6. Grime JP. Компромисс между масштабом и точностью при поиске корма растениями в пространстве: восстановление перспективы. Энн Бот. 2007;99:1017–1021. [PMC free article] [PubMed] [Google Scholar]
7. Schwinning S, Weiner J. Механизмы, определяющие степень размерной асимметрии в конкуренции между растениями. Экология. 1998; 113:447–455. [PubMed] [Google Scholar]
8. Grams TEE, Andersen CP. Конкуренция за ресурсы у деревьев: физиологическая и морфологическая пластичность. Прогр Бот. 2007; 68: 356–381. [Google Scholar]
9. Валладарес Ф., Джаноли Э., Гомес Дж. М. Экологические пределы фенотипической пластичности растений. Новый Фитол. 2007;176:749–763. [PubMed] [Google Scholar]
10. Bonser SP, Aarssen LW. Распределение меристем: новая теория классификации адаптивных стратегий у травянистых растений. Ойкос. 1996; 77: 347–352. [Google Scholar]
11. Bonser SP, Aarssen LW. Аллометрия и пластичность распределения меристем на протяжении развития у Arabidopsis thaliana . J Экол. 2001; 89: 72–79. [Google Scholar]
J Экол. 2001; 89: 72–79. [Google Scholar]
12. Chenu K, Franck N, Dauzat J, Barczi J-F, Rey H, Lecoeur J. Комплексная реакция органогенеза, морфогенеза и архитектуры розеток на уменьшение падающего света в Arabidopsis thaliana приводит к более высокой эффективности перехвата света. Func Plant Biol. 2005; 32:1123–1134. [Google Scholar]
13. Puijalon S, Lena J-P, Bornette G. Интерактивное влияние питательных веществ и механических стрессов на морфологию растений. Энн Бот. 2007; 100:1297–1305. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Vasek FC. Креозотовый куст: долгоживущие клоны в пустыне Мохаве. Амер Джей Бот. 1980; 67: 246–255. [Google Scholar]
15. Джерлинг Л. Похожи ли растения и животные? Заметка об эволюционной экологии растений. Ойкос. 1985;45:150–153. [Google Scholar]
16. Craine JM. Согласование теорий стратегии растений Грайма и Тилмана. J Экол. 2005;93:1041–1052. [Google Scholar]
17. Sage RF, McKown AD. Является ли фотосинтез C 4 менее фенотипически пластичным, чем фотосинтез C 3 ? J Опытный бот. 2006; 57: 303–317. [PubMed] [Google Scholar]
2006; 57: 303–317. [PubMed] [Google Scholar]
18. Корпелайнен Х. Лабильные проявления пола у растений. Биол Rev. 1998; 73: 157–180. [Google Scholar]
19. Munday PL, Buston PM, Warner RR. Разнообразие и гибкость стратегий смены пола у животных. Тенденции Экол Эвол. 2006;21:89–95. [PubMed] [Google Scholar]
20. Vizoso DB, Schärer L. Распределение пола в зависимости от ресурсов у одновременного гермафродита. Eur Soc Evol Biol. 2007; 20:1046–1055. [PubMed] [Google Scholar]
21. Harder LD, Johnson SD. Адаптивная пластичность размеров цветков у растений, опыляемых животными. Proc R Soc Lond B. 2005;272:2651–2657. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Fordyce JA. Эволюционные последствия экологических взаимодействий, опосредованных фенотипической пластичностью. J Эксперт Биол. 2006;209: 2377–2383. [PubMed] [Google Scholar]
23. Blackstone NW, Bridge DM. Модельные системы для сигнализации об окружающей среде. Интегр Комп Биол. 2005; 45: 605–614. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
24. Касселл Э., Лейблер С. Фенотипическое разнообразие, рост населения и информация в изменчивой среде. Наука. 2005;309:2075–2078. [PubMed] [Google Scholar]
