
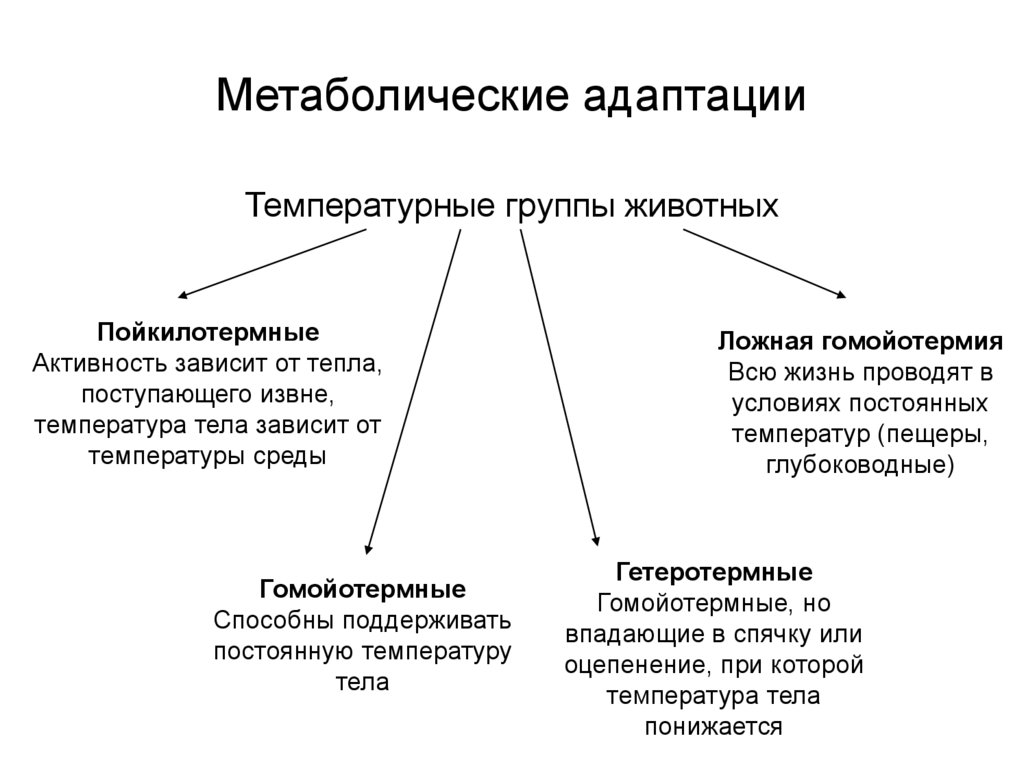
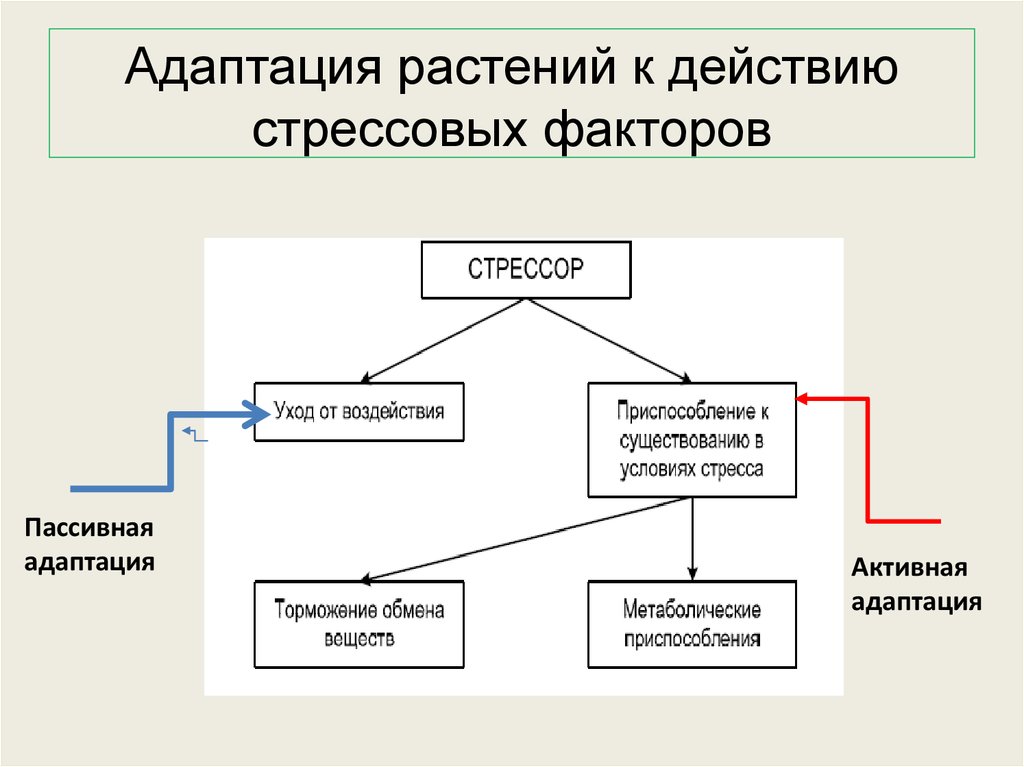
Содержание
Приспособления растений к высоким и низким температурам
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
1. Приспособления растений к высоким и низким температурам
2. высокие температуры
• В горячих источниках обнаружены:
Синезеленые
водоросли (+85°)
Тионовые бактерии
(до + 90°)
Водоросль Oscillatoria, живущая на о.
