Клеточная стенка грибов. Входят в состав клеточной стенки растений и грибов
Химический состав и структура клеточной стенки грибов
Клеточная стенка грибов многослойная, при этом разные слои образованы различающимися по химическому составу структурными углеводами, которые по химическому составу можно разделить на 3 группы:
полимеры глюкозы (глюкан, хитин, целлюлоза). Глюканы составляют наружный слой клеточной стенки большинства грибов. Внутренний слой грибной клеточной стенки образован цепочками хитина, придавая ей жесткость. Хитин замещает целлюлозу, которая у большинства грибов отсутствует, но входит в состав клеточной стенки оомицетов, которые в настоящее время к типичным грибам не относятся. Деацетилированный хитин получил название хитозан, который в комплексе с хитином образует клеточную стенку зигомицетов.
полимеры других моносахаридов (маннозы, галактозы и др.) в отличие от высших растений, где они составляют основу матрикса под общим названием гемицеллюлоза, менее характерны для грибов. Исключение составляют дрожжи, в клеточных стенках которых особенно много полимеров маннозы, называемых маннаны. Предполагают, что такой состав стенки лучше обеспечивает почкование.
полимеры углеводов, ковалентно связанные с пептидами (гликопротеины) формируют срединный слой многослойной клеточной стенки и играют важную роль, как в поддержании структурной целостности клеток, так и в ее обменных процессах с окружающей средой.
К другим специфическим особенностям грибной клетки относятся: отсутствие пластид, что сближает ее с клеткой животных;
Отсутствие крахмала, который у эумицетов замещается полисахаридом, близким к животному крахмалу гликогену, у оомицетов – полисахаридом, близкому к ламинарину бурых водорослей. Вырабатывается и целый ряд специфических для грибов запасных углеводов.
Выработка специфических вторичных метаболитов, из которых большую роль играют антибиотики, фито– и микотоксины, фитогормоны.
К специфическим особенностям грибов относятся также гетерокариоз и парасексуальный процесс.
У грибов очень широко распространено явление гетерокариоза или разноядерности, при котором в одной клетке длительное время сохраняются ядра, гетероаллельные по некоторым генам. Гетерокариоз заменяет гаплоидным грибам гетерозиготность и способствует быстрой адаптации грибов к меняющимся условиям. Наличие разноядерности обусловлено рядом специфических особенностей грибов:
1. наличие более одного ядра в клетке
2. специфическое строение клеточной перегородки, в которой имеется одно или несколько сквозных отверстий, называемых порами, через которые ядра могут мигрировать из одной клетки в другую
3. гифы внутри одной колонии и даже разных близко расположенных колоний, выросших из разных спор одного вида гриба часто срастаются, в результате чего возможен обмен ядрами разных штаммов.
Парасексуальный (псевдополовой) процесс. Если в гетерокариотичных клетках сливаются ядра, гетероаллельные по какому-либо локусу, возникает гетерозиготное диплоидное ядро. Оно может попасть в спору и дать начало диплоидному гетерозиготному клону. В процессе митозов диплоидные ядра могут возвращаться в гаплоидное состояние вследствие потери одного набора хромосом или в них может происходить обмен участками хромосом (митотический кроссинговер). Оба процесса сопровождаются рекомбинацией родительских генов и, следовательно, фенотипов. Парасексуальная (бесполовая) рекомбинация очень редкое явление, не превышающее одного ядра на миллион, но вследствие огромного числа ядер в мицелии постоянно наблюдается в популяциях грибов.
Размножение – вегетативное, бесполое, половое.
Вегетативное – фрагментация таллома, образование хламидоспор, которые после периода покоя прорастают в мицелий, почкование у дрожжей.
Бесполое размножение у разных грибов может осуществляться подвижными и неподвижными спорами. Зооспоры образуют относительно небольшая группа грибов и грибоподобных организмов – водные и некоторые наземные, у которых отчетливо прослеживаются генетические связи с водными грибами и водорослями. Структура жгутиков является важным диагностическим признаком при отнесении к определенному царству. У подавляющего большинства грибов при бесполом размножении образуются неподвижные споры, что указывает на их очень давний выход на сушу. По месту формирования и локализации различают эндогенные, образующиеся в спорангиях спорангиоспоры и экзогенные (конидии), развивающиеся на специальных гифах – конидиеносцах. Конидии образуются у большинства грибов (аскомицеты, базидиомицеты, некоторые зигомицеты) составляя конидиальное спороношение, которое отличается огромным многообразием и широко используется для диагностики грибов.
Половое размножение грибов имеет свою специфику как в морфологии полового процесс, так и в механизмах генетической и физиологической регуляции пола и передачи наследственной информации.
Соматогамия – самый распространенный и наиболее простой тип полового процесса, заключается в слиянии двух не дифференцированных на гаметы соматических клеток. Иногда он протекает даже без слияния клеток – сливаются ядра внутри клетки. Встречается у большинства базидиомицетов, сумчатых дрожжей и некоторых др. таксонов.
Гаметангиогамия – на гаплоидном мицелии обособляются гаметангии, при половом процессе сливается их содержимое. Такой половой процесс характерен для большинства сумчатых грибов. Вариантом гаметангиогамии является зигогамия у зигомицетов.
Гаметогамия в виде изо- гетеро и оогамии у грибов встречается значительно реже, чем у других эукариот. Изо- и гетерогамия встречается лишь у хитридиомицетов. Классическая оогамия с образованием сперматозоидов и яйцеклеток у грибов не выражена, а встречаются сильно измененные варианты.
По особенностям регуляции пола у грибов выделяют несколько типов полового процесса
Гинандромиксис можно рассмотреть на примере двудомных оомицетов, у которых оогонии и антеридии развиваются на разных талломах, например фитофтора или картофельный гриб. Если генетически однородный штамм выращивать в монокультуре, то он размножается только бесполым способом. Если рядом оказываются мицелии двух штаммов, то у них прослеживаются морфогенетические изменения под действием химических выделений стероидной природы – половых феромонов. Антеридиол индуцирует образование антеридиев у партнера, а оогониол –оогониев. При этом регуляция пола имеет относительный характер: будет ли штамм образовывать антеридии или же оогонии, зависит от количественного соотношения соответствующих феромонов у него и его партнера. Отсюда название полового процесса – гинандромиксис.
