
Разница между водорослями и растениями. Отличие водорослей от высших растений
Чем водоросли отличаются от растений
Водоросли и растения относятся к одному царству Растений. Благодаря водорослям, автотрофным пионерам, смогли появиться более специализированные и высокоорганизованные подданные царства Флоры.
Водоросли
Водоросли зародились на нашей планете 800 миллионов лет назад. Известно около 50-ти тысяч видов этих организмов. Изучением этих живых организмов занимается наука альгология. Их-то в первую очередь и удивляет эта группа растений своим видовым и морфологическим разнообразием – водоросли могут быть одноклеточными, колониальными, многоклеточными. Размер отдельных одноклеточных не позволяет рассмотреть их без микроскопа. Некоторые представители многоклеточных – например, бурая макроциста, вырастают в длину на несколько десятков метров. Но даже у этих гигантов тело не дифференцировано на органы. Кстати, «тело» водорослей именуется слоевищем или талломом.
На сегодняшний день описано 12 видов слоевища: амебоидный (хламидомонада), монадный (эвглена зеленая), коккоидный (вольвокс), капсальный, нитчатый (улотрикс, спирогира) и разнонитчатый, пластинчатый, сифоновый, сифонокладальный, харофитный (хара), сарциноидный и ложнотканевый.
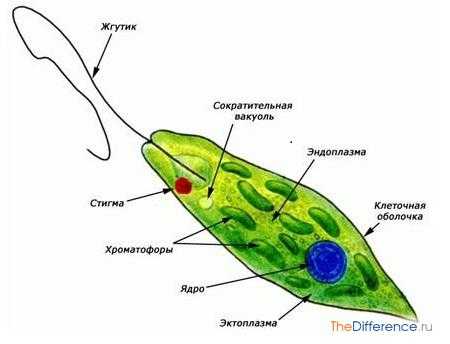 Эвглена зеленая. Строение
Эвглена зеленая. СтроениеДля поддержания существования и распространения своей группы на Земле водоросли размножаются 3 способами: бесполым с помощью зооспор или апланоспор, вегетативным – делением или выводковыми веточками и половым в виде изогамии, хологамии или коньюгации.
Общим признаком, позволившим исследователям отнести эти организмы к подцарству Низших растений, является наличие в клетках слоевища пластид с хлорофиллом. Поэтому растения способны к фотосинтезу. В клетках некоторых видов содержатся другие пигменты – красные, бурые, желто-зеленые, поэтому их талломы имеют характерные окраски. Водоросли не имеют органов, тканей. Питательные вещества и газы они поглощают и выделяют благодаря клеточным мембранам.
Большинство водорослей живут в пресных и соленых водоемах. Есть такие, которые плавают на поверхности, и те, что прикреплены ко дну, но лишь на глубине, куда доходит солнечный свет. Те организмы, которые находятся за пределами водной среды, могут поселиться в коре деревьев, на снегу, на шерсти животных, на скалах, в верхних слоях грунта. Водоросли вступают в симбиоз с коралловыми полипами и грибами, образуя лишайники.
Растения
Растения — это царство, объединившие эукариотические организмы со специфическим набором характеристик. Изучением этого таксона занимается наука ботаника.
Главной особенностью растений является наличие пигмента хлорофилла в пластидах, которые находятся в клетках. Благодаря ему и солнечному свету они могут самостоятельно синтезировать органические вещества, являются автотрофами и базой для пищевой пирамиды Земли.
Растения могут быть одноклеточными и многоклеточными. Тело высших растений состоит из тканей: основной, запасающей, механической, проводящей, покровной. У большинства растений тело состоит из органов – корень, стебель, цветок, листок, плод. Растения могут быть Низшими – водоросли и Высшими – мхи, папоротники, плауны, голосеменные и покрытосеменные. Последние представлены разными жизненными формами – травами, кустарниками и деревьями.
Выводы TheDifference.ru
- В количестве видов: водорослей меньше, чем растений.
- В уровне организации: водоросли самые примитивные представители царства Растений.
- Тело водорослей не дифференцировано на ткани и органы, в отличие от большинства растений.
- Большинство водорослей живет в водной среде, а большинство растений – на суше.
thedifference.ru
Чем водоросли отличаются от растений |
Водоросли и растения относятся к одному царству Растений. Благодаря водорослям, автотрофным пионерам, смогли появиться более специализированные и высокоорганизованные подданные царства Флоры.