Ява в воде с температурой 64°С,
погибает при 68°С уже через 5—10 мин!
4. высокие температуры
Жаровыносливые растения пустынь
Синезеленые водоросли
Верблюжья
колючка
Джузгун
Янтак
(70 — 80°)
5.
 Опасность перегрева
Опасность перегрева
Потеря воды
Появление ожогов
Разрушение хлорофилла
Нарушения обмена веществ
Гибель растения
6. Приспособления к высоким температурам
• Высокий уровень
транспирации (Растения
сильнее испаряют воду – это
охлаждает и защищает от
перегрева).
• Состояние летнего покоя
(надземные побеги отмирают, а
в почве остаются подземные с
некоторым запасом воды)
7. Приспособления к высоким температурам
• Листья покрыты либо пушком либо
восковым налетом, что уменьшает
площадь испарения листьев.
Коровяк
«медвежье ухо»
8. Приспособления к высоким температурам
• Листовая пластинка повернута ребром к
горячим полуденным лучам.
Дикий салат — Латук
9. Приспособления к высоким температурам
• Весенняя вегетация (созревание
семян и луковиц до наступления летней
жары и сухости)
Крупка, или веснянка
весенняя
Лук гадючий
Мятлик луковичный
10.
 Приспособления растений к низким температурам
Приспособления растений к низким температурам
• Перед зимой у растений увеличивается
содержание сахаров и жиров.
• Выделение специальных веществ
(глицерина и многоатомных спиртов),
которые защищают молодые побеги
(побеги становятся красно-бурого цвета)
Приспособления растений к
низким температурам
Уход от повреждающего действия
фактора (пассивная адаптация) — растения
до наступления осенних холодов успевают
дать семена
Закаливание:
• замедление или полная остановка
ростовых процессов в ночное время;
• «перестройка» цитоплазмы – запас
устойчивых к холоду белков для
уменьшения внутриклеточного
льдообразования
12. Классификация растений по отношению к теплу и холоду
13. Термофилы 1. Нехолодостойкие растения
■
Переносят довольно высокие температуры,
но могут пострадать при невысоких
положительных температурах
Какао
Сенполия
14. 2. Неморозостойкие растения
■
Переносят низкие температуры, пока в
теле растения не образуется лед
Лимон
Чай
15.
 Психрофилы Льдоустойчивые растения
Психрофилы Льдоустойчивые растения
■
В тканях этих растений образуются
кристаллики льда, сначала в межклетниках,
а потом и в клетках
осина
ель
береза
16. Морозобойные трещины
■
Образуются в холодные зимы при
t = — 35 — 43°С
Проверка ДЗ:
Как человек изменяет
температурные условия
для культурных
растений?
Проверка ДЗ:
Подготовиться к тесту по
приспособлениям
растений к наземновоздушной среде
обитания
English
Русский
Правила
Приспособления растений к высоким и низким температурам презентация, доклад
Приспособления растений к высоким и низким температурам
высокие температуры
В горячих источниках обнаружены:
Синезеленые водоросли (+85°)
Тионовые бактерии (до + 90°)
Водоросль Oscillatoria, живущая на о.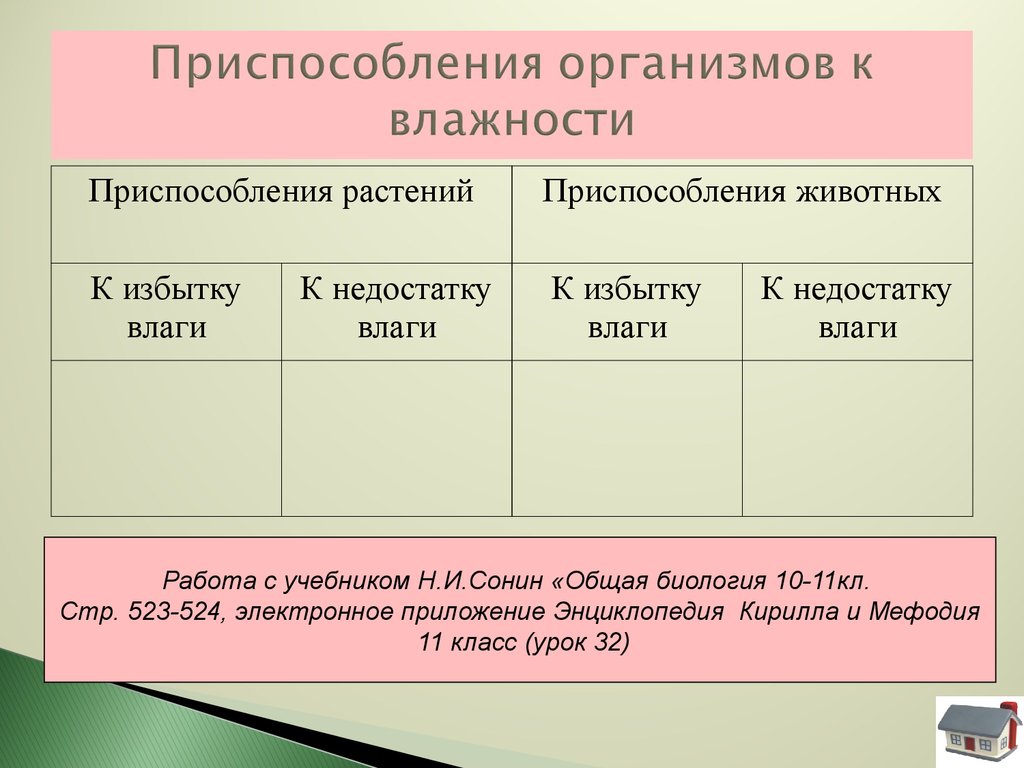 Ява в воде с температурой 64°С, погибает при 68°С уже через 5—10 мин!