Димиксис или гетероталлизм. Уже давно было подмечено, что грибы могут быть гомо- или гетероталличными. У гомоталличных видов при половом процессе сливаются генетически идентичные ядра внутри мицелия. У гетероталличных видов для прохождения полового цикла необходимо на каком-то этапе (различном у разных видов грибов) слияние потомков спор (точнее, их ядер). Для половой совместимости двух штаммов необходимо генетическое различие (гетероаллельность) в определенных локусах, называемых локусами спаривания. У большинства грибов (зигомицеты, аскомицеты, часть базидиомицетов) имеется один локус спаривания с двумя аллелями. Локус спаривания состоит из нескольких генов, котрые управляют синтезом половых феромонов. Такой гетероталлизм называется однофакторным или биполярным. Потомство таких грибов после мейоза разделяется на две самонесовместимые, но взаимно совместимые группы в соотношении 1:1, т.е. вероятность родственного(инбридинга) и неродственного(аутбридинга) скрещивания составляет как и у двуполых высших эукариот 50%.
В геноме высших базидиальных грибов имеется два локуса спаривания – A и B, причем совместимы друг с другом только штаммы гетероаллельные по обоим локусам (Ax Bx совместим с Ay By, но не с Ax By и Ay Bx). Такой гетероталлизм называют двухфакторным или тетраполярным. Он снижает вероятность инбридинга до 25 %.
Диафоромиксис – у высших базидиомицетов имеется не два, а много аллелей локуса спаривания, случайно встречающихся у разных штаммов, слагающих популяцию. Такой контроль спаривания обеспечивает 100% вероятность аутбридинга, так как штаммы, имеющие разные аллели, взаимносовместимы, а аллелей очень много. В результате образуются панмиксные гибридные популляции.
Жизненные циклы грибов также разнообразны как и сами грибы. Основные циклы, их принадлежность к отделам грибов
1.Бесполый цикл характерен для огромной группы несовершенных или митогрибов, утративших половое размножение. Деление их ядер исключительно митотические. Подавляющее большинство митогрибов относится к сумчатым грибам, но в связи с утратой полового процесса, составляют формальную группу несовершенных грибов или дейтеромицетов.
2. Гаплоидный цикл. Вегетативный таллом несет гаплоидные ядра. После полового процесса (сингамии) диплоидная зигота (обычно после периода покоя) делится мейотически – зиготический мейоз. Характерен для зигомицетов и многих хитридиомицетов.
3. Гаплоидно-дикариотический цикл характеризуется тем, что после слияния содержимого гаметангиев (гаметангиогамия) или соматических клеток гаплоидного мицелия (соматогамия) ядра образуют дикарионы (пары генетически разных ядер). Они синхронно делятся, образуя дикариотический мицелий. Половой процесс завершается слиянием ядер дикариона, образующаяся зигота делится мейозом без периода покоя. Мейоспоры составляют половое спороношение сумчатых и базидиальных грибов в виде аскоспор и базидиоспор. При их прорастании образуется гаплоидный мицелий. У подавляющего большинства сумчатых грибов (кроме дрожжей и тафриновых грибов) в жизненном цикле преобладает гаплоидная фаза в виде вегетативного мицелия (анаморфа), дикариотическая фаза кратковременна и представлена аскогенными гифами, на которых образуются сумки (телеоморфа). У базидиальных грибов в жизненном цикле преобладает дикариотическая фаза, гаплоидная фаза кратковременна.
4. Гаплоидно-диплоидный цикл в виде изоморфной смены поколений у грибов встречается редко (некоторые дрожжи и водные хитридиомицеты).
5. Диплоидный цикл характерен для оомицетов и некоторых сумчатых дрожжей. Вегетативный таллом диплоидный, мейоз гаметический, наблюдается при формировании гаметангиев или гамет.
3. Экологические группы грибов.
Грибы и грибоподобные организмы входят во все наземные и водные экосистемы, как важнейшая часть гетеротрофного блока, вместе с бактериями занимая трофический уровень редуцентов. Широкое распространение грибов в биосфере определяется рядом важнейших особенностей:
1. Наличие у большинства мицелиальной структуры таллома. (позволяет быстро осваивать субстрат, иметь большую поверхность контакта со средой).
2. Большая скорость роста и размножения, позволяющие в короткие сроки заселять большие массы субстрата, образуя огромное число спор и распространять их на большие расстояния.
3. Высокая метаболическая активность, проявляющаяся в широком диапазоне значений экологических факторов.
4. Высокая скорость генетической рекомбинации, значительная биохимическая изменчивость, экологическая пластичность.
5. Способность быстро переходить к состоянию анабиоза, переживать неблагоприятные условия в течение длительного времени.
Главным экологическим фактором для грибов является пищевой субстрат. По отношению к этому фактору выделяют основные группы грибов, которые называются трофическими группами.
1. Сапротрофы – обитают на растительных остатках
4. Краткая характеристика отделов грибов.
Похожие статьи:
poznayka.org
Клеточная стенка | Учеба-Легко.РФ - крупнейший портал по учебе
 Клеточная стенка
Клеточная стенка
Клеточная стенка (оболочка) является неотъемлемым компонентом клеток растений и грибов и представляет собой продукт их жизнедеятельности. Она придает клеткам механическую прочность, защищает их содержимое от повреждений и избыточной потери воды, поддерживает форму клеток и их размер, а также препятствует разрыву клеток в гипотонической среде. Клеточная стенка участвует в поглощении и обмене различных ионов, т. е. является ионообменником. Через клеточную оболочку осуществляется транспорт веществ.
Клеточная стенка, формирующаяся во время деления клеток и их роста путем растяжения, называется первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои, и образуется прочная вторичнаяклеточная оболочка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений и хитин у грибов), компоненты матрикса (гемицеллюлоза, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности оболочки (кутин и воск).
Молекулы целлюлозы за счет водородных связей объединяются в пучки —микрофибриллы. Переплетенные микрофибриллы составляют каркас клеточной оболочки. У большинства грибов микрофибриллы клеточной стенки состоят из хитина.
Микрофибриллы погружены в матрикс клеточной стенки. Матрикс состоит из смеси различных химических веществ, среди которых преобладают полисахариды (гемицеллюлозы и пектиновые вещества).
Гемицеллюлозы — это группа полисахаридов (полимеры пен-тоз и гексоз — ксилозы, галактозы, маннозы, глюкозы и др.). Молекулы гемицеллюлоз, как и целлюлозы, имеют форму цепи, но в отличие от последней их цепи короче, менее упорядочены и сильно разветвлены. Они легче растворяются и разрушаются ферментами.
Пектиновые вещества — это полимеры, построенные из моносахаридов (арабинозы и галактозы), галактуроновой кислоты (сахарной кислоты) и метилового спирта. Длинные молекулы пектиновых веществ могут быть линейны ми или разветвленными. Молекулы пектиновых веществ содержат большое количество карбоксильных групп и поэтому способны соединяться с ионами Mg2+ и Са2-. При этом образуются клейкие, студнеобразные пектаты магния и кальция, из которых затем складываются срединные пластинки, скрепляющие стенки двух соседних клеток.
Ионы двухвалентных металлов могут обмениваться на другие катионы (Н-, К+ и т. д.). Это обусловливает катионообменную способность клеточных оболочек.
