
какого внутренее строение стебля дерева или кустарника ? Утолщение стебля и корня растений зависит от
21.Сравнительная характеристика первичного строения корня и стебля.
Первичное строение:
Корень:ризодерма или эпиблема всасывающая. Корневой волосок - вырост клетки ризодермы. Кутикула отсутствует. Смена покровной ткани от эпиблемы до экзодермы.
Стебель: У стебля эпидерма выполняет функции: защита, газообмен, фотосинтез (собирает свет и направляя его в хлоренхиму).
Первичная кора.
У корнязанимает более 50 % объема корня, четко дифференцирована на 3 слоя. Функции: проводящая (с регуляцией), синтетическая, симбиотическая и запасающая.
У стебля первичная кора занимает периферическую зону и по объему 20-30%. Функции: запасающая, синтетическая, иногда механическая. Слабо дифференцирована на зоны энтодерма выражена биохимически.
Центральный цилиндр.
У корня- радиальный сложный пучок, проводящие ткани отделены паренхимой. Функции: проводящая.
У стеблядо 50% объема проводящей ткани собраны в пучки стандартного строения.В качестве сопровождающей ткани выступает склеренхима и паренхима. Функции: проведение веществ.
Сердцевина.
У корня -либо склеренхима (однодольные) либо звезда ксилемы (двудольные).
У стебля - либо заполнена паренхимой, либо отсутствует.
22. Анатомическое строение корня, корневища и клубня.
Анатомия корня.
Зона деления клеток, представляющая собой апикальную меристему, прикрытую корневым чехликом.
Зона роста, или растяжения, а которой клетки, уже вышедшие за пределы меристемы, находятся в разных стадиях их растяжения в продольном направлении.
Зона поглощения вещества и начала дифференциации постоянных тканей,- ризодермы, характеризующейся развитием корневых волосков и появлением проводящих элементов ксилемы и флоэмы, которая закладывается еще в зоне роста.
Зона первичного строения с заканчивающейся дифференциацией постоянных тканей, в которой четко выражены первичная кора и центральный цилиндр или стела.
Зона ветвления, в нижней части которой закладываются зачатки боковых корней, в верхней части они уже заметны на поверхности корня.
Анатомическое строение клубня. На продольном разрезе спелого клубня под микроскопом видно такие элементы: шкурку (у молодого клубня - эпидермис), кору, камбий, сосудистые щепотки, сердцевину. Шкурка – внешний защитный слой клубня, состоит из нескольких рядов опробкованных клеток вторичной покровной ткани – перидермы. Под шкуркой размещается кора, которая состоит из паренхимных клеток, заполненных крахмальными зернами, и ведущих элементов луба – ситоподобных трубок флоэмы. За корой находится слой клеток камбия, с которого до центра клубня образуются элементы ксилемы. Центральная часть клубня заполнена паренхимными клетками сердцевины, которая радиальными лучами расходится к леткам в местах их размещения. В паренхимных клетках клубней содержатся крахмальные зерна. Наибольшее количество их находится во внутренних клетках коры и внешних – сердцевины, наименьшая – в составе водянистых клеток центральной сердцевины.
Анатомия корневища
Анатомически стеблевая часть корневища сходна со стеблем надземного побега, отличаясь от него более широкой первичной корой, в которой откладываются запасные вещества и обычно хорошо выраженной эндодермой, оболочки клеток которые имеют пояски Каспари. У ландыша внутренние и боковые стенки клеток эндодермы со временем утолщаются и одревесневают. Подковообразные утолщения стенок нередко образуются и у прилегающих к эндодерме паренхимных клеток первичной коры. Особенность корневища ландыша состоит так же в наличии двух типов проводящих пучков примыкающих к перициклу коллатеральных с дугообразно изогнутой на поперечном срезе ксилемой, и внутренних, в которых ксилема со всех сторон окружает флоэму. Есть и переходные формы между этими типами пучков. Корневища со вторичным утолщением, свойственные двудольным растениям, отличаются от наземных стеблей отсутствием или очень слабым развитием механических тканей, наличием активно дилатирующих сердцевинных лучей и обычно более широкой зоной первичной коры. В многолетних корневищах развита перидерма.