25. Кашиваги А., Урабе И., Канеко К., Йомо Т. Адаптивный ответ генной сети на изменения окружающей среды путем отбора аттракторов, вызванного приспособленностью. PloS Один. 2006;1:49. doi: 10.1371/journal.pone.0000049. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Winn AA. Вклад запрограммированных изменений в развитии и фенотипической пластичности во внутрииндивидуальную изменчивость признаков листьев у Dicerandra linearifolia . Дж. Эвол Биол. 1996; 9: 737–752. [Google Scholar]
27. Ghalambor CK, McKay JK, Carroll SP, Reznick DN. Адаптивная и неадаптивная фенотипическая пластичность и потенциал современной адаптации в новых условиях. Функц. экол. 2007;21:394–407. [Google Scholar]
28. Garland T, Jr, Kelly SA. Фенотипическая пластичность и экспериментальная эволюция. J Эксперт Биол. 2006; 209: 2344–2361. [PubMed] [Google Scholar]
J Эксперт Биол. 2006; 209: 2344–2361. [PubMed] [Google Scholar]
29. Weinig C, Delph LF. Фенотипическая пластичность в раннем возрасте ограничивает реакции развития в более позднем возрасте. Эволюция. 2001; 55: 930–936. [PubMed] [Google Scholar]
30. Hoverman JT, Relyea RA. Насколько гибка фенотипическая пластичность? Окна развития для индукции и реверсии признаков. Экология. 2007; 88: 693–705. [PubMed] [Академия Google]
31. Саркар С., Фуллер Т. Обобщенные нормы реакции для экологической биологии развития. Эвол Дев. 2003; 5: 106–115. [PubMed] [Google Scholar]
32. Blackstone NW. Окислительно-восстановительный контроль и эволюция многоклеточности. Биоэссе. 2000; 22:947–953. [PubMed] [Google Scholar]
33. Buss LW, Blackstone NW. Экспериментальное исследование эпигенетического ландшафта Уоддингтона. Фил Транс Р. Сок Лонд Б. 1991; 332: 49–58. [Google Scholar]
34. Blackstone NW. Окислительно-восстановительный контроль в развитии и эволюции: данные колониальных гидроидов. J Эксперт Биол. 1999;202:3541–3553. [PubMed] [Google Scholar]
J Эксперт Биол. 1999;202:3541–3553. [PubMed] [Google Scholar]
35. Blackstone NW. Окислительно-восстановительная передача сигналов в росте и развитии колониальных гидроидов. J Эксперт Биол. 2003; 206: 651–658. [PubMed] [Google Scholar]
36. Blackstone NW, Kelly MM, Haridas V, Gutterman JU. Митохондрии как интеграторы информации у раннего эволюционирующего животного: выводы из тритерпеноидного метаболита. Proc R Soc Lond B. 2005;272:527–531. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Blackstone NW. Многоклеточная окислительно-восстановительная регуляция: интеграция биологии организма и окислительно-восстановительной химии. Биоэссе. 2005; 28:72–77. [PubMed] [Академия Google]
38. Блэкстоун СЗ. Взгляд с точки зрения еды на переход от базальных многоклеточных животных к билатеральным. Интегр Комп Биол. 2007; 47: 724–733. [PubMed] [Google Scholar]
39. Bloomfield G, Pears C. Передача сигналов супероксида необходима для многоклеточного развития Dictyostelium .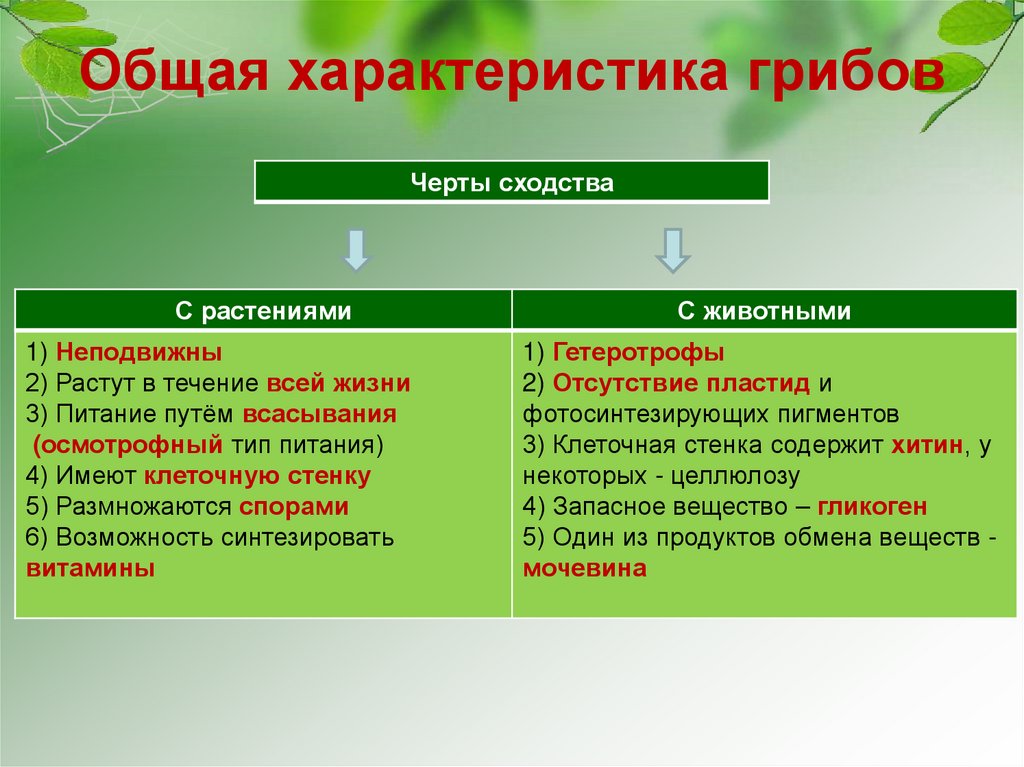 Дж. Клеточные науки. 2003; 116:3387–3397. [PubMed] [Google Scholar]
Дж. Клеточные науки. 2003; 116:3387–3397. [PubMed] [Google Scholar]
40. Nedelcu AM, Marcu O, Michod RE. Секс как ответ на окислительный стресс: двукратное увеличение клеточных активных форм кислорода активирует половые гены. Proc R Soc Lond B. 2004; 271:1591–1596. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Агирре Дж., Риос-Момберг М., Хьюитт Д., Хансберг В. Активные формы кислорода и развитие микробных эукариот. Тенденции микробиол. 2005; 13:111–118. [PubMed] [Google Scholar]
42. Terada LS. Специфика передачи сигналов реактивного оксиданта: думай глобально, действуй локально. Джей Селл Биол. 2006; 174: 615–623. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. D’Autréaux B, Toledano MB. АФК как сигнальные молекулы: механизмы, которые генерируют специфичность в гомеостазе АФК. Nature Rev Mol Cell Biol. 2007; 8: 813–824. [PubMed] [Академия Google]
