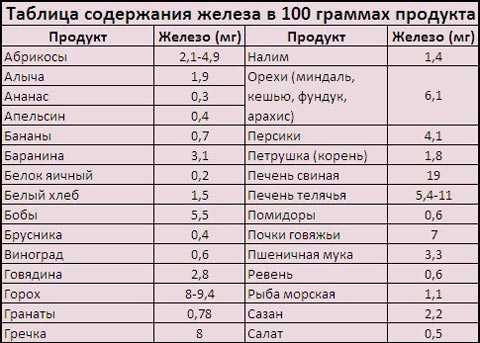
10 самых лучших продуктов богатых железом которые могут предотвратить Анемию. Железо составляет от сухой массы растения
4.3.9. Микроэлементы
Железо.Среднее содержание железа в растениях составляет 20-80 мг на 1 кг сухой массы. Ионы Fe3+почвенного раствора восстанавливаются редокс-системами плазмалеммы клеток ризодермы до Fe2+и в такой форме поступают в корень.
Железо необходимо для функционирования основных редокс-систем фотосинтеза и дыхания, синтеза хлорофилла, восстановления нитратов и фиксации молекулярного азота клубеньковыми бактериями, входя в состав нитратредуктазы и нитрогеназы. Поэтому недостаточное поступление железа в растения в условиях переувлажнения и на карбонатных почвах приводит к снижению интенсивности дыхания и фотосинтеза и выражается в пожелтении (хлорозе) листьев и быстром их опадении.
Марганецв клетки поступает в форме ионов Mn2+. Среднее его содержание составляет 1 мг на 1 кг сухой массы. Марганец накапливается в листьях. Он необходим для фоторазложения воды с выделением кислорода и восстановления углекислого газа при фотосинтезе. Марганец способствует увеличению содержания сахаров и их оттоку из листьев. Два фермента цикла Кребса - малат- и изоцитратдегидрогеназы - активируются ионами марганца. Он также необходим для функционирования нитратредуктазы при восстановлении нитратов. Марганец является кофактором РНКполимеразы и ауксиноксидазы, разрушающей фитогормон 3-индолилуксусную кислоту.
Характерный симптом марганцевого голодания - точечный хлороз листьев, когда между жилками появляются желтые пятна, а затем клетки в этих участках отмирают.
Молибден.Наибольшее содержание молибдена характерно для бобовых (0,5-20 мг на 1 кг сухой массы), злаки содержат от 0,2 до 2 мг на кг сухой массы. Он поступает в растения в форме аниона МоО2-4, концентрируется в молодых, растущих органах. Его больше в листьях, чем в корнях и стеблях, а в листе сосредоточен, в основном, в хлоропластах.
Молибден входит в состав нитратредуктазы и нитрогеназы, а также необходим для биосинтеза легоглобина. Как металл-активатор молибден участвует в реакциях аминирования и переаминирования, для включения аминокислот в пептидную цепь, работы таких ферментов как ксантиноксидаза и различных фосфатаз.
При недостатке молибдена в тканях накапливается большое количество нитратов, не развиваются клубеньки на корнях бобовых, тормозится рост растений, наблюдаются деформации листовых пластинок. При высоких дозах молибден токсичен. При недостатке молибдена молодые листья по краям приобретают серую, а затем коричневую окраску, теряют тургор, а затем ткани листа отмирают и остаются только жилки в виде хлыстиков.
Кобальт. Среднее содержание кобальта в растениях 0,02 мг на 1 кг сухой массы. Кобальт необходим бобовым растениям для обеспечения размножения клубеньковых бактерий. В растениях кобальт встречается в ионной форме и в витамине В12. Растения не вырабатывают этот витамин. Он синтезируется бактероидами клубеньков бобовых растений и участвует в синтезе метионина в бактероидах. При старении клубеньков и прекращении фиксации азота витамин выходит в цитоплазму клеток клубеньков. Наряду с магнием и марганцем кобальт активирует фермент гликолиза фосфоглюкомутазу и фермент аргиназу, гидролизующий аргинин.
Внешние признаки недостатка кобальта сходны с признаками азотного голодания.
Медь поступает в клетки в форме иона Сu2+. Среднее содержание меди в растениях 0,2 мг на кг сухой массы. Около 70 % всей меди, находящейся в листьях, сосредоточены в хлоропластах и почти половина ее в составе пластоцианина - переносчика электронов между фотосистемами II и I. Она входит в состав ферментов, катализирующих окисление аскорбиновой кислоты, дифенолов и гидроксилирование монофенолов - аскорбатоксидазы, полифенолоксидазы, ортодифенолоксидазы и тирозиназы. Два атома меди функционируют в цитохромоксидазном комплексе дыхательной цепи митохондрий. Медь входит в состав нитратредуктазного комплекса и влияет на синтез легоглобина. Для биосинтеза этилена также необходим медьсодержащий фермент. Влияя на содержание в растениях ингибиторов роста фенольной природы медь повышает устойчивость растений к полеганию. Она также повышает засухо-, морозо- и жароустойчивость. Недостаток меди вызывает задержку роста и цветения, хлороз, потерю тургора и завядание растений. У злаков при недостатке меди не развивается колос, у плодовых появляется суховершинность. При дефиците меди белеют и отмирают кончики листьев, листья и плоды плодовых деревьев покрываются бурыми пятнами.
Цинк. Содержание цинка в надземных частях бобовых и злаковых растений составляет 15-60 мг на кг сухой массы. Повышенная концентрация отмечается в листьях, репродуктивных органах и конусах нарастания, наибольшая - в семенах.
Цинк поступает в растение в форме катиона Zn2+. Он необходим для функционирования ряда ферментов гликолиза - гексокиназы, енолазы, триозофосфатдегидрогеназы, альдолазы, а также входит в состав алкогольдегидрогеназы. Цинк активирует карбоангидразу, катализирующую реакцию дегидратации гидрата оксида углерода: Н2СО3СО2+ Н2О, что помогает использованию углекислого газа в процессе фотосинтеза. Цинк участвует в образовании аминокислоты триптофана. Именно с этим связано влияние катионов цинка на синтез белков, а также фитогормона 3-индолилуксусной кислоты, предшественником которой является триптофан. Подкормка цинком способствует увеличению содержания ауксинов в тканях и активирует их рост.
При дефиците цинка у растений нарушается фосфорный обмен: фосфор накапливается в корнях, задерживается его транспорт в надземные органы, замедляется превращение фосфора в органические формы. При недостатке цинка в растениях уменьшается содержание сахарозы и крахмала, увеличивается количество органических кислот и небелковых соединений азота - амидов и аминокислот. Кроме того, в 2-3 раза подавляется скорость деления клеток, что приводит к морфологическим изменениям листьев, нарушению растяжения клеток и дифференциации тканей. Наиболее характерный признак цинкового голодания - это задержка роста междоузлий и листьев, появление хлороза и развитие розеточности.
Бор.Его среднее содержание составляет 0,1 мг на кг сухой массы. В боре наиболее нуждаются двудольные растения. Много бора в цветках. В клетках большая часть бора сосредоточена в клеточных стенках. Бор усиливает рост пыльцевых трубок, прорастание пыльцы, увеличивает количество цветков и плодов. Без него нарушается созревание семян. Бор снижает активность некоторых дыхательных ферментов, оказывает влияние на углеводный, белковый и нуклеиновый обмен. При недостатке бора нарушаются синтез, превращения и транспорт углеводов, формирование репродуктивных органов, оплодотворение и плодоношение. Он не может реутилизироваться и поэтому при борном голодании прежде всего отмирают конусы нарастания, останавливается рост побегов и корней, листовые пластинки утолщаются, скручиваются, становятся ломкими, цветки не образуются.
studfiles.net
10 самых лучших продуктов богатых железом которые могут предотвратить Анемию

Железо — это незаменимый микроэлемент, который участвует в переносе кислорода ко всем частям тела. Даже небольшой дефицит железа в организме приводит к анемии, вызывая быструю утомляемость, недомогание и слабость, а хронический дефицит железа может привести к полиорганной недостаточности.
Риск дефицита железа является наибольшим:
- Женщины с менструальным циклом — из — за потери крови во время менструации, женщины детородного возраста подвергаются постоянному риску дефицита железа.
- Люди с почечной недостаточностью — особенно подвергаются риску анемии из — за неспособности почек вырабатывать достаточное количество гормона эритропоэтина, который необходим для производства красных кровяных клеток.
- Беременные и кормящие женщины — развивающемуся плоду требуется большое количество железа.
- Младенцы и дети до 3-х лет.
- Люди с низким уровнем витамина А — витамин А (ретинол и каротиноиды) активно участвует в транспортировке железа из «складов» в организме. Без необходимых количеств витамина А организм не может регулировать нормальный уровень железа, что приводит к дефициту.
- Люди с расстройствами пищеварительного тракта — диарея, язвы желудка и другие расстройств, могут привести к неадекватному поглощению и усвоению железа.
Избыток железа может привести к увеличению производства вредных свободных радикалов, которые мешают нормальному обмену веществ и могут привести к повреждению внутренних органов, таких как сердце и печень.
Обычно передозировка железа встречается редко и может возникнуть только в случае передозировки железосодержащих препаратов. Железо из натуральных, источников питания, которые перечислены ниже считается безопасным и здоровым.

Поглощение железа из пищи стимулируется дополнительным количеством витамина С и уменьшается при приеме больших количеств кальция, цинка или магния. Нормальному всасыванию железа мешают и продукты с высоким содержанием фитиновой кислоты и ее солей — кунжут, бразильский орех, миндаль, тофу, льняное семя, овес, бобы, сою (соевый белок), кукуруза, арахис, цельная пшеница, коричневый и белый рис, горох и т.д., а также продукты содержащие танины — черный чай и красное вино, гранат, черника и клубника, сырые орехи, такие как фундук, грецкий орех и некоторые специй — гвоздика, эстрагон, тмин, тимьян, ваниль и корица.
Рекомендуемая дневная доза железа составляет 18 мг. ежедневно.А так же рекомендуем статью : Продукты снижающие холестерин и очищающие сосуды

ТОП — 10 продуктов,богатых железом
- Сушеные травы и специи.Железо из натуральных, источников питания,
Травы и специи используются на протяжении многих веков в народной медицине так как являются сокровищницей активных ингредиентов, в том числе железа. Лидером среди них является сушеный тимьян (чабрец), который содержит 125 мг железа в 100 г сухого веса или 687% от рекомендуемой суточной нормы. Это означает, что с 1 ст. л. тимьяна обеспечит 3,8 мг железо или 21% от дневной нормы.
Перец, орегано, тмин, сушеный укроп, лавровый лист, сухой базилик и кориандр, куркума, семена аниса и пажитника, эстрагон и розмарин содержат 5% от РСН железа в каждой столовой ложке.
- Какао и шоколад
Чистый какао содержит 35 мг железа на 100 г., или 200% от РСН. Это означает, что 1 ст. л. какао содержит 1,7мг железа или 10% от РСН. Сладкий молочный шоколад содержит по меньшей мере — 2,4 мг железа на 100 г. (13% РСП).

- Печень
Печень богата витаминами и железом. В начале двадцатого века ее регулярно прописывали как средство от анемии и для беременных женщин. Свиная печень имеет 17,9 мг (100% РСП) в 100 г, куриная печень (72% РСП в 100 г), баранья печень (57%) и телячья печень (36%).
- Моллюски и устрицы
Моллюски можно потреблять в пищу в сыром виде, запеченные, вареные, жареные или в супе. Устрицы содержат 12 мг железа (67% РСП) в 100 г. Или 5 мг (28% РСП) в 6 средних устриц. Большие и удлиненные голубые морские моллюски (мидии) содержат 6,72 мг. (37% РСП) железа в части в 100 грамм.
