Осмотическое давление. Осмотическое давление разных клеток и тканей растений
Биология для студентов - 07. Клетка как осмотическая система. Практическая значимость знаний по теме для земледелия
Диффузия воды через полупроницаемую мембрану называется осмосом.
Полупроницаемая мембрана – это мембрана, хорошо проницаемая для воды и непроницаемая или плохо проницаемая для растворенных в воде веществ.
Осмотическая ячейка – это пространство, окруженное полупроницаемой мембраной и заполненное каким-либо водным раствором, способным развивать определенное осмотическое давление.
Осмотическое давление (диффузное давление) – термодинамический параметр, характеризующий стремление раствора к понижению концентрации при соприкосновении с чистым растворителем вследствие встречной диффузии молекул растворённого вещества и растворителя. Если раствор отделен от чистого растворителя полупроницаемой мембраной, то возможна лишь односторонняя диффузия – осмотическое всасывание растворителя через мембрану в раствор. В этом случае осмотическое давление становится доступной для прямого измерения величиной. Оно равно избыточному давлению, приложенному со стороны раствора при осмотическом равновесии.
Осмотическое давление обусловлено понижением химического потенциала растворителя в присутствии растворённого вещества. Тенденция системы выравнивать химические потенциалы во всех частях своего объёма и переходить в состояние с более низким уровнем свободной энергии вызывает осмотический (диффузионный) перенос вещества.
Осмотическое давление в идеальных и предельно разбавленных растворах не зависит от природы растворителя и растворённых веществ; при постоянной температуре оно определяется только числом «кинетических элементов» (ионов, молекул, ассоциатов или коллоидных частиц) в единице объёма раствора.
Осмотическое давление (Р) численно равно давлению, которое оказало бы растворённое вещество, если бы оно при данной температуре находилось в состоянии идеального газа и занимало объём, равный объёму раствора.
Осмотическое давление измеряют с помощью специальных приборов (осмометров), определяя избыточное гидростатическое давление столба жидкости в трубке осмометра после установления осмотического равновесия.
Осмос является основным механизмом поступления воды в растительную клетку.
Все клеточные мембраны, в том числе плазмалемма и тонопласт, являются полупроницаемыми мембранами. Вода проходит в клетку через водные поры в плазмалемме, образованные специальными белками – аквапоринами.
Внутри вакуоли («осмотической ячейки») клеточный сок развивает осмотическое давление π:
π = i С R T,
где С – концентрация раствора в молях; Т – абсолютная температура; R – газовая постоянная 0,082 л × атм/град × моль; i – изотонический коэффициент, равный 1 + α(n–1), где α – степень электролитической диссоциации; n – число ионов, на которые распадается молекула электролита.
Благодаря осмотическому притоку воды в клетку там возникает гидростатическое давление, называемое тургорным. Это давление прижимает цитоплазму к клеточной стенке и растягивает ее. Клеточная стенка имеет ограниченную эластичность и оказывает равное противодавление. Эластическое растяжение ткани благодаря тургорному давлению ее клеток придает твердость неодревесневшим частям растений. Завядающие побеги становятся дряблыми, так как при потере воды тургорное давление падает. Тургорное давление противодействует притоку воды в клетку. Давление, с которым вода осмотически притекает в клетку, равно, таким образом, разности осмотического давления π и тургорного давления P. Эту величину называют сосущей силой S:
S = π – P.
Вода поступает в клетку из внешнего раствора, если его потенциальное осмотическое давление меньше сосущей силы клетки, и, наоборот, вода выходит из клетки в раствор с более высоким потенциальным осмотическим давлением. Величину осмотического потенциала можно определить плазмолитическим методом.
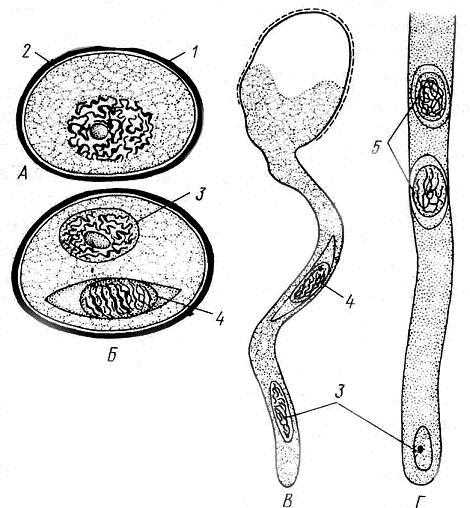
Плазмолиз – это процесс, обусловленный потерей воды клеткой. Он проявляется в отходе протопласта от клеточной стенки. При переносе плазмолизированных тканей в гипотонический раствор (или чистую воду) вода поступает в клетку и происходит деплазмолиз. Количество воды в клетке увеличивается, объем вакуоли возрастает – и она прижимает цитоплазму к клеточной стенке.
Плазмолитический метод основан на подборе изоосмотического (изотонического) раствора, то есть имеющего осмотический потенциал, равный осмотическому потенциалу клетки. Раствор, при котором начался плазмолиз, имеет осмотический потенциал, примерно равный осмотическому потенциалу клетки. Зная концентрацию наружного раствора в молях, можно вычислить осмотический потенциал клетки.
Иногда при сильном завядании протопласт не отстает от клеточной стенки, как при плазмолизе, а сжимается и тянет ее за собой. При этом клеточная стенка прогибается. Это явление называют циторризом. Развивается натяжение (или отрицательное давление стенки) – и потенциал тургорного давления приобретает отрицательное значение. В этом случае величина водного потенциала определяется уже не разностью, а суммой.
Величина осмотического потенциала позволяет судить о способности растения поглощать воду из почвы и удерживать ее, несмотря на иссушающее действие атмосферы.
Осмотический потенциал — показатель, равный по величине и размерности осмотическому давлению, но противоположный ему по знаку (т. е. осмотический потенциал является отрицательной величиной). Чем концентрироаннее раствор, тем выше его осмотическое давление и, следовательно, ниже его осмотический потенциал.
Осмотический потенциал колеблется у разных растений в пределах от –5 до –200 баров. У водных растений осмотический потенциал около –1 бара. У большинства растений средней полосы осмотический потенциал колеблется от –5 до –30 баров, растения степей и пустынь имеют более отрицательный осмотический потенциал. Осмотический потенциал различен и у разных жизненных форм. У деревьев он отрицательнее, чем у кустарников и травянистых растений. У светолюбивых растений осмотический потенциал отрицательнее, чем у теневыносливых растений.