44. Такемото Д., Танака А., Скотт Б. НАДФН-оксидазы в грибах: различные роли активных форм кислорода в дифференцировке клеток грибов. Генетика грибов Биол. 2007;44:1065–1076. [PubMed] [Google Scholar]
Генетика грибов Биол. 2007;44:1065–1076. [PubMed] [Google Scholar]
45. Lalucque H, Silar P. НАДФН-оксидаза: фермент многоклеточности? Тенденции микробиол. 2003; 11: 9–12. [PubMed] [Google Scholar]
46. Бедард К., Ларди Б., Краузе К.-Х. НАДФН-оксидазы семейства NOX: не только у млекопитающих. Биохимия. 2007; 89: 1107–1112. [PubMed] [Академия Google]
47. Форман Дж., Демидчик А., Ботвелл Дж.Ф.Ф., Милона П., Мидема Х., Торрес М.А., Линстед П., Коста С., Браунли С., Джонс Дж.Д.Г., Дэвис Дж.М., Долан Л. Активные формы кислорода, продуцируемые НАДФН-оксидазой, регулируют клетки растений. рост. Природа. 2003; 422:442–445. [PubMed] [Google Scholar]
48. Liszkay A, van der Zalm E, Schopfer P. Производство промежуточных продуктов с активным кислородом (O 2 .- , H 2 O 2 и . OH . ) корнями кукурузы и их роль в разрыхлении стенки и росте в длину. Завод Физиол. 2004; 136:3114–3123. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Гаппер С., Долан Л. Контроль развития растений с помощью активных форм кислорода. Завод Физиол. 2006; 141:341–345. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гаппер С., Долан Л. Контроль развития растений с помощью активных форм кислорода. Завод Физиол. 2006; 141:341–345. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Потоцкий М., Джонс М.А., Безвода Р., Смирнов Н., Жарский В. Активные формы кислорода, продуцируемые НАДФН-оксидазой, участвуют в росте пыльцевых трубок. Новый Фитол. 2007; 174: 742–751. [PubMed] [Google Scholar]
51. Mori IC, Schroeder JL. Активация активных форм кислорода в каналах Ca 2+ растений. Сигнальный механизм при полярном росте, гормональной трансдукции, стрессовой сигнализации и гипотетически механотрансдукции. Завод Физиол. 2004; 135: 702–708. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Маулик Н., Дас Д.К. Окислительно-восстановительная передача сигналов в сосудистом ангиогенезе. Бесплатно Рад Биол Мед. 2002;33:1047–1060. [PubMed] [Google Scholar]
53. Ушио-Фукай М., Александр Р.В. Активные формы кислорода как медиаторы передачи сигналов ангиогенеза. Роль НАД(Ф)Н-оксидазы. Мол Селл Биохим. 2004; 264:85–97. [PubMed] [Google Scholar]
Мол Селл Биохим. 2004; 264:85–97. [PubMed] [Google Scholar]
54. Такемото Д., Танака А., Скотт Б. Регулятор, подобный p67 Phox , задействован для контроля ветвления гиф в мутуалистическом симбиозе грибов и трав. Растительная клетка. 2006;18:2807–2821. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Шахтман Д.П., Шин Р. Определение питательных веществ и сигнализация: NPKS. Annu Rev Plant Biol. 2007; 58:47–69. [PubMed] [Google Scholar]
56. Walch-Liu P, Ivanov II, Filleur S, Gan Y, Remans R, Forde BG. Азотная регуляция ветвления корней. Энн Бот. 2006: 875–881. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Эшли М.К., Грант М., Грабов А. Реакция растений на дефицит калия; Роль транспортных белков калия. J Опытный бот. 2006; 57: 425–436. [PubMed] [Академия Google]
58. Делледонн М. Отсутствие новостей – хорошая новость для растений. Curr Op Plant Biol. 2005; 8: 390–396. [PubMed] [Google Scholar]