1.коллатеральный пучок.2.ксилема.3концетрический (амфивазальный) пучок.4паренхима.5первичная кора.5перицикл.6флоэма.7.эпидерма.
studfiles.net
какого внутренее строение стебля дерева или кустарника ?
В кожице стебля, как и в кожице листа, имеются устьица, через которые происходит газообмен. В пробке развиваются чечевички маленькие бугорки с отверстиями. Через них также осуществляется газообмен. У некоторых деревьев образуются толстые слои пробки. Особенно мощная пробка развивается на стволе пробкового дуба. Ее используют для разных хозяйственных нужд.
Под пробкой у трехлетней ветки липы находятся клетки первичной коры, а за ними, то есть еще глубже, расположен луб. В его состав входят ситовидные трубки, по которым перемещаются растворы органических веществ, и толстостенные лубяные волокна. Эти вытянутые клетки с разрушенным содержимым и одревесневшими стенками представляют механическую ткань стебля. В стеблях льна, липы и некоторых других растений лубяные волокна развиты особенно хорошо и очень прочны. Из лубяных волокон льна изготавливают льняное полотно, а из лубяных волокон липы — мочало и рогожу.
Древесина расположена глубже луба. Потрогайте пальцами поверхность только что обнаженной древесины, и вы почувствуете, что она влажная и скользкая. Это объясняется тем, что между лубом и древесиной залегает камбий.
Камбий — один слой узких длинных клеток с тонкими оболочками. Они рвутся при обнажении древесины. При этом содержимое клеток камбия вытекает и увлажняет поверхность древесины.
Камбий играет большую роль в жизни стебля. Его клетки делятся, и в результате по обе стороны от камбия возникают слои клеток. Клетки, которые откладываются в сторону коры, становятся новыми клетками луба, а клетки, отложенные камбием в сторону древесины, — новыми клетками древесины. Поэтому камбий считают образовательной тканью, от которой зависит утолщение стебля.
Древесина — это основная часть ствола дерева. Она образована клетками разной формы и величины. Оболочки многих клеток утолщены и пропитаны веществом, придающим им плотность. В состав древесины входят длинные трубкообразные сосуды.
Сердцевина состоит из крупных клеток с тонкими оболочками Здесь откладываются в запас питательные вещества. Обычно сердцевина состоит из рыхлой ткани.
otvet.mail.ru
Сложные соцветия
Сложными называются соцветия, в которых на главной оси располагаются не одиночные цветки, а парциальные (частные) соцветия.
Сложные рацемозные соцветия
Сложные зонтикиу укропа.
Сложные колосья, характерные для злаков (пшеница, рожь, ячмень).
Сложная кисть – борщевик сибирский.
Сложные цимозные соцветия
Цимоиды— это сложные соцветия с симподиальным нарастанием, в которых главная ось не выражена. Они делятся на три основные варианта: дихазии, монохазии и плейохазии, в зависимости от того, сколько боковых ветвей сменяют в ходе симподиального нарастания одну материнскую.
Дихазияминазывают цимозные соцветия, в которых каждая ось несёт две оси следующего порядка. Дочерние оси здесь появляются в верхней части материнской и перерастают её верхушку.
Монохазии— такие цимозные соцветия, в которых каждая материнская ось несёт только одну дочернюю. Как и у дихазиев, дочерние оси здесь располагаются в верхней части материнской оси и перерастают её верхушку. Монохазии делят наизвилиныизавитки.
Завиток— цимозное соцветие, в котором от главной оси с единственным цветком отходит другая ось с единственным цветком, а от той — ось третьего порядка и так далее, при этом все цветки направлены в одну сторону. Такой тип соцветия характерен для окопника.
Если же цветки более высоких порядков возникают попеременно то справа, то слева по отношению к цветкам более низких порядков, то такое соцветие называется извилиной(гладиолус тонкий и др.).