Поступление воды в клетку обусловлено не только осмотическим давлением, но и силой набухания. Набуханием называют поглощение жидкости или пара высокомолекулярным веществом (набухающим телом), сопровождаемое увеличением объема. Явление набухания обусловлено коллоидальными и капиллярными эффектами. В протоплазме преобладает набухание на коллоидальной основе (гидратация коллоидов), а в клеточной стенке наблюдаются оба эффекта: капиллярный – накопление воды между микрофибриллами и в межмицеллярных пространствах; коллоидальный – гидратация полисахаридов, особенно гемицеллюлоз. У некоторых частей растений поглощение воды происходит исключительно путем набухания, например, у семян.
Новые прогрессивные направления биотехнологии и биоинженерии растений призваны обеспечивать высокую продуктивность и устойчивость растений. Фактором, действующим на растения, является импульсное давление (ИД). Давление влияет на различные процессы у растений. Быстрые изменения осмотического давления могут вызывать некоторые неспецифические стрессовые симптомы в растениях, в то время как нормальное осмотическое давление - важный фактор в клеточном развитии, росте и поглощении воды. Перед посевом семена обрабатывали импульсным давлением, создаваемым ударной волной в водной среде. Каждый плод испытывал объёмное сжатие. Время прохождения ударной волны составляло 15–25 мксек. Импульсное давление (ИД) при воздействии на семена в малых дозах (11 МПа) приводит к увеличению продуктивности на 12 % без значительных повреждений и способствует адаптации растения ко второму фактору — пониженной температуре.
Изученные А.В. Шиленковым факторы повышают содержание пигментов в листьях гречихи среднего яруса, интенсивность фотосинтеза растений, что создает предпосылки для изменения структуры урожая, а также морфофизиологических характеристик. Импульсное давление и низкие температуры увеличивают засухо – и солеустойчивость семян и проростков гречихи.
Осмотическое давление — индикатор водного режима растений
Оно подвержено многим изменениям в результате колебаний содержания воды и осмотически активных веществ (коллоидно-химических изменений) в цитоплазме. Годовой ход осмотического давления у разных видов, особенно гидролабильных, четко отражает меняющиеся условия увлажнения. Осмотическое давление растет по мере увеличения сухости. При оптимальной влажности почвы и воздуха гидратура поддерживается беспрепятственно. Однако затруднение в водоснабжении обычно ведет к пассивному повышению осмотического давления, затем к изменению структуры цитоплазмы, активному поднятию осмотического давления, повышению засухоустойчивости на клеточном уровне и далее к усилению ксероморфной структуры новых органов.
Минимальное, оптимальное и максимальное значения осмотического давления — кардинальные точки гидратуры, свойственные отдельным видам растений и являющиеся их экологической характеристикой. Для сравнения осмотических свойств видов и сообществ разных местообитаний Г. Вальтер (1960) предложил составлять осмотические спектры (рис. 8.3). Измерение диапазонов осмотического давления клеточного сока листьев показало низкие значения осмотического давления у кустарников и эпифитов влажных тропических лесов (от 3 до 11 бар). Высоких значений достигает осмотическое давление у галофитов. У растений пустыни Каракумы оно в пределах 13,6—58,9 бар. В период засухи уровень максимального осмотического давления повышается. Наибольшая его амплитуда отмечена у пустынных эфемероидов.
Осмотическое давление закономерно меняется при онтогенетическом и сезонном развитии растений. В благоприятных условиях молодые листья имеют, как правило, более низкие его значения, что оптимально для ранних стадий развития. По мере развития листа осмотическое давление постепенно повышается и достигает значений, оптимальных для вида. Способность поднимать осмотическое давление к зиме при накоплении в клетках сахаров способствует продвижению растений к полюсам.
У разных видов максимальное значение осмотического давления неодинаково. В пределах вида у закаленных и засухоустойчивых экземпляров его значение выше. При недостатке воды и голодании в случае снижения или прекращения фотосинтеза осмотическое давление может опуститься ниже оптимального. Это вызывает сильную перестройку цитоплазмы, потребление растворенных запасных веществ и еще большее снижение осмотического давления. При сильном его снижении растение может погиб- путь.
Формообразующее значение гидратуры. Между гидратурой рас- гспий, осмотическим давлением клеток и морфологической структурой их органов существует тесная связь. Морфология листа, анатомическое строение и физиологические особенности растений, выросших в оптимальных условиях (высокая гидратура), отличаются от растений, находящихся в условиях затрудненного водоснабжения (низкая гидратура). При затрудненном водоснабжении у растений возникают признаки ксероморфизма:

1. Мелкоклеточность, уменьшение размеров органов (как стебля, так и площади листовой пластинки) и всего организма (нанизм).
2. Устьица мелкие, но на единицу площади листа их много (обеспечение высокой интенсивности транспирации на единицу поверхности листа при благоприятном водоснабжении).
3. Утолщение листовой пластинки, при этом сильное развитие столбчатой ткани, что связано с интенсивным фотосинтезом в благоприятных условиях; губчатый мезофилл развит слабее.
4. Клетки ксилемы мелкие, в них много механических волокон, общая площадь сосудисто-волокнистых пучков относительно велика. Сеть жилок густая, промежутков между ними немного.
5. Сильное развитие опушения, кутикулы, утолщение клеточных стенок, что в засуху при закрытых устьицах исключает кути- кулярную транспирацию.
6. Развитая корневая система, превосходящая по массе надземную часть.
eco-rasteniya.ru
Осмотическое давление — Википедия
Материал из Википедии — свободной энциклопедии
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 1 января 2016; проверки требуют 16 правок. Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 1 января 2016; проверки требуют 16 правок.Осмотическое давление (обозначается π) — избыточное гидростатическое давление на раствор, отделённый от чистого растворителя полупроницаемой мембраной, при котором прекращается диффузия растворителя через мембрану (осмос). Это давление стремится уравнять концентрации обоих растворов вследствие встречной диффузии молекул растворённого вещества и растворителя.
Мера градиента осмотического давления, то есть различия водного потенциала двух растворов, разделённых полупроницаемой мембраной, называется тоничностью. Раствор, имеющий более высокое осмотическое давление по сравнению с другим раствором, называется гипертоническим, имеющий более низкое — гипотоническим.
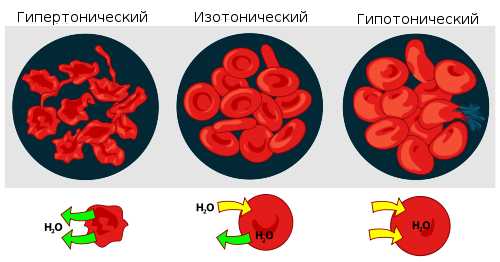 Взаимодействие эритроцитов с растворами в зависимости от их осмотического давления.