59. Лантери М.Л., Грациано М. , Корреа-Арагунде Н., Ламаттина Л. От деления клеток к форме органа: оксид азота участвует в опосредованном ауксином развитии корней. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 123–136. [Google Scholar]
, Корреа-Арагунде Н., Ламаттина Л. От деления клеток к форме органа: оксид азота участвует в опосредованном ауксином развитии корней. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 123–136. [Google Scholar]
60. Zhao D-Y, Tian Q-Y, Li L-H, Zhang WH. Оксид азота участвует в нитрат-индуцированном ингибировании удлинения корней у 9 растений.0003 Зеа майс . Энн Бот. 2007; 100: 497–503. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Де Стефано М., Феррарини А., Делледонн М. Участие оксида азота в несовместимых взаимодействиях растений и патогенов. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 111–121. [Google Scholar]
62. Коста С., Шоу П. «Непредубежденные» клетки: как клетки могут изменить судьбу. Тенденции клеточной биологии. 2006; 17: 101–106. [PubMed] [Академия Google]
63. Guyomaarch’h S, Bertrand C, Delarue M, Zhou D-X. Регуляция активности меристем путем ремоделирования хроматина. Тенденции Растениевод. 2005; 10: 332–338. [PubMed] [Google Scholar]
Регуляция активности меристем путем ремоделирования хроматина. Тенденции Растениевод. 2005; 10: 332–338. [PubMed] [Google Scholar]
64. Шерес Б. Ниши стволовых клеток: детские стишки в разных королевствах. Nature Rev Mol Cell Biol. 2007; 8: 345–354. [PubMed] [Google Scholar]
65. Такер М.Т., Лаукс Т. Соединение путей в регуляции стволовых клеток растений. Тенденции клеточной биологии. 2007; 17: 403–410. [PubMed] [Google Scholar]
66. Рандо Т.А. Стволовые клетки, старение и стремление к бессмертию. Природа. 2006; 441:1080–1086. [PubMed] [Академия Google]
67. Blackstone NW, Jasker BD. Филогенетические соображения клональности, колониальности и способа развития зародышевой линии у животных. J Exp Zool (Mol Dev Ecol) 2003; 297: B35–B47. [PubMed] [Google Scholar]
68. Whitham TG. Отдельные деревья как гетерогенные среды: адаптация к травоядным или эпигенетический шум? В: Denno RF, Dingle H, редакторы. Образцы истории жизни насекомых: среда обитания и географическая изменчивость. Нью-Йорк: Springer-Verlag; 1981. С. 9–27. [Академия Google]
Нью-Йорк: Springer-Verlag; 1981. С. 9–27. [Академия Google]
69. Гилл Д.Э., Чао Л., Перкинс С.Л., Вольф Дж.Б. Генетический мозаицизм у растений и клональных животных. Annu Rev Ecol Syst. 1995; 26: 423–444. [Google Scholar]
70. Томас Х. Старение растений. Механическое старение Dev. 2002; 123:747–753. [PubMed] [Google Scholar]
71. Мунне-Бош С. Старение многолетников. Crit Rev Plant Sci. 2007; 26: 123–138. [Google Scholar]
72. де Крон Х., Хубер Х., Штуфер Дж. Ф., ван Грюнендаль Дж. М. Модульная концепция фенотипической пластичности растений. Новый Фитол. 2005; 166:73–82. [PubMed] [Академия Google]
73. Orians C. Травоядные, сосудистые пути и системная индукция: факты и артефакты. Дж. Хим. Экол. 2005; 31: 2231–2242. [PubMed] [Google Scholar]
74. Шаиш Л., Абельсон А., Ринкевич Б. Насколько пластичной может быть фенотипическая пластичность? Ветвящийся коралл Stylophora pistillata как модельная система. PloS Один. 2007; 2:644. doi: 10. 1371/journal.pone.0000644. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
1371/journal.pone.0000644. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
75. Cookson SJ, Radziejwoski A, Granier C. Пластичность размеров клеток и листьев в Arabidopsis : какова роль эндоредупликации? Окружающая среда растительной клетки. 2006; 29: 1273–1283. [PubMed] [Google Scholar]