- Сушеные и жареные семена тыквы
Популярная еда на Ближнем Востоке и в Восточной Азии содержит около 15 мг железа (84% РСП) в 100 г или 4,4 мг. (23% РСП) на порцию 30 г. Для той же цели можно потреблять в пищу и сушеные семена тыквы, которые содержат больше железа, чем жареные.
- Семена кунжута и масло из него
Семена кунжута являются распространенным ингредиентом в популярной средиземноморской кухне. Жареные семена кунжута содержат 14,8 мг (82% РСП) железа на 100 г или 4,1 мг. (23% РСП) в рекомендованной порции 30 г.

- Икра рыб
100 г икры содержит 11,9 мг (66% РСП) железа.
- Чиа семена
Они содержат много полезные веществ, включая железо. Около 30% суточного потребления железа может обеспечить потребление всего двух ст. л. ч чиа. Чиа очень обильный источник омега-3 — богаче, чем рыба и другие зерновые. Семя также содержит много белка, целлюлозы, витаминов и минералов. Семена можно употреблять в пищу сырые, и добавить их в йогурт, мюсли, соус, суп, каша, рис, хлеб и десерты.
- Шпинат
Шпинат является одним из лучших источников железа. Кроме того, он содержит белок и богат магнием, фолиевой кислотой, витамин А, витамина С и хлорофиллом. Витамин С повышает всасывание железа.
- Курага
Чаще всего используется для перекуса между приемами пищи, но могут быть отличным дополнением к салатам. Курага — это хороший источник витамина А и калия, а также 6,3 мг (35% РСП) железа на 100 г.
kaksov.ru
Химический состав растений | Фитоблог
Терапевтическая ценность лекарственных растений определяется входящими в их состав биологически активными веществами - химический состав растений, к которым относятся все вещества, способные оказывать влияние на биологические процессы, протекающие в организме.
За долгую историю поисков и практического использования биологически активных веществ накопились сведения о биологической активности большого числа химических соединений растений с полностью или частично установленной структурой. Только фармакологическая активность, если судить по различным справочникам и фармакопеям, описана примерно для 12 000 различных соединений.
Для части из них известна также и физиологическая система организма или орган - мишень действия. Значительно меньше известны те биохимические или молекулярно-биологические процессы, на которые действуют эти вещества.
Любое из лекарственных растений представляет собой сложную лабораторию с различным химическим составом, в которой синтезируются одновременно сотни, если не тысячи, биологически активных веществ. Этим и объясняется так называемый шрапнельный эффект, т.е. эффект множественного воздействия на различные системы и органы, нередко возникающий в процессе лечения. Дополнительные исследования, казалось бы, вполне изученных и давно использующихся лекарственных растений иногда позволяют выявить новый аспект их биологической активности.
В связи с разносторонним лечебным эффектом лекарственных растений в известной степени условным оказывается понятие так называемых действующих веществ, обозначающее наиболее выраженную физиологическую активность. Вещества кажущиеся неактивными условно делятся на сопутствующие и балластные,
Устаревшими оказываются понятия "сопутствующие" и "балластные" вещества.
Сопутствующими веществами в фармакогнозии ранее называли продукты первичного или вторичного обмена (метаболизма), содержащиеся в лекарственных растениях наряду с действующими веществами. Их фармакологический эффект значительно менее выражен, но их присутствие нередко способствует пролонгированию лечебного эффекта, усиливает и ускоряет его наступление. Однако сопутствующие вещества могут проявлять и отрицательные свойства, поэтому приходится освобождаться от них в ходе приготовления из растительного сырья лекарственных средств и форм.
Достаточно близко понятию "сопутствующие" вещества понятие "балластные" вещества, встречающееся в старых руководствах по фармакогнозии.
Балластными веществами называли соединения, с которыми не связана терапевтическая активность того или иного лекарственного растения. Однако нередко они затрудняют изготовление или поддержание стабильности лекарственных форм.
Резкой границы между приведенными группами нет, и деление их условно, поскольку одну и ту же группу веществ мы иной раз относим к действующим, а другой раз – к балластным.
По мере развития знаний о лекарственных растениях вещества из группы кажущихся неактивных переводят в группу действующих веществ.
Растения способны синтезировать из неорганических веществ органические, необходимые для жизнедеятельности человека и животных. Химический состав растений довольно сложен: помимо воды (70-90 %), растения состоят из неорганических и органических веществ. На долю неорганических (минеральных) веществ приходится от 3 до 25% массы сухого остатка растений.
Сумма минеральных веществ (зола) остается после сжигания органической части растений. Растения содержат практически все природные элементы, причем концентрация их в растениях близка к содержанию их в почве.
Каждый минеральный элемент играет определенную роль в обмене веществ и не может быть заменен другим элементом. Минеральные элементы влияют практически на все физиологические процессы, происходящие в растениях: дыхание, рост, развитие, фотосинтез.
Неорганические вещества часто содержатся в растениях в виде комплексов с органическими соединениями. Например, кремниевая кислота в траве хвоща полевого находится в связанной с органическими соединениями растворимой форме. Слоевища ламинарии содержат органически связанный йод.
Некоторые минеральные вещества непосредственно включены в структуру органических соединений (например, магний входит в состав хлорофилла).
По количественному содержанию в растениях различают макро- и микроэлементы.
К макроэлементам относятся металлы - калий (К), кальций (Са), магний (Мg), натрий (Nа), и неметаллы - кремний (Si), сера (S), фосфор (Р), хлор (Сl). Содержание их не менее 0,01%.
Некоторые из макроэлементов участвуют в фармакологическом эффекте лекарственного растительного сырья. Например, кремнеорганические соединения хвоща полевого и горца птичьего в почках и мочевыводящих путях боль-ного образуют защитные коллоиды, которые препятствуют кристаллизации некоторых минеральных компонентов, затрудняют образование мочевых камней.
К микроэлементам относятся железо (Fе), медь (Сu), марганец (Мn), кобальт (Со), цинк (Zn), алюминий (А1), молибден (Мо), хром (Сг), золото (Аu), ртуть (Нg), свинец (РЬ), серебро (Аu), йод (J), бор (В) и др. Содержание их незначительное и обычно не превышает 0,001%.
Отдельные микроэлементы также определяют фармакологическую ак-
тивность лекарственного растительного сырья.
Например:
- слоевища ламинарии (бурые водоросли накапливают йод) используют при лечении больных с заболеваниями щитовидной железы;
- сфагнум (он концентрирует Аg) применяют для лечения ран;
- сырье крапивы, тысячелистника, зайцегуба опьяняющего, богатое Са и Mg используют при лечении больных с внутренними кровотечениями;
- побеги черники, богатые Мn и А1, применяют при лечении больных сахарным диабетом.
Сумма неорганических веществ растений как лекарственное средство в научной медицине не используется, но применяется в народной медицине.
Например, в народной медицине Бурятии для лечения ран, ожогов, трофических язв применяют золу сушеницы топяной.
Основную массу сухого остатка растений составляют органические вещества. Среди них различают вещества первичного синтеза и вещества вторичного синтеза.
При применении растений в качестве лекарственных средств на организм человека действует сложный комплекс минеральных веществ и органических соединений первичного и вторичного синтеза.
В этом комплексе различают биологически активные вещества и вещества, кажущиеся неактивными.
[ad name="ФИТО 336"]
Фармакогнозия на сайте Фитоблог
Интересные статьи:
Другие интересные статьи на сайте:
phytoblog.ru
В. И. Малиновский, "ФИЗИОЛОГИЯ РАСТЕНИЙ"
4. МИНЕРАЛЬНОЕ ПИТАНИЕ
4.1. Почва как источник питательных веществ
Растения получают углерод и кислород преимущественно из воздуха, а остальные элементы из почвы. Питательные элементы - это химические элементы, которые необходимы растению и не могут быть заменены никакими другими. Питательные вещества - это соединения, в которых имеются эти элементы. Питательные элементы содержатся в почве в 4 формах: 1) прочно фиксированные и недоступные для растения (например, ионы калия и аммония в некоторых глинистых минералах, 2) труднорастворимые неорганические соли (сульфаты, фосфаты, карбонаты) и в такой форме недоступные для растения, 3) адсорбированные на поверхности коллоидов, доступные для растений благодаря ионному обмену на выделяемые растением ионы, 4) растворенные в воде и поэтому легко доступные для растений. Ионы поступают в клетки ризодермы из почвенного раствора и благодаря контактному обмену Н+, НСО-3 и анионов органических кислот на ионы минеральных веществ почвенных частиц. Контактный обмен ионов клеточной стенки ризодермы с частицами почвы осуществляется без перехода ионов в почвенный раствор. Тесный контакт обеспечивается благодаря выделению слизи корневыми волосками и отсутствию у ризодермы кутикулы и других защитных покровов. Так как адсорбированные ионы находятся в постоянном колебательном движении и занимают определенный объем - сферу колебаний, при тесном контакте поверхностей сферы колебаний двух ближайших адсорбированных ионов могут перекрываться, в результате чего осуществляется ионный обмен. Выделяя различные вещества (углекислый газ, аминокислоты, сахара и другие), корень растения изменяет состояние питательных веществ в прикорневой зоне непосредственно, например, путем выделения СО2 (СО2 + Н2О ® Н+ + НСО-3: повышение растворимости фосфатов и карбонатов) и косвенно, создавая благоприятные условия для ризосферы, которая играет большую роль в превращении почвенных минералов.4.2. Содержание минеральных элементов в растениях
Растения способны поглощать из окружающей среды практически все элементы. Однако для нормальной жизнедеятельности растительному организму необходимо лишь 19 питательных элементов. Среди них углерод (45 % сухой массы тканей), кислород (42%), водород (6,5%) и азот (1,5%) называют органогенами. Оставшиеся 5 % приходятся на зольные элементы, которые остаются в золе после сжигания растения. Содержание минеральных элементов обычно выражают в процентах от массы сухого вещества. Все элементы в зависимости от их количественного содержания в растении принято делить на макроэлементы (содержание более 0,01 %) - к ним относятся азот, фосфор, сера, калий, кальций, магний и микроэлементы (содержание менее 0,01 %): железо, марганец, медь, цинк, бор, молибден, кобальт, хлор. Ю. Либихом было установлено, что все перечисленные элементы равнозначны и полное исключение любого из них приводит растение к глубокому страданию и гибели, ни один из перечисленных элементов не может быть заменен другим, даже близким по химическим свойствам. Макроэлементы при концентрации 200-300 мг/л в питательном растворе еще не оказывают вредного действия на растение. Большинство микроэлементов при концентрации 0,1-0,5 мг/л угнетают рост растений. Для нормальной жизнедеятельности растений должно быть определенное соотношение различных ионов в окружающей среде. Чистые растворы одного какого-либо катиона оказываются ядовитыми. Так, при помещении проростков пшеницы на чистые растворы KCL или CaCL2 на корнях сначала появлялись вздутия, а затем корни отмирали. Смешанные растворы этих солей не обладали ядовитым действием. Смягчающее влияние одного катиона на действие другого катиона называют антагонизмом ионов. Антагонизм ионов проявляется как между разными ионами одной валентности, например, между ионами натрия и калия, так и между ионами разной валентности, например, калия и кальция. Одной из причин антагонизма ионов является их влияние на гидратацию белков цитоплазмы. Двухвалентные катионы (кальций, магний) дегидратируют коллоиды сильнее, чем одновалентные (натрий, калий). Следующей причиной антагонизма ионов является их конкуренция за активные центры ферментов. Так, активность некоторых ферментов дыхания ингибируется ионами натрия, но их действие снимается добавлением ионов калия. Кроме того, ионы могут конкурировать за связывание с переносчиками в процессе поглощения. Действие одного иона может и усиливать влияние другого иона. Это явление называется синергизмом. Так, под влиянием фосфора повышается положительное действие молибдена.4.3. Физиолого-биохимическая роль основных элементов питания
