V1: Основы экологии и географии растений. Растения не относящиеся к группе криптофитов
Геофиты
Геофиты — многолетние травы с переннирующими тканями, скрытыми в почве (луковицы, клубнелуковицы, клубни или корневища), благодаря чему они более полно защищены от воздействия приземного слоя воздуха.[ ...]
Геофиты — разновидность криптофитов. См. Криптофиты.[ ...]
Криптофиты, или геофиты, подземные растения; зимующие почки скрыты в земле (клубневые или корневищные растения).[ ...]
Культурные корнеплоды — двулетние растения. Их относят к группе геофитов, у которых эпикотиль (головка), гипокотиль (шейка) и собственно корень превратились в органы накопления запасных питательных веществ, а почки возобновления, дающие начало листовым и цветоносным побегам, закладываются в надземных или подземных органах, близко от поверхности почвы.[ ...]
Криптофиты — растения, почки возобновления которых скрыты в почве (геофиты) или под водой (гидрофиты) (луковичные, клубневые и корневищные растения).[ ...]
При рассмотрении динамики флоры по структуре жизненных форм (по Раункиеру) можно сказать, что за 36 лет произошло увеличение групп фанерофитов (с 2 до 15 видов) и гемикриптофитов (с 25 до 54) и, в меньшей степени, увеличение групп геофитов (с 13 до 16) и терофитов (с 17 до 23), появилась группа хамефитов (6 видов). В группе гелофитов на первых этапах формирования происходило увеличение числа видов (от 1 до 5 за 1967-1972 гг.), а в последующие периоды - уменьшение до 1 вида, что связано с изменением эдафических условий.[ ...]
КРИП [англ. creep — ползать], ползучесть — то же, что оползни. КРИПТОФИТЫ [от гр. kryptos тайный, скрытый и phyton — растения] — многолетние травянистые растения, у которых почки возобновления закладываются на корневищах, клубнях, луковицах и находятся под землей (геофиты, напр, тюльпан) или под водой (гидрофиты). См. Жизненная форма.[ ...]
Однако это не значит, что каждое конкретное семейство или род обязательно прошли в ходе своей эволюции весь путь «соматической редукции». Некоторые семейства, видимо, с самого начала были травянистыми, и в некоторых случаях от травянистых предков возникли более специализированные древесные формы (бамбуки в семействе злаков). В крайних условиях эволюция приводила то к стланикам, то к подушкам, то к луковичным геофитам, то к однолетним эфемерам. Эфемеры считаются эволюционно наиболее молодой группой жизненных форм, свойственной области древнего Средиземья, которая стала сушей при высыхании древнего Средиземного моря — Тетис.[ ...]
В предгорьях и низкогорьях значительные пространства занимают пустынные типы растительности, особенно эфемеровая растительность. Для горных склонов характерны заросли листопадных кустарников, которые в большинстве случаев представляют собой вторичные сообщества, возникшие в результате уничтожения лесной растительности. Значительные пространства в горах занимают своеобразные формации степного типа с участием крупных зонтичных, эфемеров и геофитов и редколесья или реже леса, образованные можжевельниками.[ ...]
Известны морфологические приспособления растений к низким температурам, так называемые жизненные формы растений, которые, например, можно выделить по положению почек возобновления растительных видов по отношению к поверхности почвы и к защите, которую они получают от снежного покрова, лесной подстилки, слоя почвы и т. п. Вот некоторые из форм (по Раункеру): эпифиты — растут на других растениях и не имеют корней в почве; фанерофиты (деревья, кустарники, лианы) — их почки остаются над поверхностью снега и нуждаются в защите покровными чешуйками; криптофиты, или геофиты, теряют всю видимую растительную массу и прячут свои почки в клубнях, луковицах или корневищах, скрытых в почве; терофиты — однолетние растения, отмирающие с наступлением неблагоприятного сезона, выживают лишь их семена или споры.[ ...]
ru-ecology.info
Основы экологии и географии растений — МегаЛекции
51.Термин «экология» был введен:
-: К. Раункиером
-: К. Линнеем
-: Э. Геккелем
-: Э. Вармингом
52.Методы исследования экологии растений, позволяющие изучать объекты в естественных условиях, называются:
-: полевые
-: экспериментальные
-: комбинированные
53.Эдафические факторы относятся к экологическим факторам:
-: антропогенным
-: абиотическим
-: биотическим
54.Топографические факторы относятся к экологическим факторам:
-: антропогенным
-: абиотическим
-: биотическим
55.Фитогенные факторы относятся к экологическим факторам:
-: антропогенным
-: абиотическим
-: биотическим
56.Зоогенные факторы относятся к экологическим факторам:
-: антропогенным
-: абиотическим
-: биотическим
57.Виды растений с узкой экологической амплитудой называются стенобионты.
58.Виды растений с широкой экологической амплитудой называются эврибионты.
59.Синантропные виды растений по отношению к большинству экологических факторов являются:
-: эврибионтами
-: стенобионтами
60.Эндемичные виды растений по отношению к большинству экологических факторов являются:
-: эврибионтами
-: стенобионтами
61.Характерным способом преодоления неблагоприятных условий для растений является:
-: приобретение выносливости
-: избегание
62.Для травяного яруса в широколиственном лесу лимитирующим фактором является:
-: богатство почв
-: температурный режим
-: освещенность
-: увлажненность
63.Наиболее физиологически активные для растений лучи солнца имеют длину волны:
-: 220-380 нм
-: 380-710 нм
-: 710-870 нм
64.Экологическая группа растений хорошо освещенных местообитаний называется гелиофиты (светолюбивые).
65.Экологическая группа растений, адаптированных к мало освещённым местообитаниям, называется сциофиты (теневыносливые).
66.Наличие воскового налета на листьях растений является адаптацией к условиям излишнего:
-: освещения
-: увлажнения
-: богатства почв
67.Наличие мясистых стеблей или листьев у растения является адаптацией к условиям недостаточного:
-: освещения
-: увлажнения
-: богатства почв
68.Хорошо развитая палисадная паренхима с обоих сторон листа характерна для:
-: эпифитов
-: эфемероидов
-: гелиофитов
-: сциофитов
69.Тип климата при котором испарение превышает поступление влаги называется аридный.
70.Тип климата при котором осадки превышают испарении влаги называется гумидный.
71.Экологическая группа растений переувлажненных местообитаний называется гигрофиты.
72.Экологическая группа растений водных местообитаний называется гидрофиты.
73.Экологическая группа растений местообитаний с нормальным увлажнением называется мезофиты.
74.Экологическая группа растений сухих местообитаний называется ксерофиты.
75.Ксерофиты, запасающие воду в своих вегетативных органах, называются суккуленты.
76. Ксерофиты, имеющие адаптацию к засушливым условиям в виде уменьшения транспирации, называются склерофиты.
77.Соответствие между экологической растением и группой:
R1: гигрофит L3: череда трехраздельная
R2: ксерофит L1: ковыль перистый
R3: гидрофит L4: ряска малая
R4: эпифит
R5: мезофит L2: чина весенняя
78.Правильная последовательность экологических групп растений по отношению к увлажнению (начиная от самых засухоустойчивых):
1: ксерофиты
2: мезофиты
3: гигрофиты
4: гидрофиты
79.Растение Икотник серый имеет стержневую корневую систему, мелкие опушенные листья, хорошо развитую механическую ткань. Исходя из вышесказанного, Икотник серый является:
-: гигрофитом
-: гидрофитом
-: ксерофитом
-: мезофитом
80.Растение Липучка щетинистая имеет стержневую корневую систему, мелкие опушенные листья, хорошо развитую механическую ткань. Исходя из вышесказанного, Липучка щетинистая является:
-: гигрофитом
-: гидрофитом
-: ксерофитом
-: мезофитом
81.Растения, способные в определенных границах регулировать расход воды путем закрывания устьиц и складывания листьев, называются:
-: гомеогидридные
-: пойкилогидридные
82.Растения, предпочитающие богатые почвы, называются:
-: мезотрофы
-: олиготрофы
-: эутрофы
83.Растения, имеющие адаптации к произрастанию на бедных почвах, называются:
-: мезотрофы
-: олиготрофы
-: эутрофы
84.Растение, использующее другое растение в качестве субстрата, но не наносящее ему вреда называется эпифиты.
85.Растительноядные животные, питающиеся одним видом растений, называются:
-: полифаги
-: бактериофаги
-: монофаги
-: олигофаги
86.Растительноядные животные, питающиеся многими видами растений, называются:
-: монофаги
-: бактериофаги
-: полифаги
-: олигофаги
87.Формирование розеточной формы листьев у растений является адаптацией к:
-: распространению плодов
-: собирательству людьми
-: выпасу скота
-: опылению
88.Птицы, как правило, распространяют семена растений:
-: внутри организма
-: на покровах тела
89.Произрастание эпифитов на стволах древесных растений является примеров фитогенных факторов:
-: косвенных
-: комбинированных
-: прямых
90.Выпас животных является примером зоогенных факторов:
-: прямых
-: косвенных
-: комбинированных
91.Вырубка человеком леса является примером антропогенных факторов:
-: комбинированных
-: прямых
-: косвенных
92.Затенение деревьями травянистых растений леса является примеров фитогенных факторов:
-: комбинированных
-: прямых
- косвенных
93.Термин «жизненная форма» был введен:
-: Г. Вальтером
-: Л.Г. Раменским
-: Б.М. Миркиным
-: Е. Вармингом
94.Система жизненных форм, в основе которой лежит положение почек возобновление была разработана:
-: К. Линнеем
-: К. Раункиером
-: И.Г. Серебряковым
-: Л.Г. Раменским
95.Растения, не формирующие почек возобновления, по системе жизненных форм К. Раункиера называются:
-: терофиты
-: хамефиты
-: фанерофиты
-: криптофиты
96.Растения, имеющие почки возобновления на поверхности земли, по системе жизненных форм К. Раункиера называются:
-: фанерофиты
-: криптофиты
-: гемикриптофиты
-: хамефиты
97.Растения, имеющие почки возобновления на зимующих побегах невысоко над поверхностью земли, по системе жизненных форм К. Раункиера называются:
-: гемикриптофиты
-: фанерофиты
-: криптофиты
-: хамефиты
98.Растения, почки возобновления которых расположены под землей (водой), по системе жизненных форм К. Раункиера называются:
-: гемикриптофиты
-: фанерофиты
-: криптофиты
-: хамефиты
99.Растительный покров умеренного пояса Земли характеризуются преобладанием:
-: фанерофитов
-: гемикриптофитов
-: криптофитов
-: хамефитов
100.Растительный покров тропического пояса Земли характеризуются преобладанием:
-: фанерофитов
-: хамефитов
-: гемикриптофитов
-: криптофитов
101.Жизненная форма по системе К. Раункиера, к которой относятся луковичные растения называется криптофиты.