Плейохазияминазывают цимоиды, в которых каждую материнскую ось сменяют несколько более или менее мутовчато расположенных дочерних, перерастающих её верхушку (некоторые виды лютика, бузина).
Соцветия различного типа, у которых главная ось является повислой, называются серёжками. Среди серёжек встречаются (ольха, берёза, лещина).
45 Атипическое вторичное утолщение стебля двудольных растений.
АТИПИЧНОЕ УТОЛЩЕНИЕ – вторичное утолщение стебля или корня, происходящее с отклонениями от обычного. Может быть результатом одновременной деятельности нескольких камбиев, разновременного функционирования нескольких поочередно возникающих камбиев, а также деятельности особой меристематической зоны в отсутствие камбия и т. д.
Это изменение соотношения ксилемы и флоэмы.
У большинства древесных лиан – лазящих видов, пользующихся стволами и ветвями других растений как опорой, - вторичный прирост атипичен; он обладает особыми чертами, в силу которых стебель гибок, подобно канату. Общей чертой строения стебля лиан является раздробленность древесинного тела на участки с мягкими паренхимными участками в промежутках. Число участков с неравномерной деятельностью камбия увеличивается. Это создает необходимую для этих экологических форм растений гибкость стебля.
Атипическое утолщение связано с 4-мя возможными механизмами:
одновременная закладка нескольких слоев (массивов) камбия.
последовательная закладка нескольких слоев камбия.
образование вместо камбия меристематического кольца.
атипическая работа камбия.
46 Лист как орган, его признаки и функции. Возникновение и развитие листа в эволюции.
Внешнее (морфологическое) разнообразие листьев так велико, что невозможно создать единую систему классификации листьев по одному или нескольким признакам, поэтому существует несколько разнообразных классификаций листьев, основанных на различных признаках.
Существуют три способа прикрепления листьев к стеблю. Если лист имеет черешок, такой лист называют черешковым. Если лист не имеет черешка, такой лист называют сидячим. У некоторых растений основание листа разрастается и охватывает стебель над узлом. Такой лист называют влагалищным, влагалище может быть закрытым (если края срастаются) и закрытым (если края свободны).
Листовые пластинки могут быть очень разнообразны по величине, по форме, структуре и многим другим признакам.
Листья могут состоять из одной или нескольких листовых пластинок. Если лист состоит из одной листовой пластинки, такой лист называют простым. Если на одном черешке расположено несколько листовых пластинок, такой лист называют сложным. Среди сложных листьев встречаются: тройчатосложный, пальчатосложный, парноперистосложный, непарноперистосложный, дваждыперистосложный, триждыперистосложный, прерывистоперистосложный.
Простые листья также могут быть расчлененными. Если листовая пластинка рассечена не более чем на 1/4 ширины, такой лист называют лопастным. Если листовая пластинка рассечена на 1/3, такой лист называют раздельным. Если же рассечение достигает главной жилки листа, такой лист называют рассеченным. Но и количество рассечений на одном листе может быть различным. Если лист рассечен на три части, такой лист называют тройчато-, если на пять частей — пальчато-, если на большое количество частей — перисто- (лопастным, раздельным, рассеченным).
Листовые пластинки можно классифицировать по форме. Существует огромное количество форм листовых пластинок: округлые, яйцевидные, линейные, ланцетные, копьевидные, стреловидные, сердцевидные, продолговатые и др. Можно классифицировать по форме края. Кроме листьев с цельным краем, выделяют несколько основных видов формы края: зубчатый, пильчатый, шиповатый (колючезубчатый), городчатый, выямчатый, извилистый.
Необходимо заметить, что на одном побеге могут встречаться листья различной формы, величины, окраски. Это явление называется гетерофилией. Гетерофилия характерна для лютика, стрелолиста и многих других растений.