Ява в воде с температурой 64°С, погибает при 68°С уже через 5—10 мин!
высокие температуры
Жаровыносливые растения пустынь
Синезеленые водоросли
(70 — 80°)
Янтак
Джузгун
Верблюжья колючка
Опасность перегрева
Потеря воды
Появление ожогов
Разрушение хлорофилла
Нарушения обмена веществ
Гибель растения
Приспособления к высоким температурам
Высокий уровень транспирации (Растения сильнее испаряют воду – это охлаждает и защищает от перегрева).
Состояние летнего покоя (надземные побеги отмирают, а в почве остаются подземные с некоторым запасом воды)
Приспособления к высоким температурам
Листья покрыты либо пушком либо восковым налетом, что уменьшает площадь испарения листьев.
Коровяк
«медвежье ухо»
Приспособления к высоким температурам
Листовая пластинка повернута ребром к горячим полуденным лучам.
Дикий салат — Латук
Приспособления к высоким температурам
Весенняя вегетация (созревание семян и луковиц до наступления летней жары и сухости)
Крупка, или веснянка
весенняя
Бурачок,
или алиссум
Тюльпан
Лук гадючий
Мятлик луковичный
Приспособления растений к низким температурам
Перед зимой у растений увеличивается содержание сахаров и жиров.
Выделение специальных веществ (глицерина и многоатомных спиртов), которые защищают молодые побеги (побеги становятся красно-бурого цвета)
Уход от повреждающего действия фактора (пассивная адаптация) — растения до наступления осенних холодов успевают дать семена
Закаливание:
замедление или полная остановка ростовых процессов в ночное время;
«перестройка» цитоплазмы – запас устойчивых к холоду белков для уменьшения внутриклеточного льдообразования
Приспособления растений к низким температурам
Классификация растений по отношению к теплу и холоду
Термофилы
1. Нехолодостойкие растения
Нехолодостойкие растения
Переносят довольно высокие температуры, но могут пострадать при невысоких положительных температурах
Какао
Сенполия
2. Неморозостойкие растения
Переносят низкие температуры, пока в теле растения не образуется лед
Лимон
Чай
Психрофилы
Льдоустойчивые растения
В тканях этих растений образуются кристаллики льда, сначала в межклетниках, а потом и в клетках
ель
осина
береза
Морозобойные трещины
Образуются в холодные зимы при
t = — 35 — 43°С
Проверка ДЗ:
Как человек изменяет температурные условия для культурных растений?
Проверка ДЗ:
Подготовиться к тесту по приспособлениям растений к наземно-воздушной среде обитания
Скачать презентацию
Адаптация растений к частым изменениям между высокими и низкими температурами: ремоделирование мембранных липидов и поддержание уровня ненасыщенности
. 2011 сен; 34 (9): 1431-42.
2011 сен; 34 (9): 1431-42.
doi: 10.1111/j.1365-3040.2011.02341.x.
Epub 2011 20 июня.
Гуовэй Чжэн
1
, Бо Тянь, Фуцзюань Чжан, Фацин Тао, Вейци Ли
принадлежность
- 1 Ключевая лаборатория биоразнообразия и биогеографии, Куньминский институт ботаники, Китайская академия наук, Куньмин 650204, Китай.
PMID:
21486310
PMCID:
PMC3980542
DOI:
10.1111/j.1365-3040.2011.02341.x
Бесплатная статья ЧВК
Guowei Zheng et al.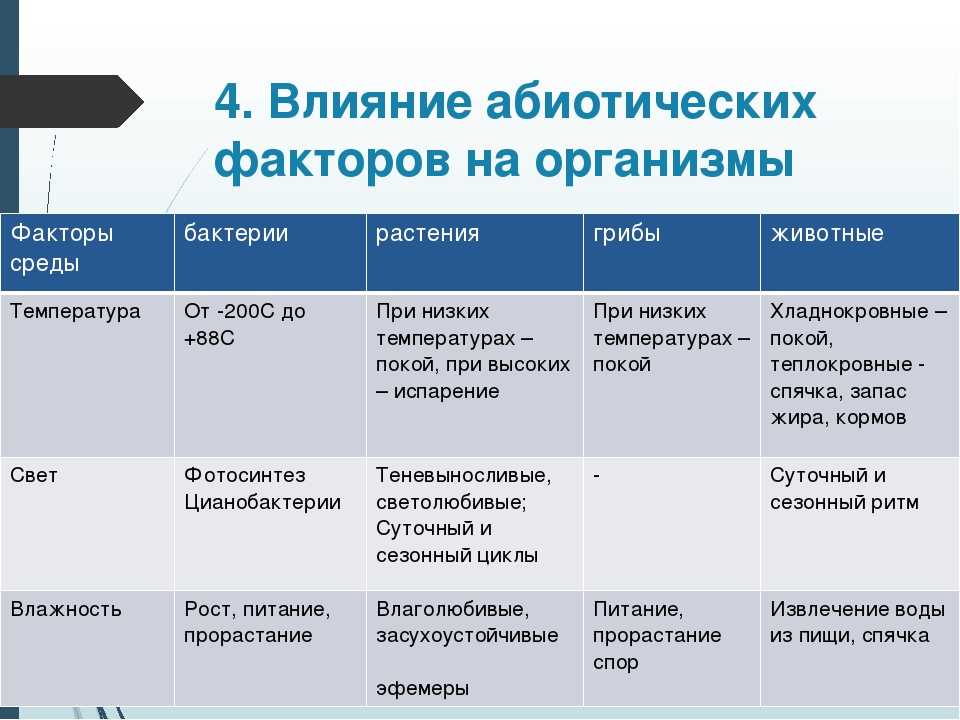
Окружающая среда растительной клетки.