102.Жизненная форма по системе К. Раункиера, к которой относятся корневищные травы называется криптофиты.
103.Правильная последовательность жизненных форм по системе К. Раункиера, начиная с самого высокого расположения почек возобновления:
1: гемикриптофиты 3
2: криптофиты 4
3: фанерофиты 1
4: хамефиты 2
104.Система жизненных форм, в основе которой лежит количество и продолжительность жизни стеблей и всего растения, его одревеснение и особенности подземных органов была разработана:
-: К. Раункиером
-: К. Линнеем
-: И.Г. Серебряковым
-: Л.Г. Раменским
105.Растения, дающие органы размножения один раз за период своей жизни, называются:
-: неокарпики
-: олигокарпики
-: поликарпики
-: монокарпики
106.Многолетние растения, не имеющие одревесневших органов и переходящие к размножению больше одного раза в течение жизни, называются:
-: монокарпики
-: поликарпики
-: полукустарнички
-: деревья
107.Многолетние растения с одним четко выраженным одревесневшим стеблем называются деревья.
108.Многолетние растения с несколькими равнозначными одревесневшими стеблями называются кустарники.
109.Многолетнее достаточно высокое растение у которого только нижняя часть побегов деревенеет и сохраняется зимой на протяжении многих лет полукустарники.
110.Правильная последовательность появления жизненных форм в процессе эволюции:
1: монокарпические травы3
2: поликарпические травы 2
3: деревья 1
111.Соответствие между жизненной формой и растением:
L1: Дерево R4: Береза повислая
L2: Кустарник R1: Черника обыкновенная R2: Карагана древовидная
L3: Травянистый поликарпик R3: Одуванчик лекарственный
L4: Травянистый монокарпик R5: Пастушья сумка обыкновенная
112.Наука, изучающая распространение растений по земной поверхности и состав флор различных территорий, называется: флористика
-: география растений
-: экология растений
-: ботаника
-: хорология
113.Учение об ареалах называется:
-: геоботаника
-: хорология
-: биоморфология
-: таксономия
114.Территория, в пределах которой встречается таксон, называется ареал.
115.Растения с очень небольшим ареалом любого происхождения называются стенохоными.
116.Очень широкий ареал, занимающий большую часть территории Земли, называется:
-: викарирующий
-: космополитный
-: эндемичный
-: дизъюнктивный
117. Мало различающиеся по морфологическим признакам, но географически разобщенные таксоны называются:
-: родственными
-: аборигенными
-: викарирующими
-: адвентивными
118.Известно, что представители семейства магнолиевые в дикой природе встречаются в Юго-восточной Азии и в Америке. Такой ареал называется:
-: дизъюнктивный
-: сплошной
-: эндемичный
-: космополитный
119.Ареал, разделенный на отдельные более или менее крупные части называется:
-: разорванным
-: неустойчивым
-: дизъюнктивным
-: сплошным
120.Известно, что в европейской части России встречается вид купальница европейская, а в азиатской части страны встречается близкий ему по морфологии и экологии вид купальница азиатская. Такие виды называются:
-: адвентивными
-: эндемичными
-: синантропными
-: викарирующими
121.Совокупность видов растений, встречающихся на какой-либо территории, называется флора.
122.Анализ систематической структуры флоры называется:
-: экологический
-: таксономический
-: биоморфологический
-: флорогенетический
123.Анализ спектра жизненных форм флоры называется:
-: таксономический
-: флорогенетический
-: биоморфологический
-: экологический
124.Совокупность растений на небольшой территории порядка 100-500 м2 называется конкретной флорой.
125.Процесс изменений во флоре под влиянием человека называется:
-: мезофитизация
-: олигомеризация
-: синантропизация
-: антропофитизация
126.Самая крупная единица флористического районирования Земли называется:
-: царство
-: империя
-: область
-: провинция
127.Территория, во флоре которой присутствуют эндемичные и характерные семейства и роды растений, а флора в целом своеобразна и оригинальна называется флористическим царством.
128. Крупнейшее по площади флористическое царство называется:
-: палеотропическое
-: капское
-: голарктическое
-: голантарктическое
129.Беднейшее по числу видов флористическое царство Земли называется:
-: палеотропическое
-: капское
-: голантарктическое
-: голарктическое
130.Наименьшее по площади флористическое царство Земли называется:
-: палеотропическое
-: голантарктическое
-: капское
-: голарктическое
131.Территория Новосибирска входит в флористическое царство: голарктическое
132.Территория, где группа видов пережила неблагоприятный период геологического времени, в течение которого на больших пространствах эти виды исчезали, называется рефугиум.
133.Совокупность растений, произрастающих совместно на однородной территории, характеризующаяся определенным составом и структурой называется фитоценоз.
134.Необратимая смена фитоценозов называется.
-: ярусность
-: мозаичность
-: сукцессия
135.Горизонтальная структура фитоценоза называется.
-: мозаичность
-: сукцессия
-: ярусность
136.Вертикальная структура фитоценоза называется.
-: ярусность
-: мозаичность
-: сукцессия
137.Совокупность фитоценозов определенной территории называется.
-: флора
-: ареал
-: растительность
-: население
138.Виды растительного сообщества, преобладающие над другими по числу особей, называются:
-: доминанты
-: эдификаторы
-: со-доминанты
-: характерные виды
139.Растительность, развивающаяся на плакорах, называется зональной.
140.Растительность, которая развивается в особых, нетипичных для данной зоны условиях среды и в то же время нигде не образует своей отдельной зоны, называется:
-: экстразональная
-: зональная
-: синантропная
-: интразональная
141.Болота умеренной зоны являются растительностью:
-: интразональной
-: зональной
-: экстразональной
142.Луга умеренной зоны являются растительностью:
-: зональной
-: интразональной
-: экстразональной
143.Растительное сообщество, эдификатором которого является сосна сибирская, относится к тайге:
-: мелколиственной
-: темнохвойной
-: светлохвойной
144.Растительное сообщество, эдификатором которого является сосна обыкновенная, относится к тайге:
-: мелколиственной
-: светлохвойной
-: темнохвойной
145.Растительное сообщество, эдификатором которого является береза повислая, носится к лесу:
-: мелколиственному
-: широколиственному
-: смешанному
146.Растительное сообщество, эдификатором которого является дуб черешчатый, носится к лесу:
-: широколиственному
-: мелколиственному
-: смешанному
147.Растительное сообщество с преобладанием травянистых ксерофитных растений, называется:
-: луг
-: лес
-: болото
-: степь
148.Растительное сообщество с преобладанием травянистых мезофитных растений, называется:
-: лес
-: болото
-: луг
-: степь
149.Растительное сообщество с преобладанием мхов и лишайников и с широким распространением карликовых деревьев, называется:
-: тундра
-: степь
-: лес
-: болото
150.Большая часть территории Новосибирской области входит в зону:
-: леса
-: степи
-: тундры
-: лесостепи
Итоговый тест
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Криптофитовые водоросли - это... Что такое Криптофитовые водоросли?
| Криптофитовые водоросли | |||||
| Rhodomonas salina | |||||
| |||||
| Cryptophyceae Pascher, 1913 | |||||
Криптофитовые водоросли (криптомонады, криптофиты, лат. Cryptophyta) — тип хромистов, включающий класс Cryptophyceae и два порядка — Cryptomonadales и Goniomonadales.
Криптофиты — небольшая, но экологически и эволюционно очень важная группа организмов, обитающих в морских и континентальных водах. Почти все они — одноклеточные подвижные жгутиконосцы, некоторые представители способны формировать пальмеллоидную стадию. Только один род Bjornbergiella (выделен из почв Гавайских островов) способен к образованию простых нитчатых талломов (положение этого рода до конца не выяснено, и в ряде систем его не относят к криптофитам), оспаривается и существование колониальных форм.
Клетки
У криптофит клетки обычно уплощённые, имеют выпуклую спинную и прямую или вогнутую брюшную стороны. Форма клеток разнообразная: яйцевидная, эллипсоидная, грушевидная, бобовидная, веретеновидная и др. Передний конец клетки более или менее скошен, от него отходит продольная борозда, которая не доходит до заднего конца. Наличие борозды и её длина — признаки, которые используются для систематики на уровне рода. Глотка мешковидная, по её краю располагаются трихоцисты. По периферии клетки также могут располагаться трихоцисты, но более мелких размеров. Трихоцисты (эжектосомы) окружены мембраной и внутри содержат 2 ленты, скрученные рулоном (цилиндры). Более крупный цилиндр состоит из намотанного по спирали лентовидного мембранного материала. Вверху он образует V-образное углубление, в котором находится второй цилиндр. Эти две части, по-видимому, связаны между собой с помощью волокон. При химическом и физическом раздражении туго скрученные ленты распрямляются, разрывая мембрану эжектосомы и перипласт. Сначала выбрасывается маленький цилиндр и, покидая клетку, он вытягивает за собой ленту большого цилиндра. Образуются эжектосомы в пузырьках аппарата Гольджи. Вначале цилиндры содержат несколько оборотов, но по мере роста увеличивают число оборотов.
Криптомонады очень чувствительны к изменению окружающих условий, таких, как изменение рН, осмотических условий или температуры. Они в массе отстреливают содержимое эжектосом, что приводит к быстрому разрушению клетки. С этим явлением связана значительная трудность по переводу криптомонад на лабораторные среды.