Пектиновыми веществами и пектатами богаты оболочки клеток многих плодов. Так как при их извлечении из оболочек и добавлении сахара образуются гели, пектины используют как желе-образующие вещества для изготовления мармелада и др.
Помимо углеводных компонентов, в состав матрикса клеточной стенки входит структурный белок экстенеин —гликонроте-ин, который по своему составу близок к межклеточным белкам животных —коллагенам.
На долю матрикса приходится до 60% сухого вещества клеточной оболочки. Матрикс оболочки не просто заполняет промежутки между микрофибриллами, а образует прочные химические (водородные и ковалентные) связи между макромолекулами и микрофибриллами, что обеспечивает прочность клеточной стенки, ее эластичность и пластичность.
Основным инкрустирующим веществом оболочки клеток растений является лигнин — полимер с неразветвленной молекулой, состоящей из ароматических спиртов.
Интенсивная лигнификация (пропитка слоев целлюлозы лигнином) клеточных оболочек начинается после прекращения роста клетки. Лигнин может откладываться отдельными участками — в виде колец, спиралей или сетки, как это наблюдается в оболочках клеток проводящей ткани — ксилемы, или сплошным слоем, за исключением тех мест, где осуществляются контакты между соседними клетками в виде плазмодесм.
Лигнин скрепляет целлюлозные волокна и действует как очень твердый и жесткий каркас, усиливающий прочность клеточных стенок на растяжение и сжатие. Он же обеспечивает клеткам дополнительную защиту от физических и химических воздействий, снижает водопроницаемость. Содержание лигнина в оболочке достигает 30%. Инкрустация им клеточных оболочек приводит к иходревеснению, которое часто влечет за собой отмирание живого содержимого клетки.
Лигнин в сочетании с целлюлозой придает особые свойства древесине, которые делают ее незаменимым строительным материалом.
На клеточную оболочку могут откладываться также жиропо-добные вещества — суберин, кутин и воск.
Суберин откладывается на оболочку изнутри и делает ее практически непроницаемой для воды и растворов. В результате протопласт клетки отмирает и клетка заполняется воздухом. Такой процесс называется опробковением. Наблюдается опробковение оболочки клеток в покровных тканях многолетних древесных растений — перидерме, корке, а также в эндодерме корня.
Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками. Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами (целлюлозой и пектином) и образует кутикулу. Воск часто откладывается в кристаллической форме на поверхности частей растений (листьев, плодов), образуя восковой налет.
Кутикула и восковой налет защищают клетки от повреждений и проникновения инфекции, уменьшают испарение воды с поверхности органов.
В оболочках эпидермальных клеток некоторых растений (злаков, осок и др.) накапливается большое количество минеральных веществ (минерализация), в первую очередь карбоната кальция и кремнезема. При минерализации листья и стебли растений становятся жесткими, твердыми и в меньшей степени поедаются животными.
Таким образом, клеточная стенка играет важную роль в жизни клеток растений и грибов и выполняет ряд специфических функций.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"
uclg.ru
Клеточная стенка грибов
из чего состоит клеточная стенка гриба?
- из хитина
- Клеточные стенки грибов состоят из хитина и глюканов
- из углевода хитина
- Самым распространенным клеточным покрытием у грибов является клеточная стенка. Ее основу составляют микрофибриллярные скелетные компоненты – хитин или целлюлоза. Над ними располагается наружный аморфный матрикс образованный глюканом, хитозаном или мананом. С микрофибриллярными и аморфными комп. повязываются различные белки, липиды, а также пигменты (меланин, хитоны), растворимые сахара, аминокислоты, разл. ионы и соли.
Клеточная стенка бактерий
специфическая по хим. составу оболочка, окружающая протопласт и тесно связанная структурно–функциональными взаимоотношениями с цитоплазматической мембраной. Толщина К. с. – 150 нм; составляет 10—5°% сухой массы клеток. У большинства бактерий в состав К. с. входит структурный полимер пептидогликан (муреин), у грамположительных бактерий он может составлять до 95 % ее массы. В качестве матрикса в состав К. с. этих бактерий входят тейхоевые и тейхуроновые кислоты, полисахариды и белки. Микобактерии в составе К. с. содержат липиды. Полисахариды и тейхоевые кислоты обладают антигенными свойствами. К. с. грамотрицательных бактерий содержит более тонкий пептидогликановый слой (толщина 3–8 нм, 5—10 % от сухой массы К. с.). Кроме одинарной муреиновой сети, она имеет снаружи трехслойную липопротеидную мембрану, компоненты которой обусловливают антигенные свойства клетки. Муреин К. с. может быть разрушен лизоцимом или другими агентами, что приводит к образованию сферопластов и протопластов. Микоплазмы полностью лишены К. с. К. с. выполняет защитную, опорную функции, придает клеткам определенную форму, а у грамотрицательных бактерий вместе с цито–плазматической мембраной обеспечивает избирательную проницаемость для веществ, поступающих внутрь клетки. В отличие от бактерий (эубактерий) археи (архебактерии) содержат в К. с. иной структурный компонент, получивший название псевдомуреин. У многих архей К. с. образована белковыми глобулами. Поэтому на археи не действуют антибиотики, подавляющие синтез муреиновой клеточной стенки эубактерий (пенициллин, ванкомицин и др.).
Отличия растений, животных и грибов
Надцарство эукариот делится на три царства: растений, животных и грибов.
1) Растения – автотрофы, т.е. делают себе питательные вещества сами в процессе фотосинтеза. Животные и грибы гетеротрофы, т.е. питательные вещества берут из пищи.
2) Животные могут передвигаться, а растения и грибы – нет. Зато растения и грибы растут всю жизнь, а животные – только до того, как начнут размножаться.
3) Только у растений в клетках есть хлоропласты. Только у животных нет твердой клеточной стенки. У растений запасной углевод – крахмал, а у животных и грибов – гликоген.
