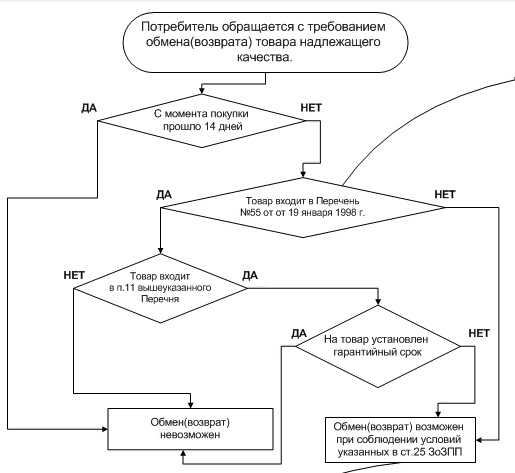
Сосудистые споровые. Наиболее примитивные споровые сосудистые растения включает отдел
Сосудистые споровые - Gpedia, Your Encyclopedia
| Cyathea dregei | |
| Сосудистые споровые растения | |
| Устаревшее таксономическое | |
| Pteridophyta | |
| Клада Высшие растения (Embryophyta) | |
Сосудистые споровые растения — собирательное название для группы высших растений, в которую входят папоротники и близкие к ним группы растений. В качестве научного названия для сосудистых споровых ранее использовалось название Pteridophyta[k 1]. В качестве русского названия для этой группы ранее использовался также термин «сосудистые тайнобрачные». Термин «папоротникообразные» обычно используется как синоним термина «сосудистые споровые растения», однако иногда под «папоротникообразными» понимают папоротниковидных — более узкую группу.
Группа Pteridophyta была общепринятой в системах классификации растений XIX века, позже она была признана искусственной[1].
Совокупность сосудистых споровых растений и мохообразных образуется группу высших споровых растений (ранее рассматривавшуюся как таксон, а в настоящее время — как неформальную группу).
Состав группы
Обычно в состав сосудистых споровых растений включают две современные группы, рассматриваемые как самостоятельные ботанические отделы:
Кроме этих групп, к сосудистым споровым растениям относят три вымерших отдела:
В 35-м издании знаменитого немецкого учебника ботаники Э. Страсбургера для высшей школы Lehrbuch der Botanik für Hochschulen (нем.)русск. (2002, русское издание — 2007) сосудистым споровым растениям соответствует подотдел Pteridophytina — один из четырёх подотделов в составе отдела Streptophyta: помимо Pteridophytina, в этот отдел в ранге других подотделов включены Зелёные водоросли (Streptophytina), Мхи (Bryophitina) и Семенные растения (Spermatophytina). В подотдел Pteridophytina в этом учебнике включено пять классов: †Псилофиты (Psilophytopsida), Плауны (Lycopodiophyta), Хвощевидные (Equisetopsida), Псилотовые (Psilotopsida) и Папоротники (Pteridopsida)[2].
Общая характеристика
Спорофит (бесполое поколение) является для сосудистых споровых доминирующей формой жизненного цикла (в отличие от мохообразных, в жизненном цикле которых доминирует половое поколение). Для спорофита современных сосудистых споровых характерна дифференциация на корни, стебель и листья. Нередко имеются сосуды. Размножение происходит с помощью спорангия — специального органа, в котором развиваются споры. Из спор развивается половое поколение растений[1][2].
Для заростка — полового поколения сосудистых споровых (гаметофита) характерно отсутствие дифференциации тканей. На заростках развиваются половые органы размножения — женские (архегонии) и мужские (антеридии). Заростки имеют существенно меньшие размеры по сравнению со спорофитами того же вида (обычно не более нескольких сантиметров, заростки некоторых видов имеют микроскопические размеры). Время жизни заростка обычно очень небольшое (несколько недель): он отмирает вскоре после того, как на нём разовьются половые органы и произойдёт оплодотворение[2].
Птеридология
Изучением сосудистых споровых растений (папоротникообразных) занимается наука птеридология. Учёные, специалисты в области птеридологии, называются птеридологами.
См. также
Комментарии
Примечания
Литература
- Жизнь растений. В 6-ти т. / Гл. ред. Ал. А. Фёдоров. — М.: Просвещение, 1978. — Т. 4. Мхи. Плауны. Хвощи. Папоротники. Голосеменные растения. Под ред. И. В. Грушвицкого и С. Г. Жилина. — 447 с. — 300 000 экз.