На переднем конце клетки расположены сократительные вакуоли, изливающие своё содержимое в глотку или борозду.
В клетках криптомонад имеются два тельца Mona, функции которых неизвестны. Так как тельца Mona содержат много мембран и фибрилл, они, возможно, вовлечены в процесс деструкции клеточных органелл, особенно отработанных трихоцист.
Клеточные покровы
У криптофит клеточные покровы представлены перипластом, который состоит из плазмалеммы и дополнительного белкового материала, расположенного над плазмалеммой. Внутриклеточный компонент перипласта может иметь форму чехла, как у Chilomonas, или состоять из пластинок (шестиугольных, как у Cryptomonas, или прямоугольных, как у Chroomonas). Белковые пластинки прикрепляются к плазмалемме с помощью округлых или многоугольных внутримембранных частиц. Наружный компонент перипласта может состоять из пластинок, чешуек, слизи или их комбинации. Перипласт у криптомонад отсутствует в области глотки.
Жгутиковый аппарат
На переднем конце клеток расположены два жгутика, длина которых сопоставима с длиной клетки. При движении они или оба направлены вперёд, или один — вперёд, а другой — назад. На жгутиках имеются особые криптофитовые мастигонемы, которые могут располагаться в два супротивных ряда на длинном жгутике (длина мастигонем до 2 мкм), в один ряд на коротком (длина мастигонем 1 мкм) либо на каждом жгутике. Мастигонемы криптомонад двухчастные, состоят из трубчатой части и тонкого терминального волоска. У некоторых видов короткий жгут свободен от мастигонем. На длинном жгутике при основании может находиться дополнительный пучок, состоящий приблизительно из 45 волосков. На жгутиках могут располагаться небольшие органические чешуйки в виде семиугольных розеток (140—170 нм в диаметре).
Переходная зона жгутиков содержит две или более пластинчатые структуры (септы) ниже точки, где заканчиваются две центральные микротрубочки. Корешковая система представлена ризостилем (состоит из 6—10 микротрубочек, соединённых сократительными фибриллами), который идёт в глубь клетки, трёх микротрубочковых корешков и фибриллярного корешка, прилегающего к одному из микротрубочковых. В разных родах имеются различия в строении корешкового аппарата жгутиков.
Глазок (стигма) у криптомонад связан с хлоропластом, но не связан со жгутиковым аппаратом, поэтому может располагаться и в центре клетки. Он состоит из большого числа пигментных глобул, расположенных параллельными рядами. Для некоторых криптомонад показан положительный фототаксис. Стигма встречается не у всех представителей.
Хлоропласты
В клетках криптомонад расположены один или два хлоропласта, реже больше, окрашенные в разные цвета — от сине-зелёного и оливково-зелёного до жёлто-бурого, коричневого и тёмно-красного. Иногда хлоропласты отсутствуют.
В хлоропластах присутствует ряд пигментов. Помимо хлорофилла а имеется хлорофилл с2, который передаёт энергию света от фикобилинов к хлорофиллу а. У криптомонад отсутствует аллофикоцианин, выполняющий эту функцию у красных и синезелёных водорослей. Фикобилипротеиновые пигменты криптомонад криптофикоциан и криптофикоэритрин отличаются организацией полипептидных компонентов и по спектральным характеристикам от фикобилипротеиновых пигментоы синезелёных и красных водорослей. Они локализованы не в фикобилисомах, а во внутритилакоидных пространствах, из-за этого тилакоиды криптомонад толще, чем у других водорослей. Каждая фотосинтезирующая криптомонада имеет только один тип фикобилипротеина — или фикоэритрин, или фикоцианин, но никогда вместе.
Криптомонады образуют несколько уникальных ксантофиллов, таких, как аллоксантин, который может быть использован для их обнаружения в смеси планктонных водорослей. У них встречаются α- и β-каротины, крококсантин, зеаксантин и монадоксантин.
В хлоропластах могут быть пиреноиды, в которые могут заходить ламеллы (каждая из 1—2 тилакоидов).
Оболочка хлоропласта состоит из четырёх мембран, наружная на своей поверхности несёт рибосомы. Между двумя парами мембран в оболочке хлоропласта криптомонад находится перипластидное пространство, в котором располагаются 80S рибосомы с эукариотным типом рРНК, зёрна крахмала и сильно редуцированное ядро, известное как нуклеоморфа. Нуклеоморфа окружена двухмембранной оболочкой с порами и содержит ядрышко. Размер ДНК нуклеоморфы составляет только 660 тыс. пар нуклеотидов. Она имеет всего три хромосомы (240, 225 и 195 тыс. пар нкулеотидов), которые кодируют процессы, необходимые только для функционирования нуклеоморфы. Нуклеоморфа делится во время деления ядра вслед за репликацией базального тела, но раньше, чем хлоропласт. Деление амитотическое, и хотя в её геноме обнаружены гены α-, β- и γ-тубулинов, веретено деления не образуется из микротрубочек, но имеются фибриллярные структуры, появляющиеся только во время деления. Дочерние нуклеоморфы мигрируют в противоположные концы пластиды, таким образом после деления пластиды каждая дочерняя пластида получает свою нуклеоморфу.
Наличие нуклеоморфы, перипластидного пространства и строение оболочек хлоропласта свидетельствуют о вторичном симбиотическом происхождении хлоропластов криптофитовых, когда бесцветная фаготрофная флагеллата поглотила («съела») эукариотную красную водоросль, которая затем трансформировалась в пластиду. Тогда две внутренние мембраны хлоропласта криптофитовых интерпретируют как мембраны хлоропласта красной водоросли, третью мембрану — как плазмалемму эндосимбионта (красной водоросли), наружную, четвёртую, как мембрану пищеварительной вакуоли клетки-хозяина. Симбиотическое происхождение хлоропласта криптомонад от красных водорослей подтверждается филогенетическим анализом, основанным на сравнении нуклеотидных последовательностей гена 16S рРНК нуклеоморфы и ядер красных водорослей.
Запасные продукты
Основное запасаемое вещество у криптофит — крахмал, откладывается между второй и третьей мембранами оболочки хлоропласта. Криптомонады — единственная группа водорослей с таким местом откладывания крахмала. Как и у красных водорослей, он содержит больше амилопектина (разветвлённые молекулы с α-1,4- и α-1,6-связями), чем амилозы (линейные цепочки с α-1,4-связями). С йодом такой крахмал даёт красное окрашивание. В цитоплазме могут присутствовать капли липидов.
Особенности питания
Среди криптомонад встречаются автотрофы, гетеротрофы (сапротрофы и фаготрофы) и миксотрофы. Большинство из них требуют витамин В12 и тиамин, некоторые нуждаются в биотине. Криптомонады могут использовать аммоний и органические источники азота, но морские представители менее способны к превращениям нитратов в нитриты по сравнению с другими водорослями. Органические вещества стимулируют рост криптомонад. Для различных бесцветных представителей и для окрашенного Cryptomonas ovata показана фаготрофность. Окрашенный в сине-зелёный цвет Chroomonas pochmanni обладает миксотрофным питанием. Для захвата и удержания бактериальных клеток образуются специализированные вакуоли. Бактерии втягиваются в вакуоль через маленькую пору в глотке, где отсутствуют перипластные пластинки. Перевариваются они в небольших вакуолях.
Ядро
Единственное ядро расположено в задней части клетки, или, реже, посередине. Митоз начинается после деления хлоропласта и имеет ряд особенностей: он открытый, ядрышко не исчезает, центриоли, по-видимому, отсутствуют. Область полюса веретена уплощена и ограничена цистернами эндоплазматической сети.
Число хромосом у криптомонад очень большое — от 40 до 210. В метафазе хромосомы объединяются в массивную пластинку. В ней образуются каналы, свободные от хроматина, через которые проходят микротрубочки веретена.
Размножение
Основной способ размножения криптомонад — вегетативный, за счёт деления клетки пополам с помощью борозды деления. При этом впячивание плазмалеммы идёт начиная с заднего конца клетки. Чаще всего делящаяся клетка сохраняет подвижность. Максимальная скорость роста для многих криптомонад — одно деление в день при температуре около 20 °С. Дефицит азота и избыточное освещение стимулируют образование покоящихся стадий. Покоящиеся цисты окружены толстым экстрацеллюлярным матриксом.
Для Ch. acuta известен хологамный половой процесс, при котором вегетативные клетки функционируют как гаметы. Слияние клеток происходит на брюшной стороне, причём задний конец одной клетки прикрепляется к середине другой. Когда клетки полностью сольются, формируется четырёхжгутиковая зигота. Затем она делится; первое деление, предположительно, мейотическое.
У криптомонады Proteomonas sulcata в определённой степени гетероморфный гаплодиплонтный жизненный цикл, в котором и диплоидная, и гаплоидная стадии жгутиковые. Эти стадии отличаются друг от друга по размерам клеток, структуре перипласта, конфигурации жгутикового аппарата. До сих пор в этом жизненном цикле для этого вида не известны оплодотворение и мейоз.
Экология и значение
Криптофитовые — типичные представители планктона, изредка они встречаются в иле солёных озёр и среди детрита в пресных водоёмах. Они занимают видное положение в олиготрофных, умеренных и высокоширотных, пресных и морских водах.
Большинство криптомонад встречается в широком температурном диапазоне, но некоторые виды Rhodomonas относятся к стенотермным. Среди стенотермных представителей криофильные (холодолюбивые) виды Cryptomonas способны вегетировать на поверхности льда и снега. Криптофиты играют важную роль в холодных водах, особенно зимой и ранней весной. Например, они могут доминировать в весеннем цветении фитопланктона в Северном море, где, возможно, являются важным звеном в пищевых цепях. Локализованные цветения, вызванные развитием криптомонад, встречаются в антарктических водах. Цветения воды коррелируют с притоком воды от таяния ледников. В покрытых льдами озёрах Антарктиды виды криптомонад могут доминировать в альгофлоре, давая более 70 % биомассы фитопланктона. Очень редко криптомонады встречаются в океанических водах при температуре 22 °С и выше. Они отсутствуют в горячих источниках и гиперсолёных водоёмах.