Также Выделяют несколько типов жилкования: параллельное, дуговое, дихотомическое, сетчатое (пальчатое и перистое). Параллельное или дуговое жилкование характерно для однодольных растений, а сетчатое — для двудольных.
studfiles.net
Вторичное строение корня
После образования камбия у некоторых растений из наружных слоев перицикла, успевшего к этому моменту из однослойного стать многослойным, или из тканей коры развивается перидерма.
Начиная с этого структурного состояния, центральный цилиндр корня в основе своего строения мало чем отличается от центрального цилиндра стебля. В отличие от стебля в корнях многих растений сохраняются, однако, группы сосудистых элементов протоксилемы, и первичные сердцевинные лучи упираются в них.
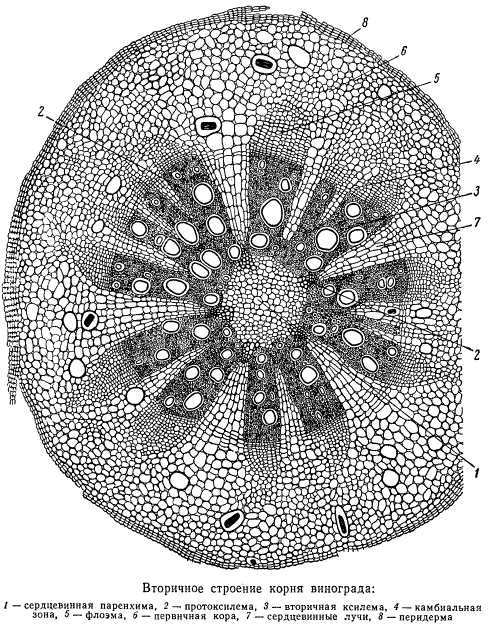
Во вторичной коре корней некоторых растений образуются лубяные волокна, не столь обильные, как в стебле, но достаточно определенно выраженные. Вторичная ксилема корней отличается от ксилемы стебля обилием широкополостных сосудов, напоминая в некоторой степени строение вторичной ксилемы стебля лиан, чему благоприятствует в известных пределах характер разрастания корня среди частиц почвы, необходимость обходить различные препятствия, что уподобляет его лиане.
Корни однодольных за очень редкими исключениями к вторичным утолщениям не способны. Только у таких растений, как драцены и юкки, корни, подобно стеблям, утолщаются. Утолщение корней у них происходит по такому же принципу, как и утолщения стеблей. В паренхиме коры сначала дифференцируется слой меристематических клеток, кольцом окружающий центральную часть корня. В этом меристематическом образовательном кольце возникают отдельные сосудистые пучки. За первым кольцом, снаружи его, дифференцируется второе кольцо меристематической ткани и т. д.
В жизни растения корень выполняет несколько функций: поддерживает в прямом состоянии стебель, поглощает воду и растворенные в ней вещества, а также служит местом отложения запасных веществ.
Общая структура корня обладает отчетливо выраженными чертами приспособления к откладыванию запасов. Обычно в корне сердцевинные лучи шире, чем в стебле, паренхимы больше, а механических элементов меньше. Сельскохозяйственная практика путем отбора добивается получения форм растений с гипертрофически разросшимися корнями, так как такие корни накапливают в себе большие количества питательных веществ (корнеплоды). Всем известны гипертрофированные корни редьки, турнепса, моркови, свеклы. В связи с гипертрофией такие корни значительно уклоняются от нормальной структуры. Разумеется, черты структуры, связанные с отложением запасов, были свойственны и дикорастущим предкам современных культурных растений, образующих корнеплоды, но только искусственный отбор усилил эти свойства, доведя гипертрофию корней до грандиозных размеров.
Остановимся кратко на рассмотрении особенностей структуры корня свеклы.
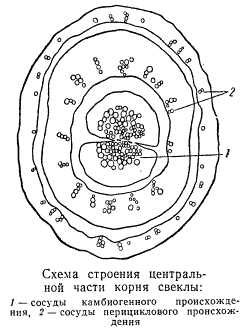
На поперечном разрезе корня свеклы, особенно некоторых столовых сортов ее, хорошо видны концентрические круги. Число концентрических кругов у различных корней неодинаково. Каждое концентрическое кольцо состоит из паренхимы и расположенных в паренхиме сосудистых пучков. Каждый пучок включает только флоэму и ксилему, не имея ни камбия, ни механических элементов.