- Папоротникообразные, птеридофиты (Pteridophytina) // Ботаника. Учебник для вузов: в 4 т. = Lehrbuch der Botanik für Hochschulen. Begründet von E. Strasburger, F. Noll, H. Schenck, A. F. W. Schimper. / 35. Auflage neubearbeitet von Peter Sitte, Elmar W. Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner / П. Зитте, Э. В. Вайлер, Й. В. Кадерайт, А. Брезински, К. Кёрнер; на основе учебника Э. Страсбургера [и др.]; пер. с нем. Е. Б. Поспеловой, К. Л. Тарасова, Н. В. Хмелевской. — М.: Издательский центр «Академия», 2007. — Т. 3. Эволюция и систематика / под ред. А. К. Тимонина, И. И. Сидоровой. — С. 305—350. — 576 с. — 3000 экз. — ISBN 978-5-7695-2741-8 (рус.). ISBN 978-5-7695-2746-3 (Т. 3) (рус.), ISBN 3-8274-1010-X (Elsevier GmbH) — УДК 58(075.8)
Ссылки
www.gpedia.com
| Устаревшее таксономическое | |||
| Pteridophyta | |||
| подцарство Высшие споровые растения (Embryophytes) | |||
| См. текст | |||
| |||
www.turkaramamotoru.com
Сосудистые растения - это... Что такое Сосудистые растения?
Сосу́дистые ра́стения (лат. Tracheophyta, Plantae Vasculares, Tracheobionta) — подцарство растений, ткани которых пронизаны сосудисто-волокнистыми пучками. К этой группе относятся все высшие растения за исключением мохообразных[1].
Остальные растения, не заключающие в себе сосудистых пучков, называются клеточными (лат. Plantae Cellulares). Это деление предложено Огюстеном Пирамом Декандолем в 1813 году. К сосудистым растениям относятся как высшие споровые (Псилотовидные, Папоротниковидные, Плауновидные, Ужовниковидные и Хвощевидные), так и все семенные растения.
См. также
Примечания
Литература
- Черепанов С. К. Сосудистые растения России и сопредельных государств. — СПб., 1995.
Ссылки
Категории:- Растения по алфавиту
- Высшие растения
Wikimedia Foundation. 2010.
- Гершфельд, Давид Григорьевич
- Уборки
Смотреть что такое "Сосудистые растения" в других словарях:
СОСУДИСТЫЕ РАСТЕНИЯ — СОСУДИСТЫЕ РАСТЕНИЯ, растения, имеющие сосуды или трубки, по которым внутри них переносится вода и питательные вещества. Все высшие растения СПОРОВЫЕ, ХВОЙНЫЕ, ЦВЕТКОВЫЕ имеют сосудистую систему (КСИЛЕМУ и ФЛОЭМУ). Эта система расположена в… … Научно-технический энциклопедический словарь
СОСУДИСТЫЕ РАСТЕНИЯ — (Plantae vasculares, или Tracheophyta), растения, в органах к рых имеются сосуды или трахеиды, проводящие воду, а также растворённые в ней минеральные соли, и ситовидные элементы, проводящие органич. вещества. Для С. р. характерно преобладание в… … Биологический энциклопедический словарь
Сосудистые растения — растения, в органах которых имеются сосуды или трахеиды, проводящие воду и растворённые в ней минеральные соли, и ситовидные трубки, проводящие органические вещества. К С. р. относят псилотовые, плауновидные, хвощевидные,… … Большая советская энциклопедия
Сосудистые растения — (Plantae Vasculares) отдел растений, ткани которых пронизаны сосудисто волокнистыми пучками (см. это сл.). Остальные растения, не заключающие в себе сосудистых пучков, называются клеточными (Pl. cellulares). Это деление предложено Августом… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Растения — Многоо … Википедия
Сосудистые споровые — Группа растений Cyathea dregei … Википедия
Растения ископаемые — Сведения наши об ископаемых Р. представляют значительные пробелы как ввиду нежности многих органов Р., лишь изредка и не вполне совершенно сохраняющихся в пластах земной коры, так и ввиду полного отсутствия сходства и определенных соотношений… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Сосудистые споровые — или сосудистые тайнобрачные (Pteridophyta) такие споровые растения, у которых бесполое поколение вполне развито и расчленено на главнейшие органы: корень, стебель и лист; в органах проходит сосудисто волокнистые пучки. Бесполое поколение… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Сосудистые болезни растений — болезни, характеризующиеся потерей Тургора всем растением или отдельными его частями вследствие нарушения функций сосудистой системы растений. Причины: недостаток воды в почве; нарушение деятельности корневой системы; закупорка проводящей … Большая советская энциклопедия