Пресноводные представители предпочитают искусственные и естественные водоёмы со стоячей водой — отстойники, различные пруды (биологические, технические, рыбоводные), реже встречаются в водохранилищах и озёрах. Они образуют крупные популяции в озёрах на глубине 15—23 м, в месте соединения слоёв воды, богатых и бедных кислородом, где уровень света значительно ниже, чем у поверхности. Бесцветные представители распространены в водах, загрязнённых органическими веществами, их много в сточной воде, поэтому они могут служить показателем загрязнения воды органическими веществами.
Среди криптофит встречаются типичные сфагнофиллы, живущие в болотах с низким значением рН, в то время как ряд видов может развиваться в широком диапазоне рН (например, Chilomonas встречается при рН 4,1—8,4).
Криптофиты играют значительную роль в водоёмах в круговороте кислорода, углерода, азота и фосфора, в синтезе органического вещества из минеральных веществ. Ими питаются различные представители зоопланктона, которыми в свою очередь питаются рыбы. Сообщалось об интенсивном поедании криптомонад кладоцерами в эвтрофных прудах в зимнее время года. В то же время отмечались случаи цветения воды в прудах, вызванных Cyanomonos, которые приводили к массовой гибели сомов.
Криптомонады обитают в качестве эндосимбионтов в миксотрофных инфузориях Myrionecta rubra (прежние названия — Mesodinium rubrum или Cyclotytrichum meunieri). Эта инфузория может образовывать нетоксичное красное цветение воды на побережье Перу, Калифорнии, Камчатки и др. (в основном в зоне апвеллинга). У неё отсутствует клеточный рот (у других представителей этого рода инфузорий, не имеющих эндосимбионтов, на переднем конце клетки имеется клеточный рот), и в своём питании она зависит от эндосимбионта, который на свету способен ассимилировать углерод и фосфор. Каждая инфузория может содержать несколько криптомонад, которые отделены от цитоплазмы хозяина двойной мембраной. Наружная мембрана рассматривается как мембрана пищеварительной вакуоли хозяина, а внутренняя — как плазмалемма симбионта. Симбионты содержат ядро, митохондрии, пластиды с нуклеоморфой и некоторые другие структуры. Хлоропласты имеют все характерные особенности хлоропластов криптофитов. В то же время у эндосимбионта отсутствует перипласт, жгутиковый аппарат, трихоцисты.
Некоторые динофиты (например, Dinophysis) приобрели хлоропласты в результате симбиоза с криптомонадами. В одних случаях у них может сохраняться ядро криптомонады, в других — только пластиды.
Эволюция и филогения
В эволюции криптофит исходными считаются формы, близкие к роду Goniomonas. Эти бесцветные представители способны к фагоцитозу, у них отсутствует нуклеоморфа и пластиды. Полагают, что пластиды криптомонад появились в результате вторичного симбиоза с эукариотом из Rhodophyta, что подтверждается филогенетическим анализом (сравнение пластидного генома криптомонад и красных водорослей, анализ генов нуклеоморфы, кодирующих α- и β-тубулины). Эволюция внутри криптофит выглядит следующим образом. Наличие борозды без глотки у Goniomonas — наиболее примитивная особенность. Глотка формировалась за счёт смыкания краёв борозды, начиная с заднего конца. Полное исчезновение борозды, но наличие глотки представлено у Chroomonas. В процессе эволюции криптомонад происходил повторный переход к гетеротрофности, что привело к образованию рода Chilomonas, у которого редуцирована борозда, развита глотка и имеются лейкопласты. Эта схема подтверждается эволюцией жгутикового аппарата и анализом нуклеотидных последовательностей гена 18S рРНК.
Положение криптомонад среди водорослей и их объём до сих пор дискутируются. А. Пашер полагал, что они произошли от хризомонад, и от криптомонад выводил перидинеи. Он объединил криптомонады и перидинеи в отдел Pirrophyta. Большинство альгологов не соглашаются с таким объдинением и исключают криптофитовых из пиррофитовых, рассматривая их как самостоятельную группу на основании того, что криптофиты отличаются от динофит строением ядра, жгутикового аппарата, наличием нуклеоморфы, составом пигментов, биохимическими особенностями. Все эти данные свидетельствуют против их близкого филогенетического родства.
Разнообразие
К настоящему времени описано более 200 видов криптофитовых (приблизительно 100 пресноводных и 100 морских). В последние годы в связи с использованием электронного микроскопа значительные изменения претерпела их систематика. Были переописаны и описаны новые таксоны, однако недостаточная конкретность и неоднозначность использованных признаков затрудняет не только определение родовой, но и видовой принадлежности этих организмов.
Систематика
- порядок Cryptomonadales
- род Campylomonas
- род Chilomonas
- род Chroomonas
- род Cryptomonas
- род Falcomonas
- род Geminigera
- род Guillardia
- род Hemiselmis
- род Plagioselmis
- род Proteomonas
- род Rhodomonas
- род Storeatula
- род Teleaulax
- порядок Goniomonadales
Примечания
dic.academic.ru
Криптофиты Википедия
Систе́ма жи́зненных форм Ра́ункиера — система, классифицирующая растения по положению и способу защиты почек возобновления в течение неблагоприятного периода (холодного или сухого). Предложена Х. Раункиером в 1905 году.
Раункиер классифицировал 5 основных типов жизненных форм, которые отражают разнообразие экологических условий, в которых сформировалась растительность. Подсчитывая процент видов, относящихся к той или иной жизненной форме, получают так называемые спектры жизненных форм в различных областях земного шара или в различных типах растительности планеты.
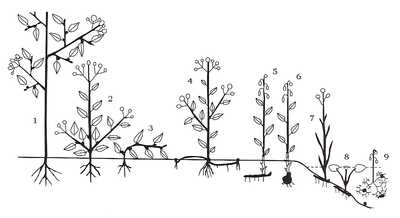 Жизненные формы растений по Раункиеру (терофиты, аэрофиты и эпифиты не показаны):
Жизненные формы растений по Раункиеру (терофиты, аэрофиты и эпифиты не показаны): - 1. Фанерофиты
- 2—3. Хамефиты
- 4. Гемикриптофиты
- 5—9. Криптофиты
- 5, 6. Геофиты
- 7. Гелофиты
- 8, 9. Гидрофиты
Фанерофиты
Фанерофиты — растения, у которых почки и концевые побеги, предназначенные для переживания неблагоприятного периода, расположены высоко над землёй. Этот тип разделяется на 15 подтипов и включает в себя деревья, кустарники и лианы. Подразделение на подтипы осуществляется в зависимости от размера (мега-, мезо-, микро-, нанофанерофиты), типа почек (с защищенными и открытыми почками) и листопадности (вечнозелёные и листопадные).
Хамефиты
Хамефиты — растения, у которых предназначенные для перенесения неблагоприятного периода почки и концевые побеги развиваются на побегах, лежащих на поверхности земли или расположенных близко к ней. В областях, где зимой поверхность земли покрыта снегом, он закрывает почки и концевые побеги, а в тёплых областях их частично закрывают отмершие остатки растений, лежащие на поверхности земли. Эту жизненную форму Раункиер подразделил на 4 подтипа: полукустарники, пассивные хамефиты, активные хамефиты и растения-подушки. Но существуют растения, очень похожие на хамефиты, не являющиеся ими. Важно уметь их различать.
Гемикриптофиты
Гемикриптофиты.
Эту жизненную форму автор подразделяет на 3 подтипа: протогемикриптофиты, частично розеточные гемикриптофиты и розеточные гемикриптофиты.
Криптофиты
Криптофиты — растения, у которых почки или окончания побегов, предназначенные для перенесения неблагоприятного периода, расположены в почве или на дне водоёма. Эта жизненная форма подразделяется на 3 подтипа: геофиты (корневищные, клубневые, луковичные, корневые геофиты)[1], гелофиты и гидрофиты.
Терофиты
Терофиты — растения, переживающие неблагоприятный сезон исключительно в виде семян. Сюда относятся все однолетние травы, крайней степенью в этой категории являются вынужденные терофиты (как правило, пустынь) — эфемеры.
Примечания
Литература
- Raunkiær Ch. Plant life forms / transl. from Danish by H. Gilbert-Carter. — Oxford : Clarendon Press, 1937. — vi, 104 p. (англ.)
wikiredia.ru
Криптофит Википедия
Систе́ма жи́зненных форм Ра́ункиера — система, классифицирующая растения по положению и способу защиты почек возобновления в течение неблагоприятного периода (холодного или сухого). Предложена Х. Раункиером в 1905 году.
Раункиер классифицировал 5 основных типов жизненных форм, которые отражают разнообразие экологических условий, в которых сформировалась растительность. Подсчитывая процент видов, относящихся к той или иной жизненной форме, получают так называемые спектры жизненных форм в различных областях земного шара или в различных типах растительности планеты.
Жизненные формы растений по Раункиеру (терофиты, аэрофиты и эпифиты не показаны): - 1. Фанерофиты
- 2—3. Хамефиты
- 4. Гемикриптофиты
- 5—9. Криптофиты
- 5, 6. Геофиты
- 7. Гелофиты
- 8, 9. Гидрофиты
Фанерофиты
Фанерофиты — растения, у которых почки и концевые побеги, предназначенные для переживания неблагоприятного периода, расположены высоко над землёй. Этот тип разделяется на 15 подтипов и включает в себя деревья, кустарники и лианы. Подразделение на подтипы осуществляется в зависимости от размера (мега-, мезо-, микро-, нанофанерофиты), типа почек (с защищенными и открытыми почками) и листопадности (вечнозелёные и листопадные).
Хамефиты
Хамефиты — растения, у которых предназначенные для перенесения неблагоприятного периода почки и концевые побеги развиваются на побегах, лежащих на поверхности земли или расположенных близко к ней. В областях, где зимой поверхность земли покрыта снегом, он закрывает почки и концевые побеги, а в тёплых областях их частично закрывают отмершие остатки растений, лежащие на поверхности земли. Эту жизненную форму Раункиер подразделил на 4 подтипа: полукустарники, пассивные хамефиты, активные хамефиты и растения-подушки. Но существуют растения, очень похожие на хамефиты, не являющиеся ими. Важно уметь их различать.