Тесты
1. Выберите признаки растений
а) автотрофы, могут расти всю жизнь
б) автотрофы, растут только до начала размножения
в) гетеротрофы, могут расти всю жизнь
г) гетеротрофы, растут только до начала размножения
2. Выберите признаки животных
а) автотрофы, могут расти всю жизнь
б) автотрофы, растут только до начала размножения
в) гетеротрофы, могут расти всю жизнь
г) гетеротрофы, растут только до начала размножения
3. Выберите признаки грибов
а) автотрофы, могут расти всю жизнь
б) автотрофы, растут только до начала размножения
в) гетеротрофы, могут расти всю жизнь
г) гетеротрофы, растут только до начала размножения
4. Выберите признаки грибов
а) есть твердая клеточная стенка, запасное вещество гликоген
б) нет твердой клеточной стенки, запасное вещество гликоген
в) есть твердая клеточная стенка, запасное вещество крахмал
г) нет твердой клеточной стенки, запасное вещество крахмал
5. Выберите признаки растений
а) есть твердая клеточная стенка, запасное вещество гликоген
б) нет твердой клеточной стенки, запасное вещество гликоген
в) есть твердая клеточная стенка, запасное вещество крахмал
г) нет твердой клеточной стенки, запасное вещество крахмал
6. Выберите признаки животных
а) есть твердая клеточная стенка, запасное вещество гликоген
б) нет твердой клеточной стенки, запасное вещество гликоген
в) есть твердая клеточная стенка, запасное вещество крахмал
г) нет твердой клеточной стенки, запасное вещество крахмал
Грамположительные бактерии. клеточная стенка грамположительных бактерий. строение клеточной стенки грамположительных бактерий. аутолизины бактерий. сферопласты. протопласты.
Оглавление темы "Анатомия бактериальной клетки. Физиология бактерий.":
1. Анатомия бактериальной клетки. Поверхностные структуры бактерии. Капсула бактерий. Организация капсул. Окраска капсул бактерий. Состав капсул. Антигенные свойства капсул.
2. Жгутики бактерий. Расположение жгутиков. Перитрихи. Монотрихи. Политрихи. Лофотрихи. Амфитрихи. Феномен роения. Диагностика подвижности бактерий.
3. Микроворсинки бактерий. Фимбрии бактерий. F-пили ( секс-пили ) бактерии. Клеточная оболочка бактерий. Гликокаликс.
4. Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
5. Грамотрицательные бактерии. Клеточная стенка грамотрицательных бактерий. Строение клеточной стенки грамотрицательных бактерий.
6. Грамположительные бактерии. Клеточная стенка грамположительных бактерий. Строение клеточной стенки грамположительных бактерий. Аутолизины бактерий. Сферопласты. Протопласты.
7. Цитоплазматическая мембрана ( ЦПМ ) бактерии. Состав цитоплазматической мембраны бактерий. Транспортные системы. Мезосомы. Периплазматическое пространство.
8. Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
9. Физиология бактерий. Питание бактерий. Тип питания бактерии. Голозои. Голофиты. Вода. Значимость воды для бактерий.
10. Усваиваемые бактериальной клеткой соединения. Пути поступления веществ в бактериальную клетку. Пассивный перенос. Диффузия.
Грамположительные бактерии. Клеточная стенка грамположительных бактерий. Строение клеточной стенки грамположительных бактерий. Аутолизины бактерий. Сферопласты. Протопласты.
Грамположительные бактерии имеют сравнительно просто организованную, но мощную клеточную стенку. Она состоит преимущественно из множества слоев пептидогликана, составляющего до 90% её сухой массы (см. рис. 4-4), часто включающих вместо диаминопимелиновой кислоты лизин и уникальные водорастворимые полимеры тейхоевых кислот, состоящих из 8-50 остатков глицерина или рибита, связанных между собой фосфодиэфирными связями. Тейхоевые кислоты [от греч. teichos, стенка] могут составлять до 50% сухого веса клеточной стенки. Известно два типа кислот — рибиттейхоевые (состоят из остатков рибитфосфата и 10-50 остатков спирта) и глицеринтейхоевые (состоят из остатков глицерофосфата и 20 остатков спирта). Клеточная стенка каждого вида содержит только один тип теихоевых кислот (за исключением вида Streptomyces). Тейхоевые кислоты — основные поверхностные Аг многих бактерий. У большей части грамноложительных бактерий также имеются периплазматические теихоевые кислоты, располагающиеся между клеточной стенкой и ЦПМ. Периплазматические кислоты, выявляемые горячей кислотной экстракцией, являются группоспецифичными Аг.
Клеточная стенка грамположительных бактерий не содержит ЛПС, но может включать различные белки. Содержание последних весьма вариабельно. Для некоторых бактерий (например, стрептококков группы А) белки служат серовароспецифичными Аг.
Аутолизины бактерий. Клеточные стенки бактерий содержат аутолизины — ферменты, растворяющие пептидогликановый слой. Их активность необходима для процессов роста клеточной стенки, деления клеток, споруляции и достижения состояния компетентности при трансформации (см. ниже).
Удаление клеточной стенки, защищающей прилежащую ЦПМ, приводит к лизису бактерии либо к образованию протопластов или сферопластов (из грамположительных или грамотрицательных бактерий соответственно). Бактерии, лишённые клеточной стенки, лишь в изотонической среде способны поглощать 02 и выделять С02, а также размножаться.
Сферопласты более устойчивы к изменениям осмотического давления и длительно сохраняются в неизотонической среде.
- Читать далее "Цитоплазматическая мембрана ( ЦПМ ) бактерии. Состав цитоплазматической мембраны бактерий. Транспортные системы. Мезосомы. Периплазматическое пространство."
Клеточная стенка
Клеточная стенка (оболочка) является неотъемлемымкомпонентом клеток растений и грибов и представляет собойпродукт их жизнедеятельности. Она придает клеткам механическуюпрочность, защищает их содержимое от повреждений и избыточнойпотери воды, поддерживает форму клеток и их размер, а такжепрепятствует разрыву клеток в гипотонической среде. Клеточнаястенка участвует в поглощении и обмене различных ионов, т. е.является ионообменником. Через клеточную оболочкуосуществляется транспорт веществ.
Клеточная стенка, формирующаяся во время деления клеток и ихроста путем растяжения, называется первичной. Послепрекращения роста клетки на первичную клеточную стенку изнутриоткладываются новые слои, и образуется прочнаявторичная клеточная оболочка.
В состав клеточной стенки входят структурные компоненты(целлюлоза у растений и хитин у грибов), компоненты матрикса(гемицеллюлоза, пектин, белки), инкрустирующие компоненты(лигнин, суберин) и вещества, откладывающиеся на поверхностиоболочки (кутин и воск).
Молекулы целлюлозы за счет водородных связей объединяются впучки —микрофибриллы. Переплетенные микрофибриллысоставляют каркас клеточной оболочки. У большинства грибовмикрофибриллы клеточной стенки состоят из хитина.
Микрофибриллы погружены в матрикс клеточной стенки. Матрикссостоит из смеси различных химических веществ, среди которыхпреобладают полисахариды (гемицеллюлозы и пектиновыевещества).
Гемицеллюлозы — это группа полисахаридов (полимерыпен-тоз и гексоз — ксилозы, галактозы, маннозы, глюкозы идр.). Молекулы гемицеллюлоз, как и целлюлозы, имеют формуцепи, но в отличие от последней их цепи короче, менееупорядочены и сильно разветвлены. Они легче растворяются иразрушаются ферментами.