76. Джейкобс Т. Почему клетки растений делятся? Растительная клетка. 1997; 9: 1021–1029. [Бесплатная статья PMC] [PubMed] [Google Scholar]
77. Цукая Х. Форма и размер органов: уроки исследований морфогенеза листьев. Curr Op Plant Biol. 2003; 6: 57–62. [PubMed] [Google Scholar]
78. Peters WS, Hagemann W, Tomos AD. Чем отличаются растения? Принципы функционирования внеклеточного матрикса в «мягких» тканях растений. Comp Biochem Physiol A. 2000;125:151–167. [PubMed] [Академия Google]
79. Балушка Ф., Фолькманн Д., Барлоу П. Эукариотические клетки и их клеточные тела: пересмотренная клеточная теория. Энн Бот. 2004; 94:9–32. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Fleming AJ. Интеграция клеточной пролиферации и роста в морфогенез листа. J Завод Res. 2006; 119:31–36. [PubMed] [Google Scholar]
Fleming AJ. Интеграция клеточной пролиферации и роста в морфогенез листа. J Завод Res. 2006; 119:31–36. [PubMed] [Google Scholar]
81. Балушка Ф., Войташек П., Фолькманн Д., Барлоу П. Архитектура роста поляризованных клеток: уникальный статус удлиняющихся растительных клеток. Биоэссе. 2003;25:569–576. [PubMed] [Google Scholar]
82. Войташек П., Фолькманн Д., Балуска Ф. Полярность и клеточные стенки. В: Линдси К., редактор. Полярность растений. Оксфорд: Блэквелл; 2004. стр. 72–103. [Google Scholar]
83. Барлоу П., Фолькманн Д., Балушка Ф. Полярность корней. В: Линдси К., редактор. Полярность растений. Оксфорд: Блэквелл; 2004. стр. 192–241. [Google Scholar]
84. Фримл Дж. Транспорт ауксина — формирование растения. Curr Op Plant Biol. 2003; 6: 7–12. [PubMed] [Академия Google]
85. Де Смет И., Юргенс Г. Паттернирование оси у растений — контроль над ауксином. Curr Op Genet Dev. 2007; 17: 337–343. [PubMed] [Google Scholar]
86. Балушка Ф., Хазенштейн К. Х. Цитоскелет корня: его роль в восприятии и реакции на гравитацию. Планта. 1997; 203:S69–S78. [PubMed] [Google Scholar]
Х. Цитоскелет корня: его роль в восприятии и реакции на гравитацию. Планта. 1997; 203:S69–S78. [PubMed] [Google Scholar]
87. Blancaflor EB, Masson PH. Гравитропизм растений. Распутывание взлетов и падений сложного процесса. Завод Физиол. 2003; 133:1677–1690. [Бесплатная статья PMC] [PubMed] [Google Scholar]
88. Harrison BR, Masson PH. ARL2, ARG1 и PIN3 определяют путь передачи гравитационного сигнала в корневых статоцитах. Плант Дж. 2008; 53: 380–392. [PubMed] [Google Scholar]
89. Telewski FW. Единая гипотеза механорецепции у растений. Амер Джей Бот. 2006; 93: 1466–1476. [PubMed] [Google Scholar]
90. Ingber DE. Тенсегрити 2. Как структурные сети влияют на сотовые сети обработки информации. Дж. Клеточные науки. 2003; 116:1397–1408. [PubMed] [Академия Google]
91. Ван Н., Батлер Дж. П., Ингбер Д.Э. Механотрансдукция через клеточную поверхность и через цитоскелет. Наука. 1993; 260:1124–1127. [PubMed] [Google Scholar]
92. Балушка Ф., Главачка А. Растительные формины достигают совершеннолетия: что-то особенное в поперечных стенках. Новый Фитол. 2005; 168: 499–503. [PubMed] [Google Scholar]
Растительные формины достигают совершеннолетия: что-то особенное в поперечных стенках. Новый Фитол. 2005; 168: 499–503. [PubMed] [Google Scholar]
93. Мур SW. Яичница-болтунья: механические силы как экологические факторы раннего развития. Эвол Дев. 2003; 51: 61–66. [PubMed] [Google Scholar]
94. Jaffe MJ, Leopold AC, Staples RC. Тигмо-ответы у растений и грибов. Амер Джей Бот. 2002;89: 375–382. [PubMed] [Google Scholar]