СОСУДИСТЫЕ БОЛЕЗНИ РАСТЕНИЙ — характеризуются потерей тургора всем р нием или отд. его частями вследствие нарушения функций сосудистой системы р ний. Причины: недостаток воды в почве; нарушение деятельности корневой системы; закупорка проводящей системы р ний мицелием… … Сельско-хозяйственный энциклопедический словарь
dic.academic.ru
Отделы споровых сосудистых растений - СПОРОВЫЕ СОСУДИСТЫЕ РАСТЕНИЯ - РАЗНООБРАЗИЕ - ВВЕДЕНИЕ В БОТАНИКУ - СОВРЕМЕННАЯ БОТАНИКА
РАЗДЕЛ IV. РАЗНООБРАЗИЕ
ГЛАВА 17. СПОРОВЫЕ СОСУДИСТЫЕ РАСТЕНИЯ
Отделы споровых сосудистых растений
Три отдела этих растений — риниофиты (Rhyniophyta), зостерофиллофиты (Zosterophyllophyta) и тримерофиты (Тrimerophyta) — процветали в девонский период и вымерли к его концу, т. е. около 360 млн. лет назад, причем риниофиты появились по крайней мере 420 млн. лет назад, в позднем силуре. Все это относительно простые по структуре организмы. Четвертый отдел споровых Progymnospermophyta, или прогимноспермы, будет рассматриваться в гл. 18, так как его представители, возможно, были предками голосеменных и покрытосеменных (рис. 17-7). Кроме вымерших растений мы обсудим в этой главе отделы Рsilotophyta, Lycophyta, Sphenophyta и Pterophyta, в которых имеются ныне живущие представители.
Рис. 17-7. Возможный ход эволюции сосудистых растений, начиная от зостерофиллофитов и риниофитов. Продолжительность девонского периода в общем масштабе временной шкалы увеличена, не показаны связи с цветковыми растениями. Не ясно, от кого произошли псилотофиты и хвощевидные. Описано всего несколько родов ископаемых растений, являющихся, возможно, промежуточными между риниофитами, появившимися в силурийском периоде (около 420 млн. лет назад), и
зостерофиллофитами, древнейшие остатки которых примерно на 10 млн. лет моложе. Точную природу взаимосвязей между этими двумя группами и их общего предка еще предстоит установить
Общую модель нарастания разнообразия флоры можно представить, как поочередное наступление господства четырех крупных групп, сопровождающееся резким вытеснением предыдущих доминантов. В каждом случае группы, захватывая ведущее положение, достигали высокого видовою богатства. Основное участие в этом историческом процессе принимали:
1. Первые сосудистые растения с простой и, вероятно, исходной для прочих групп морфологией. К ним относятся риниофиты, зостерофиллофиты и тримерофиты (рис. 17-8). Эти примитивные формы доминировали с позднего силура до среднего девона, т. е. примерно 420 — 370 млн. лет назад (см. рис. 17-1).
2. Папоротники, плауновидные, хвощевидные, прогимноспермы. Эти более сложно устроенные растения господствовали с позднего девона до конца каменноугольного периода (см. рис. 17-9, 18-1), примерно 380 — 290 млн. лет назад. Приложение к главе дает представление о растениях той эпохи (см. с. 307).
3. Семенные растения, появившиеся в позднем девоне (по крайней мере, 360 млн. лет назад) и к началу пермскою периода уже представленные многими эволюционными линиями. Голосеменные преобладали во флорах суши на протяжении большей части мезозойской эры, примерно до 100 млн. лет назад.
4. Цветковые растения, древнейшие остатки которых имеют возраст около 127 млн. лет. Они завоевали сушу за 20 — 30 млн. лет и с тех пор сохраняют господствующее положение.