Пектиновые вещества — это полимеры, построенные измоносахаридов (арабинозы и галактозы), галактуроновой кислоты(сахарной кислоты) и метилового спирта. Длинные молекулыпектиновых веществ могут быть линейны ми или разветвленными.Молекулы пектиновых веществ содержат большое количествокарбоксильных групп и поэтому способны соединяться с ионамиMg2+ и Са2-. При этом образуютсяклейкие, студнеобразные пектаты магния и кальция, из которыхзатем складываются срединные пластинки, скрепляющие стенкидвух соседних клеток.
Ионы двухвалентных металлов могут обмениваться на другиекатионы (Н-, К+ и т. д.). Этообусловливает катионообменную способность клеточных оболочек.
Пектиновыми веществами и пектатами богаты оболочки клетокмногих плодов. Так как при их извлечении из оболочек идобавлении сахара образуются гели, пектины используют какжеле-образующие вещества для изготовления мармелада и др.
Помимо углеводных компонентов, в состав матрикса клеточнойстенки входит структурный белок экстенеин —гликонроте-ин,который по своему составу близок к межклеточным белкамживотных —коллагенам.
На долю матрикса приходится до 60% сухого вещества клеточнойоболочки. Матрикс оболочки не просто заполняет промежуткимежду микрофибриллами, а образует прочные химические(водородные и ковалентные) связи между макромолекулами имикрофибриллами, что обеспечивает прочность клеточной стенки,ее эластичность и пластичность.
Основным инкрустирующим веществом оболочки клеток растенийявляется лигнин — полимер с неразветвленной молекулой,состоящей из ароматических спиртов.
Интенсивная лигнификация (пропитка слоев целлюлозы лигнином)клеточных оболочек начинается после прекращения роста клетки.Лигнин может откладываться отдельными участками — в видеколец, спиралей или сетки, как это наблюдается в оболочкахклеток проводящей ткани — ксилемы, или сплошным слоем, заисключением тех мест, где осуществляются контакты междусоседними клетками в виде плазмодесм.
Лигнин скрепляет целлюлозные волокна и действует как оченьтвердый и жесткий каркас, усиливающий прочность клеточныхстенок на растяжение и сжатие. Он же обеспечивает клеткамдополнительную защиту от физических и химических воздействий,снижает водопроницаемость. Содержание лигнина в оболочкедостигает 30%. Инкрустация им клеточных оболочек приводит к иходревеснению, которое часто влечет за собой отмираниеживого содержимого клетки.
Лигнин в сочетании с целлюлозой придает особые свойствадревесине, которые делают ее незаменимым строительнымматериалом.
На клеточную оболочку могут откладываться также жиропо-добныевещества — суберин, кутин и воск.
Суберин откладывается на оболочку изнутри и делает еепрактически непроницаемой для воды и растворов. В результатепротопласт клетки отмирает и клетка заполняется воздухом.Такой процесс называется опробковением. Наблюдаетсяопробковение оболочки клеток в покровных тканях многолетнихдревесных растений — перидерме, корке, а также в эндодермекорня.
Поверхность эпидермальных клеток растений защищенагидрофобными веществами — кутином и восками.Предшественники этих соединений секретируются из цитоплазмы наповерхность, где и происходит их полимеризация. Слой кутинаобычно пронизан полисахаридными компонентами (целлюлозой ипектином) и образует кутикулу. Воск часто откладываетсяв кристаллической форме на поверхности частей растений(листьев, плодов), образуя восковой налет.
Кутикула и восковой налет защищают клетки от повреждений ипроникновения инфекции, уменьшают испарение воды с поверхностиорганов.
В оболочках эпидермальных клеток некоторых растений (злаков,осок и др.) накапливается большое количество минеральныхвеществ (минерализация), в первую очередь карбонатакальция и кремнезема. При минерализации листья и стеблирастений становятся жесткими, твердыми и в меньшей степенипоедаются животными.
Таким образом, клеточная стенка играет важную роль в жизниклеток растений и грибов и выполняет ряд специфическихфункций.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов"Пособие по биологии для поступающих в ВУЗы"
Сравнительная характеристика моносахаридного состава полисахаридов клеточной стенки гриба aspergillus niger и семян пшеницы triticum aestivum
* От исходного веса сухих семян. ** Весовые проценты.*** сл. — Следовое количество.
При экстракциях семян пшеницы было получено пять полисахаридных фракций (ПШ-1 — ПШ-5), выход которых варьирует в достаточно широком диапазоне — от 0,3 до 5,1 %. Суммарный выход полисахаридов составляет 9,2 %.
Во фракции ПШ-1, которая получена в результате экстракции семян пшеницы водой при 20 °С, доминирующими моносахаридами являются остатки ксилозы (7,0 %) и арабинозы (6,2 %). В составе полисахаридной фракции ПШ-2, полученной при экстракции семян водой при 70 °С, главными моносахаридными компонентами являются остатки ксилозы (12,9 %), арабинозы (11,3 %) и глюкозы (8,8 %). В обеих водных фракциях отмечено высокое содержание белка — 36,5 и 26,5 % соответственно, присутствуют гликуроновые кислоты — 13,8 и 3,5 % соответственно. По-видимому, можно предположить, что фракции ПШ-1 и ПШ-2могут представлять собой водорастворимые арабиноксиланы, арабиногликуроноксиланы или ксилоглюканы.
В результате дальнейшей обработки сырья водным раствором соляной кислоты получена фракция ПШ-3, в которой среди моносахаридов доминируют остатки глюкозы (65,5 %). Остатки ксилозы, арабинозы, галактозы, маннозы, рамнозы присутствуют в небольших количествах. Содержание гликуроновых кислот составляет 17,5 %. В составе фракции ПШ-4,выделенной с помощью экстракции сырья водным раствором оксалата аммония, среди нейтральных моносахаридов остатки глюкозы (11,9 %) являются доминирующими. При дальнейшей экстракции семян пшеницы раствором щелочи получили полисахаридную фракцию ПШ-5с высоким содержанием остатков глюкозы (51,6 %). Вероятно, полисахаридные фракции ПШ-3, ПШ-4 и ПШ-5 представляют собой глюканы, которые входят в состав гемицеллюлоз злаков.
Таким образом, в зернах пшеницы обнаружены полисахариды гемицеллюлозы, которые включают в себя арабиноксиланы, арабиногликуроноксиланы, ксилоглюканы и глюканы.
В заключение, в составе полисахаридных фракций, экстрагированных из мицелия гриба Aspergillusniger водой и растворами щелочи, преобладают в разных соотношениях три нейтральных моносахарида — глюкоза (до 44,7 %), манноза (до 28,5 %) и галактоза (до 15,6 %). В зернах пшеницы Triticumaestivum в составе полисахаридов, выделенных разными экстрагентами, доминирует другой спектр моносахаридов, а именно: глюкоза (до 65,5 %), ксилоза (до 12,7 %) и арабиноза (до 8,5 %). Указанные отличия в моносахаридном составе полисахаридов гриба и пшеницы, по-видимому, отражают разный состав и структуру доминирующих полисахаридов грибов и растений.