В начале развития корня свеклы в нем залягается камбий, образующий элементы вторичной ксилемы и флоэмы. Однако продолжительность деятельности камбия в корне свеклы весьма незначительна и вскоре замирает. Таким образом, только центральная часть корня камбиогенного происхождения. После прекращения камбиальной деятельности рост корня свеклы в толщину продолжается, но уже за счет деятельности перицикла. Из перицикла в закономерной последовательности возникают все новые кольца, состоящие из сосудистых пучков.
Образование колец сосудистых пучков непосредственно зависит от листьев прикорневой розетки, число же колец кратно числу листьев розетки.
При этом у таких сортов свеклы, как египетская и эрфуртская, число колец в корне равно числу листьев разетки, деленному на 3, а у сортов эккендорфская и маммутна — деленному на 6. Вообще если листьев розетки развертывается мало, то и число колец будет соответственно незначительно, как бы долго свекла ни росла, и наоборот. Эти факты свидетельствует о том, что структура корня свеклы испытывает воздействие листьев и что сосудистые пучки в кольцах корня есть листовые следы листьев прикорневой розетки.
Несомненно, участие листьев в построении корня довольно значительно. Все мясистые корни в большинстве случаев имеют более или менее большую прикорневую розетку, состоящую из большого числа листьев. Чем число листьев или размеры их значительнее, тем толще корень.
Существуют разнообразные модификации структуры различных частей корня в соответствии со специфическими особенностями экологии растений, с условиями их произрастания.
Корни растений, произрастающих в воде или в сильно увлажненной почве, вообще развиваются слабо и особенно слабо выявлена у них проводящая система. У некоторых таких растений первичная кора по сравнению с центральным цилиндром имеет очень большие размеры и пронизана обширными воздухоносными полостями.
У некоторых тропических растений, например орхидных, произрастающих эпифитно, корни спускаются к почве с большой высоты. Воздушная часть таких корней имеет специфическую структуру. Она состоит из двух разделов, дифференцирующихся как наружная и внутренняя кора. В наружной коре, происшедшей из наружного слоя клеток конуса нарастания корня, называемого дерматогеном, образуются особые паренхимные клетки, имеющие вторичные утолщения в виде тонких спиральных полос и лишенные живого содержимого. Эти клетки, возможно, способны поглощать воду из паров воздуха и удерживать ее в виде запасов, используемых растением в дальнейшем.
Настоящей корой является внутренняя кора, возникшая из следующего меристематического слоя конуса нарастания — его периблемы. Внутренняя кора отделена от наружной коры однорядным слоем клеток — экзодермой.
В окончательно сформировавшемся виде клетки экзодермы толстостенные, лишены живого содержимого, но среди них есть пропускные клетки, через которые продвигаются водные растворы из наружной коры во внутреннюю кору и центральный цилиндр.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Реферат Строение растения. Корень, побег, стебель.
Реферат по биологии
Строение растения. Корень, побег, стебель.
Корень вегетативный орган с неограниченным ростом, который обеспечивает закрепление растений в субстрате, погл-ние и транспорт воды и растворенных в ней минеральных веществ и продуктов жизнедеятельности почвенных микро-организмов и корней других растений, первичный синтез органичным веществ, выделение в почву продуктов обмена ве щества и вегетативное размножение.