Водоросли
Водоросли зародились на нашей планете 800 миллионов лет назад. Известно около 50-ти тысяч видов этих организмов. Изучением этих живых организмов занимается наука альгология. Их-то в первую очередь и удивляет эта группа растений своим видовым и морфологическим разнообразием – водоросли могут быть одноклеточными, колониальными, многоклеточными. Размер отдельных одноклеточных не позволяет рассмотреть их без микроскопа. Некоторые представители многоклеточных – например, бурая макроциста, вырастают в длину на несколько десятков метров. Но даже у этих гигантов тело не дифференцировано на органы. Кстати, «тело» водорослей именуется слоевищем или талломом.На сегодняшний день описано 12 видов слоевища: амебоидный (хламидомонада), монадный (эвглена зеленая), коккоидный (вольвокс), капсальный, нитчатый (улотрикс, спирогира) и разнонитчатый, пластинчатый, сифоновый, сифонокладальный, харофитный (хара), сарциноидный и ложнотканевый.Для поддержания существования и распространения своей группы на Земле водоросли размножаются 3 способами: бесполым с помощью зооспор или апланоспор, вегетативным – делением или выводковыми веточками и половым в виде изогамии, хологамии или коньюгации.Общим признаком, позволившим исследователям отнести эти организмы к подцарству Низших растений, является наличие в клетках слоевища пластид с хлорофиллом. Поэтому растения способны к фотосинтезу. В клетках некоторых видов содержатся другие пигменты – красные, бурые, желто-зеленые, поэтому их талломы имеют характерные окраски. Водоросли не имеют органов, тканей. Питательные вещества и газы они поглощают и выделяют благодаря клеточным мембранам.Большинство водорослей живут в пресных и соленых водоемах. Есть такие, которые плавают на поверхности, и те, что прикреплены ко дну, но лишь на глубине, куда доходит солнечный свет. Те организмы, которые находятся за пределами водной среды, могут поселиться в коре деревьев, на снегу, на шерсти животных, на скалах, в верхних слоях грунта. Водоросли вступают в симбиоз с коралловыми полипами и грибами, образуя лишайники.
Растения
Растения – это царство, объединившие эукариотические организмы со специфическим набором характеристик. Изучением этого таксона занимается наука ботаника.Главной особенностью растений является наличие пигмента хлорофилла в пластидах, которые находятся в клетках. Благодаря ему и солнечному свету они могут самостоятельно синтезировать органические вещества, являются автотрофами и базой для пищевой пирамиды Земли.Растения могут быть одноклеточными и многоклеточными. Тело высших растений состоит из тканей: основной, запасающей, механической, проводящей, покровной. У большинства растений тело состоит из органов – корень, стебель, цветок, листок, плод. Растения могут быть Низшими – водоросли и Высшими – мхи, папоротники, плауны, голосеменные и покрытосеменные. Последние представлены разными жизненными формами – травами, кустарниками и деревьями.
TheDifference.ru определил, что отличие водорослей от растений заключается в следующем:
В количестве видов: водорослей меньше, чем растений.В уровне организации: водоросли самые примитивные представители царства Растений.Тело водорослей не дифференцировано на ткани и органы, в отличие от большинства растений.Большинство водорослей живет в водной среде, а большинство растений – на суше.
altaiinter.org
Общая хар-ка высших или наземных растений. Важнейшие отличия высших растений от водорослей. Причины повышения общего уровня организации в процессе выхода растений на сушу
Первые достоверные наземные растения, известные только по спорам, датируются началом силурийского периода. Из верхнесилурийских и нижнедевонских отложений по сохранившимся макроостаткам или по отпечаткам органов описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов. Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид, и наличие многоклеточных спорангиев с кутинизированными спорами. Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше - в кембрии или в ордовике. Предпосылок для появления наземных растений было, по-видимому, несколько. Во-первых, независимый ход эволюции растительного мира подготовил появление новых, более совершенных форм. Во-вторых, за счет фотосинтеза морских водорослей в атмосфере земли произошло увеличение количества кислорода; к началу силурийского периода оно достигло такой концентрации, при которой оказалась возможной жизнь на суше. В-третьих, в начале палеозойской эры на обширных территориях земли происходили крупнейшие горообразовательные процессы, в результате которых возникли Скандинавские горы, горы Тянь-Шань, Саяны. Это вызвало обмеление многих морей и постепенное появление суши на месте бывших мелких водоемов. Если раньше водоросли, населявшие литоральную зону, только в отдельные кратковременные периоды жизни оказывались вне воды, то по мере обмеления морей они переходили к более длительному пребыванию на суше. Это, очевидно, сопровождалось массовой гибелью водорослей; выживали лишь те немногие растения, которые смогли противостоять новым условиям жизни. В ходе длительного эволюционного процесса возникали новые виды, постепенно формировавшие типичные наземные растения.