4.3.1. Углерод
Все органические соединения построены, в основном, из углерода. Растение получает углерод из воздуха, поглощая углекислый газ, но 2 – 5 % углерода усваивается корнями в виде углекислоты из почвы. Растения поглощают углекислый газ и в процессе фотосинтеза синтезируют органическое вещество. В ходе диссимиляции, то есть расщепления органических веществ с использованием заключенной в них энергии, растения потребляют кислород и выделяют углекислый газ. Таким образом, растения участвуют в круговороте углерода на нашей планете. Основная масса (примерно 57 %) углекислоты атмосферы имеет растительное происхождение. Почва в результате жизнедеятельности почвенных микроорганизмов поставляет около 58 млрд. т углекислоты в год, то есть 38 %. Промышленная деятельность человечества (сжигание угля, нефти и другие) занимает 3 % в балансе выделяемой углекислоты. Остальные источники - дыхание людей и животных, вулканы, фумаролы и другие - вместе выделяют менее 2 % углекислоты. Мировой океан принимает участие в регуляции содержания углекислого газа в атмосфере. Морская и пресная вода, кроме карбонатов и оснований, содержит также растворенную углекислоту и бикарбонаты. При изменении давления СО2 в воздухе часть его для достижения динамического равновесия между свободным газом атмосферы и растворенным в воде переходит в воду или обратно в атмосферу. Однако постоянство парциального давления углекислого газа в атмосфере достигается, главным образом, соответствием между выделением углекислоты и потреблением ее растениями. Ежегодно в процессе фотосинтеза наземные и морские растения поглощают около 15,6 х 1010 т углекислоты, то есть 1/16 всего мирового запаса.4.3.2. Азот
Азот входит в состав белков, нуклеиновых кислот, пигментов, коферментов, фитогормонов и витаминов. При недостатке азота тормозится рост растений, ослабляется образование боковых побегов и кущение у злаков, наблюдается мелколистность, уменьшается ветвление корней. Симптомом азотного дефицита является хлороз листьев - бледно-зеленая окраска листьев, вызванная ослаблением синтеза пигмента хлорофилла. Длительное азотное голодание ведет к гидролизу белков и разрушению хлорофилла в нижних более старых листьях и оттоку растворимых соединений азота к молодым листьям, точкам роста и генеративным органам. Вследствие разрушения хлорофилла окраска нижних листьев в зависимости от вида растения приобретает желтые, оранжевые или красные тона, а при сильно выраженном азотном дефиците возможно высыхание и отмирание тканей.(…)4.3.3. Фосфор
Растения поглощают из почвы свободную ортофосфорную кислоту и ее двух- и однозамещенные соли, растворимые в воде, а также и некоторые органические соединения фосфора, такие как фосфаты сахаров и фитин. Содержание фосфора в растениях составляет около 0,2 % на сухую массу. Фосфор входит в состав нуклеиновых кислот, нуклеотидов, фосфолипидов и витаминов. Многие фосфорсодержащие витамины и их прозводные являются коферментами. (…) Основной запасной формой фосфора у растений является фитин - кальций-магниевая соль инозитфосфорной кислоты. Содержание фитина в семенах достигает 2 % от сухой массы, что составляет 50 % от общего содержания фосфора. При дефиците фосфора снижается скорость поглощения кислорода, снижается активность дыхательных ферментов, локализованных в митохондриях, и активируются ферменты (оксидаза гликолевой кислоты, аскорбатоксидаза) немитохондриальных систем окисления, происходит распад фосфорорганических соединений, тормозится синтез белков и свободных нуклеотидов. Наиболее чувствительны к недостатку фосфора молодые растения. Симптомом фосфорного голодания является синевато-зеленая окраска, в первую очередь, старых листьев нередко с пурпурным из-за накопления антоцианов или бронзовым оттенком (свидетельство задержки синтеза белка и накопления сахаров). Листья становятся мелкими и более узкими. Приостанавливается рост растений, задерживается созревание урожая.4.3.4. Сера
В почве сера находится в органической и неорганической формах. Органическая сера входит в состав растительных и животных остатков. Основные неорганические соединения серы в почве – сульфаты (CaSO4, MgSO4, Na2SO4). В затопляемых почвах сера находится в восстановленной форме в виде FeS, FeS2 или h3S. Растения поглощают из почвы сульфаты и в очень незначительных количествах серосодержащие аминокислоты. Содержание серы в растениях составляет около 0,2 %. Однако в растениях семейства крестоцветных ее содержание значительно выше. Сера содержится в растениях в двух основных формах - окисленной в виде неорганического сульфата и восстановленной (аминокислоты, глутатион, белки). Процесс восстановления сульфата происходит в хлоропластах. (…)Сера входит в состав многих витаминов и коферментов, таких как биотин, коэнзим А, глутатион, липоевая кислота. В связи с этим сера необходима для многих процессов обмена веществ (например, аэробная фаза дыхания, синтез жиров и так далее). Сера участвует в образовании полиаминов, которые влияют на структуру нуклеиновых кислот и рибосом, регулируют процессы деления клеток. Недостаточное снабжение растений серой тормозит синтез серосодержащих аминокислот и белков, снижает фотосинтез и скорость роста растений, приводит к разрушению хлоропластов. Симптомы дефицита серы - побледнение и пожелтение молодых, а затем и старых листьев.4.3.5. Калий
Калий поглощается растениями в виде катиона. Его содержание в растениях составляет, в среднем, 0,9 %. Концентрация калия высока в огурцах, томатах и капусте, но особенно много его в подсолнечнике. В растениях калий больше сосредоточен в молодых растущих тканях. Около 80 % калия содержится в вакуолях и 1 % калия прочно связан с белками митохондрий и хлоропластов. Калий стабилизирует структуру этих органелл. Калий участвует в создании разности электрических потенциалов между клетками. Он нейтрализует отрицательные заряды неорганических и органических анионов. Калий в значительной мере определяет коллоидные свойства цитоплазмы, так как способствует поддержанию состояния гидратации коллоидов цитоплазмы, повышая ее водоудерживающую способность. Тем самым калий увеличивает устойчивость растений к засухе и морозам. Калий необходим для работы устьичного аппарата. Известно более 60 ферментов, активируемых калием. (…) При недостатке калия он может заменяться натрием, но некоторые активируемые калием ферменты ингибируются натрием. При недостатке калия листья желтеют снизу вверх - от старых к молодым. Их края и верхушки приобретают бурую окраску, иногда с красными пятнами, затем происходит отмирание этих участков. Снижается функционирование камбия, нарушается развитие сосудистых тканей, уменьшается толщина кутикулы и стенок эпидермальных клеток, тормозятся процессы деления и растяжения клеток, что приводит к появлению розеточных форм растений. Недостаток калия вызывает остановку развития и гибель верхушечных почек, в результате чего активируется рост боковых побегов и растение принимает форму куста.4.3.6. Кальций
В почве содержится много кальция и кальциевое голодание встречается редко, например, при сильной кислотности или засоленности почв и на торфяниках. Общее содержание кальция у разных видов растений составляет 5-30 мг на 1 г сухой массы. Много кальция содержат бобовые, гречиха, подсолнечник, картофель, капуста, гораздо меньше - зерновые, лен, сахарная свекла. В тканях двудольных растений кальция больше, чем у однодольных. Кальций накапливается в старых органах и тканях. Это связано с тем, что реутилизация кальция затруднена, так как он из цитоплазмы переходит в вакуоль и откладывается в виде нерастворимых солей щавелевой, лимонной и других кислот.(…) Кальций используется в растительных клетках как вторичный посредник для контролирования многих процессов (закрытие устьиц, тропизм, рост пыльцевых трубок, акклиматизация к холоду, экспрессия генов, фотоморфогенез).(…) При недостатке кальция у делящихся клеток не образуются клеточные стенки и образуются многоядерные меристематические клетки. Недостаток кальция вызывает прекращение образования боковых корней и корневых волосков, приводит к набуханию пектиновых веществ, что вызывает ослизнение клеточных стенок и разрушение клеток. Также нарушается структура плазмалеммы и мембран клеточных органелл. Симптомами дефицита кальция является побеление с последующим почернением кончиков и краев листьев. Листовые пластинки искривляются и скручиваются. На плодах, в запасающих и сосудистых тканях появляются некротические участки.4.3.7. Магний
Недостаток в магнии растения испытывают на песчаных и подзолистых почвах. Много магния в сероземах, черноземы занимают промежуточное положение. Водорастворимого и обменного магния в почве 3-10 %. Магний поглощается растением в виде иона Mg2+. При снижении рН почвенного раствора магний поступает в растения в меньших количествах. Кальций, калий, аммоний и марганец действуют как конкуренты в процессе поглощения магния растениями. У высших растений среднее содержание магния составляет 0,02-3 %. Особенно много его в растениях короткого дня - кукурузе, просе, сорго, а также в картофеле, свекле и бобовых. Много магния в молодых клетках, а также в генеративных органах и запасающих тканях. Около 10-12 % магния находится в составе хлорофилла. Магний необходим для синтеза протопорфирина IX - непосредственного предшественника хлорофиллов. (…) Недостаток магния приводит к уменьшению содержания фосфора в растении, даже если фосфаты в достаточных количествах имеются в питательном субстрате. При недостатке магния тормозится превращение моносахаров в крахмал, слабо функционирует механизм синтеза белков, нарушается формирование пластид: матрикс хлоропластов просветляется и граны слипаются, ламеллы стромы разрываются и не образуют единой структуры. При магниевом голодании между зелеными жилками появляются пятна и полосы светло-зеленого, а затем желтого цвета. Края листовых пластинок приобретают желтый, оранжевый, красный или темно-красный цвет и такая как бы мраморная окраска наряду с хлорозом служит характерным симптомом нехватки магния. Признаки магниевой недостаточности сначала появляются на старых листьях, а затем распространяются на молодые листья.4.3.8. Кремний
Он обнаружен у всех растений. Особенно много его в клеточных стенках. Растения, накапливающие кремний, имеют прочные стебли. Недостаток кремния задерживает рост злаков (кукуруза, овес, ячмень) и двудольных растений (огурцы, томаты, табака, бобы). Исключение кремния во время репродуктивной стадии уменьшает количество семян, при этом снижается число зрелых семян, и нарушается ультраструктура клеточных органелл.4.3.9. Микроэлементы