2011 Сентябрь
Бесплатная статья ЧВК
. 2011 сен; 34 (9): 1431-42.
doi: 10.1111/j.1365-3040.2011.02341.x.
Epub 2011 20 июня.
Авторы
Гуовэй Чжэн
1
, Бо Тянь, Фуцзюань Чжан, Фацин Тао, Вэйци Ли
принадлежность
- 1 Ключевая лаборатория биоразнообразия и биогеографии, Куньминский институт ботаники, Китайская академия наук, Куньмин 650204, Китай.
PMID:
21486310
PMCID:
ЧВК3980542
DOI:
10.
 1111/j.1365-3040.2011.02341.x
1111/j.1365-3040.2011.02341.x
 1111/j.1365-3040.2011.02341.x
1111/j.1365-3040.2011.02341.xАбстрактный
Одна из основных стратегий адаптации растений к изменению температуры заключается в снижении степени ненасыщенности мембранных липидов при высокой температуре и повышении ее при низкой температуре. Мы предполагаем, что эта стратегия не может быть принята растениями в экосистемах и средах с частыми изменениями между высокими и низкими температурами, потому что изменения ненасыщенности липидов сложны и требуют больших затрат энергии. Чтобы проверить эту гипотезу, мы использовали липидомный подход для определения профиля изменений в молекулярных видах мембранных глицеролипидов у двух видов растений, взятых с альпийских осыпей, и у двух других видов растений, выращенных в камере для выращивания, с ежедневным циклическим изменением температуры между теплом и замораживанием. Мы обнаружили, что шесть классов фосфолипидов и два класса галактолипидов показали значительные изменения, но степень ненасыщенности общих липидов и трех классов лизофосфолипидов осталась неизменной. Этот характер изменений мембранных липидов отличался от такового при медленных изменениях температуры. Мы предлагаем два типа моделей адаптации растений к изменению температуры: (1) ремоделирование мембранных липидов, но сохранение степени ненасыщенности используются для адаптации к частым изменениям температуры; и (2) ремоделирование и изменение степени ненасыщенности для адаптации к нечастым изменениям температуры.
Этот характер изменений мембранных липидов отличался от такового при медленных изменениях температуры. Мы предлагаем два типа моделей адаптации растений к изменению температуры: (1) ремоделирование мембранных липидов, но сохранение степени ненасыщенности используются для адаптации к частым изменениям температуры; и (2) ремоделирование и изменение степени ненасыщенности для адаптации к нечастым изменениям температуры.
© 2011 Блэквелл Паблишинг Лтд.
Цифры
Рисунок 1
Изменение температуры воздуха с…
Рисунок 1
Изменение температуры воздуха в зависимости от времени суток и времени отбора проб (обозначается…
фигура 1
Изменение температуры воздуха в зависимости от времени суток и времени отбора проб (указано стрелками) в альпийских осыпях гор Хэндуань в октябре. Температуры во время отбора проб составляли -1°С в 00:00, -3°С в 06:00, 32°С в 13:00 и 6°С в 18:00.
Температуры во время отбора проб составляли -1°С в 00:00, -3°С в 06:00, 32°С в 13:00 и 6°С в 18:00.
Рисунок 2
Суточный цикл температуры в пределах…
Рисунок 2
Суточный температурный цикл в ростовой камере. Стрелки указывают выборку…
фигура 2
Суточный температурный цикл в ростовой камере. Стрелки указывают время отбора проб после того, как A. thaliana и C. himalaica выращивались в камере в течение 30 дней. Температуры во время отбора проб составляли -7°С в 00:00, 22°С в 12:00, 45°С в 15:00 и 22°С в 20:00.
Рисунок 3
Изменения молекулярных видов…
Рисунок 3
Изменения молекулярных видов мембранных липидов у S. medusa (А) и…
medusa (А) и…
Рисунок 3
Изменения молекулярных видов мембранных липидов у S. medusa (А) и S.-L. linearifolia (B) при смене высоких и низких температур на альпийских осыпях. Значения являются средними ± SD ( n = 3, 4 или 5).
Рисунок 3
Изменения молекулярных видов…
Рисунок 3
Изменения молекулярных видов мембранных липидов у S. medusa (А) и…
Рисунок 3
Изменения молекулярных видов мембранных липидов в S. medusa (A) и S.-L. linearifolia (B) при смене высоких и низких температур на альпийских осыпях. Значения являются средними ± SD ( n = 3, 4 или 5).
Рисунок 4
Изменения молекулярных видов…
Рисунок 4
Изменения молекулярных видов лизоФЛ в S. medusa и S.-L. линейнолистный…
medusa и S.-L. линейнолистный…
Рисунок 4
Изменения молекулярных видов lysoPL у S. medusa и S.-L. linearifolia при смене высоких и низких температур на альпийских осыпях. Значения являются средними ± SD ( n = 3, 4 или 5).