Прежде всего новая зона характеризовалась повышенной солнечной радиацией, дефицитом влаги и сложными контрастами двухфазной воздушно-наземной среды. Вполне вероятно допустить, что у некоторых переходных форм в процессе обмена веществ мог вырабатываться кутин, который откладывался, на поверхности растений. Это было первым этапом формирования эпидермы. Чрезмерное выделение кутина неминуемо приводило к гибели растений, так как сплошная пленка кутина препятствовала газообмену. Только те растения, у которых выделялось умеренное количество кутина, смогли сформировать сложную специализированную ткань - эпидерму с устьицами, способную и защищать растение от высыхания, и осуществлять газообмен. Таким образом, важнейшей тканью наземных растений, без которой невозможно освоение суши, следует считать эпидерму. Однако возникновение эпидермы лишало наземные растения возможности поглощать воду всей поверхностью, как это происходит у водорослей.
У самых первых наземных растений, имевших еще небольшие размеры, поглощение воды осуществлялось с помощью ризоидов - одноклеточных или многоклеточных однорядных нитей. Однако по мере увеличения размеров тела происходил процесс формирования сложных специализированных органов - корней с корневыми волосками. По-видимому, образование корней, начавшееся с верхнедевонского периода, в разных систематических группах растений происходило разными путями. Активное поглощение воды ризоидами и корнями стимулировало возникновение и совершенствование водопроводящей ткани - ксилемы. У нижнедевонских растений ксилема состояла только из кольчатых и спиральных трахеид. Начиная с верхнего девона прослеживается тенденция к "оживлению" ксилемы за счет развития древесной паренхимы, которая способствовала более активному проведению воды.
Выход растений на сушу сопровождался улучшением их освещения, что активизировало процесс фотосинтеза. Это приводило к увеличению количества ассимилятов, а следовательно, к увеличению объема растений, что вызвало необходимость их морфологического расчленения.
Существует несколько взглядов на исходную форму первых наземных растений. Одни авторы считают первичной формой пластинчатую - талломную, другие - напротив, радиальную. Есть третья точка зрения, согласно которой первые наземные растения выводятся из гетеротрихальных зеленых водорослей типа хетофоровых. Их стелющиеся части тела дали начало талломным формам, а восходящие - радиальным, т.е. талломные и радиальные структуры возникали одновременно и развивались параллельными путями. Пластинчатые талломы оказались малоперспективными в биологическом отношении, так как они очень быстро опутали бы всю поверхность земли тонким слоем, вызвав обостренную конкуренцию за свет. Восходящие структуры, напротив, получили дальнейшее развитие и сформировали радиальные ветвящиеся осевые органы. Вертикальное положение тела растения оказалось возможным только при условии возникновения механических тканей.
Независимо от формы тела во всех группах наземных растений очень рано (с середины девона) начала проявляться тенденция к образованию плоских боковых фотосинтезирующих органов - листьев. Подобно корням листья возникали разными путями, т.е. в разных систематических группах они имеют разное происхождение. (Своеобразие происхождения листьев нашло отражение в терминологии; так, все листья мохообразных иногда называют филлидами, листья плауновидных - микрофиллами, или филлоидами, папоротникообразных - макрофиллами, или вайями.Однако эти термины не всегда раскрывают специфику листьев разных групп растений.) Большая поверхность листьев в сочетании с наиболее совершенной, зернистой формой хлоропластов способствовала увеличению эффективности процессов фотосинтеза, т.е. накоплению органического вещества. Быстрое и равномерное распределение пластических веществ по всему телу растения оказалось возможным только при наличии совершенной проводящей ткани - флоэмы, которая обнаруживается уже у нижнедевонских риниофитов. Таким образом, у высших растений произошло формирование важнейших тканей - эпидермы, ксилемы, флоэмы, механической ткани и важнейших органов - листостебельного побега и корня. По-латыни побег - кормос (cormos), поэтому многие авторы называют высшие растения кормофитами. Однако А.Л. Тахтаджян справедливо отрицает универсальность этого названия, так как среди высших растений есть талломные мохообразные, а первые наземные растения - риниофиты еще не имели сформированных побегов.
Палеозойская эра характеризовалась интенсивными видообразовательными процессами, что привело к расцвету папоротникообразных, членистых, плауновидных. В мезозойскую эру господство в растительном мире переходит к голосеменным растениям, а в сменившую ее кайнозойскую эру расцвета достигают покрытосеменные растения. Быстрое освоение суши наземными растениями объясняется не только совершенством их вегетативных органов, но теми принципиальными изменениями, которые претерпели репродуктивные органы - гаметангии и спорангии. Основное назначение репродуктивных органов заключается в образовании и расселении потомства, обеспечивающего сохранение и процветание вида. Поэтому в условиях наземной среды обитания они должны быть более надежно защищены, чем одноклеточные гаметангии и спорангии водорослей. В ходе эволюции у всех высших растений сформировались многоклеточные спорангии и гаметангии, имеющие стенку из живых клеток. Цитоплазма клеток стенки, прочно удерживая влагу, предохраняет развивающиеся гаметы и споры от высыхания. Мужской половой орган - антеридий - имеет овальную или шаровидную форму (рис. 13). Под однослойной стенкой располагается сперматогенная ткань, из которой формируются жгутиковые сперматозоиды. Ко времени их созревания при наличии воды стенка вскрывается, и сперматозоиды по воде двигаются по направлению к архегонию. Женский половой орган - архегоний - имеет колбовидную форму; верхняя узкая часть называется шейкой, нижняя расширенная - брюшком. Внутри шейки под защитой стенки расположены шейковые канальцевые клетки, в брюшке - одна-две брюшные канальцевые клетки. На дне брюшка помещается одна крупная яйцеклетка. К моменту ее созревания шейковые и брюшные канальцевые клетки, а также верхние клетки стенки ослизняются, и часть слизи выходит за пределы архегония. Она содержит вещества, действующие хемотаксически на сперматозоиды, которые подплывают к архегонию, по слизи шейки двигаются по направлению к яйцеклетке и оплодотворяют ее.
