Содержание
Анатомическое строение стебля растения
ФГБОУ ВПО «Чувашский государственный
педагогический университет»
Кафедра биологии и методики преподавания
РЕФЕРАТ
по ботанике
на тему: «Анатомическое строение стебля
растения»
Выполнила: студентка 1 курса
ФЕиДС группы ЕХБ-1
Зарипова А.Ф.
Проверил: доцент
Дмитриева О. Ф.
Ф.
Содержание
Глава
1. Анатомическое строение стебля однодольных
растений…………3-6
Глава
2. Анатомическое строение стебля двудольных
растений…………..7-9
Глава
3. Анатомическое строение стебля древесных
растений……………..10-12
Использованная
литература………………..
Глава 1
Анатомическое строение
стебля однодольных
растений
Однодольные растения (Monocotyleclones)-
класс покрытосемянных цветковых растений,
который характеризуется тем, что зародыш
снабжен только одной семядолею. Сюда
Сюда
принадлежат Частуховые, Осоковые, Злаки,
Пальмы, Ароидные, Лилейные, Имбирные,
Орхидные и др.
Стебли однодольных,
особенно травянистых злаков, имеют первичное
строение. Его мы рассмотрим на примере
ржи и кукурузы.
На поперечном разрезе
стебля кукурузы можно видеть, что
с поверхности стебель покрыт однослойным
эпидермисом, стенки клеток эпидермиса
утолщены. Снаружи эпидермис покрыт слоем
кутикулы. Под эпидермисом располагается
сплошное кольцо мелкоклеточной склерифицированной
паренхимы, постепенно переходящей в обычную
крупноклеточную паренхиму. Ширина
склерифицированного кольца неодинакова;
в молодых стеблях хлорофиллоносная паренхима
подходит непосредственно к эпидермису,
и в этих местах можно заметить устьица.
В молодом стебле
стенки клеток субэпидермального цилиндра
тонки и слабо одревеснены; процесс одревеснения
клеточных стенок только начинается. С
возрастом происходят такие изменения:
увеличивается толщина склерифицированного
цилиндра, стенки клеток всё более древеснеют
и становятся толще, различия между этими
клетками и клетками обычной паренхимы
становятся всё более резкими.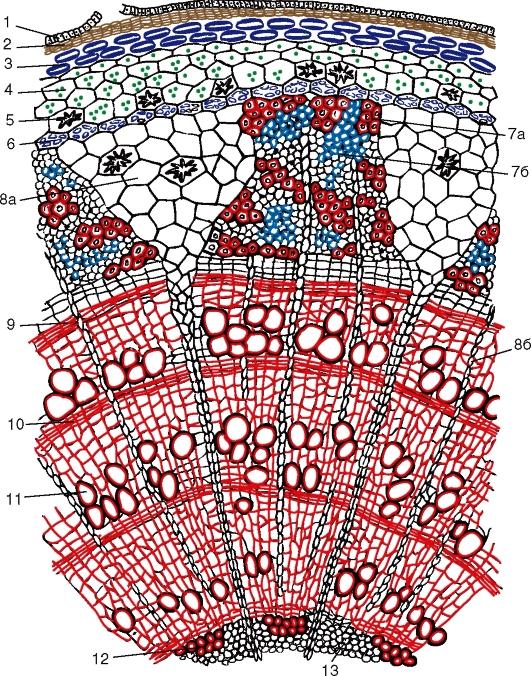
Основную массу
стебля составляет паренхима. Клетки паренхимы
крупные, с тонкими стенками; между ними
— межклетники. В молодых стеблях паренхима
сочная, клетки сильно вакуолизированы,
вакуоли заполнены сладким клеточным
соком. К концу вегетации большая часть
паренхимных клеток заполняется воздухом,
благодаря которому вся сердцевина становится
белой, очень мягкой, почти как вата.
В паренхимной ткани
без заметной закономерности разбросано
большое количество сосудисто — волокнистых
пучков. Те из них, которые находятся ближе
к периферии, размещены более густо, — они
непосредственно примыкают к склерифицированному
цилиндру; ближе к центру стебля пучки
размещены реже, но каждый пучок становится
крупнее. Пучков очень много: даже в молодых
стеблях их можно насчитать свыше сотни.
Проводящие пучки
злаков коллатерального типа, они не содержат
камбия, следовательно, это закрытые пучки.
Такие пучки формируются
в результате дифференциации прокамбиальных
тяжей, закладываемых в меристеме в процессе
формирования стеблей и листьев.
Первоначальные
проводящие элементы ксилемы представлены
несколькими узкими кольчатыми и спиральными
сосудами протоксилемы, окружёнными мелкоклетной
паренхимой. К моменту окончательного
формирования пучка эта паренхима разрывается,
и под сосудами образуется воздухоносная
полость. Позднее образуется два широких
пористых сосуда метаксилемы, расположенных
симметрично по бокам, ближе к флоэме,
которая занимает часть пучка, ориентированную
к периферии поперечного разреза стебля.
Каждый сосудисто
— волокнистый пучок у злаков окружён склеренхимной
обкладкой, или чехлом, в виде сплошной
трубочки, внутри которой находятся проводящие
и паренхимные элементы пучка. Мощность
склеренхимной обкладки пучков возрастает
по направлению к периферии стебля, так
что механически оснащённые пучки занимают
периферийное положение.
Стебель ржи, и вообще
большинства злаков, полый; такой стебель
называется соломиной. Полость образуется
на ранних этапах формирования стебля
в результате отмирания сердцевидной
паренхимы. В оставшемся периферическом
В оставшемся периферическом
слое паренхимы размещены закрытые коллатеральные
пучки, однако их значительно меньше, чем
в стебле кукурузы.
Так же, как и у кукурузы,
пучки одеты склеренхимным чехлом. Наиболее
периферийные из них размещаются в общем
склерифицированном цилиндре, примыкающем
к эпидермису. Стенки клеток эпидермиса
и склеренхимы с возрастом древеснеют.
Непосредственно
под эпидермисом в толще склерифицированного
цилиндра располагаются парные тяжи хлорофиллоносной
паренхимы, на поперечном разрезе они
имеют вид двух продолговатых участков,
прилегающих к эпидермису длинной стороной.
В эпидермисе стебля над хлорофиллоносными
участками имеются устьица. Поверхностные
тяжи хлорофиллоносной паренхимы хорошо
заметны при рассмотрении ещё не пожелтевшей
соломины; поверхность стебля полосатая,
тёмно- зелёные полосы чередуются с более
светлыми.
В узлах стебель
несколько утолщён. У растений с полым
стеблем в этом месте перегородка.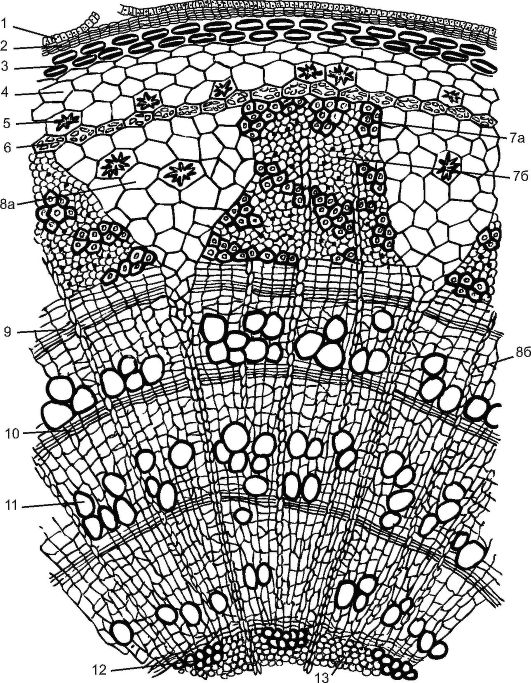 Утолщения
Утолщения
в узлах связаны с тем, что здесь находятся
обильные сплетения мелких стеблевых
пучков с пучками, выходящими из листьев;
между этими пучками имеются многочисленные
анастомозы. Стебель на узлах оснащён
добавочным количеством склеренхимной
ткани.