Рисунок 5
Изменения молекулярных видов…
Рисунок 5
Изменения молекулярных видов мембранных липидов у A. thaliana (А) и…
Рисунок 5
Изменения молекулярного состава мембранных липидов у A. thaliana (A) и C. himalaica (B) при изменении высоких и низких температур в ростовой камере. «R» и «F» означают, что температура повышалась или понижалась соответственно. Значения являются средними ± SD ( n = 4 или 5).
Значения являются средними ± SD ( n = 4 или 5).
Рисунок 5
Изменения молекулярных видов…
Рисунок 5
Изменения молекулярных видов мембранных липидов у A. thaliana (А) и…
Рисунок 5
Изменения молекулярных видов мембранных липидов в A. thaliana (A) и C. himalaica (B) при чередовании высоких и низких температур в ростовой камере. «R» и «F» означают, что температура повышалась или понижалась соответственно. Значения являются средними ± SD ( n = 4 или 5).
Рисунок 6
Изменения молекулярных видов…
Рисунок 6
Изменения молекулярных видов lysoPL у A. thaliana и C. himalaica в течение…
Рисунок 6
Изменения молекулярных видов lysoPL у A. thaliana и C. himalaica при изменении высоких и низких температур в ростовой камере. «R» и «F» означают, что температура повышалась или понижалась соответственно. Значения являются средними ± SD ( n = 4 или 5).
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Ремоделирование мембранных липидов Meconopsis racemosa после его интродукции в низменности из альпийской среды.
Чжэн Г., Тянь Б., Ли В.
Чжэн Г и др.
ПЛОС Один. 3 сентября 2014 г .; 9 (9): e106614. doi: 10.1371/journal.pone.0106614. Электронная коллекция 2014.
ПЛОС Один. 2014.PMID: 25184635
Бесплатная статья ЧВК.Дифференциальное ремоделирование липидома при холодовой акклиматизации у природных образцов Arabidopsis thaliana.
Дегенколбе Т., Джавалиско П., Цутер Э., Зайверт Б., Хинча Д.К., Уиллмитцер Л.
Дегенколбе Т. и соавт.
Плант Дж. 2012 г., декабрь 72(6):972-82. doi: 10.1111/tpj.12007. Epub 2012 19 октября.
Завод Дж. 2012.PMID: 23061922
Различия в липидном гомеостазе и ненасыщенности мембранных липидов придают разную толерантность к низким температурам у двух видов Cycas.
Чжэн И, Ян И, Ван М, Ху С, Ву Дж, Ю З.
Чжэн И и др.
BMC Растение Биол. 2021 16 августа; 21 (1): 377. doi: 10.1186/s12870-021-03158-4.
BMC Растение Биол. 2021.PMID: 34399687
Бесплатная статья ЧВК.Профилирование мембранных липидов в реакциях растений на стресс. Роль фосфолипазы D-альфа в изменениях липидов, вызванных замораживанием, у арабидопсиса.
Welti R, Li W, Li M, Sang Y, Biesiada H, Zhou HE, Rajashekar CB, Williams TD, Wang X.
Велти Р. и др.
Дж. Биол. Хим. 2002 г., 30 августа; 277(35):31994-2002. doi: 10.1074/jbc.M205375200. Epub 2002 19 июня.
Дж. Биол. Хим. 2002.PMID: 12077151
Температурная адаптация у двух видов двустворчатых моллюсков из разных термальных местообитаний: энергетика и ремоделирование мембранных липидов.
Перне Ф., Тремблей Р., Комо Л., Гудерлей Х.
Перне Ф. и др.
J Эксперт Биол. 2007 сен; 210 (часть 17): 2999-3014. doi: 10.1242/jeb.006007.
J Эксперт Биол. 2007.PMID: 17704075
Посмотреть все похожие статьи
Цитируется
Подход GWAS с использованием липидомики (lGWAS) для улучшения устойчивости сельскохозяйственных культур к высокотемпературному стрессу.
Пранешрадж В., Сангха М.К., Джалович И., Миладинович Дж., Джанагираман М.
Пранешрадж В. и др.
Int J Mol Sci. 2022 авг 20;23(16):9389. doi: 10.3390/ijms23169389.
Int J Mol Sci. 2022.PMID: 36012660
Бесплатная статья ЧВК.Обзор.
Анализ сети регуляции генов на основе транскриптома дифференциальной холодоустойчивости двух сортов табака.
Луо З., Чжоу З., Ли И., Тао С., Ху З. Р., Ян Дж. С., Ченг С., Ху Р., Чжан В.
Луо Зи и др.
BMC Растение Биол. 2022 26 июля; 22 (1): 369. doi: 10.1186/s12870-022-03767-7.
BMC Растение Биол. 2022.PMID: 35879667
Бесплатная статья ЧВК.Модуль CPK28-NLP7 объединяет индуцированный холодом сигнал Ca 2+ и перепрограммирование транскрипции в Arabidopsis .
Дин Ю, Ян Х, Ву С, Фу Д, Ли М, Гонг З, Ян С.
Дин Ю и др.
Научная реклама 2022 июль;8(26):eabn7901. doi: 10.1126/sciadv.abn7901. Epub 2022 29 июня.
Научная реклама 2022.PMID: 35767615
Бесплатная статья ЧВК.Повышенное накопление масла за счет совместной экспрессии транспортеров жирных кислот и ABC в Chlamydomonas при стандартных условиях роста.