Взаимодействие эритроцитов с растворами в зависимости от их осмотического давления. Если же подобный раствор находится в замкнутом пространстве, например, в клетке крови, то осмотическое давление может привести к разрыву клеточной мембраны. Именно по этой причине лекарства, предназначенные для введения в кровь, растворяют в изотоническом растворе, содержащем столько хлорида натрия (поваренной соли), сколько нужно, чтобы уравновесить создаваемое клеточной жидкостью осмотическое давление. Если бы вводимые лекарственные препараты были изготовлены на воде или очень сильно разбавленном (гипотоническом по отношению к цитоплазме) растворе, осмотическое давление, заставляя воду проникать в клетки крови, приводило бы к их разрыву. Если же ввести в кровь слишком концентрированный раствор хлорида натрия (3—10 %, г
ru.wikipedia.org
Осмотическое давление живых клеток - Справочник химика 21
С помощью точного и устойчивого количества электролитов сохраняется равное осмотическое давление между клеткой и окружающей ее жидкостью и, таким образом, одинаковое количество воды в тканях. Такое же равновесие достигнуто между кровью и межклеточной жидкостью, окружающей кровеносные капилляры. До тех пор, пока будет сохраняться равновесие осмотического давления различных жидкостей, содержание воды в живых тканях будет оставаться неизменным. Это жидкостное напряжение (тургор или тонус) обеспечивает коже вместе с волокнами соединительной ткани упругость, или эластичность. [c.197]
Осмос имеет большое значение в процессах жизнедеятельности животных и растений. Он обусловливает поднятие воды по стеблю растения, рост клетки и многие другие явления. Осмотическое давление в клетках обусловливает их своеобразную упругость и эластичность, а также способствует сохранению определенной формы стеблями и листьями растений. Каждая живая клетка имеет либо оболочку, либо поверхностный слой протоплазмы, обладающие свойством полупро-ницаемости. Если клетку поместить в раствор, концентрация которого равна концентрации клеточного сока, то состояние клетки не изменится, так как осмотическое давление в клетке и в растворе одинаково. [c.123]
Осмос и осмотическое давление играют огромную роль в процессах жизнедеятельности организмов. Например, с ним связано распределение воды в организме. Каждая живая клетка — микроскопическая осмотическая система. [c.82]
Состав внеклеточной жидкости близок к составу морской воды в пред-кембрийскую эпоху, когда появились животные с замкнутой системой кровообращения. С тех пор соленость моря продолжала возрастать, тогда как состав внеклеточной жидкости остался постоянным. Основным катионом во внеклеточной жидкости является ион Ка , а из анионов преобладают СГ и НСОВнутри клеток преобладают катион и анион НРО Для соблюдения физико-химического закона электронейтральности, которому подчиняется любой живой организм в целом, некоторый недостаток неорганических анионов компенсируется анионами органических кислот (молочной, лимонной и др.) и кислых белков, несущих отрицательный заряд при физиологических значениях pH. Если вне клетки органические анионы компенсируют незначительную нехватку отрицательного заряда, то внутри клетки они должны компенсировать около 25 % положительных зарядов, создаваемых неорганическими катионами. Поскольку клеточные мембраны легко проницаемы для воды, то они могут разрушаться при незначительных различиях в давлении жидкости внутри и снаружи клеточной мембраны. Поэтому осмотическое давление внутри клетки должно быть равно таковому во внеклеточной жидкости, т. е. живая клетка подчиняется закону изоосмоляльности. Повышенное содержание катионов по отношению к концентрации анионов во внеклеточных жидкостях в сравнении с внутриклеточными средами приводит к тому, что наружная поверхность мембран клеток оказывается заряжена положительно относительно ее внутренней поверхности, и это имеет огромное биологическое значение (см. главу 15). В биологических жидкостях концентрацию осмотически активных частиц (независимо от их заряда, размера и массы) выражают в единицах осмоляльности — миллиосмомолях на 1 кг воды. Так как главные катионы и анионы внутриклеточных жидкостей многозарядные, то (при одинаковых осмоляльностях) концентрация электролитов, выраженная в миллиэквивалентах на 1 л, будет значительно выше внутри клетки, чем во внеклеточных жидкостях, где в основном содержатся однозарядные ионы. [c.180]
Для жизнедеятельности микроорганизмов необходимо наличие в питательной среде минеральных веществ. С их помощью регулируется осмотическое давление внутри клетки, они определяют скорость и направление биохимических реакций и коллоидные свойства живой протоплазмы. Из минеральных элементов важнейшую роль в питании микробов играет фосфор, необходимый для синтеза сложных фосфорсодержащих соединений с белками. Лучшими источниками фосфора являются различные фосфорнокислые соли калия, натрия, аммония. [c.514]
Обычно биологи, говоря о соединениях натрия, калия, магния и кальция, в первую очередь имеют в виду их хлориды, фосфаты, сульфаты и карбонаты. Все они имеют довольно сильно ионизированные связи. Соединяясь с катионами водорода, образуют кислые соли и кислоты. Соли имеют большое значение не только как составные части жизненной среды, но и как активные участники процессов в живых клетках. Ионы натрия благодаря небольшому размеру играют важную роль в поддержании водного режима организма, и увеличение концентрации Ма+ способствует удерживанию воды. Соли натрия наряду с солями других металлов определяют осмотическое давление в клетках и влияют на работу ферментных систем. Ионы натрия вместе с ионами калия служат для передачи нервного импульса через мембраны нервных клеток (см. 38). Нормальный ритм работы сердца и головного мозга зависит от строгого выдерживания соотношения концентраций ионоз калия и натрия. Ионов калия в организмах животных меньше, и повышение концентрации К+ оказывает вредное действие. В растениях калий способствует фотосинтезу и стимулирует процессы, связанные с прорастанием семян. Поэтому так важны калийные удобрения. Роль магния не ограничивается только участием в структуре хлорофилла. В организмах животных и человека он уменьшает спазмы сосудов и регулирует работу сердца. В периодической системе М занимает промежуточное положение между бериллием и щелочноземельными металлами. Его свойства уникальны имея высокий заряд и небольшой радиус, он в то же время образует в большинстве своих соединений не ионные, а кова- [c.180]
Такое явление происходит в живой растительной и животной ткани. Здесь создается разность осмотических давлений в клетке и межклеточном пространстве эта разность играет решающую роль в прижизненном обмене веществ. В мертвой ткани не происходит обмена веществ, следовательно, не может быть различных концентраций водного раствора неорганических и органических веществ в клетке и межклеточном пространстве, т. е. различных температур замерзания. [c.113]
Осмотические явления, наблюдаемые в организмах, обусловливаются теми же причинами, какие действуют в рассмотренных нами осмотических моделях. Осмотическое давление в клетках подчиняется всем основным газовым законам. Однако в явления осмоса вносят значительные коррективы сама структура живых образований и процессы жизнедеятельности, непрерывно протекающие в живых организмах. [c.131]