Возможный путь возникновения многоклеточных архегониев и антеридиев высших растений рисует теория Дэвиса, созданная в 1903 г. Согласно этой теории, гаметангии высших растений произошли из многокамерных гаметангиев водорослей, подобных тем, которые имеются у эктокарпуса (Ectocarpus) из отдела бурых и у хетонемы (Chaetonema) из отдела зеленых. У них каждая клетка многокамерного гаметангия оказывается фертильной, т.е. способной образовывать изогаметы. Первые наземные растения, по представлению Дэвиса, обладали изогамным половым процессом. Увеличение количества сперматозоидов повышало вероятность полового процесса, а уменьшение их размеров способствовало передвижению сперматозоидов в самых тонких пленках воды. При формировании архегониев происходило увеличение размеров яйцеклеток и постепенное сокращение их числа до одного. Этот процесс биологически оправдан, так как одна крупная яйцеклетка, содержащая большой запас питательных веществ, способна обеспечить развитие наиболее полноценного потомства. Стерилизация внутренних клеток архегония и превращение их в брюшные и шейковые канальцевые клетки способствовало лучшему проведению сперматозоидов к яйцеклетке. Эта теория находит подтверждение в наличии так называемых смешанных гаметангиев, которые встречаются среди мохообразных. В одних случаях в гаметангиях могут развиваться одновременно яйцеклетки и сперматозоиды, в других случаях в архегонии развивается несколько яйцеклеток.
В результате полового процесса образуется диплоидная зигота. Она формирует диплоидный спорофит, который заканчивает свое развитие образованием многоклеточного спорангия со спорами. У всех высших растений, за исключением мохообразных, спорангии возникают на специальных органах, которые широко трактуются как спорангиофоры, т.е. носители спорангиев. Форма спорангиофоров в разных группах может быть разнообразной; в тех случаях, когда они имеют плоскую листовидную форму, их называют спорофиллами. Стенка спорангия может быть однослойной либо многослойной. Ее формирование в ходе эволюции могло происходить аналогично формированию стенки гаметангиев, т.е. путем стерилизации периферических спорогенных клеток.
В спорангиях всех высших растений в результате мейотического (редукционного) деления возникают тетрады гаплоидных спор. Характерной чертой всех высших растений является наличие в оболочке спор спорополлинина - вещества, близкого по физическим и химическим свойствам к кутину, поэтому обычно оболочки спор называют кутинизированными. Благодаря большой стойкости к химическим воздействиям и водонепроницаемости оболочек споры могут длительное время (иногда десятилетиями) сохранять свою жизнеспособность. Из гаплоидных спор формируется гаплоидное половое поколение - гаметофит. Мохообразные унаследовали гаметофитный тип цикла воспроизведения, при котором ведущее положение занимает гаметофит. Спорофит представлен лишь коробочкой и гаусторией (иногда развивается ножка), т.е. он не является самостоятельно живущим поколением. Поскольку половой процесс у мохообразных осуществляется сперматозоидами, т.е. при участии воды, гаметофит должен быть связан с влажными местообитаниями и не может по той же причине достигать больших размеров. Кроме того, гаплоидный гаметофит обладает меньшим генетическим потенциалом, чем диплоидный спорофит. Поэтому эта линия эволюции оказалась боковой, тупиковой. У всех остальных высших растений спорофит занимает ведущее место в цикле воспроизведения. Поскольку он обеспечивает размножение кутинизированными спорами, то является типично наземным растением.
Диплоидный набор хромосом расширил возможности формообразовательных процессов, поэтому огромное разнообразие жизненных форм высших растений принадлежит бесполому диплоидному поколению. Обильное ветвление и создание крупных размеров спорофита приводило к колоссальной продуктивности спор и к их эффективному рассеиванию.