Появление корня в процессе эволюции растений важный ароморфоз, одно из приспособлений к жизни на суше. Впер-е настоящие корни появляются в папоротникообразных. Затем в цветочных растений благодаря идиоадаптации фор-муються различные типы корней, способных выполнять дополни-тельно функции. Так, у некоторых растений в корнях откладывается запас питательных веществ, что приводит к их утолщение, ут-разования корнеплодов (морковь, редис, свекла) или коре-невых клубней (георгины, чистяк). Корни эпифитных растений (используют другие растения в качестве субстрата, но не паразит-тують на них: орхидеи, мхи, лишайники) могут нагрев-маджуваты воду. В тропических деревьев, живущих на грун-тах, бедных кислородом, или на болотах, образуются дыхательные корни пневматофоры (мангровые), растущие вверх; они поднимаются над поверхностью субстрата и обеспечивают дыхание. Ходульные корни образуются на надземных па-бегах, закрепляются в почве и прочно удерживают растение (фикус-баньян, кукуруза). Некоторые растения-паразиты (повы-тычет) или полупаразиты (омела) образуют корни-присоски. В вьющихся и лазящих растений формируются цепкие воздушные корни (плющ). У многих (около 90%) цветочных рос-линь корни вступают в симбиоз с грибами, образуя мико- ризу, или с бактериями, образуя бактериоризу. Микро-организмы-симбионты входят в состав ризосферы осно-тового слоя толщиной 2 3 мм, прилегающей к корням растений. Скопление большого количества грибов и бактерий в ризосфере связано с выделением корнями веществ, яки-ми питаются эти микроорганизмы.
Зародышбоковых ответвлений. Это свойство корня используют при выращивании рассады культурных растений с стержневым корнем. У молодых растений отщипывают кончик главного корня, прекращает его рост в длину и вызывает рост боковых и дополнительных корней в верхнем, плодороднейшем слое почвы. После отщипывание части в основном-го корня рассаду высаживают на постоянное место зростан ния с помощью заостренного колышка пинетки, отсюда этот процесс получил название пикирования.
Морфология корней, глубина и ширина их проникновения в почву зависят от вида растений, условий их существования, ме-тогда в искусственного влияния на рост растений. По объему коре-неве системы растений всегда больше, чем их надземные ча-стини.
Побег. В процессе эволюции побег сформировался как ком-комплексных орган, приспособленный для эффективного фотосин-тетичних процесса, транспирации, образования репродуктив-ных органов (спорангиев, шишек, цветов), опорной и транс-портных функций. При этом происходило отделение осевой стеблевой структуры с ответвлениями и форму-ем плоских боковых выростов с ограниченным ростом листьев, которые обеспечили увеличение контактов растений с воздушной средой и поглощения световой энергии. Характерной структурной особенностью типичного побега высших растений является наличие на нем почек Зачатки вых побегов, которые способны длительное время сохранять жизне- способность меристем и обеспечивать защиту их от неспры-ятливих внешних воздействий. Итак, корень и побег равноценные по рангу органы растения. Вследствие ветвления они образуют надземные и подземные системы, которые захватывают жизненное пространство. Стебли, листья и почки е зависимыми частями сложного органа побега.
Учитывая все сказанное выше, при дальнейшем изложении анатомического материала мы будем принимать термин "стебель" только в понимании его как осевой части побега.
Стебель как осевая часть побега осуществляет связь всех частей растения, увеличивает ее поверхность за счет ав-ния, образует и несет на себе брунькы и листья, обеспечивает транспорт воды, минеральных и органических веществ, веге-тативне размножения и фотосинтез, запасает питательные ве-вине. По сравнению с корнем побег растения имеет сложное строение, поскольку он на ранних этапах развития розчлено-ся на специализированные части и состоит из оси и листьев, расположенных на нем. Стебли без листьев, хотя бы зачаточных или рудиментарных, как и листья без стебля, хотя бы укороченного, образовываться не могут. Стебель при оптимальных условиях растет постоянно, давая боковых ответвлений ния и увеличивая листовую поверхность.
Первый (основной) побег развивается из почки по-родка семени. На побеге выделяются узлы и междоузлия. Узел это место прикрепления листа к стеблю. Мижвуз-ля расстояние между соседними узлами. Обычно на побеге несколько, иногда много узлов и междоузлий, они повторяются вдоль оси побега. Междоузлия могут быть длинными, тогда побег называют удлиненным (ростовым) укороченный побег имеет короткие междоузлия. На стеблях некоторых растений с очень короткими междоузлиями сближены листья образуют при-корневую розетку (одуванчик, морковь). Плодовые деревья и кусты образуют оба типа побегов: укороченные с малым приростом "Плодушки", на которых формируются цветки, согла-дом плоды, и удлиненные, как правило, бесплодны. Длинные междоузлия бывают в лиан.