Гемикриптофиты
Гемикриптофиты.
Эту жизненную форму автор подразделяет на 3 подтипа: протогемикриптофиты, частично розеточные гемикриптофиты и розеточные гемикриптофиты.
Криптофиты
Криптофиты — растения, у которых почки или окончания побегов, предназначенные для перенесения неблагоприятного периода, расположены в почве или на дне водоёма. Эта жизненная форма подразделяется на 3 подтипа: геофиты (корневищные, клубневые, луковичные, корневые геофиты)[1], гелофиты и гидрофиты.
Терофиты
Терофиты — растения, переживающие неблагоприятный сезон исключительно в виде семян. Сюда относятся все однолетние травы, крайней степенью в этой категории являются вынужденные терофиты (как правило, пустынь) — эфемеры.
Примечания
Литература
- Raunkiær Ch. Plant life forms / transl. from Danish by H. Gilbert-Carter. — Oxford : Clarendon Press, 1937. — vi, 104 p. (англ.)
wikiredia.ru
Классификация микроводорослей
В основу разграничения водорослей по основным таксонам (царствам, отделам, классам и др.) положены следующие признаки: тип фотосинтетических пигментов, а следовательно, и окраска клеток; наличие жгутиков, их строение, количество и способ прикрепления к клетке; химический состав клеточной стенки и дополнительных оболочек; химический состав запасных веществ; число клеток в талломе и способ их взаимодействия.
К началу 20 века различали следующие классы водорослей по Пашеру:
– сине-зеленые водоросли – Cynophceae;
– красные водоросли – Rhodophyceae;
– зеленый водоросли – Chlorophyceae;
– золотистые водоросли – Chrysophyceae;
– желто-зеленые, или разножгутиковые, водоросли – Xanthophyceae, или Heterocontae;
– диатомовые водоросли – Bacillariophyceae, или Diatomeae;
– динофициевые водоросли – Dinophyceae;
– криптофициевые водоросли – Cryptophyceae;
– эвгленофициевые водоросли – Euglenophyceae.
Каждый класс характеризуется специфическим набором пигментов, запасным продуктом, откладывающимся в клетке в процессе фотосинтеза, и если есть жгутики, то их строением.
Прокариотические микроводоросли объединены в подцарство – цианеи (Cyanobionta). К нему относятся все сине-зеленые водоросли или цианеи. Это простого строения организмы приспособленные жить в воде. Исторические связи этих водорослей с бактериями проявляются в строении клеток. Но они отличаются от бактерий по наличию хролофилла "а" и очень редко – "b". В процессе фотосинтеза они выделяют кислород.
Отдел Cyаnophyta – сине-зеленые водоросли или цианеи
Большинство цианобактерий — облигатные фототрофы, которые, однако способны к непродолжительному существованию за счёт расщепления накопленного на свету гликогена в окислительном пентозофосфатном цикле и в процессе гликолиза.
Цианобактерии, по общепринятой версии, явились "творцами" современной кислородсодержащей атмосферы на Земле, что привело к "кислородной катастрофе" — глобальному изменению состава атмосферы Земли, произошедшему в самом начале протерозоя (около 2,4 млрд лет назад) которое привело к последующей перестройке биосферы и глобальному гуронскому оледенению. В настоящее время, являясь значительной составляющей океанического планктона, цианобактерии стоят в начале большей части пищевых цепей и производят значительную часть кислорода (вклад точно не определен: наиболее вероятные оценки колеблются от 20 % до 40 %). Цианобактерия Synechocystis стала первым фотосинтезирующим организмом, чей геном был полностью расшифрован. Рассматривается возможное применение цианобактерий в создании замкнутых циклов жизнеобеспечения, а также как массовой кормовой или пищевой добавки. Классификация:
– Порядок Chroococcales — Хроококковые:
Класс Gloeobacteria;
– Порядок Nostocales — Ностоковые;
– Порядок Oscillatoriales — Осциллаториевые;
– Порядок Pleurocapsales — Плеврокапсовые;
– Порядок Prochlorales — Прохлорофиты;
– Порядок Stigoneomatales — Стигонемовые.
Эукариотические микроводоросли одно- или многоклеточные, различно окрашенные, первично фотоавтотрофные растения, большей частью живущей в воде. Пластиды этих водорослей содержат хролофилл и чаще всего дополнительный хролофиллы "b", "с" каротиноиды, ксантофиллы и редко фикобилины. Донором электронов для фотосинтеза служит вода. Ранее водоросли классифицировали в составе Царства Растений, где они составляли обособленную группу. Однако с развитием молекулярно-генетических методов систематики стало ясно, что эта группа филогенетически очень неоднородна. В настоящее время водоросли относят к двум Царствам эукариот: Chromista и Protista.
Водоросли, относящиеся к Царству Chromista
Фотосинтезирующие хромисты обычно содержат в хлоропластах не свойственный растениям каротиноид фукоксантин, а иногда и другие специфические пигменты, а также хлорофилл с. Еще одной особенностью клеток хромистое является наличие двух эукариотических жгутиков, один из которых обычно перистый – имеет трубчатые ответвления основной нити. Хлоропласт и ядро чаще окружены общей мембраной, при этом в хлоропласте имеются светочувствительные гранулы (стигма), воспринимающие изменения интенсивности освещения и обусловливающие фототаксис. Запасными веществами хромистое является не крахмал, как у растений, а жироподобное вещество лейкозин, полисахарид ламинарии или другие специфические полисахариды.
– Подцарство Водоросли (Phycobionta):
Отдел Bacillariophyta– диатомовые водоросли:
Являясь важнейшей составляющей морского планктона, диатомовые создают до четверти всего органического вещества планеты.
Только коккоиды, форма разнообразна. В основном одиночные, реже — колониальные. Большинство представителей этого отдела одноклеточные, однако встречаются ценоцитные и нитчатые формы. Диатомовые водоросли играют очень важную роль в трофических связях водных организмов, являясь основным компонентом фитопланктона, а также придонных осадков. Будучи фотосинтезирующими организмами, они служат главным источником пищи для пресноводных и морских животных. Считается, что на их долю приходится до четверти всего совершаемого на нашей планете фотосинтеза.
Хлоропласты диатомей содержат хлорофиллы а и с, а также фукоксантин. Размножение в основном бесполое — путем деления клетки. Запасным веществом служит лейкозин.У диатомовых водорослей жгутиковая стадия представлена только мужскими гаметами (у некоторых видов). Поэтому подвижные формы передвигаются за счет направленного перетекания цитоплазмы в районе шва панциря, в котором цитоплазма и мембрана граничат с окружающей средой. Эти организмы имеют уникальные двустворчатые панцири, состоящие из кремнезема (SiO2∙nН2O) и формирующие две половинки, вставляющиеся одна в другую. Створки панциря имеют тонкую орнаментацию и выглядят очень красиво. Известно более 10 тысяч видов диатомей, которые отличаются поразительным разнообразием и тончайшей филигранностью.
При отмирании клеток кремниевые скелеты не разрушаются, они накапливаются в течение десятков миллионов лет на дне водоемов. Эти отложения называют "диатомовым илом" и используют в качестве абразивного материала для полировки, а также для фильтрования.
Отдел Chrysophyta – золотистые водоросли:
Включают в себя преимущественно микроскопические водоросли различных оттенков жёлтого цвета.
Большинство золотистых водорослей — одноклеточные, реже колониальные, ещё реже многоклеточные организмы.
В основном золотистые водоросли — миксотрофы, то есть, имея пластиды, способны поглощать растворённые органические соединения и/или пищевые частички. У некоторых тип питания (автотрофный, миксотрофный или гетеротрофный) зависит от условий окружающей среды или клеточного состояния.
Вегетативное размножение происходит путём продольного деления клетки пополам или фрагментами колонии слоевища. Бесполое размножение осуществляется с помощью одно- или двужгутиковых зооспор, или, реже, апланоспор и амёбоидов. Половое размножение лучше всего описано у представителей с домиками благодаря хорошо наблюдаемому образованию зигот. Клетки прикрепляются друг к другу в зоне отверстия домика, и их протопласты сливаются, образуя зиготу.
Насчитывается более тысячи описанных видов золотистых водорослей, большинство из которых представлено подвижными за счет жгутиков одноклеточными формами, однако встречаются и нитевидные, и колониальные виды. Некоторые представители имеют амебоидное строение клеток и отличаются от амеб только наличием хлоропластов.
Многие хризофиты лишены клеточной стенки, но имеют кремнеземовые чешуйки или скелетные элементы. Запасным веществом служит хризоламинарин. Фотосинтетические пигменты представлены хлорофиллами а и с, а также каротинами и ксантофиллами, которые придают клеткам золотисто-коричневый оттенок.
Золотистые водоросли, как правило, обитают в планктоне, но встречаются и донные, прикреплённые формы. Входят в состав нейстона. Большинство золотистых водорослей встречается в основном в пресноводных бассейнах умеренного климата, достигая наибольшего видового многообразия в кислых водах сфагновых болот, что связано с образованием у них кислых, а не щелочных фосфатаз. Они необычайно требовательны к содержанию железа в воде, которое используют для синтеза цитохромов. Меньшее количество видов обитает в морях и солёных озёрах, единичные обнаружены в почве. Максимума развития золотистые водоросли достигают в холодное время года: в планктоне они доминируют ранней весной, поздней осенью и зимой. В это время они играют значительную роль как продуценты первичной продукции и служат пищей зоопланктону. Некоторые золотистые водоросли (р. Uroglena, Dinobryon, Mallomonas, Synura; Prymnesium parvum), развиваясь в массовом количестве, способны вызывать цветение воды.
Цисты золотистых водорослей, встречаемые в осадках со дна водоёмов, используют в качестве экологических индикаторов для изучения условий окружающей среды в прошлом и настоящем. Золотистые водоросли улучшают газовый режим водоёмов, имеют значение в образовании илов и сапропелей.