Гаметофит, назначение которого состоит в осуществлении полового процесса, подвергся у большинства высших растений значительной редукции и существует в виде недифференцированного таллома, получившего название заростка, или проталлиума (prothallium). Широко распространено мнение, что предковые формы высших растений обладали изоморфным чередованием поколений, и от этого типа в дальнейшем эволюция пошла по двум направлениям - гаметофитному и спорофитному. Однако высказывается мысль и о том, что уже исходная группа водорослей обладала разными типами цикла воспроизведения и дала две независимые линии эволюции.
Итак, все высшие растения обладают рядом общих признаков: наличием функционально сходных тканей, морфологически сходных вегетативных органов, однотипных (за исключением покрытосеменных) многоклеточных половых органов и спорангиев, кутинизированных спор, правильным чередованием поколений. Это позволяет сделать вывод о единстве происхождения высших растений от какой-то одной группы водорослей. Длительное время в качестве исходной группы рассматривались бурые водоросли, так как они имеют расчлененный, часто весьма специализированный таллом. У некоторых представителей формируются ткани и встречаются многокамерные гаметангии. Однако различия пигментного состава и запасных питательных веществ вызывают серьезные возражения этим взглядам. Большинство современных ученых в качестве предковой группы рассматривают зеленые многоклеточные водоросли, обладавшими гетеротрихальным талломом. В этом убеждает сходство пигментного состава, запасных питательных веществ, наличие у некоторых современных хетофоровых многокамерных гаметангиев.
Водоросли – это прокариотические и эукариотические талломные споровые организмы, содержащие, как правило, в своих клетках хлорофилл, обладающие оксигенным фотосинтезом, являющиеся аэробами и не достигшие уровня дифференциации, характерной для архегониальных растений.
| Водоросли | Высшие растения |
| Возникли раньше (архейская эра, 4 млрд лет). | Возникли позже (силур палеозойской эры). |
| Первичноводные. | Наземные и вторичноводные. |
| Произошли в разное время от разных предков. Распределенысреди 4 царств (Сyanobiontes, Euglenobiontes, Chromista, Plantae). | Произошли от водорослевого предка и относятся к 1 царству Plantae. |
| Различные типы питания: автотрофное, гетеротрофное, миксотрофное (включая голозойное). | Преимущественно автотрофное питание, реже вторично гетеротрофное, либо авто-гетеротрофное. |
| Различный уровень организации (одноклеточный, колониальный, многоклеточный). | Только многоклеточный уровень организации. |
| Талломные (слоевищные) организмы. | Листостебельные, реже талломные, организмы. |
| Разнообразие морфоструктуры таллома (монадный, амебоидный, пальмеллоидный, коккоидный, нитчатый, разнонитчатый, паренхиматозный, псевдопаренхиматозный, харофитный, сифональный, сифонокладальный) (см. ниже). | У талломных только паренхиматозный (тканевой) таллом. |
| Ткани только у водорослей с паренхиматозным талломом (часть зеленых, бурых, красных, желтозеленых, золотистых). Из тканей только паренхима, реже (бурые) покровная, образовательная, проводящая (“ситовидные трубки”), механическая. | Ткани у всех. Различные ткани. |
| У большинства отсутствуют механические структуры. Есть только у бурых в сердцевине). Вода – плотнее воздуха. | Имеются механические ткани. |
| У подавляющего большинства нет проводящих тканей. | Есть проводящие ткани. |
| Нет устьиц, газообмен всей поверхностью тела. | Есть устьица. |
| У большинства нет покровных тканей. | Есть покровные ткани. |
| Всасывание минеральных веществ всей поверхностью тела. Ризоиды только для прикрепления. | Всасывание воды и минеральных веществ с помощью корневых волосков у большинства. |
| Покровы клеток разнообразны (цитоплазматическая мембрана, пелликула, перипласт, тека, панцирь, домик, клеточная оболочка). | Клетки имеют клеточную оболочку. |
| Клеточная стенка имеет разнообразный состав: целлюлоза, каллоза, муреин, пектин, углекислая известь, альгиновая кислота, фикоколлоиды. Лигнина нет. | Клеточная оболочка из целлюлозы, гемицеллюлозы, пектина. Могут откладываться лигнин, суберин. |
| Прокариоты и эукариоты. В клетке одно или несколько ядер. | Эукариоты. Ядро чаще одно. |
| Хроматофоры (хлоропласты) разнообразны по форме, числу, строению, набору пигментов, местоположению в клетке. Лейкопласты редко. У прокариотических водорослей нет хроматофоров. | Линзовидные хлоропласты. Есть хромопласты, лейкопласты. |
| Тилакоиды в хроматофорах по одному, в ламеллах и реже в гранах (зеленые, харовые). | Тилакоиды только в гранах. |
| Разнообразные хлорофиллы: a, b, c, d, e у водорослей разных отделов. | Только a и b хлорофиллы. |