Осмотическое давление в клетках подчиняется газовым законам. При этом следует учитывать, что на осмос значительное влияние оказывает структура живых образований и процессы жизнедеятельности в организмах. В организм постоянно должны поступать питательные вещества. Если бы оболочка клеток была идеальной мембраной, пропускающей только молекулы растюр ителя, но препятствующей проникновению молекул растворенного вещества, то процессы питания были бы невозможны, что естественно привело бы к гибели организма. [c.97]
Абсолютная величина осмотического давления в клетках живых растений достаточно высокая и колеблется чаще всего от 506 до 1010 кПа у наземных растений и от 101 до 304 кПа у водных. В плодах фруктовых деревьев, ягодах винограда, корнеплодах сахарной свеклы осмотическое давление бывает в пределах 2026—4052 кПа, у растений, которые выращивают на засоленных почвах, оно может достигать 6078—10 130 кПа, Величина сосущей силы в разных органах и тканях имеет важное. значение для поступления и движения воды по растению,, [c.115]
Красные кровяные тельца—это живые клетки, ограничивающая поверхность которых обладает свойствами полупроницаемых перегородок. Эта поверхность отличается от остальной клетки богатством липоидов и весьма чувствительна к разного рода влияниям. Так, щелочи и растворители жиров разрушают ее. При набухании, обусловленном уменьшением осмотического давления в внешней среде, т. е. в плазме [c.192]
Ферментативные синтезы полисахаридов класса крахмала. Глюкоза является неподходящим материалом для образования запасов углеводов в живых клетках, так как она легко растворима в воде, причем ее растворы обладают высоким осмотическим давлением. Отсюда вытекает необходимость отложения глюкозы в полимерной форме в печени или мышцах. В настоящее время механизм этого важного физиологического процесса в большой степени выяснен. [c.320]
Для растения осмотическое давление играет огромную роль. Оно поддерживает устойчивость тканей, создавая в клетках тургор, т. е. набухание. При отмирании протоплазмы клеток последние теряют свои полупроницаемые свойства и свободно пропускают в обе стороны не только воду, но и раствор тургор прекращается и растение теряет свою эластичность (вянет). Гистологам хорошо известно, что только после того как клетка убита (фиксирована), удается окрасить ее содержимое погружением ее в раствор красок. Живая клетка, поглощая воду, задерживает большинство растворенных в ней красок. То же осмотическое давление обусловливает движение жидкостей по сосудам растения от корней и т, д. [c.237]
Скорость поглощения воды живыми клетками при осмотических изменениях неодинакова для различных типов клеток, но вообще она чрезвычайно велика окончание процесса часто требует всего нескольких минут. Осмотическое давление внешней среды можно контролировать прибавлением электролита или разбавлением, причем осмотическая реакция клетки находится в согласии с уравнением (10). Это показывает, что скорость проникновения воды через клеточные мембраны значительно превышает скорость проникновения электролитов. [c.372]
Осмотическое давление играет большую роль в процессах жизнедеятельности животных и растений. Процессы усвоения пищи, обмена веществ и т. д. тесно связаны с различной проницаемостью тканей для воды и тех или иных растворенных веществ. Типичные клетки образованы из протоплазменных мешочков, наполненных водными растворами разных веществ (клеточный сок), осмотическое давление которых на границе с водой обыкновенно лежит в пределах 4—20 атм. Если эти клетки погружены в воду или в раствор меньшей концентрации, чем их содержимое, то вода проникает в клетки, создавая в них гидростатическое давление, называемое тургором. Это давление создает прочесть и устойчивость живых тканей растений и животных. Осмотическое давление играет роль механизма, подающего клеткам питательные растворы и воду в высоких деревьях они поднимаются на десятки метров вверх от корней, что отвечает осмотическому давлению в несколько десятков атмосфер. [c.110]
Все живые клетки вообще проницаемы для воды без какого-либо особого активного механизма вода переносится в клетку и из нее в соответствии с различием в осмотическом давлении. [c.372]
Живые клетки не только способны набухать или сокращаться с увеличением или уменьшением осмотического давления внешней среды, но это изменение имеет количественный характер [c.372]
Вант-Гофф начал свой рассказ. От полупроницаемых перегородок он перешел к живым клеткам, к огромной силе, которая возникает в них в результате осмотического давления. [c.60]
Осмотическое давление представляет собой величину, тесно связанную с нормальной жизнедеятельностью клетки. В этом легко убедиться, сопоставив концентрации сока, выжатого из живых клеток и из тех же клеток, предварительно убитых. [c.73]
Наружный слой клетки, т. е. ее мембрана, пропускает не только воду, о и в какой-то мере растворенные в ней вещества. Живая клетка активно регулирует осмотическое давление, изменяя концентрации осмотически активных веществ. У одноклеточных животных, обитающих в пресной воде, выработались специальные приспособления (пульсирующие вакуоли), удаляющие из клеток избыточную воду. Одноклеточные организмы, не имеющие пульсирующих вакуолей, излишнюю воду удаляют через клеточную мембрану. У высших животных осмотическое давление е целом организме регулируется системой органов выделения (почками). [c.55]
Из последнего уравнения видно, что уменьшение свободной энергии при постоянной температуре и постоянном давлении равно общей работе, которую может совершить система, за вычетом ЯАУ. Работа, обозначаемая —АО, которую часто называют чистой работой или полезной работой, может быть химической, электрической, фотохимической или осмотической. К ней не относится работа расширения газа против постоянного дав- ления, за исключением случаев, когда этот вид работы используется полезным образом. Примером такой непродуктивной работы РАУ может служить расширение нагреваемого воздуха. В живых системах вопрос о полезной работе, сопровождающейся изменением свободной энергии, имеет особо важное значение, поскольку растительные и животные клетки совершают механическую работу, необходимую для поддержания жизни, не. за счет поглощения тепла из внешней среды. В случае теплокровных животных совершение работы при постоянной температуре за счет поглощения тепла просто запреш,ено вторым законом. Работа клеток зависит от химической энергии, получаемой ими из питательных веществ этот вид работы может осуществляться, очевидно, за счет изменения свободной энергии АС. Рис. 2.11 иллюстрирует некоторые виды полезной работы, с которыми мы сталкиваемся, изучая живые организмы. [c.107]
Высшие растения состоят из огромного числа клеток, определенным образом скрепленных друг с другом окружающими их клеточными стенками. Многие характерные свойства растений прямо или косвенно связаны с наличием этих клеточных стенок. Состав и внешний вид клеточных стенок непосредственно определяются тем, к какому типу принадлежит данная клетка и каковы ее функции. Вместе с тем основные принципы построения всех клеточных стенок поразительно сходны жесткие волокна целлюлозы погружены в матрикс, содержащий множество поперечных сшивок и состоящий из таких полисахаридов, как пектины и гемицеллюлозы, а также из гликопротеинов. Благодаря такому строению первичная клеточная стенка обладает большим запасом прочности при растяжении и способна пропускать лишь молекулы относительно небольшого размера. Если растительную клетку, лишенную клеточной стенки (протопласт), поместить в воду, то она осмотическим путем наберет воду, набухнет и лопнет. В то же время живое содержимое клетки, заключенное в оболочку, набухает и давит на последнюю, в результате чего возникает давление, известное под названием тургорного. Тургор строго регулируется и жизненно необходим как для увеличения размеров клетки, так и для механической жесткости молодого растения. [c.398]