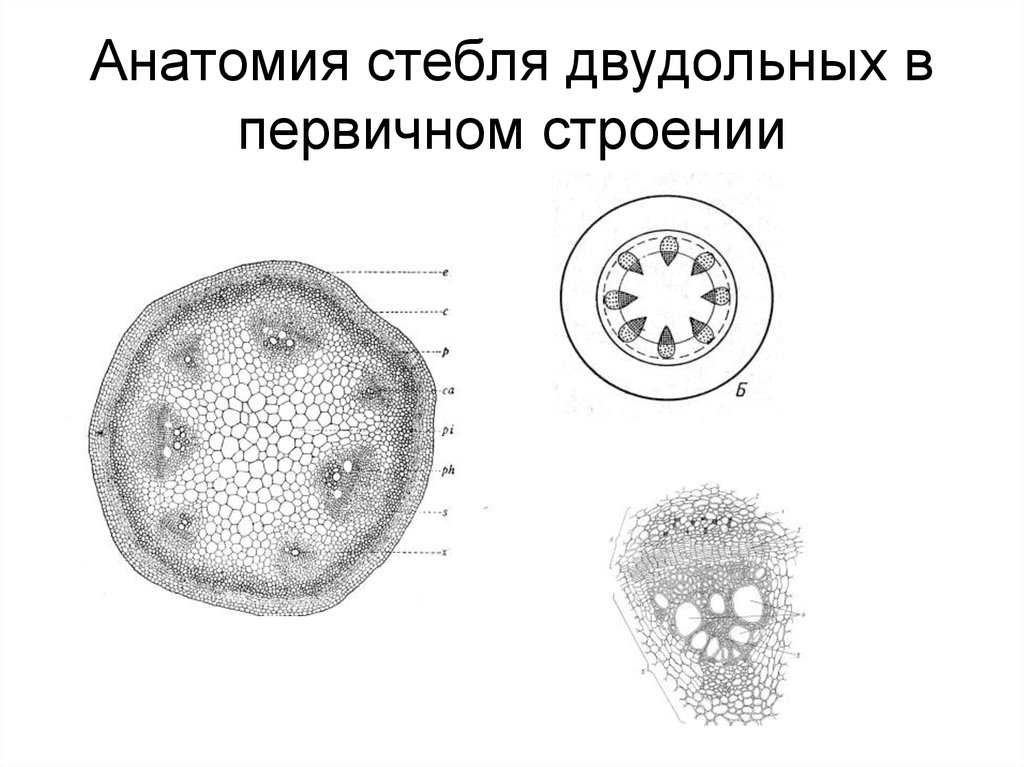
Глава 2
Анатомическое
строение стебля двудольных
растений
Двудо́льные
(устар. варианты: двусемядольные, двусемянодольные)
(лат. Dicotylédones), или Магнолиопси́ды
(лат. Magnoliópsida) — класс покрытосеменных
растений, у которых зародыш семени имеет
две боковые супротивные семядоли.
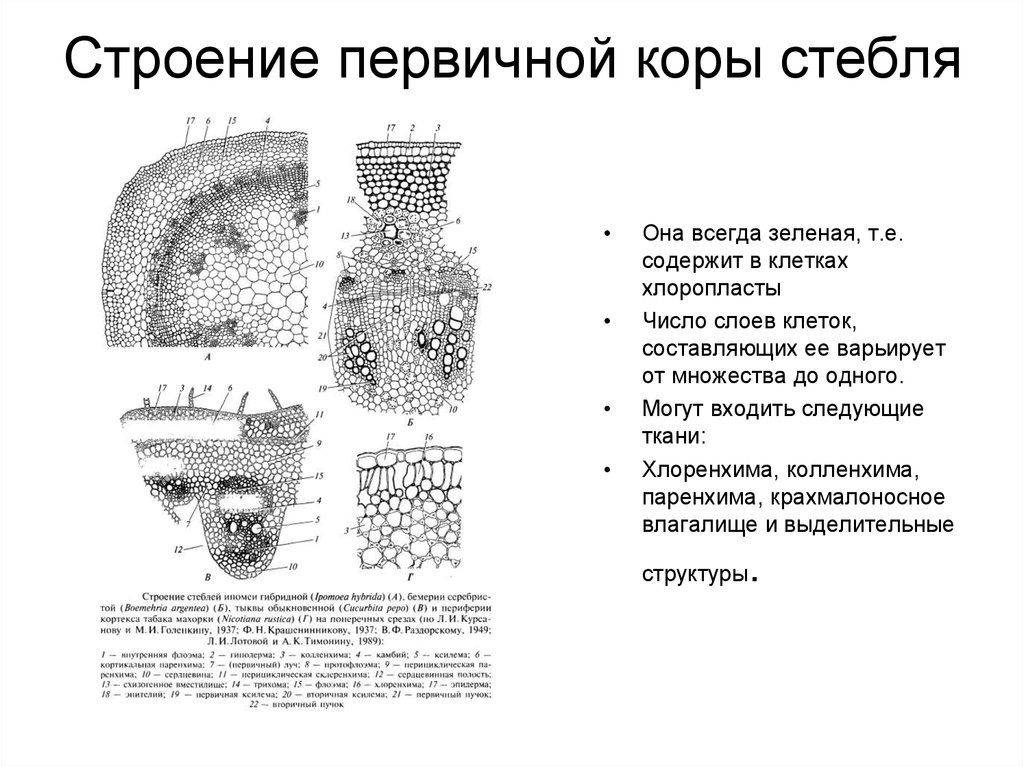
В анатомическом
строении стебля двудольных растений
принято различать первичную и вторичную
структуру. Первичная структура возникает
в результате дифференциации аппикальной
меристемы, а переход ко вторичной структуре
начинается с момента деятельности камбия.
С поверхности
стебель покрыт эпидермисом. На небольших
выростах эпидермиса, состоящих из группы
мелких клеток, сидят жёсткие волоски,
состоящие из ряда клеток. Всю остальную
массу стебля на поперечном разрезе разделяют
на 2 слоя: первичную кору и центральный
осевой цилиндр.
Первичная кора
состоит из разнообразных элементов. Непосредственно
под эпидермисом несколько слоёв пластинчатой
колленхимы, клетки которой содержат хлорофилловые
зёрна; затем идёт тонкостенная паренхима,
в её клетках количество хлоропластов
постепенно уменьшается по направлению
к центру;переход между колленхимой и
парнехимой не резкий.
Последний, внутренний
слой клеток первичной коры называется
эндодермой, или крахмалоносным влагалищем;
в клетках этого слоя имеются крахмальные
зёрна, роль которых недостаточно выяснена.
Крахмальные зёрна эндодермы не расходуются
даже при голодании растений. В коровой
паренхиме разбросаны смоляные ходы- узенькие
канальцы, окружённые мелкими клетками
основной ткани. У других видов травянистых
У других видов травянистых
растений могут быть различные отклонения:
колленхима или отсутствует вовсе, или,
если стебь имеет ребристую систему, распологается
тяжами, в таком случае колленхима обычно
уголковая или переходного характера.
Выдающиеся ребрышки, в основном, и состоят
из такой колленхимы. Вместо колленхимы
может быть сплошной цилиндр или отдельные
прослойки склеренхимы; последняя размещается
всегда глубже колленхимы, между склеренхимной
и эпидермисом остаётся слой хлорофиллоносной
паренхимы.
Первый слой паренхимных
клеток центрального осевого цилиндра,
ли стелы, непосредственно примыкающий
к эндодерме, относится к перициклу, клетки
которого способны к возобновлению меристематической
деятельности. Клетки перицикла могут
превращаться в лубяные волокна, которые
называются первичными, в отличии от вторичных,
образованных камбием. Перицикл даёт начало
придаточным корням. Глубже перицикла
располагается основная паренхима, в которой
размещены коллатеральные пучки, расположённые
здесь закономерн, по кругу. Пучки возникают
Пучки возникают
из прокамбиальных тяжей, закладываемых
ещё апикальной меристемой.
В отличии от
однодольных, у которых весь прокамбий
расходуется на образование постоянных
элементов пучка, у двудольных прокамбий
полностью не дифференцируется; клетки
узенькой прослойки прокамбия между
сформировавшимися первичными ксилемой
и флоэмой сохраняют способность делиться
и превращаются в постоянную действующую
прослойку первичного камбия.
Камбий продолжает
формировать элементы ксилемы и флоэмы.
В отличие от элементов, образованных
прокамбием, ксилема и флоэма, возникающие
в результате действия камбия, называется
вторичным. С момента деятельности камбия
начинается формирование вторрчной структуры
стебля.
Между пучками,
образованными прокамбием, остаются прослойки
сердцевиннной паренхимы, играющие роль
в горизонтальном перемещении растворов.
Эти прослойки называют сердцевинными,
радиальными лучами или межпучковыми
зонами. Центральный участок стебля, состоящий
из паренхимы, ограниченный внутренними
концами пучков, называется сердцевиной.
Сердцевинные пучки соединяют сердцевину
с перициклом.
У некоторых
растений (лён, сурепка,мыльнянка и др.)отдельные
тяжи прокамбия ещё в конусе нарастания
соединятся и дают сплошное кольцо камбия,
который продуцирует кнаружи элементы
вторичной флоэмы, внутрь-вторичной ксилемы.
Такое строение стебля называется беспучковым.
Вторичные изменения
в стебле пучкового строения не ограничиваются
деятельностью пучкового камбия. Уже в
самом начале вторичных изменений в том
участке первичного сердцевинного
луча, который расположен между камбиальными
прослойками двух соседних пучков,
клетки основной ткани преобретают свойство
энергично делиться. В результате этого
деления появляется участок образовательной
ткани, соединяющейся в конце концов с
соседними участками пучкового камбия.