| Каротиноиды (в том числе специфические – фукоксантин, сифонеин, сифоноксантин, вашериаксантин), фикобилины (фикоцианин, аллофикоцианин, фикоэритрин). | Каротиноиды. |
| Хроматическая адаптация в водоемах. | Нет. |
| У многих есть пиреноид. Пиреноид – это белковая структура, пространственно и функционально связанная с хроматофором, представляющая собой скопление фермента фиксации углекислоты при фотосинтезе. | Пиреноид есть только у некоторых мохообразных. |
| Разнообразные запасные вещества: крахмал, хризоламинарин (лейкозин), парамилон, ламинарин, багрянковый крахмал, сахариды, многоатомные спирты, цианофициновые гранулы, цианофициновый крахмал, белки, липиды (масла, жиры), волютин, гликоген. | Часто крахмал, масло, белки. |
| Кристы митохондрий разные: пластинчатые, трубчатые, дисковидные. | Кристы митохондрий только пластинчатые (ламеллярные). |
| Центриоли есть у немногих (хлорококковые зеленые, желтозеленые, бурые). | Центриолей нет. |
| Жгутики у подвижных стадий (зооспор, гамет) разные: изоморфные, изоконтные, гетероморфные, гетероконтные. | Жгутики есть только у сперматозоидов (только изоморфные и изоконтные). |
| Размножение разнообразное: – вегетативное (деление клетки, фрагментация колонии и многоклеточного таллома, столоны, выводковые почки, гормогонии, акинеты, гонидии, кокки, планококки, клубеньки), – собственно бесполое (подвижные и неподвижные споры), – половое (хологамия, гаметогамия, конъюгация, автогамия). Изогамия, гетерогамия, оогамия истинная или примитивная. | Выводковыми почками, частями вегетативных органов и д.р. Имеются только неподвижные споры. Половой процесс - только гаметогамия – истинная оогамия, сифоногамия. Семенное размножение. |
| Спорангии одноклеточные. | Спорангии многоклеточные. |
| Споры гаплоидные и диплоидные. | Споры только гаплоидные. |
| Все споры одинаковые по размеру. | Есть одинаковые споры, но есть и разные споры – микроспоры и мегаспоры. |
| Гаметангии у большинства одноклеточные. Только у харовых исключение. | Гаметангии только многоклеточные. |
| Для копуляции гамет нужна капельно-жидкая влага. | Для копуляции гамет капельно-жидкая влага нужна только споровым. У семенных мужские гаметы доставляют гаусториальная или пыльцевая трубки. |
| Разнообразные (все из возможных) жизненные циклы и типы мейоза. Есть циклы без полового процесса и мейоза. | Только один тип цикла и мейоза – гапло-диплобионтный со спорическим (промежуточным) мейозом. |
| В гапло-диплобионтном цикле есть 1) изоморфное чередование поколений, 2) гетероморфное с преобладанием гаметофита, 3) гетероморфное с преобладанием спорофита. | Только гетероморфное чередование поколений: 1) гетероморфное с преобладанием гаметофита (только у мохообразных), 3) гетероморфное с преобладанием спорофита (у всех других высших растений). |
1859 году канадский геолог Джеймс Досон в девонских отложениях на полуострове Гаспе в Канаде обнаружил остатки удивительно примитивного высшего растения не похожего ни на одно из известных науке в то время. Его вильчато разветвленные стебли были лишены листьев, а проводящая система представляла собой типичную протостелу, а это, как мы узнаем, был самый примитивный тип организации проводящей системы. Досон дал своему растению название Psilophyton princeps, что можно перевести на русский язык как «голорос первичный». Позднее в Шотландии были найдены и другие, еще более примитивные растения: риния, хорнеофит и куксония. Эти древние растения были выделены в особый отдел, для обозначения которого прежде употреблялось название «псилофиты» (Psilophyta), но в настоящее время многие ученые предпочитают название «риниофиты» (Rhiniophyta).
Остатки самых древних сухопутных растений были найдены в породах раннедевонского возраста, образовавшихся более 415 млн. лет назад. Породы состоят из окаменелых слоев торфа, который подвергался периодическим затоплениям. Вертикально стоящие растения заносились слоями песка прямо на месте произрастания. Благодаря тому, что в процессе захоронения растения эти пропитывались раствором кремнекислоты, сохранилась не только их внешняя форма, но и тончайшие детали внутреннего строения. Так что, мы имеем возможность воссоздать морфологическую эволюцию высших растений с большой достоверностью.
Надо отметить, что уже и до этого сушу населяли различные виды бактерий, водорослей и грибов. Но эти организмы в своей жизнедеятельности были полностью зависимы от наличия влаги в окружающей среде. Если наступала засуха ? они впадали в состояние анабиоза. Такие растения можно назвать пойкилогидрическими, по аналогии с пойкилотермными (холоднокровными) животными, неспособными регулировать температуру своего тела.
Высшие растения приобрели способность регулировать водный режим своего тела независимо от содержания влаги в почве и в воздухе. Поэтому их можно назвать гомойогидрическими.