До сих пор мы предполагали, что коллоид не является электролитом, а это действительно верно для растворов макромолекул в неполярных растворителях. Однако в водных растворах многие макромолекулы, и прежде всего различные биоколлоиды, как правило, находятся в виде ионов. Если же раствор, кроме того, содержит обычные электролиты, то картина еще более усложняется. Здесь осмотическое равновесие сочетается с электростатическими взаимодействиями. Макроионы, которые не проходят через поры мембраны, частично удерживают около себя противоионы и нарушают их равномерное распределение возникает так называемый мембранный потенциал (играющий важную роль в процессах обмена живой клетки). Электростатически обусловленная повышенная концентрация ионов с одной стороны мембраны является причиной более высокого осмотического давления. Добавка электролита экранирует мембранный потенциал (эффект сжатия противоионной атмосферы), а тепловое движение понижает неравномерное распределение ионов, и осмотическое давление понижается. Предельный случай полностью подавленного мембранного потенциала (равномерное распределение всех ионов около мембраны) соответствует осмотическому давлению раствора неэлектролита той же концентрации. Теорию этого эффекта предложил Доннан (1911г.). Допустим, что слева от мембраны находится раствор полиэлектролита N31 с концентрацией с , а справа — раствор обычного электролита, например ЫаС1, с концентрацией с . Мембрана свободно пропускает молекулы растворителя (воды), ионы Ыа+ и С1 , но не пропускает ионы Для простоты вслед за Доннаном примем, что объемы растворов, находящихся с обеих сторон мембраны, одинаковы. Это делает вывод наглядным, не лишая его общности. Предположим также, что оба электролита полностью диссоциированы. Когда в системе установится равновесие, в ту часть раствора, где находится ЫаК, перейдет х молей ЫаС1, так что концентрация N3+ в нем повысится до - + х, концентрация К останется, как и прежде, равной с , а концентрация С1 , которая вначале была равна нулю, составит х. По другую сторону мембраны концентра- [c.45]
Эффект Доннана (т. е. нерав1юмерное распределение электролитов между клетками и омывающей их жидкостью) оказывает большое влияние на жизнедеятельность клеток, на величину биопотенциалов и т. п. Однако в целостном организме на распределение ионов влияет ряд физиологических регуляторных механизмов, процессы обмена веществ и т. п. Поэтому в живом организме эффект Доннана является лишь одной из причин сложных процессов возникновения осмотического давления, электрических явлений, распределения электролитов и пр. [c.196]
Оболочка живых клеток - всегда полупроницаемая мембрана. Она задерживает молекулы многих веществ, растворенных в воде, но воду пропускает. Поэтому каждая животная и растительная клетка - это микроскопическая осмотическая система, а осмотическое давление ифает очень важную роль в жизнедеятельности организмов. [c.68]
Осмотическое давление имеет важное значение в биологических процессах. К стенкам клеток прилегает, как известно, слой протоплазмы, которая играет роль полупроницаемой перегородки. Заключенная внутри клеток жидкость (клеточный сок) содержит в растворенном состоянии различные вещества, которые не проникают через слой протоплазмы. Если клетка находится в воде, то последняя проникает внутрь клетки и создает в ней давление, называемое тургором. Благодаря тургору ткани живых растений приобретают свойственную им прочность и упругость. При отмирании клеток протоплазма теряет свойство полупрони-цаемости, что приводит к исчезновению тургора и к увяданию растения. Когда живая клетка попадает в раствор, концентрация которого больше концентрации клеточного сока, вода перемещается из клетки в наружный раствор, протоплазма сжимается, наступает явление плазмолиза. [c.27]
Ионы щелочных металлов (Na , К )- Натрий распределен в основном снаружи, а калий - внутри клетки. Оба катиона вносят вклад в поддержание осмотического давления, передачу нервных импульсов, активный перенос сахаридов и аминокислот. Катионы Na" " и К , представляющие собой сильные кислоты, образуют комплексы с лигандами, содержащими донорные атомы кислорода (эти лиганды являются сильными основаниями). Но в живых системах эти ионы переносятся свободно, поскольку in vivo взаимодействие ионов с лигандами сравнительно слабое. Роль антибиотиков-ионофоров в активном переносе ионов через клеточные мембраны, например, в избирательном переносе натрия и калия при возбуждении мембран нервных клеток или [c.269]
Осмотическое давление играет большую роль в жизнедеятельности человека, животных и растений. Клетки живого организма состоят из протоплазменных мешочков, наполненных водными растворами различных веществ (клеточным соком). Осмотическое давление клеточного сока на границе с водой лёжит в пределах 4-10 —20-10 н м . Если клетка погружена в воду или раствор меньшей концентрации, чем концентрация клеточного сока, то вода проникает в клетку, создавая в ней гидростатическое давление, называемое тургором. Если клетка погружена в раствор, более концентрированный, чем клеточный [c.142]
Первичным восстановителем при обычном фотосинтезе в зеленых растениях является вода. Активность воды в клетках может быть изменена посредством прямого оводнения и обезвоживания или путем помещения клеток в растворы с различным осмотическим давлением. Обе эти операции оказывают значительное влияние на фотосинтез. Однако это влияние не может рассматриваться как кинетическое явление, подчиняющееся закону действующих масс, так как оно связано с изменениями проницаемости и других коллоидных свойств протоплазмы и клеточных мембран, от которых зависит в различной степени всякая активность живой клетки. Действие обезвоживания обсуждалось поэтому в гл. XIII (т. I, стр. 341), где мы имели дело с различного рода физическим и химическим ингибированием и стимулированием фотосинтеза. [c.365]
Дрожжевую- клетку, помещенную в раство1р различных веществ, каким является осахаренная масса, условно можно рассматривать как ячейку, окруженную полупроницаемой перегородкой. В отношении такой ячейки будут до известной степени справедливы законы осмотического давления с соответствующими поправками на коллоиды внутри клетки и на живую плазму. [c.270]
Явление осмоса в живых клетках. Диффузия, осмос и осмотическое давление играют важную роль в животных и растительных организмах. Протоплазма клеток представляет собой идеальную полупроницаемую перегородку, через которую в клетку могут проникать или удаляются из нее только определенные вещества, но она непроницаема для других веществ. Так, стенки эритроцитов (красных кровяных телец) непроницаемы для хлорида натрия, но проницаемы для воды. Если эритроциты ввести в раствор хлорида натрия, осмотическое давление которого больше, чем осмотическое давление внутри клеток гипертонический раствор), то вода диффундирует из клеток наружу, и клетки сжимаются. В растворе, осмотическое давление которого меньше внутреннего давления гипотонический раствор), осмос происходит в противоположном направлении. Вода извне проникает в клетки, при этом они набухают. Объем клеток увеличивается до тех пор, пока их стенки не лопаются и содержимое не вытекает в окружающую жидкость. Два раствора, которые имеют одинаковое осмотическое давление независимо от состава растворенных веществ, обусловливающих это давление, называются изотоническими или изоосмоти-ческими растворами. Жидкость крови, в которой взвешены красные кровяные тельца,— плазма крови — является изотонической с жидкостью, находящейся в красных кровяных тельцах и других клетках организма. [c.161]