Вновь образованные
прослойки образовательной ткани называется
межпучковым камбием. Таким образом, межпучковый
камбий по своему происхождению является
вторичным камбием. Вторичный камбий соединяется
с первичным пучковым камбием, образуя
общее камбиальное кольцо, клетки которого
продолжают функционировать согласлванно,
подстилая вторичную флоэму под первичную
и наслаивая вторичную ксилему на первичную.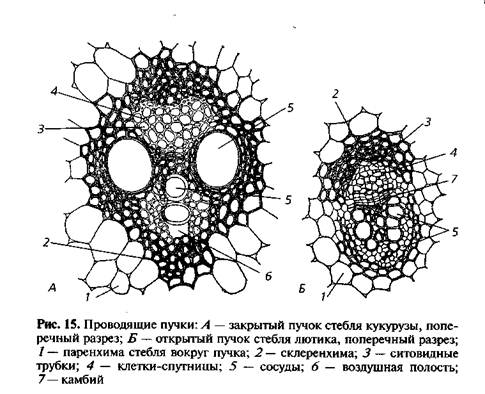
Межпучковый
камбий обычно вначале продуцирует паренхиму,
наращивая таким образом первичные сердцевинные
лучи; у некоторых растений он формирует
механическую ткань- склеренхиму. При
существенном утолщении стебля , как, например,
у подсолнечника, межпучковый камбий формирует
добавочные пучки вторичного происхождения;
у этих пучков нет первичных элементов,
а ксилема и флоэма у них только вторичные.
В результате разрастания старых и вновь
возникающих пучков сердцевинные лучи
настолько стискиваются, что ксилемы соседних
пучков почти сливаются, образуя сплошной
древесинный цилиндр.
В стеблях беспучкового
строения сплошное кольцо камбия образуется
прямо из прокамбия и , следовательно,
на всём протяжении имеет первичное происхождении.
Деятельность этого кольца камбия ничем
не отличается от деятельности камбиального
кольца, возникающего более сложным путём
при пучковом типе строения стебля. Таким
образом, независимо от того, какова
была первичная структура стебля растений,
в результате вторичных изменений у них
может образоваться одинаковая структура:
общее камбиальное кольцо , внутрь от него
— ксилема, кнаружи — флоэма.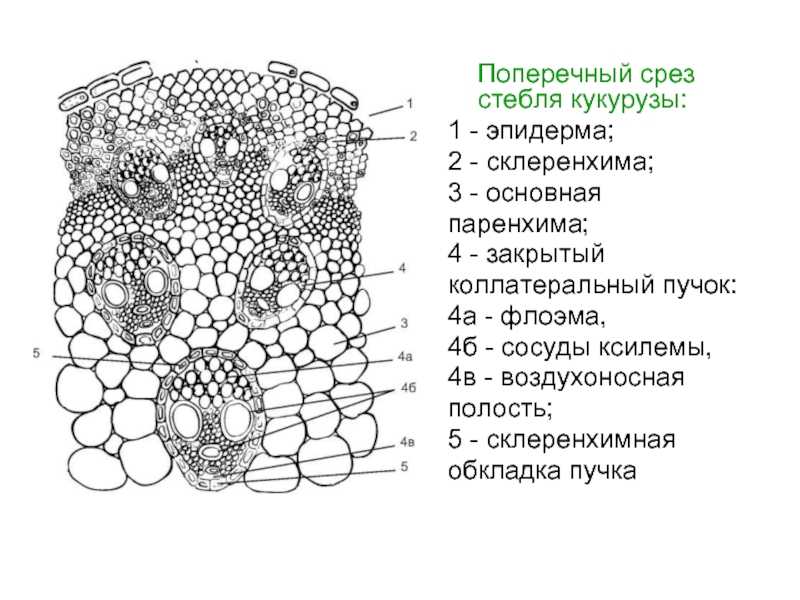 Так формируется
Так формируется
вторичная структура у большинства травянистых
растений.
404 Cтраница не найдена
Размер:
AAA
Изображения
Вкл.
Выкл.
Обычная версия сайта
К сожалению запрашиваемая страница не найдена.
Но вы можете воспользоваться поиском или картой сайта ниже
|
|
Внутренняя структура стебля двудольных растений
21 июня 2020 г.
Гаураб Карки
Ботаника
0
Анатомическое строение стебля двудольных растений
T.S. стебля двудольных показывает следующие внутренние особенности:
Эпидермис:
- Это самый внешний слой и имеет один слой паренхиматозных клеток.
- Обладает устьицами и большим количеством многоклеточных волосков (трихом).
- Наружные стенки сильно утолщены и кутинизированы.
- Клетки расположены компактно, межклеточного пространства не имеют.
- Эпидермис выполняет следующие функции:
- Минимизирует скорость транспирации благодаря толстой кутикуле
- Защищает подлежащие ткани от механических повреждений
- Предотвращает проникновение вредных организмов
- Способствует газообмену через устьица.
Гиподерма:
- Этот слой лежит ниже эпидермиса и состоит из 4 или 5 слоев колленхиматозных клеток.
- Эти клетки специально утолщены по углам на фоне межклетников за счет отложения целлюлозы и пектина.

- Клетки являются живыми в природе и могут содержать небольшое количество хлоропластов.
- Придает механическую прочность и эластичность периферической части стебля, особенно молодым и растущим органам.
- Они осуществляют фотосинтез, а также служат хранилищем пищи.

Кора:
- Расположена под гиподермой.
- Состоит из нескольких слоев тонкостенных крупных округлых или овальных живых паренхиматозных клеток с межклетниками.
- Клетки коры могут содержать некоторые хлоропласты, которые могут использоваться для производства пищевых материалов.
- Служат для хранения продуктов.
Энтодерма:
- Это единственный самый внутренний слой коры, который отделяет кору от сосудистых пучков.
- Клетки несколько бочкообразные, компактно расположенные, без межклетников, паренхиматозные.
- Обычно клетки содержат зерна крахмала, поэтому эндодерму можно назвать крахмальной оболочкой.
- Они служат запасом еды.
- Радиальная и поперечная стенки утолщены за счет отложений лигнина, образующих полоски каспария.
Перицикл:
- Находится между эндодермой и сосудистыми пучками.
- Обычно состоит из склеренхиматозных и паренхиматозных клеток.
- Склеренхима имеет форму полулунных участков над сосудистыми пучками, которые обеспечивают механическую поддержку частей растения.
- Точно так же паренхиматозный перицикл присутствует вне мозговых лучей, который служит для хранения пищи.
Сосудистые пучки:
- Расположены кольцом вокруг центральной сердцевины и внутри перицикла.
- Соединяемые, боковые, открытые и клиновидные.
- Размер пучков варьируется у разных видов.
- Каждый пучок имеет участок ксилемы по направлению к центру, участок флоэмы по направлению к периферии и полоску камбия между ними.
- Ксилема:
- Пролегает по направлению к сердцевине сосудистых пучков.
- Состоит из трахеид, сосудов, паренхимы ксилемы, волокон ксилемы.
- Трахеиды и сосуды состоят из меньшей протоксилемы и большей метаксилемы.
- Сначала формируется протоксилема, которая лежит ближе к центру, а затем формируется метаксилема, которая лежит ближе к периферии.
- Этот тип ксилемы называется эндархной ксилемой. Это помогает в проведении сока.
- Пролегает по направлению к сердцевине сосудистых пучков.
- Флоэма:
- Он расположен непосредственно под склеренхиматозным участком перицикла и состоит из следующих элементов, таких как ситовидные трубки, клетки-компаньоны и паренхима флоэмы.
- Он проводит продукты.
- Камбий:
- Расположен между ксилемой и флоэмой.
- Состоит из узкой полоски меристематических клеток с крупными ядрами и плотной цитоплазмой, называемой фасцикулярным камбием.
- Отвечает за вторичный рост толщины тела растения.

Сердцевина:
- Занимает центральную часть стержня.
- Состоит из тонкостенных паренхиматозных клеток округлой или многоугольной формы с межклетниками или без них.
- В этом регионе хранится еда.
- Сердцевинные лучи:
- Это тонкостенные радиально вытянутые паренхиматозные клетки, расположенные между сосудистыми пучками.
- Они хранят пищевые материалы и помогают во внутреннем перемещении воды.
Внутренняя структура стебля двудольных растений
- Внутренняя структура стебля двудольных растений
Copyright © 2022 | Тема WordPress от MH Themes
первичный рост — Inanimate Life
Как описано в главе 6, три органа сосудистых растений (корни, стебли и листья) имеют одинаковую базовую структуру: граница кожной ткани, включающая основную ткань, через которую проходит от одной до многих нитей сосудистой ткани. Три органа различаются по расположению сосудистой ткани: в корнях она встречается в виде одного центрального тяжа; в стеблях сосудистая ткань представлена множественными пучками, погруженными в основную ткань; а в листьях сосудистая ткань часто встречается в виде сетчатой сети жилок или в виде параллельных тяжей сосудистой ткани. В обоих случаях имеется основная ткань, заполняющая пространство между сосудистыми тяжами и кожной тканью. Эту основную анатомию легко увидеть в спарже, если обрезать основание и посмотреть на срезанный конец. Кожная ткань более жесткая снаружи, сосудистые пучки видны в виде небольших кругов, разбросанных по наружной части стебля, а основная ткань составляет остальное.
В обоих случаях имеется основная ткань, заполняющая пространство между сосудистыми тяжами и кожной тканью. Эту основную анатомию легко увидеть в спарже, если обрезать основание и посмотреть на срезанный конец. Кожная ткань более жесткая снаружи, сосудистые пучки видны в виде небольших кругов, разбросанных по наружной части стебля, а основная ткань составляет остальное.