Список литературы:
- Андриянова Д.А., Смирнова Г.П., Шашков А.С., Чижов А.О., Галанина Л.А., Феофилова Е.П., Усов А.И. Полисахаридный состав мицелия и клеточных стенок гриба Penicilliumroqueforti // Биоорг. химия. — 2011. — Т. 37, № 3. — С. 399—407.
- Билай В.И. Основы общей микологии. Киев: Выща школа, 1989. — 392 с.
- Кочетков Н.К., Бочков А.Ф., Дмитриев Б.А., Усов А.И., Чижов О.С., Шибаев В.Н. Химия углеводов. М.: Химия, 1967. — 674 с.
- Усов А.И., Бланко Ф.Ф., Иванова В.С., Бедрина Е.Н., Фирсова С.А., Седакова Л.А., Фунтикова Н.С. Строение и противоопухолевая активность полисахаридов из мицелияAspergillusoryzae //Биоорг. химия. — 1991. — Т. 17, № 1. — С. 121—125.
- Феофилова Е.П. Клеточная стенка грибов: современные представления о составе и биологической функции // Микробиология. — 2010. — Т. 79, № 6. — С. 723—733.
- Феофилова Е.П., Немцев Д.В., Терешина В.М., Меморская А.С. Состав и содержание хитин-глюканового комплекса в онтогенезе гриба Aspergillusniger // Прикл. биохим. микробиол. —2006.Т. 42, № 6. — С. 624—628.
- Шубаков А.А., Донцов А.Г., Елькина Е.А. Выделение полигалактуроназ из ферментного препарата пектофоетидин Г3х // Бутлеровские сообщения. — 2009. — Т. 18, № 7. — С. 63—68.
- Шубаков А.А., Елькина Е.А. Продуцирование полигалактуроназ мицелиальными грибами Aspergillusniger ВКМ F-1119 и Penicilliumdierckxii ВКПМ F-152 // Химия и компьютерное моделирование. Бутлеровские сообщения. — 2002. — № 7. — С. 65—68.
- Farkas V. Biosynthesis of cell walls of fungi // Microbiol. Rev. — 1979. —Vol. 43, N 2. — P. 117—144.
- Fontaine T., Simenel C., Dubreucq G., Adam O., Delepierre M., Lemoine J., Vorgias C.E., Diaquin M., Latge J.-P. Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall // J. Biol. Chem. — 2000. — Vol. 275, N 8. — P. 27594—27607.
- Schuster E., Dunn-Coleman N., Frisvad J.C., Van Dijck P.W.M. On the safety of Aspergillus niger — a review // Appl. Microbiol. Biotechnol. — 2002. — Vol. 59. — P. 426—435.
- Wasser S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides // Appl. Microbiol. Biotechnol. —2002. — Vol. 60. — P. 258—274.
Клеточная стенка — википедия
- 1 Клеточные стенки прокариот
- 2 Клеточные стенки грибов
- 3 Клеточные стенки водорослей
- 4 Клеточные стенки высших растений
- 5 См. также
Клеточные стенки прокариот[править]
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми и липотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов[править]
Клеточные стенки грибов состоят из хитина и глюканов.
Клеточные стенки водорослей[править]
Большинство водорослей имеют клеточную стенку из целлюлозы и различных гликопротеинов. Включения дополнительных полисахаридов имеют большое таксономическое значение.
Диатомовые водоросли синтезируют свою клеточную стенку из кремнезёма.
Клеточные стенки высших растений[править]
Клеточные стенки высших растений построены в основном из целлюлозы, гемицеллюлозы и пектина. В них существуют углубления — поры, через которые проходят плазмодесмы, осуществляющие контакт соседних клеток и обмен веществами между ними. Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жесткость клетки для структурной и механической поддержки, придают форму клетке, направление её роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургору, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают от патогенов, проникающих из окружающей среды, и запасают углеводы для растения. Растительные клеточные стенки строятся прежде всего из углеводного полимера целлюлозы.
См. также[править]
- Метод Грама
- Метод Пешкова
Это заготовка статьи по цитологии. Вы можете помочь проекту, исправив и дополнив её.Это заготовка статьи по бактериологии. Вы можете помочь проекту, исправив и дополнив её.
Еще записи по теме
actibo.ru
Клеточные стенки грибов
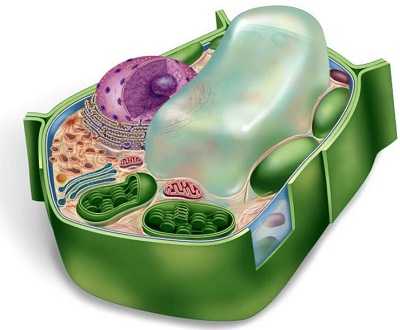
Прокариотическая клетка
Прокариоты — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариотическая клетка
Эукариоты — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов(эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
2) цитоплазма
Цитопла́зма — внутренняя среда живой или умершей клетки, кроме ядра и вакуоли, ограниченная плазматической мембраной. Включает в себя гиалоплазму — основное прозрачное вещество цитоплазмы, находящиеся в ней обязательные клеточные компоненты — органеллы, а также различные непостоянные структуры — включения.
В состав цитоплазмы входят все виды органических и неорганических веществ. В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы — вода.
Цитоплазма постоянно движется, перетекает внутри живой клетки, перемещая вместе с собой различные вещества, включения и органоиды. Это движение называется циклозом. В ней протекают все процессы обмена веществ.
Цитоплазма способна к росту и воспроизведению и при частичном удалении может восстановиться. Однако нормально функционирует цитоплазма только в присутствии ядра. Без него долго существовать цитоплазма не может, так же как и ядро без цитоплазмы.
Важнейшая роль цитоплазмы заключается в объединении всех клеточных структур (компонентов) и обеспечении их химического взаимодействия. Так же цитоплазма поддерживает тургор(объём) клетки, поддержание температуры.
3)Клеточная стенка
Клеточная стенка — жёсткая оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшиене имеют клеточной стенки.
Клеточные стенки прокариот
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми илипотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов
Клеточные стенки грибов состоят из хитина и глюканов.
studopedya.ru
Введение в биологию (VIa) - caenogenesis
Все углеводы делятся на моносахариды (простые сахара), олигосахариды (цепочки, содержащие от 2 до 10 моносахаридных остатков) и полисахариды (полимеры, в которых число моносахаридных остатков может достигать многих тысяч). Один из самых известных полисахаридов - крахмал, представляющий собой длинную цепь остатков глюкозы, соединенных гликозидными связями. Это важнейшее запасное вещество у растений.