Чен Р., Ян М., Ли М., Чжан Х., Лу Х., Доу Х., Фэн С., Сюэ С., Чжу С., Чи З., Конг Ф.
Чен Р. и др.
Биотехнология Биотопливо Биопрод. 2022 20 мая; 15 (1): 54. doi: 10.1186/s13068-022-02154-6.
Биотехнология Биотопливо Биопрод. 2022.PMID: 35596223
Бесплатная статья ЧВК.Липидомное ремоделирование в Begonia grandis при тепловом стрессе.
Sun AZ, Chen LS, Tang M, Chen JH, Li H, Jin XQ, Yi Y, Guo FQ.
Сан АЗ и др.
Фронт завод науч. 2022 17 фев; 13:843942. doi: 10.3389/fpls.2022.843942. Электронная коллекция 2022.
Фронт завод науч. 2022.PMID: 35251112
Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- P20 GM103418/GM/NIGMS NIH HHS/США
- P20 RR016475/RR/NCRR NIH HHS/США
- P20 RR16475/RR/NCRR NIH HHS/США
Как растения справляются с температурным стрессом | BMC Biology
- Комментарий
- Открытый доступ
- Опубликовано:
- Вирджиния Уолбот 1
BMC Биология
том 9 , номер статьи: 79 (2011)
Процитировать эту статью
22 тыс. обращений
14 цитирований
Сведения о показателях
Abstract
Холодная ночь может смениться жарким днем, и, поскольку они не могут двигаться, растения, подверженные таким колебаниям температуры, должны акклиматизироваться главным образом на основе ранее существовавших белков. Чжан и др. . Однако в статье BMC Plant Biology сообщается, что гибель клеток, вызванная нагреванием, является результатом транскрипционной активации киназы, связанной с факторами устойчивости к болезням и приводящей к локализованной гиперчувствительной реакции. Этот специализированный ответ отражает отказ адаптаций, которые обычно позволяют растениям выживать в значительном диапазоне температур, по механизмам, которые до конца не изучены.
См. научную статью http://www.biomedcentral.com/1471-2229/11/160
Комментарий
Когда вы разбиваете лагерь в высокогорных пустынях Южной Калифорнии, ночью замерзает. Пока вы съеживаетесь в своем спальном мешке, ожидая, пока закипит кофейная вода, встает солнце, и растения начинают фотосинтез, используя первые фотоны прохладного рассвета. Позже в тот же день, когда вы сбрасываете одежду и глотаете воду при 45-градусной жаре, те же растения проводят фотосинтез под палящим солнцем. Примечательно, что растения, адаптированные к высокогорным пустыням, хорошо себя чувствуют, несмотря на суточные перепады температуры примерно в 50°С [1]. Транскрипция, трансляция, мембранные свойства, митохондриальное дыхание, процессы, опосредованные микротрубочками и микрофиламентами, пластиды и все другие важные функции клетки сохраняют активность в широком диапазоне температур, и, кроме того, все эти процессы остаются в равновесии [2]. Как растительные клетки, ткани и органы могут поддерживать гомеостаз, несмотря на колебания температуры? Конечно, для растений и сельскохозяйственных культур умеренных и тропических зон колебания менее экстремальны в течение обычного дня, но, тем не менее, изменения на 10–15 °C в течение дня или недели легко компенсируются. Только при нарушении какого-либо процесса из-за жары или холода локальный температурный режим задает пределы распространения растений [3]. Чжан и др. [4] представляют собой пример неудачи с доказательством того, что превышение гомеостатических пределов для управления активными формами кислорода при высокой температуре приводит к локальной гибели клеток.
Эффективность в сравнении с приспособляемостью
На уровне отдельных белков должен быть компромисс между максимальной эффективностью при постоянной температуре (путем, которым пользуются млекопитающие) и достаточностью в широком диапазоне температур (рис. 1). Другими словами, у млекопитающих ферменты и составляющие процессы выходят из строя за пределами узкого диапазона оптимальных температур, и такой отказ является катастрофическим для организма, тогда как растения могут поддерживать и координировать клеточные процессы в широком диапазоне температур.
Рисунок 1
Схематическое изображение температурной устойчивости процессов растений и животных . Растительные белки (черная линия) демонстрируют широкий диапазон активности и стабильности по отношению к температуре по сравнению с белками млекопитающих (красная линия) с пиком, узким центром которого является температура тела.
Увеличенное изображение
Нарушение координации между ключевыми клеточными процессами может привести к гибели клеток. Это может происходить в растительных клетках как часть локализованной сверхчувствительной реакции на атаку патогена, стратегии ограничения роста захватчика. Подобные локализованные очаги гибели клеток могут возникать в результате нарушения гомеостатических процессов при непермиссивной температуре у мутантов растений со специфической чувствительностью к теплу или холоду. В обоих случаях это отражает неспособность управлять активным кислородом и его последствиями. В случае, о котором сообщил Zhang и др. [4] высокая температура недопустима (рис. 2). Напротив, доминантный мутант кукурузы Lesion mimic1 ( Les1 ) полностью нормален при высокой температуре, но при 22°C или ниже развиваются обширные некротические поражения листьев [5]. Ранение листьев булавками или окрашивание листьев низкомолекулярными органическими соединениями (такими, которые содержатся в маркерах, используемых для письма на листьях кукурузы) могут вызвать симптомы поражения при любой температуре. Наблюдения Zhang и др. . предполагают, что пути, активируемые при защите от патогенов, могут также играть роль в интеграции ответов на абиотические вызовы значительных колебаний температуры. Однако остается загадкой, как растениям вообще удается поддерживать гомеостаз перед лицом таких колебаний.