Корни и побеги демонстрируют две полярности: радиальную полярность, означающую, что ткани и клетки различаются по мере продвижения кнаружи от центра (по радиусу), и проксимальную/дистальную полярность, означающую, что клетки на концах органов, где они расположены производятся, отличаются от клеток вдали от кончика, клетки, которые старше. Листья имеют полярность кончика к основанию и часто имеют полярность верх/низ.
Рис. 1 Проращивание семян подсолнечника. Кончик побега находится между двумя листочками (семядолями). Место соединения корня и стебля находится на поверхности почвы. Обратите внимание, что корень саженца на переднем плане (необычно) идет горизонтально.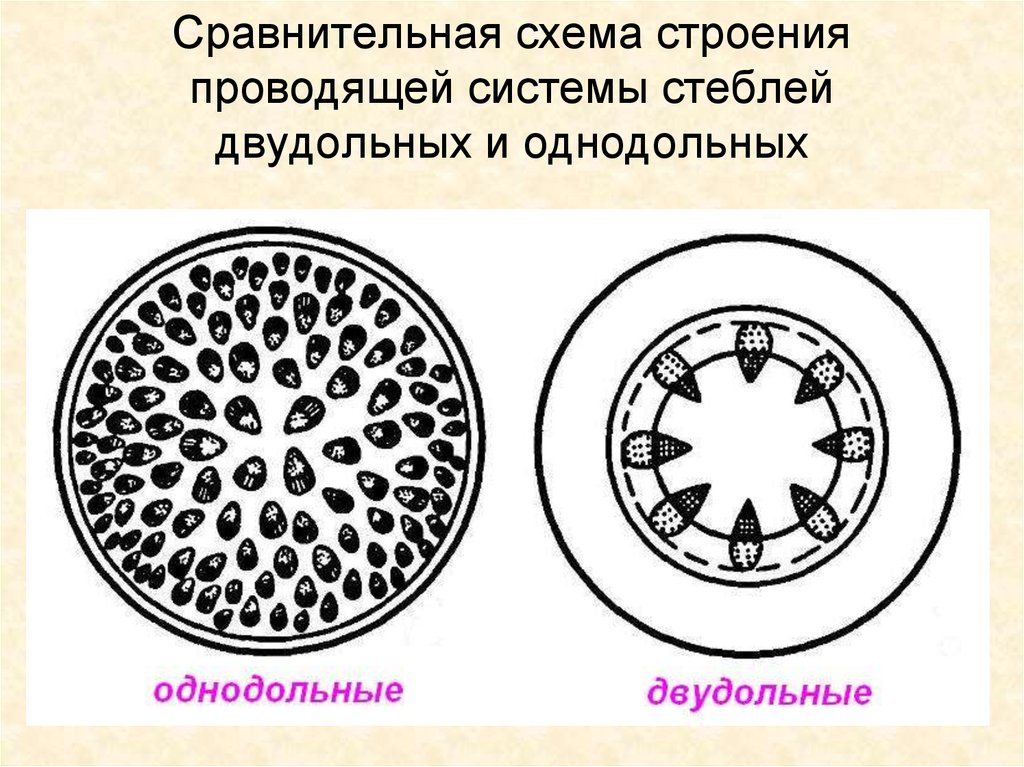 с многочисленными корневыми волосками
с многочисленными корневыми волосками
В этой главе мы более подробно описываем анатомию цветковых растений в результате первичного роста (рост за счет апикальных меристем корня или побега), а также рассматриваем изменения в развитии и, следовательно, закономерности, проявляющиеся с возрастом (расстояние от верхушки). .
- Развитие корня
- Анатомия зрелого корня
- Проявление побегов
- Анатомия зрелых побегов
- Развитие листьев
- Анатомия листьев
Развитие корня
Если корень разрезать вдоль длинной оси (т. е. продольный разрез), его развитие становится очевидным. Рядом с кончиком находится меристема, узнаваемая по небольшому размеру клеток и митотической активности, о чем часто свидетельствует появление хромосом. Двигаясь назад от кончика (проксимально, к телу растения), встречаются более старые и более зрелые клетки, узнаваемые, потому что они больше, больше не делятся и обладают чертами, которые отличают разные типы клеток, например, вторичные клеточные стенки трахеид и сосудистой трубки.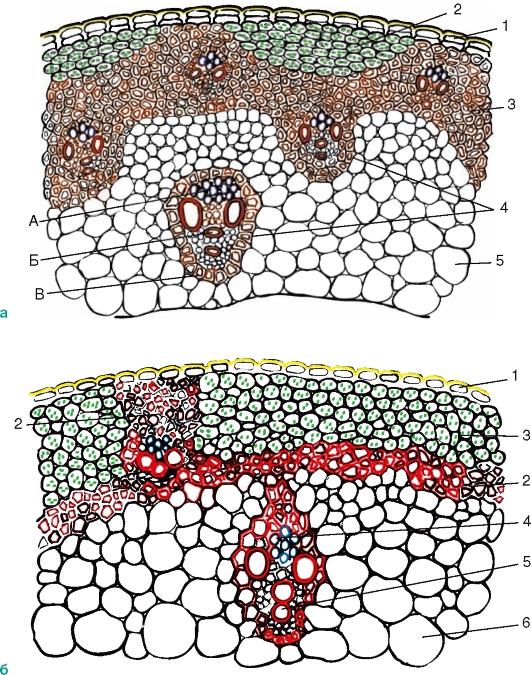 члены . Поскольку после перемещения на небольшое расстояние от верхушки корня больше не происходит деления или расширения клеток, диаметр корня, демонстрирующего только первичный рост, обычно постоянен по всей его длине, за исключением нескольких миллиметров на конце. (Корни, демонстрирующие вторичный рост, действительно увеличиваются в диаметре и обсуждаются в следующей главе). Клетки, продуцируемые апикальной меристемой корня, больше всего расширяются в дистальном/проксимальном направлении (вверх/вниз, при условии, что корень вертикальный) и производят клетки, вытянутые в этом направлении. Радиальное расширение гораздо меньше, поэтому корни в основном становятся длиннее, а не шире, и этот рост происходит возле кончика корня. Еще более важным, чем расширение отдельных клеток, является тот факт, что большинство клеточных делений в меристематической зоне делят клетки, так что большая часть образующихся дополнительных клеток находится в продольной (дистальной/проксимальной) плоскости. Это похоже на то, как одноклеточная нить делится, чтобы расшириться, с клеточными делениями, которые перпендикулярны длинной оси нити.
члены . Поскольку после перемещения на небольшое расстояние от верхушки корня больше не происходит деления или расширения клеток, диаметр корня, демонстрирующего только первичный рост, обычно постоянен по всей его длине, за исключением нескольких миллиметров на конце. (Корни, демонстрирующие вторичный рост, действительно увеличиваются в диаметре и обсуждаются в следующей главе). Клетки, продуцируемые апикальной меристемой корня, больше всего расширяются в дистальном/проксимальном направлении (вверх/вниз, при условии, что корень вертикальный) и производят клетки, вытянутые в этом направлении. Радиальное расширение гораздо меньше, поэтому корни в основном становятся длиннее, а не шире, и этот рост происходит возле кончика корня. Еще более важным, чем расширение отдельных клеток, является тот факт, что большинство клеточных делений в меристематической зоне делят клетки, так что большая часть образующихся дополнительных клеток находится в продольной (дистальной/проксимальной) плоскости. Это похоже на то, как одноклеточная нить делится, чтобы расшириться, с клеточными делениями, которые перпендикулярны длинной оси нити. Деления клеток в апикальной меристеме корня добавляют клетки в дистальном направлении, и лишь в ограниченной степени корни добавляют клетки радиально. Большинство корней имеют ширину примерно от 20 до 100 клеток (при условии только первичного роста), но корни часто имеют длину в миллионы или триллионы клеток.
Деления клеток в апикальной меристеме корня добавляют клетки в дистальном направлении, и лишь в ограниченной степени корни добавляют клетки радиально. Большинство корней имеют ширину примерно от 20 до 100 клеток (при условии только первичного роста), но корни часто имеют длину в миллионы или триллионы клеток.
Предполагая клеточное деление, которое увеличивает количество клеток в дистальной/проксимальной плоскости, второе ключевое соображение после клеточного деления заключается в том, является ли клетка, которая остается меристематической (и не растет), дистальной (по направлению к кончику) или проксимальной (по направлению к кончику). остальное растение). В подавляющем большинстве клеточных делений меристематических клеток корня клетка, которая остается меристематической, является дистальной, и расширение другой клетки выталкивает меристематический участок в почву. Однако в небольшой части клеток именно проксимальная клетка остается меристемной, а дистальная клетка созревает и становится частью защитного корневого чехлика, расположенного на крайнем дистальном конце корня. Клетки этого корневого чехлика постоянно отслаиваются по мере того, как корень проходит через почву, а корневой чехлик гарантирует, что сами меристематические клетки не отслаиваются.
Клетки этого корневого чехлика постоянно отслаиваются по мере того, как корень проходит через почву, а корневой чехлик гарантирует, что сами меристематические клетки не отслаиваются.