Уже самые примитивные из известных нам высших растений были дифференцированы на элементарные органы, хотя и напоминали больше по внешнему виду своих водорослевых предков. Выход на сушу потребовал приспособления к совершенно новым условиям. Тело растения оказалось разделенным на две части: надземную и подземную.
Одни ответвления, называемые теломами, поднимались вертикально, другие прижимались к поверхности почвы, поглощая воду и минеральные соли. Последняя задача была не слишком трудной, учитывая то, что риниофиты обитали в заболоченных переувлажненных местах и на мелководьях.
Корневищеподобные веточки риниофитов получили название ? ризомоидов, они были покрыты волосовидными ризоидами и явились прототипом корней.
В ходе дальнейшей эволюции, по мере освоения более сухих почв, происходила более глубокая специализация подземных органов. Увеличивалась их поглощающая способность, благодаря обильному ветвлению и возникновению волосконосного слоя. Защиту апикальной меристемы в плотной почве обеспечило возникновение чехлика. Боковые ответвления корня стали закладываться изнутри (эндогенно) и не мешали продвижению в почве. Образование настоящих корней у высших растений было крупным эволюционным событием.
Одновременно с обособлением корней, надземная часть ? телома риниофитов, преобразовалась в побег, расчлененный на стебли и листья. В качестве листьев дифференцировались выросты на поверхности телома либо его ответвления. Образование листьев как аппаратов транспирации и фотосинтеза явилось важным приспособлением к надземному образу жизни. Это был новый прогрессивный признак общей структуры растительных организмов.
Морфогенез листьев шел двумя путями, приводившими к формированию или микрофиллов (малых листьев) или макрофиллов (крупных листьев) (микрофильная и макрофильная линии эволюции). Микрофиллы возникали как выросты (энанции) на поверхности оси телома. Макрофиллы ? это своеобразные плосковетки, видоизмененные и в результате специализации уплощенные ответвления телома (то есть возникли в результате уплощения боковых веточек и целых систем ветвления).
В процессе эволюции совершенствовалась и система ветвления телома. У самых примитивных высших растений телом ветвился вильчато (дихотомически), причем ветвление было равнодихотомическим. Путем неравномерного роста двух развилок, из которых одна растет сильнее (этот процесс был назван перевершиниванием) выработались более совершенные системы ветвления неравнодихотомическая, моноподиальная и симподиальная.
При моноподиальном ветвлении более развитая главная ось совершенно выпрямляется. Но главная особенность ветвления этого типа заключается в том, что боковые ветви закладываются под верхушкой главной оси.
При симподиальном ветвлении боковая ветвь перевершинивает главную, сдвигает ее в сторону и занимает ее место и так далее.
Открытие древнейших растений заставило пересмотреть принципы организации тела растения.
studopedia.net
В чём отличие низших растений от высших???
Высшие растения, зародышевые растения (Embryobionta, Embryophyta), побеговые растения (Cormophyta, Cormobionta), теломные растения (Telomophyta, Telomobionta), одно из двух подцарств растительного мира. Объединяет не менее 300 тыс. видов растений следующих групп (отделов) : риниовидные (псилофитовые) , моховидные (печёночники и мхи) , псилотовидные (псилотум и тмезиптерис) , плауновидные, хвощевидные, папоротники, голосеменные и цветковые, или покрытосеменные, растения. В отличие от низших растений, Высшие растения представляют собой сложно дифференцированные многоклеточные организмы, приспособленные к жизни в наземной среде (за исключением немногочисленных и явно вторичных водных форм) , с правильным чередованием двух поколений: полового (гаметофит) и бесполого (спорофит) . На спорофитах Высшие растения развиваются многоклеточные спорангии, в которых образуются неподвижные споры, на гаметофитах — многоклеточные половые органы (гаметангии) , однако у некоторых голосеменных (роды гнетум и вельвичия) и у всех цветковых растений гаметангии в процессе эволюции исчезли. Спорангии одинаковые или чаще дифференцированы на 2 типа (микроспорангии и мегаспорангии) , гаметангии — всегда двух типов — мужские (антеридии) и женские (архегонии) . Как антеридии, так и архегонии защищены многоклеточными стенками. Женская гамета (яйцеклетка) всегда одна, неподвижна. Зигота у Высшие растения развивается в многоклеточный зародыш, который проходит первые стадии развития внутри женского гаметофита. Спорофит у Высшие растения, как правило, расчленён на 3 основных органа — лист, стебель и корень. В большинстве случаев у Высшие растения развивается специальная проводящая ткань — ксилема и флоэма, у некоторых групп редуцировавшаяся. Имеется эпидерма с кутикулой и типичными устьицами. Хлорофилл у Высшие растения не содержит дополнительных пигментов, окраска фотосинтезирующих частей зелёная. Предками Высшие растения были какие-то морские водоросли. Достоверные ископаемые остатки Высшие растения известны начиная с силура.