Эти явления могут быть использованы и для определения осмотического давления в живых клетках, особенно в клетках растений. Внешняя проницаемая оболочка из клетчатки покрыта протоплазмой, которая играет роль полупроницаемой перегородки. Если такую клетку ввести в гипертонический раствор, то вода выходит изнутри через протоплазму. Протоплазменная оболочка морщится, отрываясь от оболочки из клетчатки,— происходит плазмолиз. Более разбавленный раствор, в котором плазмолиз прекращается, является изотоническим по отношению к жидкости, находящейся внутри клетки. При введении в гипотонические растворы клетки набухают за счет проникающей извне через протоплазменную оболочку воды. Поскольку в обычных условиях вода, поглощаемая растениями, содержит мало растворенных веществ, она проникает через оболочку и создает внутри клетки давление, которое действует на стенки клетки (тургесцентность). Это давление определяет упругость растений, особенно молодых, зеленых. Когда клетки при испарении теряют больше воды, чем проникает снаружи за счет осмоса, они теряют упругость, растение как бы увядает. [c.162]
Вода представляет o6oii главный жидкий компонент животных и растительных тканей, создает среду для переноса питательных веществ к растущим клеткам и удаления вредных вешсств, а также обеспечивает регулирование тепла за счет испарения. Поскольку последние две функции связаны с выводом воды из организма, необходимо периодическое восстановление водного баланса. Вода поглощается через корневую систему растений и через мембраны клеток живых существ. Поглощение является следствием различия осмотических давлений по обе стороны мембраны и может происходить только при градиенте концентраций растворенных солей (в воде меньше, чем во внутриклеточной жидкости). [c.532]
В физиологических условиях, т. е. при тех условиях, которые имеются в живой клетке (температура, pH, осмотическое давление, концентрация реагируюшлх веществ и пр.), расщепление моля АТФ (506 г) сопровождается выделением 12 ккал, или 50 кДж энергии. [c.22]
Концентрация клеточного сока, его осмотическое давление, несомненно, имеют важное значение для поглощения воды клеткой. Это обстоятельство не дает, однако, оснований рассматривать процесс поглощения воды как чисто осмотический и тем самым исключать участие в нем живой протоплазмы. Известно, что основная часть биоколлоидов клетки принадлежит к гидрофильным соединениям, способным к обратимым изменениям степени своей оводненности. Поглощая воду, коллоидная мицелла набухает, при отдаче же воды коллоидной мицеллой происходит ее так называемое отбухание. Развиваемые в ходе этих процессов силы весьма значительны и нередко достигают сотен атмосфер. Сила, которую нужно приложить к коллоидной системе, чтобы предотвратить поглощение ею воды, называется давлением набухания. [c.73]
Корневое давление было открыто еще в первой половине XVIII в. и с тех пор довольно интенсивно изучается. Высказан ряд весьма противоречивых гипотез относительно его природы. Диапазон этих гипотез достаточно широк — от объяснений "эльса и Найта, относящихся соответственно к 1727 и 1801 гг. 259], Л, В. Можаевоп и Н. В. Пильщиковой [260—265], Д. Боса [266], А. П. Петрова [244], согласно которым воду нагнетают главным образом живые клетки корня, до сформулированных в конце прошлого столетия представлений, согласно которым живые клетки не принимают активного участия в движении воды через корневые системы или даже препятствуют этому движению [259]. Особенно широкое распространение получила предложенная в 1920-х годах схема Пристли [267, 268], уподобляющая нагнетающую деятельность корня работе осмометра, В наши дни эта схема в несколько модифицированном виде известна как осмотическая концепция экссудации. Она неизменно упоминается во всех учебниках и обзорах. Согласно данной концепции, роль находящегося внутри осмометра концентрированного раствора выполняет ксилемный сок, а роль полупроницаемой перегородки, препятствующей выходу наружу осмотически действующих веществ ксилемного сока, отводится [c.118]
По схеме Д. Пристли (рис. 14), А — живые клеткн, С — клетки сосудистой системы, в которых нет тургорного давления, следовательно, S = P. При полном насыщении водой группы клеток /1, которые погружены в сосуд В, сосущая сила вскоре будет равна 0. Пока Р>Т, вода поступает в клетку, но когда осмотическое давление уравновесится тургорным давлением, сосущая сила равна О, воду будут всасывать клетки сосудистой системы с си, 10й, равной величине их осмотического давления. Таким образом, клетк1г С будут всасывать воду из сосуда В, фильтруя се через живые клетки А. [c.118]
В тканях растений осмотич. давление составляет 0,5-2 МПа (у растений в пустынях - более 10 МПа). Гидростатич. давление, возникшее яо внутриклеточных структурах в результате осмоса, наз. тургором. Это давление придает прочность и упругость тканям живых организмов. Если клетка отмирает, оболочка теряет св-во полупроницаемос-ти, тургор исчезает (растение вянет). Осмотич. давление-главная причина, обеспечивающая движение воды в растениях и ее подъем от корней до вершины. Клетки листьев, теряя воду, осмотически всасывают ее из клеток стебля, а последний-из клеток корня, берущих, в свою очередь, воду из почвы. Для роста и развития растений важно соотношение между осмотич. давлением почвенного р-ра и клеточного сока. Растение может нормально развиваться лишь тогда, когда осмотич. давление клеточного сока больше осмотич. давления почвенного р-ра. [c.419]
Гипотеза Мюнха чисто физическая, и не объясняет, почему ситовидные трубки должны оставаться живыми и метаболически активными. Она не объясняет также, каким образом клетки мезофилла листа способны загружать ситовидные трубки ассимилятами против осмотического градиента известно, что у флоэмы /о более отрицательный, чем у фотосинтезирующей ткани. С учетом этого гипотеза Мюнха бьша впоследствии дополнена — в нее включили механизм активной загрузки растворенных веществ в ситовидные трубки. Он подразумевает, что осмотический и гидростатический градиенты начинаются не в фотосинтезирующем мезофилле, а непосредственно во флоэме. Кроме того, полагают, что разгрузка флоэмы на уровне потребителей — тоже активный процесс. Такая современная версия гипотезы Мюнха называется гипотезой тока под давлением. [c.135]
Возникающее вследствие осмотического дисбаланса этих двух сред избыточное гидростатическое давление внутри растительной клетки, называемое тургорным давлением (или просто тургором), имеет для растений жизненноважное значение. Тургор - главная сила, растягивающая клетку в период ее роста он в значительной мере ответствен также за жесткость живых растительных тканей (сравните увядший лист обезвоженного растения с упругими листьями растения, получающего достаточно воды). [c.389]
chem21.info
Работа 6. Определение потенциального осмотического давления клеточного сока методом плазмолиза
Цель:определить осмотическое давление клеточного сока синего лука плазмолитическим методом.