Рисунок 2
Некротические поражения на листьях, выращенных при недопустимой температуре . Листья риса со сверхэкспрессией NRKe , гена, кодирующего киназу, связанную с известными киназами устойчивости к болезням, которые вызывают некроз в ответ на инфекцию, выращивают в течение 10 дней при 24°C (две лопасти слева), а затем в течение 3 дней при 35°C (две лопасти справа). При 35°С, недопустимой температуре, на листьях появляются некротические поражения. Взято с рисунка 2b Чжан 9.0111 и др. [4].
Увеличить
Является ли решение дублированием колебаний температуры или стабилизацией белков?
В зависимости от вида растений аллели дуплицированных генов у тетраплоидных растений могут быть отобраны для участия в температурной буферизации; но только самый последний тетраплоид, возможно, сохраняет все дублированные гены, поэтому это не может объяснить адаптивность большинства растений. Если для температурной адаптации необходимы субфункциональные локусы, потеря генов и фиксация аллелей в отдельных локусах могут стать рецептом катастрофы в изменчивой среде, поскольку они приведут к многочисленным процессам, уязвимым к высоким и низким температурам. Вместо этого я предсказываю, что большая часть буферизации от температурных возмущений в белковых комплексах и в координации процессов потребует дополнительного, возможно, нового и специфичного для растений набора стабилизирующих белков.
Меня больше всего поразило сохранение точности в процессах на основе нуклеиновых кислот. Эмпирические лабораторные данные убедительно демонстрируют, что взаимодействия белков, такие как связывание факторов транскрипции с короткими мотивами ДНК или заряженными тРНК с тремя основаниями в мРНК, стабильны в очень узком диапазоне температур. In vivo структура хроматина может стабилизировать взаимодействие белок-ДНК, чтобы регулировать инициацию транскрипции, а рибосома аналогичным образом обеспечивает специальную нишу для трансляции. Я предполагаю, что у растений будут идентифицированы белки, которые стабилизируют специфические локальные конфигурации хроматина в пределах нормального температурного диапазона для данной среды обитания. Точно так же я предсказываю, что будут существовать белки, стабилизирующие рибосомы, чтобы позволить слабым взаимодействиям внутри молекулярных комплексов продолжаться с высокой точностью и эффективностью. Эти факторы стабильности будут предотвращать как диссоциацию факторов, которые действуют вместе, так и сохранение взаимодействий между белками, разборка которых необходима для нормальной регуляции клеточных процессов. Я предполагаю, что различные молекулярные комплексы, определенные у лабораторных дрожжей и у млекопитающих, будут иметь дополнительные белки. Я предсказываю, что они, скорее всего, будут определяться биохимическими, а не генетическими подходами, потому что в оптимальных условиях они могут быть ненужными.
Непрерывное развитие как стратегия акклиматизации органов
Стратегия непрерывного развития растений также способствует сохранению функций в изменчивой среде. Растения приспосабливаются к текущим условиям за счет интеграции информации об окружающей среде с программами развития для образования листьев и стеблей или корней из меристем и последующего роста и дифференциации. Пример акклиматизации к световым условиям используется питомниками, чтобы побудить вас купить растения. Изгоняя растения по фотонам в тусклом свете, садоводы выращивают растения с более крупными и темными зелеными листьями. После того, как вы купили растение и посадили его на солнечное окно, все роскошные теневые листья преждевременно стареют, и появляются более мелкие и бледные солнечные листья, которые больше подходят для яркого освещения. Зебра-полосатые мутанты кукурузы отражают чередование прохладных темных ночей с более теплыми яркими днями из-за неспособности стабилизировать содержание хлорофилла (рис. 3) в течение половины каждого 24-часового периода в ответ на неблагоприятные световые или температурные условия, в результате чего поперечные полосы бледной ткани чередуются с нормальной зеленой тканью в течение 10-дневного периода развития пластинок. У многих злаков дефектные полосы не восстанавливаются до фенотипа дикого типа, но остаются сообщением о колебаниях температуры за пределами оптимального диапазона в ходе роста листьев.
Рисунок 3
Полосатая полоса на полностью зрелом растении кукурузы, выращенном на Молокаи, Гавайи, зимой 2011 г. . Все взрослые листья были отмечены поперечными нормальными темно-зелеными и хлоротичными (желто-зелеными) полосами. Для развития взрослого листа требуется около 10 дней, и предполагается, что пары зеленых и желто-зеленых полос представляют 24-часовой период развития листа. Без дополнительных тестов невозможно точно определить, является ли нормальное развитие или стабильность хлоропластов чувствительными к теплу или холоду. Дневная температура окружающего воздуха составляла от 23 до 27°C, незначительно превышая ночную температуру от 17 до 21°C. Полосатая зебра очень устойчива на кукурузе, потому что зоны плохой дифференциации не восстанавливаются во время роста при допустимой температуре. Фотография предоставлена Тимом Келлихером, Стэнфордский университет.