 Животный аналог крахмала - гликоген, тоже важный запасной углевод. У нас он накапливается в первую очередь в печени и в случае надобности быстро расщепляется до мономеров глюкозы, которые уходят в кровь. Гликоген тоже состоит из остатков глюкозы, соединенных гликозидными связями. Серьезное отличие гликозидных связей, например, от пептидных - в том, что образованный с их помощью полимер может гораздо легче ветвиться. "По умолчанию" гликозидная связь образуется между гидроксилами 1-го и 4-го атомов углерода глюкозы (1-4-гликозидная связь), и тогде получается линейная цепочка. Но в глюкозе есть и другие гидроксилы, между которыми образование гликозидной связи тоже запросто возможно. На 1-6-гликозидной связи полимерная цепочка обычно как раз и разветвляется. В гликогене такое ветвление выражено сильнее, чем в крахмале, хотя оно есть и там и там.
Животный аналог крахмала - гликоген, тоже важный запасной углевод. У нас он накапливается в первую очередь в печени и в случае надобности быстро расщепляется до мономеров глюкозы, которые уходят в кровь. Гликоген тоже состоит из остатков глюкозы, соединенных гликозидными связями. Серьезное отличие гликозидных связей, например, от пептидных - в том, что образованный с их помощью полимер может гораздо легче ветвиться. "По умолчанию" гликозидная связь образуется между гидроксилами 1-го и 4-го атомов углерода глюкозы (1-4-гликозидная связь), и тогде получается линейная цепочка. Но в глюкозе есть и другие гидроксилы, между которыми образование гликозидной связи тоже запросто возможно. На 1-6-гликозидной связи полимерная цепочка обычно как раз и разветвляется. В гликогене такое ветвление выражено сильнее, чем в крахмале, хотя оно есть и там и там.
 Цвета на этой картинке, на самом деле, никакого значения сейчас не имеют, она просто красивая. Это - структура гликогена. Зеленым тут обозначен остаток глюкозы, с которого начинается боковая цепь, красным - концевые остатки, ну а все остальное нам сейчас уже должно быть понятно и так. Совершенно особый интерес представляют полисахариды, участвующие в образовании клеточных стенок. Ни в коем случае нельзя путать клеточную стенку с клеточной мембраной! Клеточная стенка - это внеклеточная структура, состоящая из полимеров, расположенная снаружи от мембраны и заключающая в себе клетку целиком (не считая отверстий, обеспечивающих межклеточные контакты, если организм многоклеточный). Клеточная стенка может состоять из целлюлозы (у растений), из хитина (у грибов), из сложных полимеров, в состав которых входят углеводы и аминокислоты (у бактерий) или из белков (у архей). У некоторых организмов, например у животных, клеточных стенок нет вообще - это позволяет их клеткам легко менять форму.
Цвета на этой картинке, на самом деле, никакого значения сейчас не имеют, она просто красивая. Это - структура гликогена. Зеленым тут обозначен остаток глюкозы, с которого начинается боковая цепь, красным - концевые остатки, ну а все остальное нам сейчас уже должно быть понятно и так. Совершенно особый интерес представляют полисахариды, участвующие в образовании клеточных стенок. Ни в коем случае нельзя путать клеточную стенку с клеточной мембраной! Клеточная стенка - это внеклеточная структура, состоящая из полимеров, расположенная снаружи от мембраны и заключающая в себе клетку целиком (не считая отверстий, обеспечивающих межклеточные контакты, если организм многоклеточный). Клеточная стенка может состоять из целлюлозы (у растений), из хитина (у грибов), из сложных полимеров, в состав которых входят углеводы и аминокислоты (у бактерий) или из белков (у архей). У некоторых организмов, например у животных, клеточных стенок нет вообще - это позволяет их клеткам легко менять форму.
 Основной компонент клеточных стенок растений - целлюлоза - это полимер глюкозы, так же как и крахмал. Но, в отличие от крахмала, она состоит не из α-глюкозы, а из β-глюкозы. Кроме того, молекулы целлюлозы не ветвятся. Образующиеся между остатками β-глюкозы β-гликозидные связи - на схеме молекулы целлюлозы они выглядят зигзагообразными - гораздо прочнее α-гликозидных и расщепляются только очень немногими ферментами. Например, никто из животных, питающихся растениями, не может самостоятельно переваривать целлюлозу; тем, кто берется ее усваивать, приходится заводить для этой цели симбионтов-бактерий, у которых есть нужный фермент - целлюлаза (Гиляров, 2008).
Основной компонент клеточных стенок растений - целлюлоза - это полимер глюкозы, так же как и крахмал. Но, в отличие от крахмала, она состоит не из α-глюкозы, а из β-глюкозы. Кроме того, молекулы целлюлозы не ветвятся. Образующиеся между остатками β-глюкозы β-гликозидные связи - на схеме молекулы целлюлозы они выглядят зигзагообразными - гораздо прочнее α-гликозидных и расщепляются только очень немногими ферментами. Например, никто из животных, питающихся растениями, не может самостоятельно переваривать целлюлозу; тем, кто берется ее усваивать, приходится заводить для этой цели симбионтов-бактерий, у которых есть нужный фермент - целлюлаза (Гиляров, 2008).

Растительная клеточная стенка может быть гораздо толще мембраны. Если растение многоклеточное, то между клетками обычно есть плазмодесмы - проходящие сквозь отверстия в клеточных стенках цитоплазматические мостики (цитоплазмой называется все внутреннее содержимое клетки, кроме ядра). Через плазмодесмы растительные клетки общаются и обмениваются разными веществами. На самом деле клеточная стенка растений вовсе не состоит из чистой целлюлозы. Во-первых, в нее еще входят короткие ветвящиеся полимеры, включающие не только глюкозу, но и другие моносахариды (эти полимеры собирательно называются гемицеллюлозами), а во-вторых - некоторые структурные белки. Целлюлоза вместе с гемицеллюлозами и белками образует сложную сеть, усиленную к тому же водородными связями - между длинными молекулами целлюлозы, в которых много гидроксильных групп, они возникают очень легко.