Отдел Cryptophyta – криптофитовые водоросли:
Криптофиты — небольшая, но экологически и эволюционно очень важная группа организмов, обитающих в морских и континентальных водах. Почти все они — одноклеточные подвижные жгутиконосцы, некоторые представители способны формировать пальмеллоидную стадию. Только один род Bjornbergiella (выделен из почв Гавайских островов) способен к образованию простых нитчатых талломов (положение этого рода до конца не выяснено, и в ряде систем его не относят к криптофитам), оспаривается и существование колониальных форм.
Среди криптомонад встречаются автотрофы, гетеротрофы (сапротрофы и фаготрофы) и миксотрофы. Большинство из них требуют витамин В12 и тиамин, некоторые нуждаются в биотине. Криптомонады могут использовать аммоний и органические источники азота, но морские представители менее способны к превращениям нитратов в нитриты по сравнению с другими водорослями. Органические вещества стимулируют рост криптомонад.
Основной способ размножения криптомонад — вегетативный, за счёт деления клетки пополам с помощью борозды деления. При этом впячивание плазмалеммы идёт начиная с заднего конца клетки. Чаще всего делящаяся клетка сохраняет подвижность. Максимальная скорость роста для многих криптомонад — одно деление в день при температуре около 20 °С. Дефицит азота и избыточное освещение стимулируют образование покоящихся стадий. Покоящиеся цисты окружены толстым экстрацеллюлярным матриксом.
Криптофитовые — типичные представители планктона, изредка они встречаются в иле солёных озёр и среди детрита в пресных водоёмах. Они занимают видное положение в олиготрофных, умеренных и высокоширотных, пресных и морских водах.
Пресноводные представители предпочитают искусственные и естественные водоёмы со стоячей водой — отстойники, различные пруды (биологические, технические, рыбоводные), реже встречаются в водохранилищах и озёрах. Они образуют крупные популяции в озёрах на глубине 15—23 м, в месте соединения слоёв воды, богатых и бедных кислородом, где уровень света значительно ниже, чем у поверхности. Бесцветные представители распространены в водах, загрязнённых органическими веществами, их много в сточной воде, поэтому они могут служить показателем загрязнения воды органическими веществами.
Среди криптофит встречаются типичные сфагнофиллы, живущие в болотах с низким значением рН, в то время как ряд видов может развиваться в широком диапазоне рН.
Отдел Haptophyta – гаптофитовые водоросли:
Гаптофиты — это группа аутотрофных, осмотрофных или фаготрофных простейших, которые населяют морские экосистемы. Гаптофиты, как правило, одноклеточные, но встречаются и колониальные формы. Несмотря на маленький размер, эти организмы играют очень большую и важную роль в геохимических циклах углерода и серы.
Многие гаптофиты помимо фототрофии способны к осмотрофному и фаготрофному поглощению питательных веществ, таким образом, миксотрофия для них — обычное явление.
Большинство примнезиофитов обитает в морях, предпочитая открытые зоны, лишь немногие встречаются в пресных и солоноватых водоёмах. Наибольшего биоразнообразия примнезиофиты достигают в водах, содержащих минимальное количество биогенов, субтропических океанических открытых водах, где встречаются даже на глубине более 200 м.
Некоторые примнезиофиты играют отрицательную роль в природе. Так, виды, образующие кокколиты, участвуют в глобальном потеплении климата как ключевые элементы в глобальном балансе улекислого газа. Они могут вызывать "цветение" воды, которое из-за способности кокколитов отражать свет, называют "белым".
Отдел Xanthophyta – желто-зеленые водоросли:
Водоросли, хлоропласты которых окрашены в жёлто-зелёный или жёлтый цвет.
Среди ксантофитов встречаются одноклеточные жгутиковые формы, а также колониальные, нитчатые и ценоцитные. Последние представлены широко распространенным родом Vaucheria ("водяной войлок"). Эти ценоцитные нитчатые слабоветвящиеся водоросли часто обнаруживаются на периодически затопляемом прибрежном иле.
У большинства жёлто-зелёных известно вегетативное и бесполое размножение.
Жёлто-зелёные водоросли входят в различные экологические группы — планктон, реже в перифитон и бентос. Подавляющее большинство из них — свободноживущие формы.
Водоросли, относящиеся к Царству Protista
Фотосинтезирующие Протесты в совокупности с гетеротрофными Протистами входят в состав смешанных типов – Dinoflagellata (динофлагелляты) и Euglenida (эвгленовые), а также представлены крупными группами зеленых и красных водорослей. Динофлагелляты. Относящиеся к типу Dinoflagellata водоросли иначе называют огненными (Pyrrhophyta) за способность к биолюминесценции – явлению свечения, или испускания видимого света.
Большинство огненных водорослей представляют собой одноклеточные формы с двумя жгутиками, часто причудливой и очень разнообразной формы, с плотными целлюлозными пластинками, образующими клеточную стенку в виде шлема или доспехов. Некоторые имеют довольно большие размеры, достигая 2 мм в диаметре. Из-за наличия под плазматической мембраной большого количества слоистых ячеек (альвеол) эти водоросли относят к особой группе Протистов – альвеолятам.
Фотосинтезирующие динофлагелляты обычно содержат хлорофиллы а и с, а также каротиноиды, их клетки бывают окрашены в золотистые и буро-коричневые тона. Запасным веществом является крахмал. Эти водоросли часто вступают в симбиотические отношения с морскими организмами (губками, медузами, актиниями, кораллами, кальмарами и др.). В этом случае они утрачивают целлюлозные пластинки и выглядят как золотистые сферические клетки, называемые зооксантеллами. В таких симбиотических системах животное обеспечивает динофлагелляты углекислотой, необходимой им для фотосинтеза, и предоставляет защиту, а водоросли снабжают животное кислородом и органикой.
Основной способ бесполого размножения динофлагеллят – продольное деление, реже встречается формирование зооспор. Некоторые виды способны к половому размножению в ходе изогамии, иногда – анизогамии.
Известно около 2000 видов ныне живущих динофлагеллят, обитающих чаще в морских, реже – в пресноводных водоемах. Фотосинтезирующие представители типа являются высокопродуктивными компонентами морского планктона, способными, однако, вызывать массовые вспышки заболевания и гибели рыбы, моллюсков, других животных. Оно объясняется необычайно бурным развитием некоторых огненных водорослей, способных образовывать яды, относящиеся к числу мощных нервных токсинов. В результате наносится огромный ущерб морским промыслам, а кроме того, получают отравления люди, употребляющие в пищу рыбу или моллюсков, питавшихся ядовитыми водорослями.
Отдел Chlorophyta – зеленые водоросли:
Самый обширный на данное время отдел водорослей. По приблизительным подсчётам сюда входит около 500 родов и от 13 000 до 20 000 видов. Все они отличаются в первую очередь чисто-зелёным цветом своих слоевищ, сходным с окраской высших растений и вызванным преобладанием хлорофилла над другими пигментами. Чрезвычайно велик и диапазон их размеров — от нескольких микрон до нескольких метров.
Преобладающими пигментами хлоропластов (как и у растений) являются хлорофиллы а и Ь, отчего талломы окрашены в зеленый цвет. Каротиноиды в хлоропластах многих одноклеточных Зеленых водорослей формируют скопление в виде "глазка" (стигмы). Многие виды содержат в клетках сократительные вакуоли, участвующие в осморегуляции. Одноклеточные формы обычно подвижны за счет двух одинаковых жгутиков, причем жгутики не покрыты трубчатыми ответвлениями, как у Хромистов.
Основным резервным материалом зеленых водорослей является крахмал, а клеточные стенки большинства видов состоят из целлюлозы. Эти признаки в совокупности с химическим составом фотосинтезирующих пигментов и некоторыми особенностями строения отдельных клеточных элементов делают зеленые водоросли очень похожими на растения. Кроме того, как и у растений, у зеленых водорослей наблюдается смена генераций в жизненном цикле. Подобное сходство позволяет считать зеленые водоросли непосредственными предками наземных растений. Исследование малых рРНК показало, что отдельные представители данной группы, в частности харовые водоросли, даже ближе по степени филогенетического родства к растениям, чем к другим водорослям.
Размножение зелёных водорослей бывает вегетативным, бесполым и половым.
Зелёные водоросли широко распространены по всему миру. Большинство из них можно встретить в пресных водоёмах (представители харофитов и хлорофициевых), но немало солоноватоводных и морских форм (большинство представителей класса ульвофициевых). Среди них есть планктонные, перифитонные и бентосные формы. Есть зелёные водоросли, которые приспособились к жизни в почве и наземных местообитаниях. Их можно встретить на коре деревьев, скалах, различных постройках, на поверхности почв и в толще воздуха. Массовое развитие микроскопических зелёных водорослей вызывает "цветение" воды, почвы, снега, коры деревьев и т. д.
Существуют паразитические представители зелёных водорослей, большинство из которых в качестве хозяина имеют высшие растения.
Отдел Euglenophyta – эвгленовые водоросли:
У эвгленовых форма тела варьирует от веретеновидной, овальной до плосколистовидной и игловидной. Передний конец тела более или менее закруглён, задний может быть вытянутым и заканчиваться заострённым отростком. Клетки могут быть спирально скручены. Длина клеток от 5 до 500 мкм и более.
Эвглениды имеют 1, 2, 3, 4 и 7 видимых жгутиков, за исключением небольшой группы безжгутиковых форм, а также прикрепленных организмов. Жгутики отходят от колбообразного впячивания на переднем конце клетки — глотки (ампулы).
Светочувствительная система эвгленовых состоит из двух структур. Первый компонент — это парафлагелларное тело (парабазальное вздутие), представляет собой вздутие при основании одного видимого жгутика и содержит синие светочувствительные флавины. Второй компонент системы — глазок (стигма), расположенный в цитоплазме около резервуара напротив парафлагелларного тела.
Эвгленовым водорослям свойственно автотрофное и гетеротрофное (сапротрофное) питание. В последнем случае питательные вещества поступают в клетку в растворенном виде, всасываясь всей ее поверхностью (осмотрофный тип). Некоторые виды характеризуются также фаготрофным способом питания. Известны ауксотрофные представители эвглен, зависимые по витаминам В12 и В,.