Объекты, реактивы, оборудование: Луковица синего лука с чешуями, 1М раствор сахарозы или KNO3, безопасные бритвы, микроскопы, предметные и покровные стекла, бюксы, градуированные пипетки на 10 мл, препаровальные иглы, часы, фильтровальная бумага, термометр.
Краткие сведения
Клеточный сок — водный раствор различных органических и неорганических веществ. Потенциальное осмотическое давление зависит от числа частиц, находящихся в этом растворе, т. е. от концентрации и степени диссоциации растворенных молекул. Потенциальное осмотическое давление выражает максимальную способность клетки всасывать воду. Величина этого показателя указывает на возможность растения произрастать на почвах различной водоудерживающей силы. Повышение осмотического давления при засухе служит критерием обезвоживания растений и необходимости полива.
Данный метод основан на подборе такой концентрации наружного раствора, которая вызывает самый начальный (уголковый) плазмолиз в клетках исследуемой ткани. В этом случае осмотическое давление раствора примерно равно осмотическому давлению клеточного сока. Такой наружный раствор называют изотоническим.
Ход работы
В бюксах готовят по 10 мл растворов согласно форме таблицы 1. Для опыта можно взять сахарозу 1 М (или KNO3) и с помощью разбавления дистиллированной водой получить нужную концентрацию. Растворы тщательно перемешивают, бюксы закрывают крышками, чтобы предотвратить испарение, и ставят в ряд по убывающей концентрации.
Лезвием безопасной бритвы делают тонкие срезы с выпуклой поверхности пигментированной чешуи луковицы размером примерно 25 мм2 из среднего хорошо окрашенного участка.
В каждый бюкс, начиная с высокой концентрации, с интервалом 3 мин опускают по два-три среза. Через 30 мин после погружения срезов в первый бюкс их исследуют под микроскопом. Затем через каждые 3 мин наблюдают срезы из последующих бюксов. Таким способом достигают равную продолжительность пребывания срезов в растворах плазмолитиков. Срезы рассматривают под микроскопом в капле раствора из того бюкса, откуда они были взяты.
Определяют стадию плазмолиза (см. рис. 1 работы 1) клеток в каждом растворе и находят изотоническую концентрацию как среднюю арифметическую между концентрацией, при которой плазмолиз только начинался, и концентрацией, которая уже не вызывает плазмолиза.
Результаты опыта записывают в таблицу 1 по приведенной форме.
Таблица 1
| Концентрация раствора, моль/л | На 10 мл раствора | Стадия плазмолиза | Изотоническая концентрация, моль/л | Потенциальное осмотическое давление, кПа | |
| 1М раствора сахарозы или КNO3, мл | Воды, мл | ||||
| 0,7 | |||||
| 0,6 | |||||
| 0,5 | |||||
| 0,4 | |||||
| 0,3 | |||||
| 0,2 | |||||
| 0,1 |
Потенциальное осмотическое давление
Π = RcTi , где R – универсальная газовая постоянная, равная 8,314 Дж/моль▪К; Т- абсолютная температура в Кельвинах; с- изотоническая концентрация, М; i- изотонический коэффициент Вант-Гоффа.
Коэффициент Вант-Гоффа характеризует ионизацию растворов: I = 1+ α(n-1), где α – степень диссоциации раствора данной концентрации, n – число ионов, на которое диссоциирует соль.
Так как неэлектролиты не диссоциируют, для сахарозы i= 1.
Степень диссоциации KNO3 разной концентрации приведена ниже.
| Концентрация | 0,5 | 0,4 | 0,3 | 0,2 | 0,1 |
| Степень | 0,71 | 0,74 | 0,76 | 0,79 | 0,83 |
В зависимости от вязкости цитоплазмы в клетках чешуи репчатого лука осмотическое давление варьирует, как правило, от 300 до 1300 кПа.
Определить осмотическое давление в клетках синего лука при комнатной температуре (200С) и при температуре 4 0С.
Работа 7. Определение сосущей силы (водного потенциала) тканей растений по изменению их размеров (метод Уршпрунга)
Цель:определить сосущую силу, тургорное и осмотическое давление растительных тканей
Объекты, реактивы, оборудование: свежие и подвявшие клубни картофеля, корнеплоды сахарной или столовой свеклы; молярные растворы сахарозы от 0,1 до 0,9 М, алюминиевые бюксы, разделочная доска, ножи, лезвия, пинцеты, линейки.
Краткие сведения
Сила, с которой клетка в данный момент поглощает воду, называется сосущей. Сосущая сила клетки зависит от ее физиологического состояния и от внешних условий. В покоящихся семенах и меристематических клетках она обусловлена главным образом давлением набухания коллоидов протоплазмы и пектиновых веществ клеточных оболочек. В клетках, закончивших рост и имеющих большую центральную вакуоль, сосущая сила в значительной степени определяется величиной осмотического давления клеточного сока π и тургорного давления Р, которое в свою очередь зависит от эластичности клеточной оболочки и содержания воды в клетке.