Железо. Среднее содержание железа в растениях составляет 20-80 мг на 1 кг сухой массы. Ионы Fe3+ почвенного раствора восстанавливаются редокс-системами плазмалеммы клеток ризодермы до Fe2+ и в такой форме поступают в корень. Железо необходимо для функционирования основных редокс-систем фотосинтеза и дыхания, синтеза хлорофилла, восстановления нитратов и фиксации молекулярного азота клубеньковыми бактериями, входя в состав нитратредуктазы и нитрогеназы. Поэтому недостаточное поступление железа в растения в условиях переувлажнения и на карбонатных почвах приводит к снижению интенсивности дыхания и фотосинтеза и выражается в пожелтении (хлорозе) листьев и быстром их опадении. Марганец в клетки поступает в форме ионов Mn2+. Среднее его содержание составляет 1 мг на 1 кг сухой массы. Марганец накапливается в листьях. Он необходим для фоторазложения воды с выделением кислорода и восстановления углекислого газа при фотосинтезе. Марганец способствует увеличению содержания сахаров и их оттоку из листьев. (…) Характерный симптом марганцевого голодания - точечный хлороз листьев, когда между жилками появляются желтые пятна, а затем клетки в этих участках отмирают. Молибден. Наибольшее содержание молибдена характерно для бобовых (0,5-20 мг на 1 кг сухой массы), злаки содержат от 0,2 до 2 мг на кг сухой массы. Он поступает в растения в форме аниона МоО2-4, концентрируется в молодых, растущих органах. Его больше в листьях, чем в корнях и стеблях, а в листе сосредоточен, в основном, в хлоропластах. (…) При недостатке молибдена в тканях накапливается большое количество нитратов, не развиваются клубеньки на корнях бобовых, тормозится рост растений, наблюдаются деформации листовых пластинок. При высоких дозах молибден токсичен. При недостатке молибдена молодые листья по краям приобретают серую, а затем коричневую окраску, теряют тургор, а затем ткани листа отмирают и остаются только жилки в виде хлыстиков. Кобальт. Среднее содержание кобальта в растениях 0,02 мг на 1 кг сухой массы. Кобальт необходим бобовым растениям для обеспечения размножения клубеньковых бактерий. В растениях кобальт встречается в ионной форме и в витамине В12. Растения не вырабатывают этот витамин. Он синтезируется бактероидами клубеньков бобовых растений и участвует в синтезе метионина в бактероидах. При старении клубеньков и прекращении фиксации азота витамин выходит в цитоплазму клеток клубеньков. Наряду с магнием и марганцем кобальт активирует фермент гликолиза фосфоглюкомутазу и фермент аргиназу, гидролизующий аргинин. Внешние признаки недостатка кобальта сходны с признаками азотного голодания. Медь поступает в клетки в форме иона Сu2+. Среднее содержание меди в растениях 0,2 мг на кг сухой массы. (…) Влияя на содержание в растениях ингибиторов роста фенольной природы медь повышает устойчивость растений к полеганию. Она также повышает засухо-, морозо- и жароустойчивость. Недостаток меди вызывает задержку роста и цветения, хлороз, потерю тургора и завядание растений. У злаков при недостатке меди не развивается колос, у плодовых появляется суховершинность. При дефиците меди белеют и отмирают кончики листьев, листья и плоды плодовых деревьев покрываются бурыми пятнами. Цинк. Содержание цинка в надземных частях бобовых и злаковых растений составляет 15-60 мг на кг сухой массы. Повышенная концентрация отмечается в листьях, репродуктивных органах и конусах нарастания, наибольшая - в семенах. (…)Подкормка цинком способствует увеличению содержания ауксинов в тканях и активирует их рост. При дефиците цинка у растений нарушается фосфорный обмен: фосфор накапливается в корнях, задерживается его транспорт в надземные органы, замедляется превращение фосфора в органические формы. При недостатке цинка в растениях уменьшается содержание сахарозы и крахмала, увеличивается количество органических кислот и небелковых соединений азота - амидов и аминокислот. Кроме того, в 2-3 раза подавляется скорость деления клеток, что приводит к морфологическим изменениям листьев, нарушению растяжения клеток и дифференциации тканей. Наиболее характерный признак цинкового голодания - это задержка роста междоузлий и листьев, появление хлороза и развитие розеточности. Бор. Его среднее содержание составляет 0,1 мг на кг сухой массы. В боре наиболее нуждаются двудольные растения. Много бора в цветках. В клетках большая часть бора сосредоточена в клеточных стенках. Бор усиливает рост пыльцевых трубок, прорастание пыльцы, увеличивает количество цветков и плодов. Без него нарушается созревание семян. Бор снижает активность некоторых дыхательных ферментов, оказывает влияние на углеводный, белковый и нуклеиновый обмен. При недостатке бора нарушаются синтез, превращения и транспорт углеводов, формирование репродуктивных органов, оплодотворение и плодоношение. Он не может реутилизироваться и поэтому при борном голодании прежде всего отмирают конусы нарастания, останавливается рост побегов и корней, листовые пластинки утолщаются, скручиваются, становятся ломкими, цветки не образуются.4.4. Применение удобрений
В естественных биоценозах поглощенные из почвы соединения частично возвращаются с опавшими листьями, ветками, хвоей. С убранным урожаем сельскохозяйственных растений поглощенные вещества из почвы устраняются. Величина выноса минеральных элементов зависит от вида растения, урожайности и почвенно-климатических условий. Овощные культуры, картофель, многолетние травы выносят больше элементов питания, чем зерновые. Для предотвращения истощения почвы и получения высоких урожаев сельскохозяйственных культур необходимо внесение удобрений. Сопоставляя количество элементов в почве и растении с величиной урожая Ю. Либих сформулировал закон минимума или закон ограничивающих факторов. Согласно этому закону величина урожая зависит от количества того элемента, который находится в почве в относительном минимуме. Увеличение содержания этого элемента в почве за счет внесения удобрений будет приводить к возрастанию урожая до тех пор, пока в минимуме не окажется другой элемент. Позже было установлено наличие у растений критических периодов по отношению к тому или иному минеральному элементу, то есть периодах более высокой чувствительности растений к недостатку этого элемента на определенных этапах онтогенеза. Это позволяет регулировать соотношение питательных веществ в зависимости от фазы развития и условий среды. Так, известно, что в осенний период для озимых культур не рекомендуется вносить азотные удобрения, так как они усиливают ростовые процессы, снижая устойчивость растений. В осенний период надо проводить подкормки фосфором и калием, а весной азотом. С помощью удобрений можно регулировать не только величину урожая, но и его качество. Так, для получения зерна пшеницы с высоким содержанием белка надо вносить азотные удобрения, а для получения продуктов с высоким содержанием крахмала (например, зерна пивоваренного ячменя или клубни картофеля) необходимы фосфор и калий. Внекорневая подкормка фосфором незадолго до уборки усиливает отток ассимилятов из листьев сахарной свеклы к корнеплодам и тем самым увеличивает ее сахаристость. Система удобрений - это программа применения удобрений в севообороте с учетом растений-предшественников, плодородия почвы, климатических условий, биологических особенностей растений, состава и свойств удобрений. Система удобрений создается с учетом круговорота веществ и их баланса в земледелии. Баланс питательных веществ учитывает поступление их в почву с удобрениями, суммарный расход на формирование урожаев и непродуктивные потери в почве. Необходимое условие функционирования системы удобрений - это предотвращение загрязнения окружающей среды вносимыми в почву химическими соединениями. Удобрения подразделяют на минеральные и органические, промышленные (азотные, калийные, фосфорные, микроудобрения, бактериальные) и местные (навоз, торф, зола), простые (содержат один элемент питания - азотные, калийные, борные) и комплексные (содержат два или более питательных элементов). Среди комплексных удобрений выделяют сложные и комбинированные. Сложные удобрения в составе одного химического соединения содержат два или три питательных элемента, например, калийная селитра (KNO3), аммофос (Nh5h3PO4) и другие. Одна гранула комбинированных удобрений включает два или три основных элемента питания в виде различных химических соединений, например, нитроаммофоска. До посева вносят 2/3 общей нормы удобрений. Они должны обеспечить растение на весь период развития элементами питания и повысить плодородие почвы. Припосевное удобрение в виде хорошо растворимых соединений вносят малыми дозами одновременно с посевом или посадкой растений для обеспечения минерального питания молодых растений. Послепосевные внекорневые подкормки, основанные на способности листьев поглощать минеральные соли в растворе, проводятся для усиления питания растений в наиболее важные периоды их развития.
элементы питания растений, питательные элементы, макроэлементы, микроэлементы, удобрения
plantators.com
1. Средний состав основных химических элементов в сухом веществе растений и животных, %
Передовая наука и практика животноводства показали, что высокопродуктивные породы сельскохозяйственных животных можно создать только при условии хорошего и целенаправленного их кормления в различные возрастные и физиологические периоды. Установлено, что ценные качества животных невозможно сохранять без хорошего кормления. Породный скот быстро «вырождается», теряет свои качества при плохом кормлении, что в полной мере согласуется с мнением академика М. Ф. Иванова:
«Корма и кормление оказывают гораздо большее влияние на организм животного, чем порода и происхождение».
Таким образом, все вышесказанное подчеркивает многообразное влияние кормления на сельскохозяйственных животных, их природу и продуктивность. Поэтому, чтобы успешно использовать в практике животноводства это могучее средство воздействия на животных, необходимо знать физиологическое значение применяемых кормов и всех питательных веществ, содержащихся в них.
Питательные вещества кормов необходимы животному как источник энергии для жизнедеятельности организма, как источник структурного материала для образования органов и тканей, секреции молока и для отложения резервных веществ в теле. Чем полнее корм удовлетворяет эти потребности животного, тем он питательнее.
Чтобы иметь объективное представление о питательности того или иного корма и ее изменчивости под влиянием разных факторов необходимо знать содержание в кормах основньк питательных и биологически активных веществ. Иначе говоря, необходимо знать химический состав кормов, их переваримость и использование животными разных видов, возраста и направления продуктивности при содержании в различных хозяйственных условиях. Следовательно, питательность корма может быть определена по результатам изменения физиологического состояния животного и его продуктивности.
Химический состав кормов и физиологическое значение отдельных веществ.Основную долю кормов, используемых в кормлении сельскохозяйственных животных, составляют растительные кормовые средства и в значительно меньшем количестве продукты животного происхождения.
Из всех известных химических элементов (105) почти все обнаруживают в том или ином количестве в растениях и теле животных. Основными же химическими элементами (органогенами), составляющими растительное и животное вещество, являются углерод, кислород, водород и азот. В среднем элементарный состав сухого вещества растений и животных характеризуется следующими данными (табл. 1).
14
| Мине | |||||
| С | 0 | Н | N | ральные | |
| вещества | |||||
| Растение 45 42 6,5 1,5 5 | |||||
| Животное 63 13,8 9,4 5,0 8,8 | |||||
Углерод преобладает в составе растений и животных. В составе растений больше кислорода, а азота, углерода и водорода больше содержится в теле животных.
По своему химическому составу организм животных существенно отличается от растительных кормов (табл. 2).
2. Химический состав сухого вещества растительных кормов и тела животных, % (по а. П. Дмитроченко)
| Корм | Животные | |||||
| зеле | зерно | сено | ||||
| ный | куку | луго | бык | сви | кури- | |
| клевер | рузы | вое | нья | ца | ||
| Вода | 77,8 13,0 14,3 | 54,0 | 58,0 | 56,0 | ||
| Сухое вещество | 22,2 87,0 85,7 | 46,0 | 42,0 | 44,0 | ||
| Протеин | 16,6 10,1 11,3 | 32,6 | 35,7 | 47,7 | ||
| Жир | 4,4 4,5 2,9 | 55,2 | 55,2 | 40,9 | ||
| Клетчатка | 22,5 2,2 30,7 | - | - | - | ||
| Безазотистые | ||||||
| экстрактивные | ||||||
| вещества | 47,9 81,6 47,9 | 2,2 | 2,5 | 1,6 | ||
| Зола | 8,6 1,6 7,2 | 10,0 | 6,6 | 9,8 | ||
В составе сухого вещества тела животных основную долю составляют протеин и жир, тогда как у большинства растений сухое вещество в основном представлено углеводами (клетчаткой, крахмалом и др.). Содержание же углеводов в животном организме очень низкое. Это связано с тем, что клеточные стенки растений состоят в основном из целлюлозы, а стенки клетки животного - из белка. У животных из белка состоят также мышцы, кожа, волосы, шерсть и др. В растениях же протеин представлен в основном различными ферментами и как структурное вещество он не характерен для растительной ткани. Кроме того, растения откладывают энергию в форме углеводов, тогда как у животных она сосредоточена в основном в жировых депо.