Если эвглен долго культивировать в подходящей питательной среде в темноте, они могут утрачивать хлоропласты и неограниченно долго демонстрировать гетеротрофный тип питания, ничем не отличаясь в этом случае от простейших. Таким образом, эвглен можно считать простейшими с нестабильным наследованием хлоропластов.
Эвгленовые водоросли обитают в основном в пресных водах, предпочитая водоёмы с замедленным стоком и богатым содержанием органических веществ. Их можно обнаружить в прибрежье озёр и рек, в мелких водоёмах, включая лужи, на рисовых полях, на сырой почве. В почвах бесцветные представители встречаются на глубине 8-25 см. Окрашенные эвгленовые могут вызывать цветение воды, образуя на её поверхности плёнку зелёного или красного цвета.
В значительной степени эвгленовые водоросли реагируют на степень минерализации воды: чем она выше, тем беднее их качественный и количественный состав. Некоторые выдерживают высокую солёность воды.
Среди эвгленовых встречаются фотоавтотрофы, гетеротрофы (фаготрофы и сапротрофы) и миксотрофы. Только треть родов способны к фотосинтезу, а остальные — фаготрофы и осмотрофы. Даже фотосинтезирующие эвгленовые способны к гетеротрофному росту. Большинство гетеротрофных форм — сапротрофы, поглощающие растворённые в воде питательные вещества.
Отдел Dinophyta – динофитовые водоросли:
Большинство представителей — двусторонне-симметричные или асимметричные жгутиконосцы с развитым внутриклеточным панцирем.
Размножаются вегетативным, бесполым и половым способом.
Морские и пресноводные формы, планктон и бентос, иногда встречаются как симбионты растений, иногда — паразиты ракообразных и червей. Могут вызывать цветение воды. Динофиты — важная группа фитопланктона в морских и пресных водах. Их значение прежде всего определяется тем, что они занимают второе место после диатомей как продуценты первичной продукции в прибрежных морских водах, являются пищей для различных простейших, коловраток, рыб. Большинство динофит (около 90 % видов) обитают в морских водах.
– Подцарство Багряники (Rhodobionta):
Отдел Rhodophyta – красные водоросли:
Обычно это довольно крупные растения, но встречаются и микроскопические. Среди красных водорослей имеются одноклеточные (крайне редко), нитчатые и псевдопаренхимные формы, истинно паренхимные формы отсутствуют. Ископаемые остатки свидетельствуют, что это очень древняя группа растений. Обычно это довольно крупные растения, но встречаются и микроскопические.
Для красных водорослей характерен сложный цикл развития, не встречающийся у других водорослей.
Отдел красных водорослей (Rhodophyta) включает виды, в клетках которых есть особый класс фотосинтетических пигментов – фикобилинов (фикоцианин и фикоэритрин), придающих им красную окраску (поэтому их называют багрянками). Эти вспомогательные пигменты маскируют цвет основного фотосинтетического пигмента – хлорофилла а. Преобладающим резервным веществом багрянок является крахмалоподобный полисахарид. Клеточные стенки этих водорослей содержат целлюлозу или другие полисахариды, погруженные в слизистый матрикс, который представлен, в свою очередь, агаром или карраги-наном. Эти компоненты делают красные водоросли гибкими и скользкими на ощупь. Некоторые багрянки откладывают в клетках карбонат кальция, что придает им жесткость. Такие формы играют важную роль в формировании коралловых рифов.
Красные водоросли не имеют жгутиков, большинство ведет неподвижный образ жизни в прикрепленном к камням или другим водорослям состоянии.
В Баренцевом море красные водоросли — типичные представители прибрежной бентосной растительности.
Некоторые виды красных водорослей употребляются в пищу. Из красных водорослей также получают гелеобразующее вещество агар-агар.
biofile.ru
Адвентивный компонент флоры города Калининграда
Введение
Город Калининград — административный центр самого западного региона России, единственного субъекта РФ, полностью отделенного от остальной территории страны сухопутными границами иностранных государств (Польша, Литва) и международными морскими водами. Калининград расположен в Приморской низменности, по берегам реки Преголя, при ее впадении в Калининградский залив.
В городе расположен единственный незамерзающий порт России на Балтийском море.
Железнодорожные и автомобильные трассы соединяют причалы порта напрямую со всей сетью транспортных путей России и других стран, а судоходные морские линии — с портами Германии, Дании, Голландии, Бельгии, Великобритании [5].
В настоящее время на территории г. Калининграда, развернуто массовое строительство жилых и деловых комплексов, ремонт и расширение дорог, строительство новых мостов. Заключительным этапом облагораживания новых и реконструированных участков идет озеленение, замена поврежденных и старых древесных растений и посадка новых, оформление клумб и газонов. Для проведения этих работ производится закупка посевного материала в Западной Европе, вместе с семенами декоративных видов и культивируемых злаков, входящих в состав газонной травы к нам в город попадают адвентивные растения.
Цель исследования
Целью нашего исследования является изучение адвентивных растений города Калининграда.
Объекты и методы исследования
Работа проводилась на территории всего города, особое внимание уделялось местам разгрузок автомобильного транспорта и железнодорожных составов, железнодорожным станциям, территории порта, газонным покрытиям улиц и скверов, а также свалкам садовых обществ, находящихся в черте города.
Для проведения работы использовался метод маршрутов. Для каждой экскурсии разрабатывался свой маршрут. В него включались территории с наиболее выраженной антропогенной нагрузкой и большей вероятностью нахождения адвентивных видов: узловые развязки и места разгрузки железнодорожного транспорта, территории складских помещений, новостройки, реконструированные скверы, новые газоны и клумбы. Работа проводилась в 2010—2014 полевых сезонах.
Материалы собственных исследований, дополнены литературными данными и материалами фондового гербария кафедры биоэкологии и биоразнообразия химико-биологического института Балтийского Федерального Университета имени Иммануила Канта [2, 3, 4, 6, 7].
Результаты исследования и их обсуждение
По нашим данным, на настоящее время флора г. Калининграда включает 201 вид адвентивных сосудистых растений, относящихся к 142-м родам и 44-м семействам.
Ядро адвентивной флоры представлено 8-ю семействами: Asteraceae — 35 видов, Poaceae — 29 видов, Brassicaceae — 24 вида, Fabaceae — 16 видов, Labiatae — 9 видов, Rosaceae — 7 видов, Polygonaceae — 6 видов, Scrophulariaceae — 6 видов, на долю которых приходится 66 % видового разнообразия адвентивного компонента.
В целом, в составе адвентивной фракции преобладают семейства с небольшим числом видов: 20 семейств по 1-му виду, в 12-ти семействах по 2—3 вида, 4 семействах по 4—5 видов.
Для выявления экологических особенностей изучаемой флоры нами были выделены группы видов растений с различным отношением к факторам увлажнения, освящения и активного богатства почв.
По отношению к фактору увлажнения были выделены следующие экологические группы растений: мезофиты — 151 вид (75 %), к группе ксеромезофитов относятся 20 видов (10 %), к группам мезоксерофиты и ксерофиты по 13 видов (6,5 %), гигромезофиты и гидрофиты по 2 вида (1 %). Преобладание группы мезофитов достигается за счет присутствия благоприятных условий увлажнения. Данная группа видов встречается преимущественно в парках, реконструированных скверах, на неухоженных газонах, на придомовых территориях старой застройки, в садовых обществах.
Адвентивная флора г. Калининграда характеризуется доминированием светолюбивых видов, встречающихся преимущественно на открытых хорошо освещенных экотопах (улицах, газонах, пустырях, железных и автодорогах).
По отношению к активному богатству почв выделены следующие экологические группы растений: мезотрофные — 161 вид (80 %), олиготрофные — 25 видов (12 %), эвтрофные — 15 видов (8 %). Повышенное участие в адвентивной фракции мезотрофных видов объясняется преобладанием среднеплодородных почв на территории города.
Биоморфологический анализ показал, что согласно классификации И. Г. Серебрякова в адвентивном компоненте флоры преобладают однолетние виды — 104 (52 %). Многолетние травы стоят на втором месте — 59 видов (29 %), затем двулетние растения — 19 (9,5 %) и кустарники — 16 (7,5 %). К деревьям относятся 3 вида (2 %): Sorbus intermedia (Ehrh.) Pers., Acer negundo L., Acer pseudoplatanus L (рисунок 1).
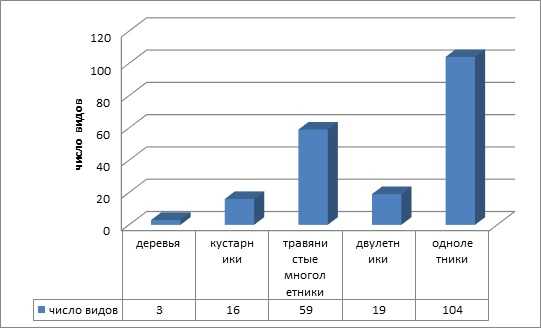
Рисунок 1 — Соотношение жизненных форм по И. Г. Серебрякову
По способу перенесения не благоприятных условий в соответствии с классификацией К. Раункиера, ведущее место занимают терофиты — 104 вида (52 %). Далее следуют гемикриптофиты 74 вида (37 %) и фанерофиты 19 видов (9 %). Группа криптофитов представлена 4-мя видами (2 %): Armoracia rusticana Gaerth., Mey. et Scherb, Heliantus tuberosus L., Elodea сanadensis Michx.,Ornithogalum umbellatum L (рисунок 2).
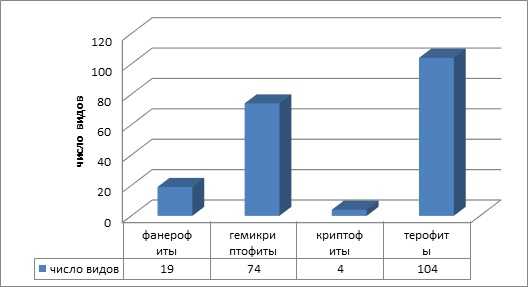
Рисунок 2 — Соотношение жизненных форм по К. Раункиеру
В адвентивном комплексе ведущими группами являются однолетние и многолетние травы, терофиты и гемикриптофиты. Адвентивные древесные виды — фанерофиты представлены интродуцированными деревьями и кустарниками, дичающими после посадок. Сравнение спектров жизненных форм показывает их стабильность.