В отличие от низших растений тело высших растений разделено на специализированные органы — листья, стебель и корень. Высшие растения — растения, которым свойственна дифференциация тканей, в отличие от низших растений — водорослей.
Низшие растения - это растения, которые низкие :) Высшие растения - это растения, которые высокие :) А на самом деле: Все растения: цветковые и нецветковые, имеющие корни, листья и стебли называются высшими растениями . Высшие растения чаще всего обитают на суше. Но есть среди них и те, которые обитают в воде (например, кувшинка, элодея) . Есть растения, у которых нет не только цветков и плодов но даже корней, стеблей и листьев. Это низшие растения. Они живут в соленой морской воде и в пресной воде рек и озер - это водоросли. Некоторые из них, например хламидомонада, хлорелла, настолько малы, что разглядеть их невооруженным глазом невозможно. Зато другие, такие как водоросль ламинария, вырастают до 60 м в длину. <img src="//otvet.imgsmail.ru/download/5eeae3a01fdf3ecc79d515e7b741db3a_i-7.jpg" >
Высшие растения — растения, которым свойственна дифференциация тканей, в отличие от низших растений — водорослей. К высшим растениям относятся мхи, и сосудистые растения (папоротникообразные, псилотовые, хвощевидные, плауновидные, голосеменные и покрытосеменные) . Эволюция высших растений тесно связано с выходом на сушу и завоеванием наземных ниш. Ни́зшие расте́ния, слоеви́щные, или талло́мные растения (лат. Thallophyta, греч. θάλλοφῠτόν: θάλλος — отпрыск, побег, молодая ветка, и φῠτόν — растение) — неформальный термин, объединяющий те растения, тело которых, в отличие от высших растений, не расчленено на части: (корень, стебель, лист) . Получается что высшие растения расчленены на части а низшие нет, в этом и есть отличие
llllllllllllllllllllllllllllllllll
Низшие растения - это растения, которые низкие :) Высшие растения - это растения, которые высокие :) А на самом деле: Все растения: цветковые и нецветковые, имеющие корни, листья и стебли называются высшими растениями . Высшие растения чаще всего обитают на суше. Но есть среди них и те, которые обитают в воде (например, кувшинка, элодея) . Есть растения, у которых нет не только цветков и плодов но даже корней, стеблей и листьев. Это низшие растения. Они живут в соленой морской воде и в пресной воде рек и озер - это водоросли. Некоторые из них, например хламидомонада, хлорелла, настолько малы, что разглядеть их невооруженным глазом невозможно. Зато другие, такие как водоросль ламинария, вырастают до 60 м в длину.
Высшие растения, зародышевые растения (Embryobionta, Embryophyta), побеговые растения (Cormophyta, Cormobionta), теломные растения (Telomophyta, Telomobionta), одно из двух подцарств растительного мира. Объединяет не менее 300 тыс. видов растений следующих групп (отделов) : риниовидные (псилофитовые) , моховидные (печёночники и мхи) , псилотовидные (псилотум и тмезиптерис) , плауновидные, хвощевидные, папоротники, голосеменные и цветковые, или покрытосеменные, растения. В отличие от низших растений, Высшие растения представляют собой сложно дифференцированные многоклеточные организмы, приспособленные к жизни в наземной среде (за исключением немногочисленных и явно вторичных водных форм) , с правильным чередованием двух поколений: полового (гаметофит) и бесполого (спорофит) . На спорофитах Высшие растения развиваются многоклеточные спорангии, в которых образуются неподвижные споры, на гаметофитах — многоклеточные половые органы (гаметангии) , однако у некоторых голосеменных (роды гнетум и вельвичия) и у всех цветковых растений гаметангии в процессе эволюции исчезли. Спорангии одинаковые или чаще дифференцированы на 2 типа (микроспорангии и мегаспорангии) , гаметангии — всегда двух типов — мужские (антеридии) и женские (архегонии) . Как антеридии, так и архегонии защищены многоклеточными стенками. Женская гамета (яйцеклетка) всегда одна, неподвижна. Зигота у Высшие растения развивается в многоклеточный зародыш, который проходит первые стадии развития внутри женского гаметофита. Спорофит у Высшие растения, как правило, расчленён на 3 основных органа — лист, стебель и корень. В большинстве случаев у Высшие растения развивается специальная проводящая ткань — ксилема и флоэма, у некоторых групп редуцировавшаяся. Имеется эпидерма с кутикулой и типичными устьицами. Хлорофилл у Высшие растения не содержит дополнительных пигментов, окраска фотосинтезирующих частей зелёная. Предками Высшие растения были какие-то морские водоросли. Достоверные ископаемые остатки Высшие растения известны начиная с силура.
touch.otvet.mail.ru