Изображение полного размера
Ключевые этапы, когда температурная устойчивость действительно имеет значение
Прорастание цветковых растений — это необратимый рост зародыша растения из защитной семенной оболочки, подпитываемый запасами питательных веществ. По мере прорастания происходит гонка между скоростью потребления запасов и созданием независимого, фотосинтетически компетентного сеянца, способного получать воду и минеральные питательные вещества из почвы. У многих видов прорастание очень чувствительно к температуре. Во-первых, многие виды запускаются для прорастания периодом высокой или низкой температуры, который разрушает ингибиторы прорастания, адаптация, позволяющая растению измерять конец зимы для весеннего появления всходов или конец лета для осеннего прорастания. Во-вторых, вода стимулирует впитывание, делая возможным рост, но последующее понижение температуры может заморозить нежный стебель проростка, в то время как высокая температура приведет к хрусту распустившихся предварительно сформированных листьев без возможности восстановления в условиях, которые могут переноситься хорошо укоренившимся растением.
Цветение представляет собой еще одно одностороннее обязательство в жизненном цикле, так как апикальная меристема, ранее производившая листья и стебли, переключается на цветочную программу и полностью расходуется на формирование цветка. Хотя жара и холод могут неблагоприятно воздействовать на эффектные части цветка, наиболее серьезное воздействие оказывается в первую очередь на развивающуюся гаплоидную пыльцу и ее питательную диплоидную поддерживающую ткань, тапетум. Параллель с прорастанием заключается в том, что пыльца отделяется от вегетативного растения вскоре после мейоза толстым слоем и должна сохраняться с фиксированным запасом питательных веществ на протяжении всего созревания, распространения и начальных стадий роста пыльцевых трубок до оплодотворения. Питательные вещества в пыльце проходят через тапетальный слой, поэтому качество этого одноклеточного тканевого кольца также имеет первостепенное значение. У томата небольшое повышение температуры, которое не влияло на биомассу растения, количество цветков или мейоз, сильно влияло на количество функциональных пыльцевых зерен и, следовательно, на урожай плодов [6]. У риса низкая температура ограничивает производство в прохладный сезон из-за негативного влияния на репродуктивную способность самцов [7]. Литература по мужскому фитнесу изобилует примерами негативного влияния переносимых вегетативным растением перепадов температур.
Принимая во внимание как вегетативные случаи (фенотип листьев), так и мужскую стерильность, становится ясно, что температуры сразу за пределами диапазона акклиматизации могут сильно повлиять как на выживание, так и на размножение. Эти случаи показывают, что растения могут процветать в широком диапазоне температур, но температуры за пределами генотипических порогов вызывают такие последствия, как гибель клеток, как показано Zhang et al . — плохое озеленение и мужское бесплодие. Эти вредные фенотипы являются отправной точкой для разгадки механизмов, лежащих в основе устойчивости к температуре, с гипотезой о том, что первый процесс, который нарушается при высокой или низкой температуре, определяет ключевой компонент жизни растений.
Ссылки
Смит С.Д., Монсон Р.
К., Андерсон Дж.Э. Физиологическая экология растений пустыни Северной Америки. 1997, Нью-Йорк: Книга SpringerGoogle ученый
Пошел FW: Влияние температуры на рост растений. Annu Rev Plant Physiol. 1953, 4: 347-362. 10.1146/аннурев.стр.04.060153.002023.
Артикул
Google ученый
Osmond CB, Austin MP, Berry JA, Billings WD, Boyer JS, Dacey JWH, Nobel PS, Smith SD, Winner WE: Физиология стресса и распространение растений. Биология. 1987, 37: 38-48. 10.2307/1310176.
Артикул
Google ученый
Zhang H, Cao Y, Zhao J, Li H, Xiao J, Wang S: пара ортологов богатого лейцином семейства генов устойчивости к киназному рецептору повторов регулирует реакцию риса на повышенную температуру. BMC Растение Биол. 2011, 11: 160-10.1186/1471-2229-11-160.
Артикул
ПабМед Центральный
КАС
пабмедGoogle ученый
Сато С., Камияма М., Ивата Т., Макита Н., Фурукава Х., Икеда Х.: Умеренное повышение среднесуточной температуры неблагоприятно влияет на завязывание плодов Lycopersicon esculentum , нарушая определенные физиологические процессы в репродуктивном развитии самцов. Энн Бот. 2006, 97: 731-738. 10.1093/аоб/mcl037.
Артикул
ПабМед Центральный
КАС
пабмедGoogle ученый
Симоно Х., Хасегава Т., Морияма М., Фумимура С., Нагата Т.: Моделирование стерильности колосков, вызванной низкой температурой в рисе. Агрономия Ж.
2005, 97: 1524-1536. 10.2134/агронь2005.0043.Артикул
Google ученый
Хойзингтон Д.А., Нойффер М.Г., Уолбот В.: Имитация поражений кукурузы. I. Влияние генетического фона, температуры, возраста развития и ранения на образование пятен некроза у Les1 . Дев биол. 1982, 93: 381-388. 10.1016/0012-1606(82)-7.
Артикул
КАС
пабмед
Google ученый
Ссылки на скачивание
Благодарности
Поддерживается грантом Национального научного фонда (PGRP 07-01880).
Информация об авторе
Авторы и организации
Факультет биологии MC5020, 385 Serra Mall, Stanford University, Stanford, CA, 94305-5020, USA
Virginia Walbot
Авторы
- Virginia Walbot
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar
Автор, ответственный за корреспонденцию
Вирджиния Уолбот.
Оригинальные файлы изображений, представленные авторами
Ниже приведены ссылки на оригинальные файлы изображений, представленные авторами.
Оригинальный файл авторов рисунка 1
Оригинальный файл авторов для рисунка 2
Оригинальный файл авторов для рисунка 3
Права и разрешения
Эта статья опубликована по лицензии BioMed Central Ltd.