В географической структуре адвентивной флоры, представлены виды, принадлежащие к 6-ти широтным и 11-ти долготным географическим элементам. Преобладают умеренный и южно-умеренный широтные и евроазиатский и европейский долготные географические элементы (рисунки 3, 4)
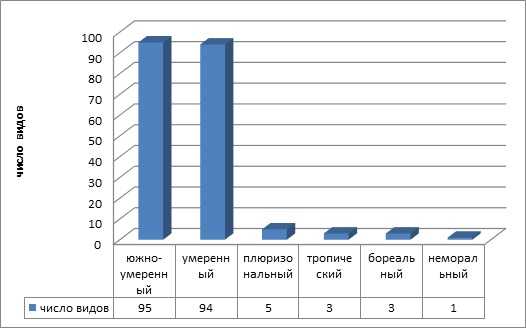
Рисунок 3 — Широтные географические элементы адвентивной фракции флоры
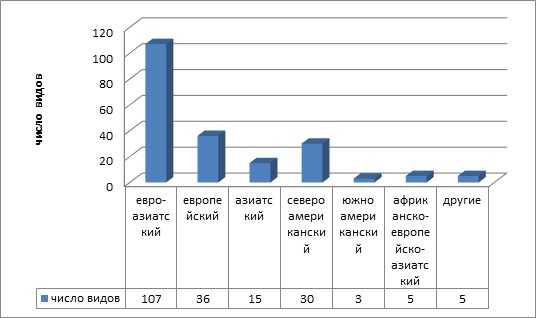
Рисунок 4 — Долготные географические элементы адвентивной фракции флоры
Именно умеренные и южно умеренные зоны Евразии являются главным источником поступления заносных видов на территорию региона. Кроме того, евроазиатские виды в силу своих эколого-биологических особенностей могут успешно закрепляться во флоре. Интенсивность заноса на городские территории азиатских, американских, африканских видов и возможность их произрастания, ниже, но они также представлены в урбанофлоре.
Анализ соотношения групп адвентивных видов флоры по времени заноса на новую территорию показывает, что главной группой являются неофиты — 184 вида, а среди них отмечены новейшие элементы флоры — эунеофиты — 10 видов. Группу археофитов представляют 7 видов распространенных на пустырях, обочинах дорог, на неухоженных газонах, в скверах, парках обычные для региона сорные растения. Расширение адвентивной фракции урбанофлоры во многом связано с застройкой городских территорий с организацией новых газонов, расширением площадей свалок, пустырей, неухоженных дворов и зеленых насаждений.
По типу осваиваемых местообитаний виды адвентивной фракции разделены на 5 групп (рисунок 5): рудеральные — 15 видов (7,5 %), сегетально-рудеральные — 6 видов (3 %), сегетальные — 14 видов (7 %), садово-парковые (дичающие) — 76 (37,5 %), случайно занесенные — 90 видов (45 %).
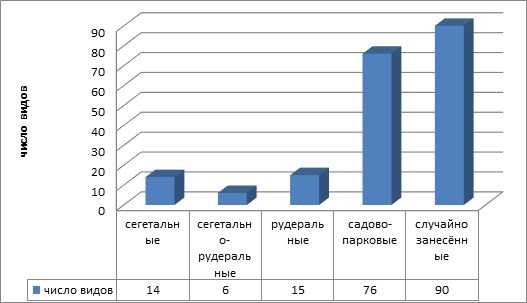
Рисунок 5 — Соотношение экотопных групп адвентивных растений
В урбанофлоре г. Калининграда основной группой являются случайно занесённые виды. Виды этой группы не всегда имеют устойчивое возобновление на городских территориях и представляют собой непостоянный компонент адвентивной фракции урбанофлоры. Второй по значению группой являются садово-парковые, дичающие из культуры виды. Они адаптируются к новым условиям произрастания, что подтверждается наличием семенного или вегетативного размножения. Именно такие виды необходимо использовать в озеленении городов, они хорошо приспособлены к городским условиям, успешно существуют в насаждениях при минимальном уходе или даже при его отсутствии [1].
Основным способом поступления адвентивных видов на городские территории, является непреднамеренный занос. Большая часть адвентивных видов принадлежит к группе ксенофитов. Это растения спутники хозяйственной деятельности человека (Galinsoga parviflora Cav., Bunias orientalis L., Sisymbrium altissimum L., Sisymbrium loeselii L., Euphorbia virgata Waldst.et Kit., Papaver rhoeas L.), приуроченные к вторичным синантропным местообитаниям. Второй источник обогащения городской флоры адвентивными видами — интродукция. Распространение эргазиофитов (Hordeum jubatum L., Saponaria officinalis L., Dianthus barbatus L.), приурочено главным образом к паркам, скверам, дворам, свалкам садовых обществ.
Из четырех групп адвентивных видов по результатам внедрения в урбанофлору преобладают две — эпекофиты и эфемерофиты. К первым относятся многочисленные сорные растения, распространяющиеся семенным и вегетативным путем только во вторичных экотопах и не внедряющиеся в естественные фитоценозы. Вторые не имеют устойчивого возобновления на новой для них территории и являются не постоянным компонентом адвентивной флоры. Колонофиты, приуроченные к изменённым экотопам и возобновляющиеся только вегетативно. Агриофиты — виды становящиеся полноправными членами естественных сообществ (Heracleum sosnowskyi Manden., Elodea сanadensis Michx.).
Выводы
Итогом работы стал конспект адвентивных видов сосудистых растений, произрастающих на территории города Калининграда. Конспект содержит сведения о 201 виде сосудистых растений изм 142-х родов и 44-х семейств.
В процессе работы был проведен систематический анализ. Наибольшее количество семейств, родов и видов относится к отделу Magnoliophyta, среди которых преобладают представители класса Magnoliopsida. Представители класса Liliopsida составляют 16 % от общего числа видов флоры.
Среди адвентивной фракции выделены 8 семейств, представленных наибольшим количеством видов, и составляющих 66 % от общего числа видов адвентивной фракции.
Экологическая структура адвентивной флоры выражена преобладанием светолюбивых мезофитных мезотрофных видов. Это виды, которые в силу экологических особенностей и низкой конкурентной способности практически не внедряются в естественные фитоценозы, но успешно заселяют открытые вторичные экотопы, формирующиеся в ходе хозяйственной деятельности.
В биоморфологическом анализе главную роль играют однолетние травы, терофиты. Они представлены 104-мя видами и составляют 52 %.
Географический анализ показал, что наибольшее количество видов адвентивной фракции флоры принадлежат к южно умеренному, умеренному Европейско-Азиатскому географическому элементу.
В комплексном анализе видов адвентивной флоры по времени заноса преобладают неофиты. Основными экотопными группами являются случайно занесенные и садово-парковые (дичающие) виды. Главными группами урбанофлоры являются ксенофиты — эпекофиты и ксенофиты — эфемерофиты, которые вместе составляют 72 % адвентивных видов.
Литература
- Антипина, Г. С. Урбанофлора Карелии. Монография // Петрозаводск. Изд-во ПетрГУ. 2002. — 200 с.
- Губарева, И. Ю. Некоторые дополнения к адвентивной флоре Калининградской области // Ботан. журн. — 1998. — Т.83, № 8. — С. 25.
- Гуджинскас, З. А. Дополнение к адвентивной флоре Калининградской области семейства Poaceae // Ботан. журн. — 1991. — Т.76, № 10. — С.1441—1446.
- Гусев, Ю. Д. Новые данные по адвентивной флоре Калининградской области // Ботан. журн. — 1974. — Т. 76, № 10. — С. 1458—1460.
- Паспорт городского округа «Город Калининград» // Калининград. Изд-во Калининград. 2013 — 83 с.
- Победимова, Е. Г. Состав, распределение и хозяйственное значение флоры Калининградской области // Тр. Бот. инст. АН СССР. — 1955. — Сер. 3(геобот.), Вып. 10. — С. 285—329.
- Abromeit J., Neuhoff W., Steffen M., Senh 2 ch A., Vogel G. Flora von Ost — und Westpreussen. // Bd. Berlin. 1903. — 1248 s.
Literature
- Antipina, G. S. Urbanoflora Karelii // Monografiya.-Petrozavodsk. Izd-vo PetrGU. 2002. — 200 s.
- Gubareva, I. Yu. Nekotorye dopolneniya k adventivnoy flore Kaliningradskoy oblasti // Botan. zhurn. — 1998. — T.83, № 8. — S. 25.
- Gudzhinskas, Z. A. Dopolnenie k adventivnoy flore Kaliningradskoy oblasti semeystva Poaceae // Botan. zhurn. — 1991. — T.76, № 10. — S. 1441—1446.
- Gusev, Yu. D. Novye dannye po adventivnoy flore Kaliningradskoy oblasti // Botan. zhurn. — 1974. — T.76, № 10. — S. 1458—1460.
- Pasport gorodskogo okruga «Gorod Kaliningrad» // Kaliningrad. Izd-vo Kaliningrad. 2013 — 83 s.
- Pobedimova, E. G. Sostav, raspredelenie i khozyaystvennoe znachenie flory Kaliningradskoy oblasti // Tr. Bot. inst. AN SSSR. — 1955. — Ser. 3(geobot.), Vyp. 10. — S. 285—329.
- Abromeit J., Neuhoff W., Steffen M., Senh 2 ch A., Vogel G. Flora von Ost — und Westpreussen. // Bd. Berlin. 1903. — 1248 s.
Библиографическая ссылка
Мацкова С. В., Адвентивный компонент флоры города Калининграда // «Живые и биокосные системы». — 2014. — № 7; URL: http://www.jbks.ru/archive/issue-7/article-14.
www.jbks.ru