95. Браам Дж. На связи: реакция растений на механические раздражители. Новый Фитол. 2005; 165: 373–389. [PubMed] [Google Scholar]
96. Маккормак Э., Веласкес Л., Делк Н.А., Браам Дж. Сенсорное поведение и экспрессия генов у растений. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 249–260. [Google Scholar]
97. Читайте Дж., Стоукс А. Биомеханика растений в экологическом контексте. Амер Джей Бот. 2006;93: 1546–1565. [PubMed] [Google Scholar]
98. Spector T, Putz FE. Биомеханическая пластичность способствует проникновению бразильского перца ( Schinus terebinthifolius ) в приморские леса на юге США. Biol Inv. 2006; 8: 255–260. [Google Scholar]
99. Робертс М.Р., Пол Н.Д. Соблазненный темной стороной: интеграция молекулярных и экологических взглядов на влияние света на защиту растений от вредителей и патогенов. Новый Фитол. 2006; 170: 677–699. [PubMed] [Академия Google]
100. Иноуэ С., Киношита Т., Такемия А., Дои М., Симадзаки К. Положение листьев Arabidopsis в ответ на синий свет. Мол завод. 2008; 1:15–26. [PubMed] [Google Scholar]
101. Izaguirre MM, Mazza CA, Biondini M, Baldwin IT, Ballaré CL. Дистанционное зондирование будущих конкурентов: влияние на защитные силы растений. Proc Natl Acad Sci USA. 2006; 103:7170–7174. [Бесплатная статья PMC] [PubMed] [Google Scholar]
102. Винсент Г. Пластичность продолжительности жизни листьев тропических саженцев, выращенных в условиях контрастного освещения. Энн Бот. 2006;97: 245–255. [Бесплатная статья PMC] [PubMed] [Google Scholar]
103. Kleiman D, Aarssen LW. Компромисс размера и количества листьев у деревьев. J Экол. 2007; 95: 376–382. [Google Scholar]
104. Mommer L, Visser EJW. Подводный фотосинтез у затопленных наземных растений: вопрос пластичности листьев. Энн Бот. 2005; 96: 581–589. [Бесплатная статья PMC] [PubMed] [Google Scholar]
105. Furla P, Allemand D, Shick JM, Ferrier-Pages C, Richier S, Plantivaux A, Merle PL, Tambutté S. Симбиотический антозой: физиологическая химера между водоросли и животные. Интегр Комп Биол. 2005;45:595–604. [PubMed] [Google Scholar]
106. Wells CL, Pigliucci M. Адаптивная фенотипическая пластичность: случай гетерофилии у водных растений. Perspect Plant Evol Ecol Syst. 2000; 3:1–18. [Google Scholar]
107. Кувабара А., Нагата Т. Клеточная основа пластичности развития, наблюдаемая при формировании гетерофильных листьев Ludwigia arcuata (Onagraceae) Planta. 2006; 224:761–770. [PubMed] [Google Scholar]
2006; 224:761–770. [PubMed] [Google Scholar]
108. Pierik R, Sasidharan R, Voesenek LACJ. Контроль роста с помощью этилена: приспособление фенотипов к окружающей среде. J Регулятор роста растений. 2007; 26: 188–200. [Академия Google]
109. Вятт ТД. Феромоны и поведение животных. Общение через запах и вкус. Великобритания: Издательство Кембриджского университета; 2002. [Google Scholar]
110. Baldwin IT, Halitschke R, Paschold A, von Dahl CC, Preston CA. Летучие сигналы во взаимодействиях между растениями: «Говорящие деревья» в эпоху геномики. Наука. 2006; 311:812–815. [PubMed] [Google Scholar]
111. Хайль М. Непрямая защита посредством тритрофических взаимодействий. Новый Фитол. 2007 г.: 10.1111/j.1469-8137.2007.02330.x. [PubMed] [CrossRef] [Академия Google]
112. Karban R, Maron J, Felton GW, Ervin G, Eichenseer H. Повреждение травоядными полыни вызывает устойчивость дикого табака: доказательства подслушивания между растениями. Ойкос. 2003; 100:325–332. [Google Scholar]
[Google Scholar]