Довольно значительные различия отмечаются и по содержанию минеральных веществ в растениях и животных организмах (табл. 3).
studfiles.net
Среднее содержание железа в растениях составляет 20-80 мг на 1 кг сухой массы
Железо необходимо для функционирования основных редокс-систем фотосинтеза и дыхания, синтеза хлорофилла, восстановления нитратов и фиксации молекулярного азота клубеньковыми бактериями, входя в состав нитратредуктазы и нитрогеназы. Поэтому недостаточное поступление железа в растения в условиях переувлажнения и на карбонатных почвах приводит к снижению интенсивности дыхания и фотосинтеза и выражается в пожелтении (хлорозе) листьев и быстром их опадении.
Марганец в клетки поступает в форме ионов Mn2+. Среднее его содержание составляет 1 мг на 1 кг сухой массы. Марганец накапливается в листьях. Он необходим для фоторазложения воды с выделением кислорода и восстановления углекислого газа при фотосинтезе. Марганец способствует увеличению содержания сахаров и их оттоку из листьев. Два фермента цикла Кребса - малат- и изоцитратдегидрогеназы - активируются ионами марганца. Он также необходим для функционирования нитратредуктазы при восстановлении нитратов. Марганец является кофактором РНКполимеразы и ауксиноксидазы, разрушающей фитогормон 3-индолилуксусную кислоту.
Характерный симптом марганцевого голодания - точечный хлороз листьев, когда между жилками появляются желтые пятна, а затем клетки в этих участках отмирают.
Молибден. Наибольшее содержание молибдена характерно для бобовых (0,5-20 мг на 1 кг сухой массы), злаки содержат от 0,2 до 2 мг на кг сухой массы. Он поступает в растения в форме аниона МоО2-4, концентрируется в молодых, растущих органах. Его больше в листьях, чем в корнях и стеблях, а в листе сосредоточен, в основном, в хлоропластах.
Молибден входит в состав нитратредуктазы и нитрогеназы, а также необходим для биосинтеза легоглобина. Как металл-активатор молибден участвует в реакциях аминирования и переаминирования, для включения аминокислот в пептидную цепь, работы таких ферментов как ксантиноксидаза и различных фосфатаз.
При недостатке молибдена в тканях накапливается большое количество нитратов, не развиваются клубеньки на корнях бобовых, тормозится рост растений, наблюдаются деформации листовых пластинок. При высоких дозах молибден токсичен. При недостатке молибдена молодые листья по краям приобретают серую, а затем коричневую окраску, теряют тургор, а затем ткани листа отмирают и остаются только жилки в виде хлыстиков.Кобальт. Среднее содержание кобальта в растениях 0,02 мг на 1 кг сухой массы. Кобальт необходим бобовым растениям для обеспечения размножения клубеньковых бактерий. В растениях кобальт встречается в ионной форме и в витамине В12. Растения не вырабатывают этот витамин. Он синтезируется бактероидами клубеньков бобовых растений и участвует в синтезе метионина в бактероидах. При старении клубеньков и прекращении фиксации азота витамин выходит в цитоплазму клеток клубеньков. Наряду с магнием и марганцем кобальт активирует фермент гликолиза фосфоглюкомутазу и фермент аргиназу, гидролизующий аргинин.
Внешние признаки недостатка кобальта сходны с признаками азотного голодания.
Медь поступает в клетки в форме иона Сu2+. Среднее содержание меди в растениях 0,2 мг на кг сухой массы. Около 70 % всей меди, находящейся в листьях, сосредоточены в хлоропластах и почти половина ее в составе пластоцианина - переносчика электронов между фотосистемами II и I. Она входит в состав ферментов, катализирующих окисление аскорбиновой кислоты, дифенолов и гидроксилирование монофенолов - аскорбатоксидазы, полифенолоксидазы, ортодифенолоксидазы и тирозиназы. Два атома меди функционируют в цитохромоксидазном комплексе дыхательной цепи митохондрий. Медь входит в состав нитратредуктазного комплекса и влияет на синтез легоглобина. Для биосинтеза этилена также необходим медьсодержащий фермент. Влияя на содержание в растениях ингибиторов роста фенольной природы медь повышает устойчивость растений к полеганию. Она также повышает засухо-, морозо- и жароустойчивость. Недостаток меди вызывает задержку роста и цветения, хлороз, потерю тургора и завядание растений. У злаков при недостатке меди не развивается колос, у плодовых появляется суховершинность. При дефиците меди белеют и отмирают кончики листьев, листья и плоды плодовых деревьев покрываются бурыми пятнами.
Цинк. Содержание цинка в надземных частях бобовых и злаковых растений составляет 15-60 мг на кг сухой массы. Повышенная концентрация отмечается в листьях, репродуктивных органах и конусах нарастания, наибольшая - в семенах.Цинк поступает в растение в форме катиона Zn2+. Он необходим для функционирования ряда ферментов гликолиза - гексокиназы, енолазы, триозофосфатдегидрогеназы, альдолазы, а также входит в состав алкогольдегидрогеназы. Цинк активирует карбоангидразу, катализирующую реакцию дегидратации гидрата оксида углерода: Н2СО3 СО2 + Н2О, что помогает использованию углекислого газа в процессе фотосинтеза. Цинк участвует в образовании аминокислоты триптофана. Именно с этим связано влияние катионов цинка на синтез белков, а также фитогормона 3-индолилуксусной кислоты, предшественником которой является триптофан. Подкормка цинком способствует увеличению содержания ауксинов в тканях и активирует их рост.
При дефиците цинка у растений нарушается фосфорный обмен: фосфор накапливается в корнях, задерживается его транспорт в надземные органы, замедляется превращение фосфора в органические формы. При недостатке цинка в растениях уменьшается содержание сахарозы и крахмала, увеличивается количество органических кислот и небелковых соединений азота - амидов и аминокислот. Кроме того, в 2-3 раза подавляется скорость деления клеток, что приводит к морфологическим изменениям листьев, нарушению растяжения клеток и дифференциации тканей. Наиболее характерный признак цинкового голодания - это задержка роста междоузлий и листьев, появление хлороза и развитие розеточности.
Бор. Его среднее содержание составляет 0,1 мг на кг сухой массы. В боре наиболее нуждаются двудольные растения. Много бора в цветках. В клетках большая часть бора сосредоточена в клеточных стенках. Бор усиливает рост пыльцевых трубок, прорастание пыльцы, увеличивает количество цветков и плодов. Без него нарушается созревание семян. Бор снижает активность некоторых дыхательных ферментов, оказывает влияние на углеводный, белковый и нуклеиновый обмен. При недостатке бора нарушаются синтез, превращения и транспорт углеводов, формирование репродуктивных органов, оплодотворение и плодоношение. Он не может реутилизироваться и поэтому при борном голодании прежде всего отмирают конусы нарастания, останавливается рост побегов и корней, листовые пластинки утолщаются, скручиваются, становятся ломкими, цветки не образуются.4.4. Применение удобрений
В естественных биоценозах поглощенные из почвы соединения частично возвращаются с опавшими листьями, ветками, хвоей. С убранным урожаем сельскохозяйственных растений поглощенные вещества из почвы устраняются. Величина выноса минеральных элементов зависит от вида растения, урожайности и почвенно-климатических условий. Овощные культуры, картофель, многолетние травы выносят больше элементов питания, чем зерновые.
Для предотвращения истощения почвы и получения высоких урожаев сельскохозяйственных культур необходимо внесение удобрений. Сопоставляя количество элементов в почве и растении с величиной урожая Ю. Либих сформулировал закон минимума или закон ограничивающих факторов. Согласно этому закону величина урожая зависит от количества того элемента, который находится в почве в относительном минимуме. Увеличение содержания этого элемента в почве за счет внесения удобрений будет приводить к возрастанию урожая до тех пор, пока в минимуме не окажется другой элемент. Позже было установлено наличие у растений критических периодов по отношению к тому или иному минеральному элементу, то есть периодах более высокой чувствительности растений к недостатку этого элемента на определенных этапах онтогенеза. Это позволяет регулировать соотношение питательных веществ в зависимости от фазы развития и условий среды. Так, известно, что в осенний период для озимых культур не рекомендуется вносить азотные удобрения, так как они усиливают ростовые процессы, снижая устойчивость растений. В осенний период надо проводить подкормки фосфором и калием, а весной азотом.
С помощью удобрений можно регулировать не только величину урожая, но и его качество. Так, для получения зерна пшеницы с высоким содержанием белка надо вносить азотные удобрения, а для получения продуктов с высоким содержанием крахмала (например, зерна пивоваренного ячменя или клубни картофеля) необходимы фосфор и калий. Внекорневая подкормка фосфором незадолго до уборки усиливает отток ассимилятов из листьев сахарной свеклы к корнеплодам и тем самым увеличивает ее сахаристость.Система удобрений - это программа применения удобрений в севообороте с учетом растений-предшественников, плодородия почвы, климатических условий, биологических особенностей растений, состава и свойств удобрений. Система удобрений создается с учетом круговорота веществ и их баланса в земледелии. Баланс питательных веществ учитывает поступление их в почву с удобрениями, суммарный расход на формирование урожаев и непродуктивные потери в почве. Необходимое условие функционирования системы удобрений - это предотвращение загрязнения окружающей среды вносимыми в почву химическими соединениями.
Удобрения подразделяют на минеральные и органические, промышленные (азотные, калийные, фосфорные, микроудобрения, бактериальные) и местные (навоз, торф, зола), простые (содержат один элемент питания - азотные, калийные, борные) и комплексные (содержат два или более питательных элементов). Среди комплексных удобрений выделяют сложные и комбинированные. Сложные удобрения в составе одного химического соединения содержат два или три питательных элемента, например, калийная селитра (KNO3), аммофос (Nh5h3PO4) и другие. Одна гранула комбинированных удобрений включает два или три основных элемента питания в виде различных химических соединений, например, нитроаммофоска.
До посева вносят 2/3 общей нормы удобрений. Они должны обеспечить растение на весь период развития элементами питания и повысить плодородие почвы. Припосевное удобрение в виде хорошо растворимых соединений вносят малыми дозами одновременно с посевом или посадкой растений для обеспечения минерального питания молодых растений. Послепосевные внекорневые подкормки, основанные на способности листьев поглощать минеральные соли в растворе, проводятся для усиления питания растений в наиболее важные периоды их развития.
5. ФОТОСИНТЕЗ
Фотосинтез – это процесс трансформации поглощенной растением электромагнитной энергии солнечного света в химическую энергию органических соединений. Чтобы свет мог быть использованным в процессе фотосинтеза, необходимо его поглощение фоторецепторами – пигментами.5.1. Пигменты
5.1.1. Хлорофиллы
У всех высших растений, водорослей и цианобактерий содержится хлорофилл a, хлорофилл b имеется у высших растений и зеленых водорослей. Хлорофилл с, лишенный фитола, содержится в бурых и диатомовых водорослях, хлорофилл d – в красных водорослях. Фотосинтезирующие зеленые бактерии имеют бактериохлорофиллы c и d, пурпурные бактерии - бактериохлорофиллы a и b.
У хлорофилла а четыре пиррольных кольца соединены между собой метиновыми мостиками (=СН-), образуя порфириновое кольцо. Кроме того, атомы азота пиррольных колец связаны с атомом магния. С порфириновым ядром соединено циклопентановое кольцо, образованное остатком кетопропионовой кислоты и содержащее активные карбонильную (С=О) и метилированную карбоксильную (О=С-О-СН3) группы. Структура, состоящая из порфиринового ядра и циклопентанового кольца, называется форбином. Боковая цепь, состоящая из пропионовой кислоты и непредельного спирта фитола, связана с атомом углерода IV пиррольного кольца (рис. 5.1). Хлорофилл, лишенный фитола, называется хлорофиллидом. Если атом магния замещен протоном, то такое соединение носит название феофитина. Активность хлорофиллов, также как и других пигментов, обусловлена наличием большого количества двойных связей с делокализованными электронами.