Изменения в развитии первичного корня
Проксимально от кончика корня встречаются следующие области, которые постепенно переходят друг в друга и перекрываются:
- зона клеточного деления, эмбриональная область, часто менее одного миллиметра
- зона расширения клеток, обычно протяженностью всего несколько миллиметров, область, в которой клетки удлиняются и, в гораздо меньшей степени, увеличиваются в диаметре; .
- зона созревания клеток, область, в которой клетки приобретают характерные черты. Эта зона простирается от менее сантиметра до нескольких сантиметров в длину. В самой молодой части этой зоны образуются корневые волоски, но они вскоре стареют и теряются из растения
Важные аспекты зоны созревания клеток включают (в порядке от кончика по мере продвижения проксимально):
- проводящие элементы флоэмы становятся функциональными
- водостойкий компаунд суберин депонируется в виде полоски каспариана
- проводящие элементы ксилемы становятся функциональными
- появление и исчезновение корневых волос. Корневые волоски являются отростками клеток дермы. Они образуются после того, как эти клетки перестали удлиняться, но присутствуют лишь короткое время перед старением. Следовательно, корневые волоски присутствуют только на относительно небольшом участке корня.
 Корневые волоски являются отростками клеток дермы. Они образуются после того, как эти клетки перестали удлиняться, но присутствуют лишь короткое время перед старением. Следовательно, корневые волоски присутствуют только на относительно небольшом участке корня.
Корневые волоски являются отростками клеток дермы. Они образуются после того, как эти клетки перестали удлиняться, но присутствуют лишь короткое время перед старением. Следовательно, корневые волоски присутствуют только на относительно небольшом участке корня.Значение полосы Каспариана
Эти гидрофобные отложения первоначально возникают в виде полосы, которая блокирует движение воды через стену снаружи внутрь. В конце концов вся энтодермальная клеточная стенка покрывается. Полоска каспария вынуждает воду и любые минералы, растворенные в воде, попадать в цитозоль в какой-то момент их пути между почвой и тканью ксилемы. До отложения каспариевой полоски, т. е. в самой молодой части корня, вода может двигаться из почвы к центру корня через апопласт ’, термин, описывающий совокупное пространство клеточных стенок и любые заполненные водой пространства между клетками, которые обычно включают не менее 10% объема ткани. Поскольку клетки эндодермы тесно связаны друг с другом, после отложения полоски каспария вода вынуждена двигаться через симпласт , чтобы пересечь эндодерму и попасть внутрь корня. Симпласт — это термин, описывающий совокупный объем цитозолей всех клеток, собирательный, поскольку все клетки связаны между собой плазмодесмами, цитоплазматическими нитями, окаймленными мембраной, которые проходят между клетками.
Поскольку клетки эндодермы тесно связаны друг с другом, после отложения полоски каспария вода вынуждена двигаться через симпласт , чтобы пересечь эндодерму и попасть внутрь корня. Симпласт — это термин, описывающий совокупный объем цитозолей всех клеток, собирательный, поскольку все клетки связаны между собой плазмодесмами, цитоплазматическими нитями, окаймленными мембраной, которые проходят между клетками.
Каспариевая полоска эндодермы после депонирования:
- позволяет растению регулировать, благодаря избирательной проницаемости клеточных мембран, какие минералы попадают и не попадают в ткань ксилемы, канал к верхушке растения
- позволяет растению при определенных условиях концентрировать растворенные вещества в ксилеме корня, потому что раствор апопласта внутри энтодермы (и связанный с тканью ксилемы) разделен двумя мембранами (одна обеспечивает вход в симпласт, другая обеспечивает выход из симпласт) из раствора апопласта вне эндодермы. Обратите внимание, что апопласт вне эндодермы непрерывен с почвой.
- уменьшает легкость, с которой вода может перемещаться из почвы в ксилему корня.
 Обратите внимание, что апопласт вне эндодермы непрерывен с почвой.
Обратите внимание, что апопласт вне эндодермы непрерывен с почвой.Рис. 2. Схема корня, показывающая корневые волоски, эпидермис, кору, эндодерму и стелу (центральное ядро сосудистой ткани). Синим цветом обозначен символический путь движения воды из почвы в стелу. Красным отмечен путь апопласта, за исключением того места, где каспариевая полоска энтодермы вынуждает воду проникать в симпласт.
Роль корневых волосков
Корневые волоски появляются, когда клетки эпидермиса образуют тонкий отросток, называемый корневым волоском, который идет перпендикулярно от корня в почву. Корневые волоски тонкие (~ 10 мкм), но могут уходить в почву на несколько миллиметров. Как обсуждалось ранее, хотя корневые волоски значительно увеличивают площадь, доступную для поглощения воды и питательных веществ, их более значительный эффект может заключаться в увеличении объема почвы на определенном расстоянии от корня.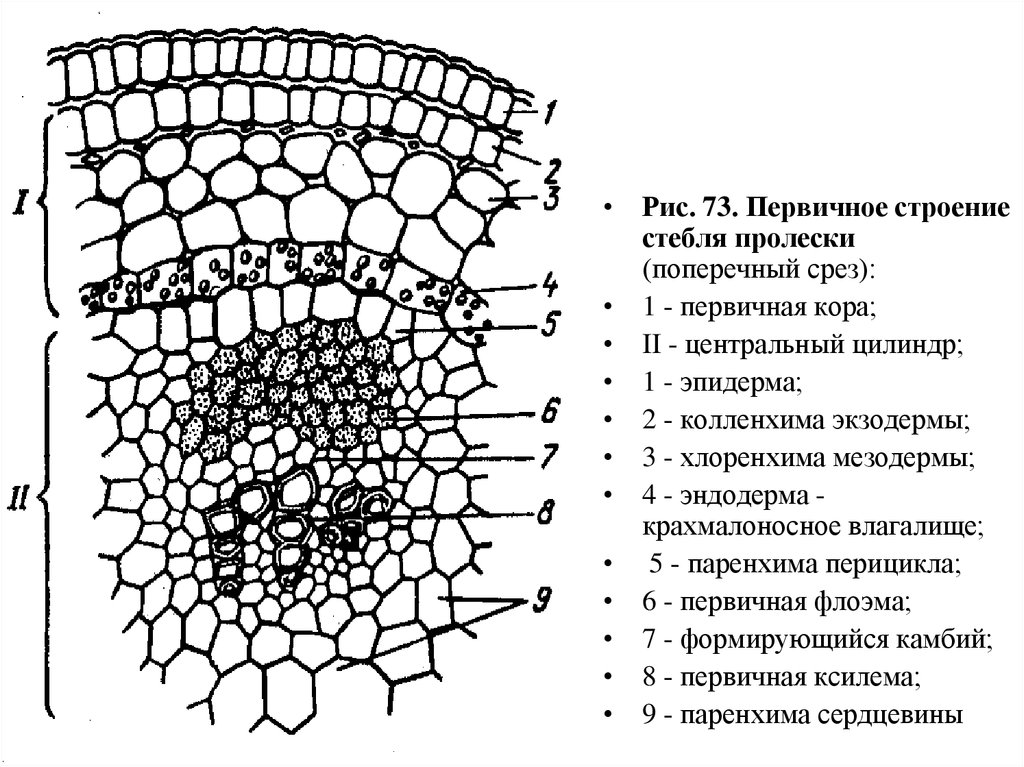 Учитывая корневые волоски, вода/питательные вещества могут:
Учитывая корневые волоски, вода/питательные вещества могут:
- проникают в цитозоль корневого волоска и затем проходят внутрь через симпласт
- проникают в клеточную стенку корневого волоска и продвигаются через апопласт внутрь корня (обратите внимание, что после отложения полоски каспария в энтодермальные клеточные стенки этот корень блокируется)
- обход корневых волосков и продвижение через почву к корню
Во влажных почвах вода может быстрее всего двигаться через почву, а не через корневые волоски, и возможно, что корневые волоски могут играть более важную роль в поглощении питательных веществ, чем в поглощении воды
Анатомия зрелого корня
Типичный поперечный срез корня показывает дермальную ткань снаружи, окружающую область основной ткани (кортикальный слой), которая окружает эндодерму, узнаваемую по тому, что опробковевший слой клеточной стенки окрашивается. Сразу внутри энтодермы находится перицикл, кольцо паренхимных клеток, которые можно стимулировать для образования корневых апикальных меристем, которые вырастают из корня и образуют боковые корни.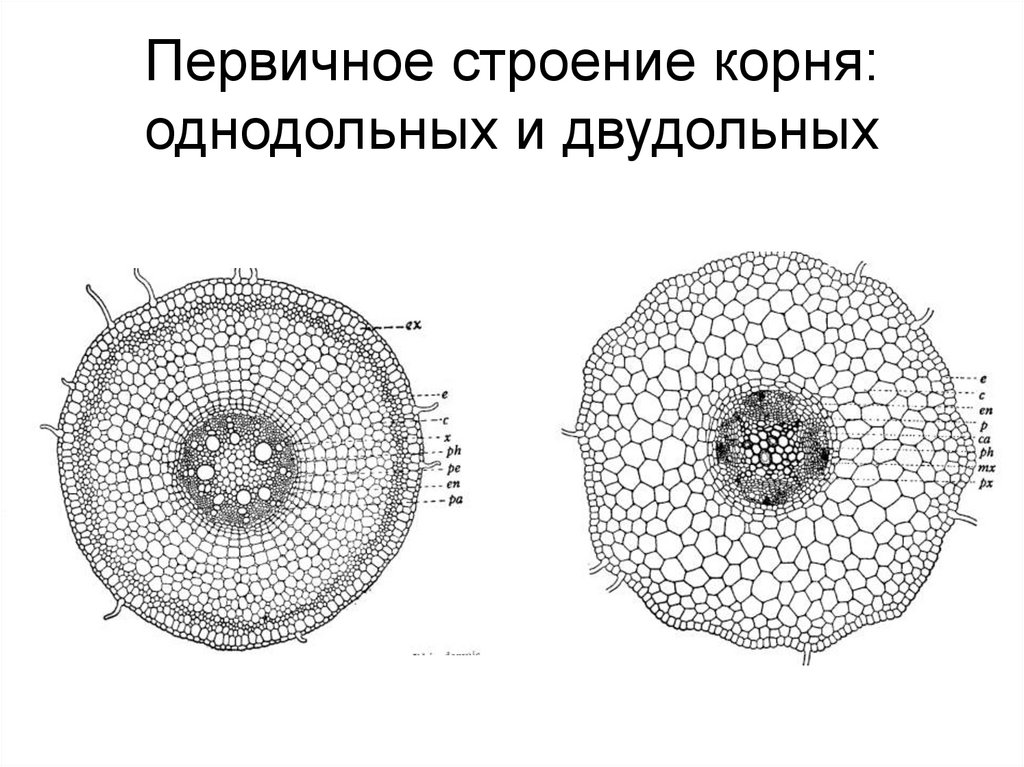 Внутри перицикла находится сосудистая ткань, которая у разных корней расположена по-разному. Некоторые корни имеют центральную сердцевину из паренхимных клеток, в то время как большинство корней имеют центральное твердое ядро из сосудистой ткани.