 С точки зрения жизни на Земле в целом самая интересная составляющая клеточной стенки растений - это лигнин. Он не имеет никакой общей формулы. Лигнин - сложный полимер, сшитый из нескольких разновидностей спиртов с ароматическими ядрами и углеводородными цепочками. Все мономеры лигнина синтезируются из аминокислоты фенилаланина, которая превращается сначала в коричную кислоту - вещество, входящее в состав масла корицы, - а потом в разнообразные спирты (на схеме показаны только два из них):
С точки зрения жизни на Земле в целом самая интересная составляющая клеточной стенки растений - это лигнин. Он не имеет никакой общей формулы. Лигнин - сложный полимер, сшитый из нескольких разновидностей спиртов с ароматическими ядрами и углеводородными цепочками. Все мономеры лигнина синтезируются из аминокислоты фенилаланина, которая превращается сначала в коричную кислоту - вещество, входящее в состав масла корицы, - а потом в разнообразные спирты (на схеме показаны только два из них):
 Образование лигнина - признак сосудистых растений, то есть папоротников, плаунов, хвощей, хвойных и цветковых. Это эволюционное "изобретение", сделанное только после выхода растений на сушу, и то далеко не сразу. Дело в том, что лигнин придает клеточным стенкам огромную механическую прочность. Он необходим, чтобы сделать ствол наземного растения высоким, вплоть до многометрового, и создать транспортную систему из микроскопических трубочек, качающую воду на всю эту высоту. Именно с "изобретением" биосинтеза лигнина связано одно из крупнейших событий, поменявших лик Земли - появление лесов (Еськов, 2000). Кроме того, появление лигнина сильно изменило глобальный круговорот углерода. Тут дело в том, что лигнин с его разнообразными мономерами и перепутанными химическими связями исключительно неподатлив к действию ферментов. Поэтому растительной тканью, в которой много лигнина, почти невозможно питаться. Из всех земных живых организмов эффективно разлагать лигнин "научились" только грибы, причем не все и не сразу (Robinson, 1990). Именно они и стали разрушителями мертвых деревьев. До этого вся огромная биомасса лигнифицированной древесины просто захоранивалась как есть, создавая залежи каменного угля, в честь которых получил название целый геологический период - каменноугольный, или карбон.
Образование лигнина - признак сосудистых растений, то есть папоротников, плаунов, хвощей, хвойных и цветковых. Это эволюционное "изобретение", сделанное только после выхода растений на сушу, и то далеко не сразу. Дело в том, что лигнин придает клеточным стенкам огромную механическую прочность. Он необходим, чтобы сделать ствол наземного растения высоким, вплоть до многометрового, и создать транспортную систему из микроскопических трубочек, качающую воду на всю эту высоту. Именно с "изобретением" биосинтеза лигнина связано одно из крупнейших событий, поменявших лик Земли - появление лесов (Еськов, 2000). Кроме того, появление лигнина сильно изменило глобальный круговорот углерода. Тут дело в том, что лигнин с его разнообразными мономерами и перепутанными химическими связями исключительно неподатлив к действию ферментов. Поэтому растительной тканью, в которой много лигнина, почти невозможно питаться. Из всех земных живых организмов эффективно разлагать лигнин "научились" только грибы, причем не все и не сразу (Robinson, 1990). Именно они и стали разрушителями мертвых деревьев. До этого вся огромная биомасса лигнифицированной древесины просто захоранивалась как есть, создавая залежи каменного угля, в честь которых получил название целый геологический период - каменноугольный, или карбон.

Карбоновые леса непрерывно вели фотосинтез и выделяли в атмосферу огромное, немыслимое в более ранние эпохи количество кислорода, который не расходовался на окисление стволов погибших деревьев, потому что перерабатывать их было еще некому. В результате доля кислорода в атмосфере достигла уникальной в истории Земли цифры 35% (Beerling et al., 2002). Как известно, современная атмосфера Земли содержит "всего" 21% кислорода. На самом деле по космическим меркам и это очень много, но в карбоне было в полтора раза больше. Связано это именно с тем, что огромная биомасса стволов деревьев со всеми содержащимися там полимерами не съедалась никакими живыми существами, в отличие от современной ситуации, когда упавшие стволы измельчаются насекомыми, перерабатываются грибами и в итоге их углеродные соединения окисляются дыханием до углекислого газа (CO2) - при этом расходуется кислород (O2), а углекислый газ уходит в атмосферу. А вот до той биомассы, которая успела захорониться в виде каменного угля до возникновения эффективных деструкторов, биосфера смогла "добраться" только с появлением человека, который неутомимо откапывает каменный уголь и жжет его. Процессы дыхания и горения описываются одним и тем же суммарным уравнением: C6h22O6 (глюкоза) + 6O2 → 6CO2 + 6h3O. Так что в итоге получается тот же самый углекислый газ, из которого фотосинтезирующие организмы (то есть растения) могут заново создать более сложные углеродные соединения, пригодные для построения тел живых существ.

Еще один очень распространенный в природе полисахарид - хитин, из которого состоят клеточные стенки грибов и наружные панцири очень многих многоклеточных животных. Это полимер, во многом похожий на целлюлозу. Он тоже состоит из остатков β-глюкозы, но только модифицированных. Хитин - азотсодержащий полисахарид. Его мономер - строго говоря, не глюкоза, а ацетилглюкозамин, производное глюкозы, где ко 2-му атому углерода вместо гидроксила присоединена аминоацетильная группа -NH-CO-Ch4.

В состав клеточных стенок бактерий входят еще более сложные азотсодержащие производные глюкозы, к которым дополнительно ковалентно "пришиты" цепочки аминокислот. Такой многокомпонентный полимер называется пептидогликаном. Запоминать детали тут не имеет никакого смысла, единственное, что стоит обязательно отметить - в состав пептидогликанов входят не только L-, но и D-аминокислоты. Это тот редкий случай, когда D-аминокислоты в живых организмах все-таки встречаются. Пептидные цепочки, входящие в пептидогликан - именно пептиды, но не белки.
Со времен работавшего еще в XIX веке ученого-медика Ганса Христиана Грама (Hans Christian Joachim Gram) бактерий делят на грамположительных и грамотрицательных, в зависимости от того, окрашиваются ли они определенным химическим методом, который Грам изобрел. Чем они отличаются по строению клеток - показано на картинке; из еще не встречавшихся нам слов здесь стоит пояснить липопротеин (белок с липидной частью), липотейхоевую кислоту (спиртовой полимер, связанный с липидами) и порины - транспортные белки, создающие в мембране как бы поры для воды и растворенных в ней мелких молекул. Но эти детали не должны заслонять от нас интереснейшую проблему. У грамположительных бактерий снаружи от мембраны находится толстая пептидогликановая клеточная стенка - в этом плане их клетка похожа, скажем, на растительную, не считая того, что материал клеточной стенки другой. А вот у грамотрицательных бактерий есть две полноценные билипидные мембраны - внутренняя и наружная - и относительно тонкая пептидогликановая клеточная стенка между ними! Так не устроены никакие другие клетки. Есть гипотеза, что первые на Земле живые организмы были именно грамотрицательными бактериями, и только у их потомков вторая - наружная - мембрана исчезла (Cavalier-Smith, 2006). Независимо от того, верна эта гипотеза или нет, эволюционный зигзаг тут получился очень занятный.
caenogenesis.livejournal.com