Рис. 5.1. Структурные формулы хлорофиллов a и b (по В. В. Полевому).
Хлорофиллы хорошо растворимы в органических растворителях (этиловом эфире, бензоле, хлороформе, ацетоне, этиловом спирте) и нерастворимы в воде. Хлорофиллы имеют максимумы поглощения света в красной и синей частях спектра. Растворы хлорофиллов обладают флуоресценцией и фосфоресценцией.
5.1.2. Каротиноиды
Каротиноиды – жирорастворимые пигменты, присутствующие в хлоропластах всех растений. Они входят в состав хромопластов в незеленых частях растений, например, корнеплодов моркови. К каротиноидам относят 3 группы соединений: 1) оранжевые или красные каротины, 2) желтые ксантофиллы, 3) каротиноидные кислоты. Каротины и ксантофиллы состоят из 8 остатков изопрена, которые образуют цепь конъюгированных двойных связей (рис. 5.2). Основные каротиноиды - -каротин, лютеин, виолаксантин и неоксантин.
Рис. 5.2. Структурные формулы каротиноидов и последовательность их превращений (по В. В. Полевому).
Каротины и ксантофиллы растворимы в хлороформе, бензоле, сероуглероде, ацетоне. Каротины хорошо растворяются в эфирах, но плохо в спиртах, а ксантофиллы наоборот. Каротиноиды имеют максимумы поглощения в фиолетово-синей и синей частях спектра света. Они не способны к флуоресценции.
Главные функции каротиноидов: поглощение света в качестве дополнительных пигментов, защита молекул хлорофиллов от необратимого фотоокисления, тушение активных радикалов, участие в фототропизме, так как способствуют определению направления роста побега.
5.1.3. Фикобилины
Сине-зеленые и красные водоросли помимо хлорофилла а и каротиноидов содержат пигменты фикобилины. Их молекула состоит из 4 последовательных пиррольных колец (рис. 5.3). Фикобилины являются хромофорными группами глобулиновых белков фикобилипротеинов. Они делятся на 3 группы: 1) фикоэритрины – белки красного цвета, 2) фикоцианины – сине-голубые белки и 3) аллофикоцианины – синие белки. Все они обладают флуоресценцией и растворимы в воде.
Рис. 5.3. Структурные формулы фикобилинов (по В. В. Полевому).
Фикобилины имеют максимумы поглощения в оранжевой, желтой и зеленой частях спектра света. Это позволяет водорослям полнее использовать свет, проникающий в воду. Вода обладает светопоглощающей способностью. На глубине около 30 м полностью исчезают красные лучи, около 180 м – желтые, 320 м – зеленые, а на глубину более 500 м не проникают синие и фиолетовые лучи. Фикобилины – это дополнительные пигменты, участвующие в светособирающем комплексе. Около 90 % энергии света, поглощенного фикобилинами, передается на хлорофилл а.У растений имеется фикобилин фитохром. Он не участвует в фотосинтезе, но является фоторецептором красного и дальнего красного света и выполняет регуляторные функции в клетках растений.
5.2. Световая фаза фотосинтеза
Фотосинтез имеет две фазы – световую и темновую. В световой фазе молекулы пигментов поглощают фотоны, передают поглощенную энергию молекулам хлорофилла а, происходит трансформация энергии света в химическую энергию АТФ и восстановленного НАДФН, выделяется кислород в результате фоторазложения воды. Все эти процессы происходят на мембранах хлоропластов. При темновой фазе фотосинтеза в строме хлоропластов восстанавливается поглощенный СО2 с образованием углеводов и других органических соединений.
5.2.1. Поглощение света и возбуждение хлорофилла
Электрон представляет собой заряженную частицу, которая движется по орбите вокруг ядра атома. Электрон обладает вектором магнитного момента – спином, который отражает направление вращения электрона вокруг своей оси. В стабильном состоянии валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули: два электрона с антипараллельными спинами на каждой орбитали. Такое состояние молекулы называют основным синглетным состоянием S0. При поглощении молекулой пигмента фотона электрон переходит на более высокую орбиталь за счет энергии фотона. Если у возбужденного электрона сохраняется направление спина, то считают, что молекула находится в синглетном возбужденном состоянии S*. Если возбужденный электрон меняет спин, то такое возбужденное состояние называется триплетным Т*.
Поглощение молекулой хлорофилла кванта красного света приводит к синглетному возбужденному состоянию S*1. При поглощении кванта синего цвета с более высоким уровнем энергии электрон переходит на более высокую орбиталь S*2. Время жизни электрона на S*2 составляет не более 10-12 секунды. За такое короткое время энергия электронного возбуждения не может быть использована и электрон возвращается на S*1, потеряв немного энергии в виде тепла. Время жизни электрона в состоянии S*1 равно 10-9. Если молекула пигмента не участвует в фотохимической реакции, то электрон возвращается в стабильное состояние. При этом часть энергии теряется в виде тепла и излучается квант света с большей по сравнению с поглощенным фотоном длиной волны – явление флуоресценции.Время жизни электрона в состоянии Т* равно 10-4-10-2 секунды. Из триплетного возбужденного состояния молекула может вернуться в стабильное состояние, излучив кроме тепла еще более длинноволновый, чем при флуоресценции, квант света. Это слабое свечение называют фосфоресценцией. Если молекула в возбужденном состоянии участвует в фотохимических реакциях с передачей электрона акцептору, то флуоресценции и фосфоресценции не происходит.
Для более полного использования падающего на листья света энергия фотонов улавливается 200-400 молекулами пигментов светособирающего (антенного) комплекса и передается к одной молекуле хлорофилла а, являющейся реакционным центром, которая и участвует в фотохимических реакциях. Передача энергии молекулами пигментов происходит по принципу индуктивного резонанса. Молекула пигмента, поглотившая фотон, создает переменное электрическое поле, которое усиливает колебания электрона в соседней молекуле. Это происходит из-за перекрытия частот колебаний и очень маленького расстояния между молекулами пигментов. Тем более, что при освещении листьев тилакоиды и граны сдвигаются и уплотняются, поэтому хлоропласты уменьшаются в объеме. Передача энергии молекулами пигментов идет с большой эффективностью. Так, от хлорофилла b к хлорофиллу а передается примерно 90 %, а от каротиноидов к хлорофиллу а – 40 % поглощенной энергии.
Совокупность молекул светособирающего комплекса и реакционного центра составляет фотосистему. Предположение о существовании в хлоропластах двух фотосистем высказал Р. Эмерсон в 1957 г, изучая влияние света на квантовый выход фотосинтеза у водоросли хлореллы. Квантовый выход фотосинтеза – это количество выделившегося кислорода или связанного углекислого газа на 1 квант поглощенной энергии. Он установил, что при одновременном освещении хлореллы коротковолновым (650 нм) и длинноволновым (700 нм) красным светом эффект выше, чем суммарный квантовый выход при воздействии красным светом этих длин в отдельности. Это явление получило название эффекта усиления Эмерсона. Позже предположение Р. Эмерсона о наличии двух фотосистем получило экспериментальное подтверждение.5.2.2. Нециклический и циклический транспорт электронов
В фотосистеме II реакционный центр, состоящий из димера хлорофилла а с максимумом поглощения при 680 нм (П680), последовательно поглотив энергию, равную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, передает 2 электрона феофитину. От феофитина электроны последовательно передаются на пластохиноны QA и QB, расположенные на другой стороне мембраны тилакоида, затем на две липидорастворимые молекулы пластохинона PQ. Он является переносчиком протонов и электронов, выполняющего в мембране роль челнока. Принимая 2 электрона и захватывая из стромы 2 Н+, 2 молекулы PQh3 диффундируют через липидную фазу мембраны. Электроны от 2 PQh3 поступают в цепь:
железосерный белок FeSr цитохром f Cu-содержащий белок пластоцианин П700,
а протоны попадают в полость тилакоида.
Вакантные места в П+680 заполняются 2 электронами из содержащего Mn переносчика электронов, который восстанавливается с участием белкового комплекса S. Окисленный комплекс S связывает воду и восстанавливается за счет электронов воды. При фотоокислении воды комплексом S вторая пара протонов освобождается в полость тилакоида, а атом кислорода диффундирует из хлоропласта.
После возбуждения реакционного центра фотосистемы I - молекулы хлорофилла а с максимумом поглощения при 700 нм (П700), 2 квантами длинноволного красного света 2 электрона передаются мономерной форме хлорофилла а – А1 и затем переносчикам электронoв железосерным белкам А2 и АВ, ферредоксину на наружной стороне мембраны тилакоида, НАДФ-редуктазе. Этот фермент восстанавливает НАДФ в НАДФН. На вакантные места в П+700 переходят электроны с пластоцианина и нециклическая цепь транспорта электронов замыкается.В мембранах хлоропластов может функционировать циклический транспорт электронов, когда действует только фотосистема I. Возбужденные молекулы П700 передают электроны на А1, железосерные белки А2 и АВ, переносящие электроны на другую сторону мембраны, ферредоксин, пластохинон, цитохром b6, железосерный белок FeSr, цитохром f, пластоцианин и П700 (рис. 5.4). Энергия, освобождающаяся при транспорте электронов, используется для фосфорилирования АДФ.
Рис. 5.4. Локализация электрон- и протонтранспортных реакций в тилакоидной мембране хлоропласта (по В. В. Полевому).
Механизм фотофосфорилирования АДФ объясняет теория П. Митчелла. Согласно этой теории, трансмембранный перенос пластохинонами электронов и протонов в одну сторону чередуется с переносом цитохромной системой в обратную сторону электронов. Поэтому по одну сторону мембраны накапливается избыток протонов и возникает электрохимический мембранный потенциал. Его энергия используется для синтеза АТФ при разрядке мембраны в результате транспорта протонов через мембрану посредством Н+-АТФазы, которая действует как АТФсинтетаза. Этот фермент выглядит как грибовидная частица на поверхности мембраны тилакоидов. Его молекула состоит из двух частей: головки, выступающей с наружной стороны мембраны и содержащей активный центр фермента, и ножки, погруженной в мембрану и представляющей канал, через который передвигаются протоны.
5.3. Темновая фаза фотосинтеза
5.3.1. С3-путь фотосинтеза или цикл Кальвина
Этот путь ассимиляции СО2 обнаружен у всех фотосинтезирующих растений. Он был назван циклом Кальвина в честь американского биохимика М. Кальвина, который с сотрудниками открыл и изучил его в 1946-1956 годах с помощью метода меченых атомов и хроматографии. Растения разное время подкармливали 14СО2 и хроматографировали экстракты. Радиоактивные изотопы по химическим свойствам не отличаются от стабильных. Принимая участие в реакциях, они помечают те соединения, в которые входят. Цикл состоит из трех этапов: карбоксилирования, восстановления, регенерации первичного акцептора СО2 и синтеза конечного продукта фотосинтеза (рис. 5.5).
Рис. 5.5. Цикл Кальвина (С3-путь фотосинтеза.