Внутри перицикла находится сосудистая ткань, которая у разных корней расположена по-разному. Некоторые корни имеют центральную сердцевину из паренхимных клеток, в то время как большинство корней имеют центральное твердое ядро из сосудистой ткани.
Рис. 3. Поперечный разрез зрелого корня.
Развитие побегов
Основная схема развития побегов такая же, как и у корней: терминальная область клеточного деления над областью роста клеток над областью созревания клеток. Но рост побега более сложен, чем рост корня, по нескольким причинам, один из которых заключается в том, что апикальная меристема побега не только удлиняет стебель, но также производит зародышевые листья (зачатки листьев) и ветвящиеся побеги (зачатки почек), расположенные непосредственно над листьями. Наличие этих структур делит стебель на узлы, места соединения листьев со стеблем, и междоузлия, промежутки между узлами. Как в зачатках листьев, так и в зачатках почек развивается сосудистая ткань, которая должна быть связана с сосудистой тканью основного стебля. Если бы кто-то проследил за сосудистой тканью в листе или ветке обратно к основному стеблю, то увидел бы один или несколько пучков сосудистой ткани, простирающихся от листа/ветви к стеблю. Этим объясняется наличие «сосудистых пучков» на стеблях цветковых растений: в основном они представляют собой следы сосудистой ткани, идущей к листьям и ветвям. В узлах видны сосудистые следы, отходящие от стебля и входящие в зачатки листьев и ветвей. Папоротники, хвощи, косолапые и ряд групп ископаемых растений имеют различные модели распределения сосудистой ткани внутри стебля (описываемые как «звездчатая структура»), и, поскольку сосудистая ткань часто представлена в окаменелостях, ее распределение было полезно при классификации окаменелых сосудистых растения.
Если бы кто-то проследил за сосудистой тканью в листе или ветке обратно к основному стеблю, то увидел бы один или несколько пучков сосудистой ткани, простирающихся от листа/ветви к стеблю. Этим объясняется наличие «сосудистых пучков» на стеблях цветковых растений: в основном они представляют собой следы сосудистой ткани, идущей к листьям и ветвям. В узлах видны сосудистые следы, отходящие от стебля и входящие в зачатки листьев и ветвей. Папоротники, хвощи, косолапые и ряд групп ископаемых растений имеют различные модели распределения сосудистой ткани внутри стебля (описываемые как «звездчатая структура»), и, поскольку сосудистая ткань часто представлена в окаменелостях, ее распределение было полезно при классификации окаменелых сосудистых растения.
Зачатки листьев образуются по характерному образцу, который зависит от вида растений. Наиболее легко описываемая модель — это когда листья образуются парами на противоположных сторонах стебля. Последовательно листья образуются парами с поворотом на 90 90 284 o 90 285 относительно предыдущей пары листьев, т. е. если вы смотрите на стебель сверху и первая пара листьев находится на севере и юге, следующий набор листья были бы восток и запад. Следующий набор произведенных листьев вернется к исходной ориентации север-юг. Большинство растений имеют более сложный филлотаксия , то есть расположение листьев, которое можно описать, подсчитав количество новых листьев и количество оборотов вокруг стебля, прежде чем вы окажетесь с листом прямо под другим листом.
е. если вы смотрите на стебель сверху и первая пара листьев находится на севере и юге, следующий набор листья были бы восток и запад. Следующий набор произведенных листьев вернется к исходной ориентации север-юг. Большинство растений имеют более сложный филлотаксия , то есть расположение листьев, которое можно описать, подсчитав количество новых листьев и количество оборотов вокруг стебля, прежде чем вы окажетесь с листом прямо под другим листом.
В то время как удлинение корней, вызванное расширением вновь продуцируемых клеток, обычно устойчиво во времени и пространстве, происходит вскоре после продуцирования клеток и очень близко к апикальной меристеме корня, удлинение стеблей не столь устойчиво. Удлинение побегов происходит за счет удлинения междоузлий. У некоторых растений рост междоузлий происходит близко к верхушке побега, в результате чего образуется стебель с листьями, которые отделяются от каждого вблизи верхушки побега со скоростью, соответствующей образованию новых зачатков листьев. У других растений рост междоузлий задерживается или отсутствует, в результате чего образуются очень короткие стебли с множеством листьев, расположенных очень близко друг к другу (розетки). Такая структура может быть постоянной или может быть временной до тех пор, пока не будет получен конкретный сигнал, и стебель не «вытянется», быстро удлиняясь за счет увеличения пространства между листьями, например, у салата или шпината. Некоторые однодольные (см. ниже), в частности злаки, имеют меристематические зоны в узлах стеблей и в основании листовой пластинки травы. Эти меристемы активизируются, если стебель или лист над ними повреждаются (обычно от выпаса).
У других растений рост междоузлий задерживается или отсутствует, в результате чего образуются очень короткие стебли с множеством листьев, расположенных очень близко друг к другу (розетки). Такая структура может быть постоянной или может быть временной до тех пор, пока не будет получен конкретный сигнал, и стебель не «вытянется», быстро удлиняясь за счет увеличения пространства между листьями, например, у салата или шпината. Некоторые однодольные (см. ниже), в частности злаки, имеют меристематические зоны в узлах стеблей и в основании листовой пластинки травы. Эти меристемы активизируются, если стебель или лист над ними повреждаются (обычно от выпаса).
В отличие от корней, на дистальной стороне апикальной меристемы побега не происходит созревания клеток, т. е. нет «чепчика побега» (ср. корневой чехлик), происходящего из апикальной меристемы побега. Как правило, эта апикальная меристема не проталкивается через почву, поэтому функция, которую обеспечивает корневой чехлик, обычно не требуется.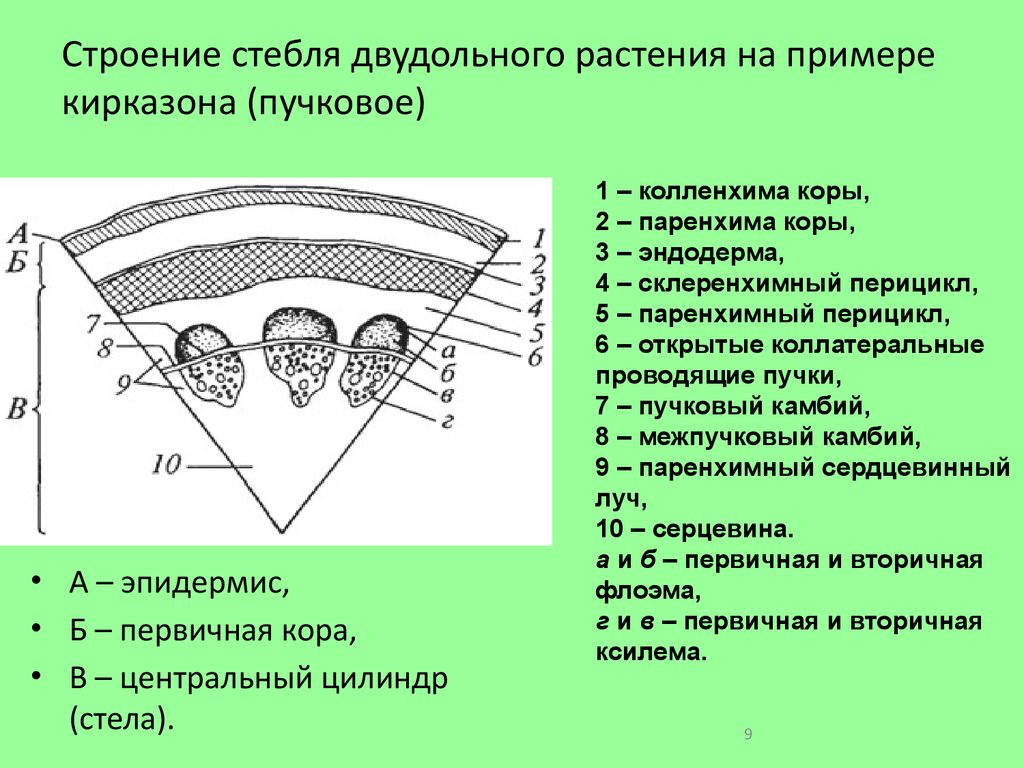 Однако верхушечная меристема побега покрыта молодыми листьями, которые растут очень близко к верхушке побега и способны его покрыть. Некоторые стебли, называемые корневищами, действительно прорастают сквозь почву, и они защищены модифицированными листьями, называемыми катафиллами, которые защищают апикальную меристему побега и образуют заостренную структуру, которую легче протолкнуть через почву. Подобные видоизмененные листья, называемые почковыми чешуями, покрывают верхушечные меристемы побегов древесных растений в течение продолжительных периодов, когда они не растут. В то время как большинство корней часто активно растут большую часть года, в том числе в течение вегетационного периода, рост побегов растений в сезонных местообитаниях часто длится очень долго. Многие растения, особенно деревья, демонстрируют рост (удлинение стеблей) в течение короткого периода, часто менее 30 дней весной. Однако деление клеток побегов происходит намного раньше, чем рост клеток, часто на восемь месяцев раньше, то есть некоторые новые клетки, разрастающиеся весной, образовались в конце лета предыдущего года.