Угол между листом и стеблем называют листовой па-зухою. В пазухе листа размещаются почки ха-терний образование стебля зачатки новых побегов, которые виника ют в определенном порядке на оси. Они обеспечивают длительное нарастание побега и его ветвления. Различают Бруни-ки по месту их расположения (пазушные, верховые) и по функциональному назначению (вегетативные, цветочные, спящие, дополнительные). Верхушечные почки размещены на вер-хивках стебля и его боковых ответвлений. Внешне почки защищены чешуйками (видоизмененные листья). В зимующих бру-ньок внешние чешуи плотные, кожистые, могут укрываться кутикулой или клейкими смолистыми веществами. во хлопками находится укороченный зачаточный побег с тесно сближенными зачаточными листьями, которые покрывают образующую ткань конуса нарастания. В пазухах зачатков нижних листьев есть зачатки пазушных почек, из которых развивают-ся боковые побеги.
В деревьев и кустов пазушные почки бывают ростовыми (вегетативными), с зачатками листьев и стебля, и цветочно-мы с зачатками цветков или соцветий, их можно различить по форме: ростовые удлиненные, с заостренной верхов-кой, а цветочные округлые и больше.
Некоторые пазушные почки могут оставаться в состоянии спо-кой неопределенно долго. Это спящие почки. Они развивает-ся и дают побеги в случае повреждения верхушечных почек, взлома стебля над ними.
Важным является формирование дополнительных почек, которые по-кладаються на стеблях, листьях, корнях и обеспечивают вегетативное размножение цветковых растений.
Рост стебля в высоту обеспечивает верхушечная почка, или почка зародыша семени. Клетки образующей ткани кону са нарастания постоянно делятся. В процессе разделения образует ются новые зачатки листьев и почек. После разделения клетками-ны растут, вследствие чего удлиняются междоузлия и весь стебель. По мере удаления от конуса нарастания способность клеток к делению уменьшается и начинается дифференции-ция их, что сопровождается образованием основных тканей.
Возможен и другой путь роста стебля: вставной или интеркалярный. В этом случае образующая ткань разделы-на участками клеток, способных к делению. Участок разделения размещена обычно в основе междоузлий. Такой рост ха-рактерно для злаковых.
Видоизменения побегов. Кроме основной функции, о которой уже говорилось, побеги выполняют ряд дополнительных функций, связанных с их видоизменениями (метаморфозами), которые возникли в процессе приспособительной эволюции. Это ву-сики длинные тонкие побеги с редуцированными листьями. Усиками (виноград, огурец и другие растения с лазая или ползучими стеблями) растения прикрепляются к различным предметам. Колючки укороченные побеги без листьев, что захищають растение от поедания животными (боярышник, дикая яблоня). Усики и колючки это видоизменения надземного побега. Они размещаются в пазухах листьев или на письмо-ном узле напротив листа, что свидетельствует о стеблевые происхождения их.
Кроме видоизменений надземного побега у растений есть видоизменения и подземного корневище, клубень, луковица, которые выполняет ют функции накопления питательных веществ и вегета-ного размножения.
Корневище это видоизмененный побег с небольшими чешуйчатыми листьями буроватого цвета и почками.
Корневище внешне похоже на корень, но отличается на-личие рудиментарных листьев, боковых и верхушечных бру-ньок и отсутствием корневого чехлика. По форме оно может быть длинным и тонким (пырей, осока) или коротким и толстым (щавель, ирисы). Ежегодно из верхушечной почки вырастает надземный побег. Во время обработки коре-не выше может быть измельченное, тогда каждая его часть с почкой даст новое растение. Особенно быстро происхо-ся вегетативное размножение с разветвленными ко-реневищамы (пырей).