Отдел Rhyniophyta
К этому отделу принадлежат самые древние из детально известных нам сосудистых растений. Они появились в позднем силуре (не менее 420 млн. лет назад) и вымерли в среднем девоне (около 380 млн. лет назад). Еще более древние сосудистые растения, ископаемые остатки, которых по крайней мере на 15 млн. лет старше, были, вероятно, сходны с риниофитами. Отдел включает споровые растения, состоящие из простых, дихотомически ветвящихся осей с верхушечными спорангиями. Дифференцировка на корни, стебли или листья отсутствовала, виды были равноспоровыми. Название отдела происходит от деревни Райни1 в Шотландии, близ которой остатки этих примитивных растений прекрасно сохранились в кремнистых породах.
1По-латыни ее название (Rhynie) читается «Риниэ». — Прим. ред.
Куксония (Cooksonia), представитель риниофитов, обитавший, как полагают, на заболоченных низинах, — древнейшее из известных нам сосудистых растений (см. рис. 1-5, 17-1). Ее тонкие безлистные надземные стебли достигали в длину 6,5 см; спорангии были шаровидными. Хотя о нижних частях этих растений ничего не известно, вполне вероятно, что куксония имела корневище, т. е. подземный стебель, от которого отходили надземные ветви. В мацерированных кусочках осей обнаружены трахеиды. Куксония вымерла к раннему девону, около 390 млн. лет назад.
Наиболее известный представитель риниофитов — риния (Rhynia) (рис. 17-8, A), вероятно болотное растение, безлистные, дихотомически ветвящиеся стебли которого отходили от корневища с пучками водопоглощающих ризоидов. Надземные стебли ринии длиной 20 — 50 см и толщиной 3 — 6 мм были покрыты кутикулой, имели устьица и служили фотосинтезирующими органами.
Рис. 17-8. Древние сосудистые растения. А. Риниофит Rhynia major — одно из наиболее простых известных нам сосудистых растений. Его безлистный стебель дихотомически ветвился. Герминальные спорангии высвобождали споры, вероятно, за счет продольного расщепления. Б. У рода Zosierophyllum и других зостерофиллофитов собранные в терминальные колоски спорангии расщеплялись по определенным бороздам вокруг своего дистального кран. Более крупные, чем риниофиты, эти растения, как и последние, как правило, дихотомически ветвились, были голыми, шиповатыми или покрытыми зубчиками. R. Тримерофиты — еще более крупные и сложнее устроенные растения с крепкой центральной осью и боковыми ветвями меньшего размера, которые дихотомически ветвились и часто заканчивались группами парных, суженных к концам спорангиев. Наиболее известные роды из этой группы — Psilophyton и Trimerophyton. На рисунке приведена реконструкция вида Psiíophyton princeps. Отдельные экземпляры Rhynia major достигали в высоту 0,5 м, а некоторые тримерофиты — более 1 м (см. также рис. 17-1)
Рис. 17-9. Реконструкция болотистого леса каменноугольного периода (см. также рис. 18-1)
Рис. 17-10. Psilotum nudum. А. Поперечный разрез стебля. показывающий сформировавшиеся ткани. Б. Протостела; видны ксилема и флоэма
Рис. 17-11. Подземный гаметофит Psilotum nudum. Гаметофиты обоеполые, т. е. несут и антеридии, и архегонии
Рис. 17-12. A. Tmesipteris parva на стволе древовидного папоротника Cyathea australis в Новом Южном Уэльсе, Австралия. Б. Tmesipteris lanceolate из Новой Каледонии
Рис. 17-13. Жизненный цикл псилота. У этого рода из корневищ вырастают дихотомически ветвящиеся надземные побеги. На них в пазухах чешуевидных выростов группами по три развиваются спорангии. Для псилота, как и для всех сосудистых растений, характерно чередование гетероморфных поколений с доминированием спорофита. В результате мейоза внутри спорангия образуются споры, и начинается гаметофитная фаза. Псилот равноспоровый; его споры дают начало обоеполым, т. е. несущим одновременно антеридии и архегонии, гаметофитам. Чтобы достичь яйцеклетки, многожгутиковый спермий нуждается в воде. При сингамии образуется зигота, из которой развивается спорофитное поколение. Зародыш формируется в брюшке архегония, причем молодой спорофит получает питание от гаметофита через свою стопу, внедренную в его ткань. В конечном итоге спорофит отделяется от стопы и начинает независимое существование
Рис. 17-14. А. Поперечный срез стебля Lycopodium complanatum, на котором видны зрелые ткани. Б. Протостела этого же вида; видны ксилема и флоэма. И стебель, и корень плаунов протостелические (см. также рис. 17-2)
Рис. 17-15. Lycopodium lucidulum —представитель плаунов, не имеющих стробила; спорофиллы рассеяны среди стерильных микрофиллов
По внутренней структуре риния напоминала современные сосудистые растения. Единственный слой поверхностных клеток — эпидерма — окружал ассимиляционную ткань коры, а центр оси состоял из сплошного тяжа ксилемы, окруженного двумя слоями клеток, возможно, флоэмных. По- видимому, первыми развивались клетки ксилемы в центральной части тяжа, а последними — на его периферии.