Однако верхушечная меристема побега покрыта молодыми листьями, которые растут очень близко к верхушке побега и способны его покрыть. Некоторые стебли, называемые корневищами, действительно прорастают сквозь почву, и они защищены модифицированными листьями, называемыми катафиллами, которые защищают апикальную меристему побега и образуют заостренную структуру, которую легче протолкнуть через почву. Подобные видоизмененные листья, называемые почковыми чешуями, покрывают верхушечные меристемы побегов древесных растений в течение продолжительных периодов, когда они не растут. В то время как большинство корней часто активно растут большую часть года, в том числе в течение вегетационного периода, рост побегов растений в сезонных местообитаниях часто длится очень долго. Многие растения, особенно деревья, демонстрируют рост (удлинение стеблей) в течение короткого периода, часто менее 30 дней весной. Однако деление клеток побегов происходит намного раньше, чем рост клеток, часто на восемь месяцев раньше, то есть некоторые новые клетки, разрастающиеся весной, образовались в конце лета предыдущего года.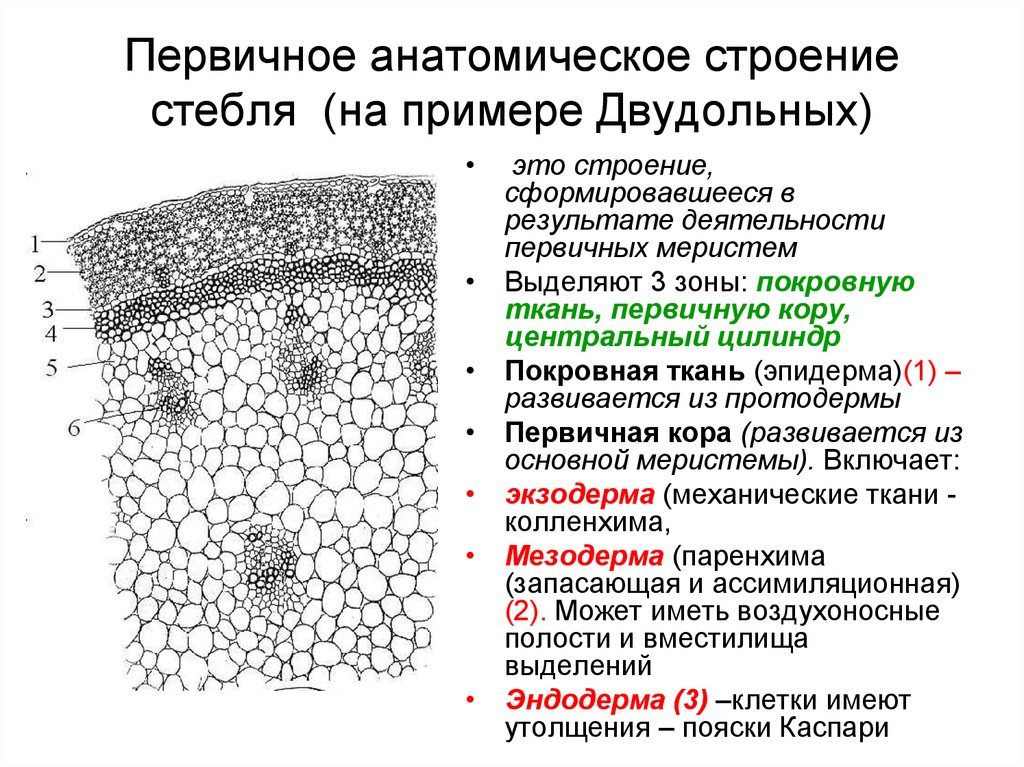
Анатомия зрелых побегов
Традиционно цветковые растения делили на две группы, однодольные и двудольные, на основании ряда признаков, одним из которых была анатомия стебля. В то время как группа однодольных по-прежнему считается допустимой филогенетической единицей, большинство исследователей считают «двудольные» искусственной группой и разделили двудольные на «евдидольные» и несколько других групп. Подавляющее большинство представителей бывшей группы двудольных являются эвдикотами, и в приведенном ниже обсуждении мы будем использовать термин двудольные.
Стебли двудольных имеют сосудистые пучки, расположенные кольцом близко к краю стебля. К тканям, идущим снаружи внутрь, относятся: эпидермис, кора, сосудистые пучки (в кольце, с переменным количеством основной ткани между пучками), сердцевина.
Стебли однодольных растений отличаются от стеблей двудольных тем, что по всему стеблю разбросаны сосудистые пучки.
Рис. 4 Клевер (Trifolium), двудольное растение, стебель с сосудистыми пучками, расположенными в виде кольца с большой центральной сердцевиной.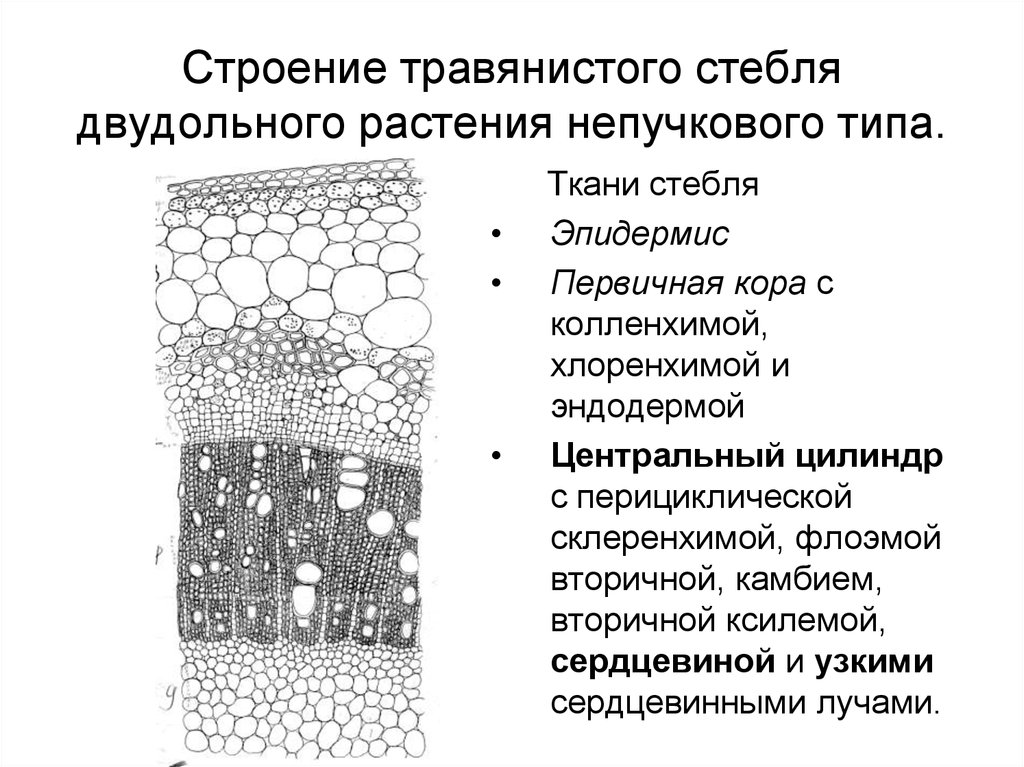 Сосудистые пучки имеют клетки склеренхимы снаружи и ткани флоэмы и ксилемы по мере продвижения внутрь. Рис. 5 Zea (кукуруза, однодольное растение), поперечный срез стебля, показывающий многочисленные сосудистые пучки, погруженные в основную ткань. Рис. 6. Поперечный разрез стебля двудольных, показывающий сосудистый пучок. Снаружи: кожная ткань, кора, склеренхима (окрашена зеленым цветом с толстыми стенками), ткань флоэмы (окрашена в синий цвет), ткань ксилемы (окрашена в красный цвет с толстыми клеточными стенками) окрашенные клетки. Это в увеличенном виде ниже. Рис. 8. Продольный разрез, показывающий сосуды. Обратите внимание, что обычно вторичная клеточная стенка трубчатых элементов сосудов не откладывается непрерывно и часто имеет форму колец или спиралей. Также в красный цвет окрашены волокна склеренхимы, вторичные стенки которых отложены непрерывно.