Бульба это утолщенный верхушка подземного стебля столона. Наиболее известными являются клубни картофеля. О стеб-лове их происхождения свидетельствует наличие и спиральное разме-ние почек-глазков. Хлорофилла клубни не имеют, но на солнце зеленеют. Клубни могут быть и надземными на-пример утолщенный стебель капусты кольраби. Такие клубни имеют зеленый цвет.
Луковица очень укороченный подземный побег с вы-дозминенимы листьями. Луковицы бывают шаровидные, яйцо-образные, удлиненные и тому подобное. Стеблевой часть луковицы зайцы имеет незначительную зону, ее называют донышком. На нижней по-верхние донышка размещены многочисленные дополнительные корни, на верхней видоизмененные мясистые листья (чешуи), плотно прилегают друг к другу и имеют запас питательных ве-вин. Внешние листья (чешуи) выполняют защитную функ-цию, поэтому становятся кожистыми. В пазухах некоторых соков-х чешуек является почки, из которых развиваются или дочерние ци-были не (дети), или надзерастения, чечевички, которые имеют вид маленьких бугорков на поверхности стебля. Чечевички образованы крупными клетками основной ткани с большими межклетниками.
утолщение стебля происходит за счет пе-ной деятельности камбия. Клетки камбия постоянно действуют в период вегетации растения и могут функционировать упро-ние нескольких лет. Особенно интенсивно размножаются клетки камбия весной, поэтому кора из побегов легко снимает ться. Схематично процесс роста клеток камбия можно воображения-ти так: клетка камбия растет и делится. С двух новых клеток внутренняя специализируется и превращается в кли-типа древесины, прилегающей к старым ее элементов, а клетка камбия, отодвинута наружу, сохраняет способность к дальнейшей-го деления (т.е. остается камбиального). После разделения этой клетки внешняя дочерняя клетка начинает пре-рюватися на постоянную клетку коры, а внутренняя дочерняя остается камбиального. Эта камбиальных клеток снова образует клетку древесины, а дальше коры. Итак, камбий откладывает клетки непрерывно: внутрь клетки, из которых строится дерево, наружу клетки, из которых строится кора. Новые клетки древесины нарастают наружу, а клетки коры изнутри. При этом камбий все время кольцо-образное охватывает древесину.
Как правило, клеток древесины образуется значительно больше, чем клеток коры, поэтому древесина занимает значительную часть на поперечном разрезе стебля. Клетки древесины и коры, образовавшиеся из камбия, могут разрастаться как в поз-продольных, так и в поперечном направлении.
Рост клетки камбия прекращается с наступлением зимо- ного периода, а весной возобновляется. Только в посушлы вых районах (саваннах) растения летом теряют листья и вступают в летний период покоя. В условиях влажных тропиков действие камбия не прекращается, поэтому растения там растут постоянно.
На поперечном разрезе трехлетнего побега липы видно, что древесина имеет четко выраженные слои. Это годичные кольца, которые четко выделяются вследствие того, что древесина,которая образования-лась в разное время года, имеет различные цвет, блеск и плотность. Весенняя древесина, которая образовалась после восстановления дея-тельности камбия, состоит из крупных клеток с тонкими обо-ки. Она обычно богаче на сосуды и трахеиды, поэтому кажется пышнее и окрашена в светлый цвет. Осенняя древесина состоит из более толстостенных, но узких волокон и вместо элементов, которые проводят воду, содержит больше механической ткани, поэтому кажется темнее. В нас-лидок различия в структуре поздняя древесина функционирует преимущественно как механическая ткань, а весенняя проводит воду с растворенными минеральными солями. Переход от весенней древесины к осенней происходит постепенно, а переход от осенней к весенней всегда внезапный.
Ширина годовых колец зависит от вида дерева, его возраста (в начале жизни растения образуются широкие кольца, с возрастом они постепенно сужаются), от условий среды-выше. По ширине колец можно прочитать историю условий жизни дерева.
myreferat.net