113. Кесслер А., Халичке Р., Дизель С., Болдуин И. Инициация защитных реакций растений в природе посредством передачи сигналов по воздуху между Artemisia tridentata и Nicotiana attenuata . Экология. 2006; 148: 280–292. [PubMed] [Google Scholar]
114. Karban R, Shiojiri K, Huntzinger M, McCall AC. Устойчивость полыни к повреждениям: летучие вещества играют ключевую роль во внутри- и межрастительной коммуникации. Экология. 2006;87:922–930. [PubMed] [Google Scholar]
115. Heil M, Bueno JCS. Передача сигналов внутри растений с помощью летучих веществ приводит к индукции и запуску непрямой защиты растений в природе. Proc Natl Acad Sci USA. 2007; 104: 5467–5472. [Бесплатная статья PMC] [PubMed] [Google Scholar]
116. Frost CJ, Appel HM, Carlson JE, De Moraes CM, Mescher MC, Schultz JC. Передача сигналов внутри растений через летучие вещества преодолевает сосудистые ограничения на системную передачу сигналов и инициирует ответ против травоядных. Эколь Летт. 2007;10:490–498. [PubMed] [Google Scholar]
Эколь Летт. 2007;10:490–498. [PubMed] [Google Scholar]
117. Rasmann S, Köllner TG, Degenhardt J, Hiltpold I, Topfer S, Kuhlmann U, Gershenzon J, Turlings TCJ. Рекрутирование энтомопатогенных нематод поврежденными насекомыми корнями кукурузы. Природа. 2005; 434: 732–737. [PubMed] [Google Scholar]
118. Runyon JB, Mescher MC, De Moraes CM. Летучие химические сигналы определяют местонахождение хозяина и выбор хозяина растениями-паразитами. Наука. 2006; 313:1964–1967. [PubMed] [Google Scholar]
119. Paschold A, Halitschke R, Baldwin IT. Использование «немых» растений для передачи изменчивых сигналов. Плант Дж. 2006; 45: 275–29.1. [PubMed] [Google Scholar]
120. Pierik R, Visser EJW, De Kroon H, Voesenek L. Этилен необходим в табаке, чтобы успешно конкурировать с ближайшими соседями. Окружающая среда растительной клетки. 2003; 26:1229–1234. [Google Scholar]
121. von Dahl CC, Baldwin IT. Расшифровка роли этилена во взаимодействиях растений и травоядных. J Регулятор роста растений. 2007; 26: 201–209. [Google Scholar]
J Регулятор роста растений. 2007; 26: 201–209. [Google Scholar]
122. Adie B, Chico JM, Rubio-Somoza I, Solano R. Модуляция защиты растений этиленом. J Регулятор роста растений. 2007; 26: 160–177. [Академия Google]
123. Волков А.Г. Зеленые растения: электрохимические интерфейсы. J Electroanal Chem. 2000; 483:150–156. [Google Scholar]
124. Дэвис Э. Электрические сигналы в растениях: факты и гипотезы. В: Волков АГ, редактор. Электрофизиология растений — теория и методы. Берлин, Гейдельберг: Springer-Verlag; 2006. стр. 407–422. [Google Scholar]
125. Балушка Ф., Фолькманн Д., Главачка А., Манкузо С., Барлоу П.В. Нейробиологический взгляд на растения и их строение тела. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 19.–35. [Google Scholar]
126. Фромм Дж., Лотнер С. Электрические сигналы и их физиологическое значение для растений. Окружающая среда растительной клетки. 2007; 30: 249–257. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
127. Van Bel AJE, Ehlers K. Передача электрических сигналов через плазмодесмы. В: Опарка К.Я., изд. Плазмодесмы. Оксфорд: Блэквелл; 2004. стр. 263–278. [Google Scholar]
128. Maffei M, Bossi S, Spiteller D, Mithöfer A, Boland W. Влияние подкормки Spodoptera littoralis на листья лимской фасоли. I. Мембранные потенциалы, вариации внутриклеточного кальция, ротовые выделения и компоненты регургитации. Завод Физиол. 2004; 134:1752–1762. [Бесплатная статья PMC] [PubMed] [Google Scholar]