1 – фосфорибулокиназа, 2 – рибулозодифосфаткарбоксилаза, 3 - фофсфоглицераткиназа, 4 – триозофосфатдегидрогеназа, 5 – триозофосфатизомераза, 6 – альдолаза, 7 – фосфатаза, 8 – транскетолаза, 9 – альдолаза, 10 – фосфатаза, 11 – транскетолаза, 12 – рибозофосфатизомераза, 13 – фосфокетопентоэпимераза (по В. В. Полевому).
shkolakz.ru
Среднее содержание железа в растениях составляет 20-80 мг на 1 кг сухой массы
Железо необходимо для функционирования основных редокс-систем фотосинтеза и дыхания, синтеза хлорофилла, восстановления нитратов и фиксации молекулярного азота клубеньковыми бактериями, входя в состав нитратредуктазы и нитрогеназы. Поэтому недостаточное поступление железа в растения в условиях переувлажнения и на карбонатных почвах приводит к снижению интенсивности дыхания и фотосинтеза и выражается в пожелтении (хлорозе) листьев и быстром их опадении.
Марганец в клетки поступает в форме ионов Mn2+. Среднее его содержание составляет 1 мг на 1 кг сухой массы. Марганец накапливается в листьях. Он необходим для фоторазложения воды с выделением кислорода и восстановления углекислого газа при фотосинтезе. Марганец способствует увеличению содержания сахаров и их оттоку из листьев. Два фермента цикла Кребса - малат- и изоцитратдегидрогеназы - активируются ионами марганца. Он также необходим для функционирования нитратредуктазы при восстановлении нитратов. Марганец является кофактором РНКполимеразы и ауксиноксидазы, разрушающей фитогормон 3-индолилуксусную кислоту.
Характерный симптом марганцевого голодания - точечный хлороз листьев, когда между жилками появляются желтые пятна, а затем клетки в этих участках отмирают.
Молибден. Наибольшее содержание молибдена характерно для бобовых (0,5-20 мг на 1 кг сухой массы), злаки содержат от 0,2 до 2 мг на кг сухой массы. Он поступает в растения в форме аниона МоО2-4, концентрируется в молодых, растущих органах. Его больше в листьях, чем в корнях и стеблях, а в листе сосредоточен, в основном, в хлоропластах.
Молибден входит в состав нитратредуктазы и нитрогеназы, а также необходим для биосинтеза легоглобина. Как металл-активатор молибден участвует в реакциях аминирования и переаминирования, для включения аминокислот в пептидную цепь, работы таких ферментов как ксантиноксидаза и различных фосфатаз.
При недостатке молибдена в тканях накапливается большое количество нитратов, не развиваются клубеньки на корнях бобовых, тормозится рост растений, наблюдаются деформации листовых пластинок. При высоких дозах молибден токсичен. При недостатке молибдена молодые листья по краям приобретают серую, а затем коричневую окраску, теряют тургор, а затем ткани листа отмирают и остаются только жилки в виде хлыстиков.Кобальт. Среднее содержание кобальта в растениях 0,02 мг на 1 кг сухой массы. Кобальт необходим бобовым растениям для обеспечения размножения клубеньковых бактерий. В растениях кобальт встречается в ионной форме и в витамине В12. Растения не вырабатывают этот витамин. Он синтезируется бактероидами клубеньков бобовых растений и участвует в синтезе метионина в бактероидах. При старении клубеньков и прекращении фиксации азота витамин выходит в цитоплазму клеток клубеньков. Наряду с магнием и марганцем кобальт активирует фермент гликолиза фосфоглюкомутазу и фермент аргиназу, гидролизующий аргинин.
Внешние признаки недостатка кобальта сходны с признаками азотного голодания.
Медь поступает в клетки в форме иона Сu2+. Среднее содержание меди в растениях 0,2 мг на кг сухой массы. Около 70 % всей меди, находящейся в листьях, сосредоточены в хлоропластах и почти половина ее в составе пластоцианина - переносчика электронов между фотосистемами II и I. Она входит в состав ферментов, катализирующих окисление аскорбиновой кислоты, дифенолов и гидроксилирование монофенолов - аскорбатоксидазы, полифенолоксидазы, ортодифенолоксидазы и тирозиназы. Два атома меди функционируют в цитохромоксидазном комплексе дыхательной цепи митохондрий. Медь входит в состав нитратредуктазного комплекса и влияет на синтез легоглобина. Для биосинтеза этилена также необходим медьсодержащий фермент. Влияя на содержание в растениях ингибиторов роста фенольной природы медь повышает устойчивость растений к полеганию. Она также повышает засухо-, морозо- и жароустойчивость. Недостаток меди вызывает задержку роста и цветения, хлороз, потерю тургора и завядание растений. У злаков при недостатке меди не развивается колос, у плодовых появляется суховершинность. При дефиците меди белеют и отмирают кончики листьев, листья и плоды плодовых деревьев покрываются бурыми пятнами.
Цинк. Содержание цинка в надземных частях бобовых и злаковых растений составляет 15-60 мг на кг сухой массы. Повышенная концентрация отмечается в листьях, репродуктивных органах и конусах нарастания, наибольшая - в семенах.Цинк поступает в растение в форме катиона Zn2+. Он необходим для функционирования ряда ферментов гликолиза - гексокиназы, енолазы, триозофосфатдегидрогеназы, альдолазы, а также входит в состав алкогольдегидрогеназы. Цинк активирует карбоангидразу, катализирующую реакцию дегидратации гидрата оксида углерода: Н2СО3 СО2 + Н2О, что помогает использованию углекислого газа в процессе фотосинтеза. Цинк участвует в образовании аминокислоты триптофана. Именно с этим связано влияние катионов цинка на синтез белков, а также фитогормона 3-индолилуксусной кислоты, предшественником которой является триптофан. Подкормка цинком способствует увеличению содержания ауксинов в тканях и активирует их рост.
При дефиците цинка у растений нарушается фосфорный обмен: фосфор накапливается в корнях, задерживается его транспорт в надземные органы, замедляется превращение фосфора в органические формы. При недостатке цинка в растениях уменьшается содержание сахарозы и крахмала, увеличивается количество органических кислот и небелковых соединений азота - амидов и аминокислот. Кроме того, в 2-3 раза подавляется скорость деления клеток, что приводит к морфологическим изменениям листьев, нарушению растяжения клеток и дифференциации тканей. Наиболее характерный признак цинкового голодания - это задержка роста междоузлий и листьев, появление хлороза и развитие розеточности.
Бор. Его среднее содержание составляет 0,1 мг на кг сухой массы. В боре наиболее нуждаются двудольные растения. Много бора в цветках. В клетках большая часть бора сосредоточена в клеточных стенках. Бор усиливает рост пыльцевых трубок, прорастание пыльцы, увеличивает количество цветков и плодов. Без него нарушается созревание семян. Бор снижает активность некоторых дыхательных ферментов, оказывает влияние на углеводный, белковый и нуклеиновый обмен. При недостатке бора нарушаются синтез, превращения и транспорт углеводов, формирование репродуктивных органов, оплодотворение и плодоношение. Он не может реутилизироваться и поэтому при борном голодании прежде всего отмирают конусы нарастания, останавливается рост побегов и корней, листовые пластинки утолщаются, скручиваются, становятся ломкими, цветки не образуются.4.4. Применение удобрений
В естественных биоценозах поглощенные из почвы соединения частично возвращаются с опавшими листьями, ветками, хвоей. С убранным урожаем сельскохозяйственных растений поглощенные вещества из почвы устраняются. Величина выноса минеральных элементов зависит от вида растения, урожайности и почвенно-климатических условий. Овощные культуры, картофель, многолетние травы выносят больше элементов питания, чем зерновые.
Для предотвращения истощения почвы и получения высоких урожаев сельскохозяйственных культур необходимо внесение удобрений. Сопоставляя количество элементов в почве и растении с величиной урожая Ю. Либих сформулировал закон минимума или закон ограничивающих факторов. Согласно этому закону величина урожая зависит от количества того элемента, который находится в почве в относительном минимуме. Увеличение содержания этого элемента в почве за счет внесения удобрений будет приводить к возрастанию урожая до тех пор, пока в минимуме не окажется другой элемент. Позже было установлено наличие у растений критических периодов по отношению к тому или иному минеральному элементу, то есть периодах более высокой чувствительности растений к недостатку этого элемента на определенных этапах онтогенеза. Это позволяет регулировать соотношение питательных веществ в зависимости от фазы развития и условий среды. Так, известно, что в осенний период для озимых культур не рекомендуется вносить азотные удобрения, так как они усиливают ростовые процессы, снижая устойчивость растений. В осенний период надо проводить подкормки фосфором и калием, а весной азотом.
С помощью удобрений можно регулировать не только величину урожая, но и его качество. Так, для получения зерна пшеницы с высоким содержанием белка надо вносить азотные удобрения, а для получения продуктов с высоким содержанием крахмала (например, зерна пивоваренного ячменя или клубни картофеля) необходимы фосфор и калий. Внекорневая подкормка фосфором незадолго до уборки усиливает отток ассимилятов из листьев сахарной свеклы к корнеплодам и тем самым увеличивает ее сахаристость.Система удобрений - это программа применения удобрений в севообороте с учетом растений-предшественников, плодородия почвы, климатических условий, биологических особенностей растений, состава и свойств удобрений. Система удобрений создается с учетом круговорота веществ и их баланса в земледелии. Баланс питательных веществ учитывает поступление их в почву с удобрениями, суммарный расход на формирование урожаев и непродуктивные потери в почве. Необходимое условие функционирования системы удобрений - это предотвращение загрязнения окружающей среды вносимыми в почву химическими соединениями.
Удобрения подразделяют на минеральные и органические, промышленные (азотные, калийные, фосфорные, микроудобрения, бактериальные) и местные (навоз, торф, зола), простые (содержат один элемент питания - азотные, калийные, борные) и комплексные (содержат два или более питательных элементов). Среди комплексных удобрений выделяют сложные и комбинированные. Сложные удобрения в составе одного химического соединения содержат два или три питательных элемента, например, калийная селитра (KNO3), аммофос (Nh5h3PO4) и другие. Одна гранула комбинированных удобрений включает два или три основных элемента питания в виде различных химических соединений, например, нитроаммофоска.
До посева вносят 2/3 общей нормы удобрений. Они должны обеспечить растение на весь период развития элементами питания и повысить плодородие почвы. Припосевное удобрение в виде хорошо растворимых соединений вносят малыми дозами одновременно с посевом или посадкой растений для обеспечения минерального питания молодых растений. Послепосевные внекорневые подкормки, основанные на способности листьев поглощать минеральные соли в растворе, проводятся для усиления питания растений в наиболее важные периоды их развития.
5. ФОТОСИНТЕЗ
Фотосинтез – это процесс трансформации поглощенной растением электромагнитной энергии солнечного света в химическую энергию органических соединений. Чтобы свет мог быть использованным в процессе фотосинтеза, необходимо его поглощение фоторецепторами – пигментами.5.1. Пигменты
5.1.1. Хлорофиллы
У всех высших растений, водорослей и цианобактерий содержится хлорофилл a, хлорофилл b имеется у высших растений и зеленых водорослей. Хлорофилл с, лишенный фитола, содержится в бурых и диатомовых водорослях, хлорофилл d – в красных водорослях. Фотосинтезирующие зеленые бактерии имеют бактериохлорофиллы c и d, пурпурные бактерии - бактериохлорофиллы a и b.
У хлорофилла а четыре пиррольных кольца соединены между собой метиновыми мостиками (=СН-), образуя порфириновое кольцо. Кроме того, атомы азота пиррольных колец связаны с атомом магния. С порфириновым ядром соединено циклопентановое кольцо, образованное остатком кетопропионовой кислоты и содержащее активные карбонильную (С=О) и метилированную карбоксильную (О=С-О-СН3) группы. Структура, состоящая из порфиринового ядра и циклопентанового кольца, называется форбином. Боковая цепь, состоящая из пропионовой кислоты и непредельного спирта фитола, связана с атомом углерода IV пиррольного кольца (рис. 5.1). Хлорофилл, лишенный фитола, называется хлорофиллидом. Если атом магния замещен протоном, то такое соединение носит название феофитина. Активность хлорофиллов, также как и других пигментов, обусловлена наличием большого количества двойных связей с делокализованными электронами.