Сосудистые пучки имеют клетки склеренхимы снаружи и ткани флоэмы и ксилемы по мере продвижения внутрь. Рис. 5 Zea (кукуруза, однодольное растение), поперечный срез стебля, показывающий многочисленные сосудистые пучки, погруженные в основную ткань. Рис. 6. Поперечный разрез стебля двудольных, показывающий сосудистый пучок. Снаружи: кожная ткань, кора, склеренхима (окрашена зеленым цветом с толстыми стенками), ткань флоэмы (окрашена в синий цвет), ткань ксилемы (окрашена в красный цвет с толстыми клеточными стенками) окрашенные клетки. Это в увеличенном виде ниже. Рис. 8. Продольный разрез, показывающий сосуды. Обратите внимание, что обычно вторичная клеточная стенка трубчатых элементов сосудов не откладывается непрерывно и часто имеет форму колец или спиралей. Также в красный цвет окрашены волокна склеренхимы, вторичные стенки которых отложены непрерывно.
Развитие листьев
В отличие от корней и побегов листья представляют собой детерминированные структуры, характер развития которых сравним с развитием человека и лягушки.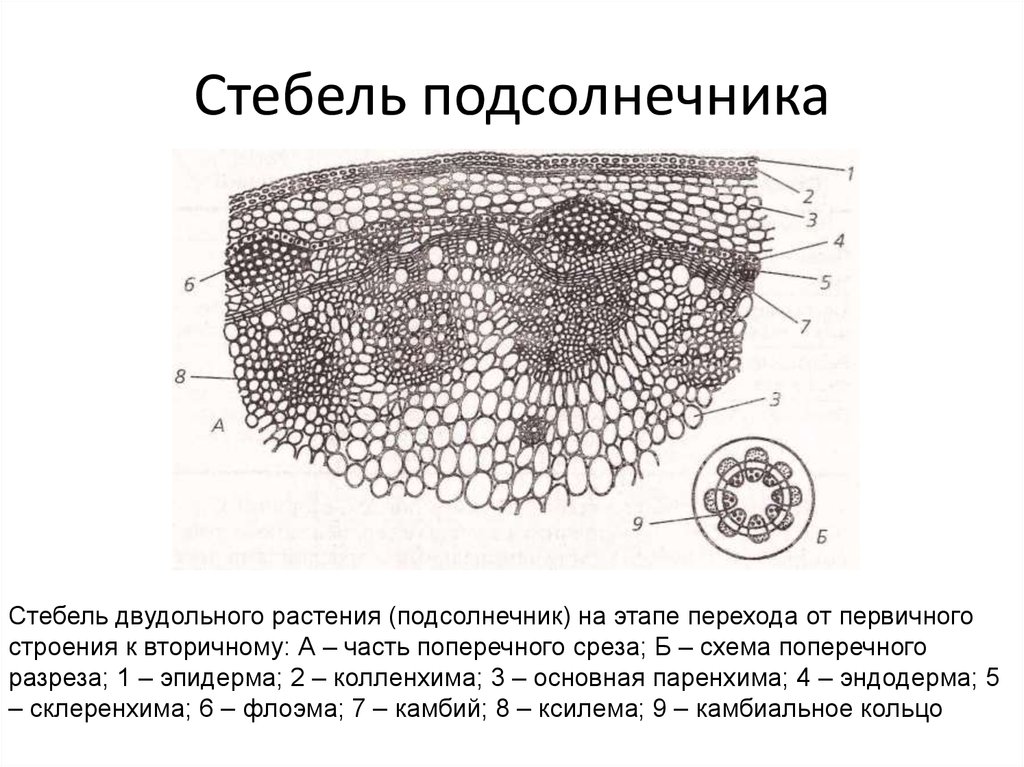 Они начинаются как зародыш, называемый зачатком листа, клетки которого делятся и впоследствии растут в течение определенного периода времени, образуя трехмерную форму, которая часто значительно сложнее, чем цилиндрическая структура корней и стеблей. Более того, разнообразие форм листьев, характерное для сосудистых растений, особенно цветковых, намного превышает разнообразие форм корней или стеблей. После прекращения клеточного деления и роста лист остается во «взрослой» форме в течение (обычно) определенного периода времени, прежде чем он претерпевает окончательный процесс старения, который в конечном итоге приводит к отделению листа от растения (опадению). Старение и опадение листьев будут обсуждаться позже, но здесь важно подчеркнуть, что эти процессы развития очень важны для жизни растения.
Они начинаются как зародыш, называемый зачатком листа, клетки которого делятся и впоследствии растут в течение определенного периода времени, образуя трехмерную форму, которая часто значительно сложнее, чем цилиндрическая структура корней и стеблей. Более того, разнообразие форм листьев, характерное для сосудистых растений, особенно цветковых, намного превышает разнообразие форм корней или стеблей. После прекращения клеточного деления и роста лист остается во «взрослой» форме в течение (обычно) определенного периода времени, прежде чем он претерпевает окончательный процесс старения, который в конечном итоге приводит к отделению листа от растения (опадению). Старение и опадение листьев будут обсуждаться позже, но здесь важно подчеркнуть, что эти процессы развития очень важны для жизни растения.
Интеркалярные меристемы присутствуют в некоторых листьях, в частности в листьях трав, и позволяют листу, кончик (дистальная часть) которого была задета, возобновить рост и заменить утраченную область фотосинтеза. Способность расти таким образом была особенно важна для успеха трав.
Способность расти таким образом была особенно важна для успеха трав.
Структура листа
Многие листья демонстрируют полярность между верхней и нижней поверхностью, при этом нижняя эпидерма имеет регулируемые поры и устьица, а верхняя поверхность — нет. Кроме того, основная ткань верхней части листа состоит из клеток, ориентированных параллельно друг другу (эта область называется палисадным мезофиллом), а клетки мезофилла нижней части листа (так называемый губчатый мезофилл) имеет клетки, которые непоследовательно ориентированы друг относительно друга и имеют гораздо большие воздушные промежутки между отдельными клетками. У многих листьев однодольных жилки идут параллельно друг другу, и это означает, что если вы сделаете поперечный разрез листа, вы, вероятно, увидите поперечные срезы всех сосудистых пучков. У большинства листьев двудольных жилки проходят под разными углами, и поперечный срез листа вряд ли покажет поперечные срезы через какой-либо другой пучок, кроме главной центральной жилки.
Рис. 9 Лист сирени, показывающий верхний и нижний эпидермис, центральную жилку в поперечном сечении с окрашенной в красный цвет сосудистой тканью, палисадный слой, состоящий из двух рядов клеток, и слой губчатого мезофилла.
Рис. 10 Поперечное сечение листа кукурузы, показывающее множественные сосудистые пучки и мезофилл, не разделенный на палисадный и губчатый компоненты. Также видны увеличенные клетки, окружающие сосудистые пучки (клетки оболочки пучков), которые важны для их пути фотосинтеза (C4).
См. следующие сайты, которые также были перечислены в Главе 6.
Веб-сайты с прекрасными изображениями клеток и тканей семенных растений:
- Лаборатория Остина Маусет Техасского университета Содержание
- Ботанические ресурсы Университета Висконсина
- «Увидеть скрытые микроскопические чудеса растительного царства» Михаила Грешко. Хорошие изображения клеток растений (и некоторых водорослей).
- https://www. nationalgeographic.com/science/article/photography-plants-cells-diatoms-xylem-phloem-microscope
- https://www.
- Ткани и органы растений из библиотеки изображений биологических наук Беркширского муниципального колледжа. Слайды по анатомии растений.
- http://blogs.berkshirecc.edu/bccoer/plants/
- Таблицы анатомии растений от BibliOdyssey. Ботанические карты.
- http://bibliodyssey.blogspot.com/2012/12/plant-anatomy-charts.html
 nationalgeographic.com/science/article/photography-plants-cells-diatoms-xylem-phloem-microscope
nationalgeographic.com/science/article/photography-plants-cells-diatoms-xylem-phloem-microscopeMedia Attributions
- Sunflower_seedlings © Bluemoose~commonswiki находится под лицензией CC BY-NC-SA (Attribution NonCommercial ShareAlike)
- В сталь: поглощение воды и минералов корнями © Kelvinsong находится под лицензией CC BY-SA (Attribution ShareAlike)
- Стебель клевера © Berkshire Community College Bioscience Image Library используется под лицензией CC0 (Creative Commons Zero)