129. Maischak H, Grigorieva PA, Vogela H, Boland W, Mithöfer A. Оральные выделения травоядных личинок чешуекрылых проявляют активность по формированию ионных каналов. ФЭБС лат. 2007; 581: 898–904. [PubMed] [Google Scholar]
130. Фейт А., Йеллин А., Фромм Х. Нейротрансмиттеры ГАМК и ГОМК в растениях и животных. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 171–185. [Google Scholar]
[Google Scholar]
131. Бланкафлор Э.Б., Чепмен К.Д. Сходство между передачей сигналов эндоканнабиноидов в системах животных и N — метаболизм ацилэтаноламина в растениях. В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 206–219. [Google Scholar]
132. Лох Т.Дж., Лукас В.Дж. Интегративная биология растений: роль флоэмы в перемещении макромолекул на большие расстояния. Annu Rev Plant Biol. 2006; 57: 203–232. [PubMed] [Google Scholar]
133. Mullineaux P, Ball L, Escobar C, Karpinska B, Creissen G, Karpinski S. Различные сигнальные пути интегрированы в регуляцию Экспрессия генов антиоксидантной защиты Arabidopsis в ответ на избыточную энергию возбуждения? Фил Транс Р. Сок Лонд Б. 2000; 355: 1531–1540. [Бесплатная статья PMC] [PubMed] [Google Scholar]
134. Филковски Дж., Йоман А., Ковальчук О., Ковальчук И. Системный сигнал растений вызывает нестабильность генома. Плант Дж. 2004; 38: 1–11. [PubMed] [Google Scholar]
Плант Дж. 2004; 38: 1–11. [PubMed] [Google Scholar]
135. Schwachtje J, Minchin PEH, Janke S, van Dongen JT, Schittko U, Baldwin IT. Киназы, связанные с SNF1, позволяют растениям переносить травоядность за счет выделения углерода корням. Proc Natl Acad Sci USA. 2006;103:12935–12940. [Бесплатная статья PMC] [PubMed] [Google Scholar]
136. Сакс Т. Как растения могут выбирать наиболее перспективные органы? В: Балушка Ф., Манкузо С., Фолькманн Д., редакторы. Коммуникация у растений. Берлин, Гейдельберг: Springer-Verlag; 2006. С. 53–63. [Google Scholar]
137. Нефф М.М., Стрит И.Х., Терк Э.М., Уорд Дж.М. Взаимодействие световой и гормональной сигнализации для опосредования фотоморфогенеза. В: Schäfer E, Nagy F, редакторы. Фотоморфогенез у растений и бактерий. Нидерланды: Спрингер; 2006. стр. 439.–473. [Google Scholar]
138. Дюме Дж. Может ли механика управлять формированием паттернов в растениях? Curr Op Plant Biol. 2007; 10:58–62. [PubMed] [Google Scholar]
139. Bjedov I, Tenaillon O, Gerard B, Souza V, Denamur E, Radman M, Taddei F, Matic I. Мутагенез, вызванный стрессом у бактерий. Наука. 2003; 300:1404–1409. [PubMed] [Google Scholar]
Bjedov I, Tenaillon O, Gerard B, Souza V, Denamur E, Radman M, Taddei F, Matic I. Мутагенез, вызванный стрессом у бактерий. Наука. 2003; 300:1404–1409. [PubMed] [Google Scholar]
140. Rando OJ, Verstrepen KJ. Временные шкалы генетического и эпигенетического наследования. Клетка. 2007; 128: 655–668. [PubMed] [Академия Google]
141. Донг X. Патоген-индуцированная системная перестройка ДНК в растениях. Тенденции Растениевод. 2004; 9: 60–61. [PubMed] [Google Scholar]
142. Бойко А., Катирия П., Земп Ф.Дж., Яо Ю., Погрибный И., Ковальчук И. Трансгенерационные изменения стабильности генома и метилирования у растений, инфицированных патогенами (Вирус-индуцированная нестабильность генома растений) Нук Кислота Рез. 2007; 35: 1714–1725. [Бесплатная статья PMC] [PubMed] [Google Scholar]
143. Molinier J, Ries G, Zipfel C, Hohn B. Трансгенерационная память о стрессе у растений. Природа. 2006; 442:1046–1049. [PubMed] [Google Scholar]
144. Schultz JC. Общие сигналы и возможность филогенетического шпионажа между растениями и животными.