Рис. 5.1. Структурные формулы хлорофиллов a и b (по В. В. Полевому).
Хлорофиллы хорошо растворимы в органических растворителях (этиловом эфире, бензоле, хлороформе, ацетоне, этиловом спирте) и нерастворимы в воде. Хлорофиллы имеют максимумы поглощения света в красной и синей частях спектра. Растворы хлорофиллов обладают флуоресценцией и фосфоресценцией.
5.1.2. Каротиноиды
Каротиноиды – жирорастворимые пигменты, присутствующие в хлоропластах всех растений. Они входят в состав хромопластов в незеленых частях растений, например, корнеплодов моркови. К каротиноидам относят 3 группы соединений: 1) оранжевые или красные каротины, 2) желтые ксантофиллы, 3) каротиноидные кислоты. Каротины и ксантофиллы состоят из 8 остатков изопрена, которые образуют цепь конъюгированных двойных связей (рис. 5.2). Основные каротиноиды - -каротин, лютеин, виолаксантин и неоксантин.
Рис. 5.2. Структурные формулы каротиноидов и последовательность их превращений (по В. В. Полевому).
Каротины и ксантофиллы растворимы в хлороформе, бензоле, сероуглероде, ацетоне. Каротины хорошо растворяются в эфирах, но плохо в спиртах, а ксантофиллы наоборот. Каротиноиды имеют максимумы поглощения в фиолетово-синей и синей частях спектра света. Они не способны к флуоресценции.
Главные функции каротиноидов: поглощение света в качестве дополнительных пигментов, защита молекул хлорофиллов от необратимого фотоокисления, тушение активных радикалов, участие в фототропизме, так как способствуют определению направления роста побега.
5.1.3. Фикобилины
Сине-зеленые и красные водоросли помимо хлорофилла а и каротиноидов содержат пигменты фикобилины. Их молекула состоит из 4 последовательных пиррольных колец (рис. 5.3). Фикобилины являются хромофорными группами глобулиновых белков фикобилипротеинов. Они делятся на 3 группы: 1) фикоэритрины – белки красного цвета, 2) фикоцианины – сине-голубые белки и 3) аллофикоцианины – синие белки. Все они обладают флуоресценцией и растворимы в воде.
Рис. 5.3. Структурные формулы фикобилинов (по В. В. Полевому).
Фикобилины имеют максимумы поглощения в оранжевой, желтой и зеленой частях спектра света. Это позволяет водорослям полнее использовать свет, проникающий в воду. Вода обладает светопоглощающей способностью. На глубине около 30 м полностью исчезают красные лучи, около 180 м – желтые, 320 м – зеленые, а на глубину более 500 м не проникают синие и фиолетовые лучи. Фикобилины – это дополнительные пигменты, участвующие в светособирающем комплексе. Около 90 % энергии света, поглощенного фикобилинами, передается на хлорофилл а.У растений имеется фикобилин фитохром. Он не участвует в фотосинтезе, но является фоторецептором красного и дальнего красного света и выполняет регуляторные функции в клетках растений.
5.2. Световая фаза фотосинтеза
Фотосинтез имеет две фазы – световую и темновую. В световой фазе молекулы пигментов поглощают фотоны, передают поглощенную энергию молекулам хлорофилла а, происходит трансформация энергии света в химическую энергию АТФ и восстановленного НАДФН, выделяется кислород в результате фоторазложения воды. Все эти процессы происходят на мембранах хлоропластов. При темновой фазе фотосинтеза в строме хлоропластов восстанавливается поглощенный СО2 с образованием углеводов и других органических соединений.
5.2.1. Поглощение света и возбуждение хлорофилла
Электрон представляет собой заряженную частицу, которая движется по орбите вокруг ядра атома. Электрон обладает вектором магнитного момента – спином, который отражает направление вращения электрона вокруг своей оси. В стабильном состоянии валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули: два электрона с антипараллельными спинами на каждой орбитали. Такое состояние молекулы называют основным синглетным состоянием S0. При поглощении молекулой пигмента фотона электрон переходит на более высокую орбиталь за счет энергии фотона. Если у возбужденного электрона сохраняется направление спина, то считают, что молекула находится в синглетном возбужденном состоянии S*. Если возбужденный электрон меняет спин, то такое возбужденное состояние называется триплетным Т*.
Поглощение молекулой хлорофилла кванта красного света приводит к синглетному возбужденному состоянию S*1. При поглощении кванта синего цвета с более высоким уровнем энергии электрон переходит на более высокую орбиталь S*2. Время жизни электрона на S*2 составляет не более 10-12 секунды. За такое короткое время энергия электронного возбуждения не может быть использована и электрон возвращается на S*1, потеряв немного энергии в виде тепла. Время жизни электрона в состоянии S*1 равно 10-9. Если молекула пигмента не участвует в фотохимической реакции, то электрон возвращается в стабильное состояние. При этом часть энергии теряется в виде тепла и излучается квант света с большей по сравнению с поглощенным фотоном длиной волны – явление флуоресценции.Время жизни электрона в состоянии Т* равно 10-4-10-2 секунды. Из триплетного возбужденного состояния молекула может вернуться в стабильное состояние, излучив кроме тепла еще более длинноволновый, чем при флуоресценции, квант света. Это слабое свечение называют фосфоресценцией. Если молекула в возбужденном состоянии участвует в фотохимических реакциях с передачей электрона акцептору, то флуоресценции и фосфоресценции не происходит.
Для более полного использования падающего на листья света энергия фотонов улавливается 200-400 молекулами пигментов светособирающего (антенного) комплекса и передается к одной молекуле хлорофилла а, являющейся реакционным центром, которая и участвует в фотохимических реакциях. Передача энергии молекулами пигментов происходит по принципу индуктивного резонанса. Молекула пигмента, поглотившая фотон, создает переменное электрическое поле, которое усиливает колебания электрона в соседней молекуле. Это происходит из-за перекрытия частот колебаний и очень маленького расстояния между молекулами пигментов. Тем более, что при освещении листьев тилакоиды и граны сдвигаются и уплотняются, поэтому хлоропласты уменьшаются в объеме. Передача энергии молекулами пигментов идет с большой эффективностью. Так, от хлорофилла b к хлорофиллу а передается примерно 90 %, а от каротиноидов к хлорофиллу а – 40 % поглощенной энергии.
Совокупность молекул светособирающего комплекса и реакционного центра составляет фотосистему. Предположение о существовании в хлоропластах двух фотосистем высказал Р. Эмерсон в 1957 г, изучая влияние света на квантовый выход фотосинтеза у водоросли хлореллы. Квантовый выход фотосинтеза – это количество выделившегося кислорода или связанного углекислого газа на 1 квант поглощенной энергии. Он установил, что при одновременном освещении хлореллы коротковолновым (650 нм) и длинноволновым (700 нм) красным светом эффект выше, чем суммарный квантовый выход при воздействии красным светом этих длин в отдельности. Это явление получило название эффекта усиления Эмерсона. Позже предположение Р. Эмерсона о наличии двух фотосистем получило экспериментальное подтверждение.5.2.2. Нециклический и циклический транспорт электронов
В фотосистеме II реакционный центр, состоящий из димера хлорофилла а с максимумом поглощения при 680 нм (П680), последовательно поглотив энергию, равную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, передает 2 электрона феофитину. От феофитина электроны последовательно передаются на пластохиноны QA и QB, расположенные на другой стороне мембраны тилакоида, затем на две липидорастворимые молекулы пластохинона PQ. Он является переносчиком протонов и электронов, выполняющего в мембране роль челнока. Принимая 2 электрона и захватывая из стромы 2 Н+, 2 молекулы PQh3 диффундируют через липидную фазу мембраны. Электроны от 2 PQh3 поступают в цепь:
железосерный белок FeSr цитохром f Cu-содержащий белок пластоцианин П700,
а протоны попадают в полость тилакоида.
Вакантные места в П+680 заполняются 2 электронами из содержащего Mn переносчика электронов, который восстанавливается с участием белкового комплекса S. Окисленный комплекс S связывает воду и восстанавливается за счет электронов воды. При фотоокислении воды комплексом S вторая пара протонов освобождается в полость тилакоида, а атом кислорода диффундирует из хлоропласта.
После возбуждения реакционного центра фотосистемы I - молекулы хлорофилла а с максимумом поглощения при 700 нм (П700), 2 квантами длинноволного красного света 2 электрона передаются мономерной форме хлорофилла а – А1 и затем переносчикам электронoв железосерным белкам А2 и АВ, ферредоксину на наружной стороне мембраны тилакоида, НАДФ-редуктазе. Этот фермент восстанавливает НАДФ в НАДФН. На вакантные места в П+700 переходят электроны с пластоцианина и нециклическая цепь транспорта электронов замыкается.В мембранах хлоропластов может функционировать циклический транспорт электронов, когда действует только фотосистема I. Возбужденные молекулы П700 передают электроны на А1, железосерные белки А2 и АВ, переносящие электроны на другую сторону мембраны, ферредоксин, пластохинон, цитохром b6, железосерный белок FeSr, цитохром f, пластоцианин и П700 (рис. 5.4). Энергия, освобождающаяся при транспорте электронов, используется для фосфорилирования АДФ.
Рис. 5.4. Локализация электрон- и протонтранспортных реакций в тилакоидной мембране хлоропласта (по В. В. Полевому).
Механизм фотофосфорилирования АДФ объясняет теория П. Митчелла. Согласно этой теории, трансмембранный перенос пластохинонами электронов и протонов в одну сторону чередуется с переносом цитохромной системой в обратную сторону электронов. Поэтому по одну сторону мембраны накапливается избыток протонов и возникает электрохимический мембранный потенциал. Его энергия используется для синтеза АТФ при разрядке мембраны в результате транспорта протонов через мембрану посредством Н+-АТФазы, которая действует как АТФсинтетаза. Этот фермент выглядит как грибовидная частица на поверхности мембраны тилакоидов. Его молекула состоит из двух частей: головки, выступающей с наружной стороны мембраны и содержащей активный центр фермента, и ножки, погруженной в мембрану и представляющей канал, через который передвигаются протоны.
5.3. Темновая фаза фотосинтеза
5.3.1. С3-путь фотосинтеза или цикл Кальвина
Этот путь ассимиляции СО2 обнаружен у всех фотосинтезирующих растений. Он был назван циклом Кальвина в честь американского биохимика М. Кальвина, который с сотрудниками открыл и изучил его в 1946-1956 годах с помощью метода меченых атомов и хроматографии. Растения разное время подкармливали 14СО2 и хроматографировали экстракты. Радиоактивные изотопы по химическим свойствам не отличаются от стабильных. Принимая участие в реакциях, они помечают те соединения, в которые входят. Цикл состоит из трех этапов: карбоксилирования, восстановления, регенерации первичного акцептора СО2 и синтеза конечного продукта фотосинтеза (рис. 5.5).
Рис. 5.5. Цикл Кальвина (С3-путь фотосинтеза.
1 – фосфорибулокиназа, 2 – рибулозодифосфаткарбоксилаза, 3 - фофсфоглицераткиназа, 4 – триозофосфатдегидрогеназа, 5 – триозофосфатизомераза, 6 – альдолаза, 7 – фосфатаза, 8 – транскетолаза, 9 – альдолаза, 10 – фосфатаза, 11 – транскетолаза, 12 – рибозофосфатизомераза, 13 – фосфокетопентоэпимераза (по В. В. Полевому).
girniy.ru