Осмотическое давление окружающего раствора (π) равно: π = iСRТ. Сосущая сила клетки обычно равна разности осмотического давления клеточного сока и тургорного давления: S кл. = π кл.сока __ Р. В зависимости от насыщения клетки водой величина тургорного давления будет меняться, соответственно изменится и сосущая сила клетки.
Водообмен между клеткой и окружающей средой определяется соотношением сосущей силы клетки и осмотическим давлением наружного раствора. Поглощение или отдача воды клетками сопровождается изменением как их размеров и веса, так и концентрации окружающего раствора. При погружении кусочка ткани растения в раствор с большим осмотическим давлением вода из клеток поступает в раствор и размеры кусочка уменьшаются (Р=0, следовательно, S = π). Если сосущая сила клеток выше, чем π окружающего раствора, клетки всасывают воду и кусочек ткани увеличивается. При равенстве сосущей силы ткани и π окружающего раствора между выходом и поступлением воды в клетку устанавливается равновесие и размеры кусочка ткани не изменяются.
Задача настоящей работы сводится к тому, чтобы из серии растворов найти такой, осмотическое давление которого равнялась бы сосущей силе клеток ткани. Зная, что S кл. = π р-ра, находим π р-ра. Осмотическое давление раствора (π р-ра) легко рассчитать, зная его молярную концентрацию.
Однако в настоящее время для характеристики энергетического уровня молекул воды (их способности диффундировать или испаряться) используется термодинамический показатель – водный потенциал, который для чистой воды принят за нуль (ΨН2О = 0), а для любого раствора – меньше нуля. При замене осмотических показателей растительной клети (Sкл =πкл.сока __ Р) термодинамическими вышеприведенное уравнение примет следующий вид:
— Ψ Н2Окл = — Ψп + Ψр,
где ΨН2Окл – водный потенциал клетки; Ψп – осмотический потенциал клеточного сока; Ψр – гидростатический потенциал.
Из уравнения видно, что осмотический потенциал понижает водный потенциал клетки, а потенциал давления повышает его. Как правило, ΨН2О клетки отрицателен, и лишь при полном насыщении клетки водой, когда Ψр = Ψп, этот показатель равен нулю.
При погружении растительной клетки в какой-либо раствор водообмен между ними определяется соотношением их водных потенциалов: вода перемещается в сторону более отрицательного водного потенциала.
Ход работы
Поставить бюксы с растворами сахарозы указанной молярности на лабораторный стол. Вырезать из клубня или корнеплода (поперек продольной оси органа) пластинку толщиной 3-4 мм в форме прямоугольника размером 30х40 мм. С помощью лезвия и линейки разрезать ее на ряд одинаковых полосок величиной 3х40мм (нарезать полоски следует быстро, не допуская подвядания). Излишки клеточного сока, вытекающие при разрезании ткани, удалить фильтровальной бумагой. Погрузить по 3 полоски в растворы сахарозы (погружение должно быть полным). Через 30 мин извлечь полоски из растворов и измерить. Данные повторностей и средние из них записать по образцу в табл. 1.
Таблица 1
| Концентрация сахарозы, М | 0,9 | 0,7 | 0,5 | 0,3 | 0,2 | 0,1 | ||
| Длина полосок, мм | Исходная (lu) | |||||||
| через 30 мин (l0) | Повтор- ность | |||||||
| В среднем | ||||||||
| Разность до и после погружения в раствор (∆l),мм | ||||||||
| ∆l, % |
На основании измерений рассчитывают % изменения длины полосок в каждом растворе по формуле:
∆
Данные заносят в таблицу. Используя их, строят график зависимости изменения длины полосок в % от концентрации наружного раствора. На оси абсцисс откладывают концентрации (С) приготовленных растворов, а на оси ординат — ∆l, %.
Объяснить причину изменения длины полосок в растворах разных концентраций. На основании графика выяснить следующие концентрации растворов, где клетки находятся: в состоянии полного тургора, частичного тургора, его отсутствия. Отмечают гипертонические, гипотонические и изотоническую концентрацию наружного раствора по отношению к концентрации клеточного сока растительной ткани.
Для определения величины сосущей силы клеток растительной ткани исходят из того, что в изотоническом растворе величина сосущей силы клеток равна осмотическому давлению наружного раствора, которое определяется по уравнению Вант-Гоффа π = iСRТ.
Определить раствор, сосущая сила которого равна сосущей силе данной ткани.
Рассчитать сосущую силу ткани по сосущей силе раствора.
Исходя из полученных результатов работы, вычертить диаграмму, отражающую зависимость между осмотическим давлением, водным потенциалом (сосущей силой) и тургорным давлением ткани. Для этого необходимо заполнить табл. 2.
Таблица 2
| Концентрация сахарозы, М | L (длина полосок ткани через 30 мин), мм | S (сосущая сила клеток), Па S=сRT | π (осмотическое давление клеточного сока), Па | Р = π – S (тургорное давление),Па |
| 0,9 0,7 0,5 0,3 0,2 0,1 |
В 1-ую графу записывается молярная концентрация использованных растворов сахарозы.
Во 2-ую __ длина полосок ткани после их 30-минутного пребывания в соответствующем растворе.
В 3-ю __ значение сосущей силы ткани в соответствующем растворе. Поскольку полоски находились в растворах достаточно продолжительное время и размеры их перестали меняться, то можно считать, что сосущая сила их уравнялась с осмотическим давлением окружающего раствора, т.е. S тк. = π р-ра; π р-ра легко вычислить по формуле π = iСRТ.
В 4-ую графу вносятся значения π клеточного сока. Принимаем, что для самой короткой полоски Р = 0, тогда, исходя из формулы S = π – Р, π1 = S1 , В растворах меньшей концентрации полоски ткани будут иметь более низкие значения π, которые уменьшаются обратно пропорционально длине полосок ткани:
π·L1 = πn. Ln , значит πn = π L1/ Ln
Имея все значения S и π, можно заполнить последнюю графу таблицы, рассчитав значения Р для исследованных полосок ткани по формуле
π = Р — S
Далее необходимо вычертить диаграмму (рис.1).
На оси абсцисс в масштабе 1см : 1мм откладывают значения длины полосок ткани в порядке возрастания; на оси ординат ___ значения π, затем Р для соответствующих полосок. Откладывать значения S нет необходимости, так как на графике они представляют отрезки πn__ Рn . Соединив точки π и Р линиями, получим график зависимости π, S и Р. На основании этого требуется сделать вывод о взаимной зависимости трех величин и объяснить, от какого показателя, π или Р зависит в основном изменение сосущей силы отрезков ткани.
Рис. 1. Диаграмма зависимости между сосущей силой, осмотическим и тургорным давлением растительных клеток, по-разному насыщенных водой.
infopedia.su