Рис. 17-16. А. Selaginella kraussiana — стелющееся, ползучее растение. Видны придаточные корни, образующиеся на стеблях. Б. S. rupestris со стробилами. В. S. willdenowii — растение тропиков Старого Света, встречающееся во Вьетнаме, Малайзии и Гималаях. Этот теневыносливый вид с переливчатыми, металлически блестящими голубыми листьями может забираться по деревьям на семиметровую высоту
Отдел Zosterophyllophyta
Ископаемые остатки представителей этого отдела вымерших сосудистых растений находят в ранне позднедевонских отложениях возрастом примерно от 408 до 370 млн. лет. Как и риниофиты, они были безлистными и дихотомически разветвленными. Возможно, растения этой группы обитали в воде. Надземные стебли покрывала кутикула, но только самые верхние ветви имели устьица. Это может свидетельствовать о том, что нижние были погружены в ил. Предполагают, что у зостерофиллума (Zosterophyllum) эти ветви часто давали боковые ответвления, вильчато разделявшиеся на две оси, одна из которых росла вверх, а другая вниз (рис. 17-8, Б). Растущие вниз ответвления, по-видимому, служили опорой растению, позволяя ему распространяться в стороны. Свое название зостерофиллофиты получили из-за их общего сходства с современным покрытосеменным Zostera, растущим в морской воде и внешне напоминающим злаки.
В отличие от риниофитов спорангии зостерофиллофитов, имевшие шаровидную или бобовидную форму, располагались латерально на коротких ножках. Растения были равноспоровыми. Хотя внутренняя структура зостерофиллофитов в основном такая же, как у риниофитов, первыми достигавшие зрелости клетки ксилемы располагались здесь по периферии ксилемного тяжа, а развивавшиеся последними — в его центре.
Можно почти с полной уверенностью считать, что зостерофиллофиты — предки плауновидных. Спорангии древних плауновидных также располагались латерально, и ксилема в обоих отделах развивалась центростремительно (от периферии к центру). Представители двух этих отделов заметно отличаются от риниофитов и тримерофитов.
Отдел Trimerophyta
Этот отдел произошел, вероятно, непосредственно от риниофитов, а его представители, по-видимому, стали предками папоротников, прогимноспермов и, не исключено, также хвощей. Тримерофиты, более крупные и более сложно устроенные по сравнению с риниофитами и зостерофиллофитами растения (рис. 17-8, В), впервые появились в раннем девоне, около 395 млн. лет назад, и вымерли к концу среднего девона, примерно 20 млн. лет спустя, просуществовав, таким образом, относительно короткий промежуток времени.
Хотя тримерофиты более продвинуты в эволюционном отношении, чем риниофиты, они еще лишены листьев. На главной оси формировались системы боковых ветвей, неоднократно разделявшихся дихотомически. Эти растения, подобно риниофитам и зостерофиллофитам, были равноспоровыми. Некоторые из более мелких ветвей заканчивались удлиненными спорангиями, тогда как другие были исключительно вегетативными. Кроме усложнившегося способа ветвления тримерофиты имели и более массивный, чем у риниофитов, проводящий тяж, который вместе с мощной корой, состоявшей из толстенных клеток, вероятно, давал опору довольно крупному растению. Как и у риниофитов, первые клетки ксилемы у тримерофитов формировались в центре (центробежное развитие). Название отдела происходит от греческих слов «tri», «meros» и «phyton» и означает «растение из трех частей». В нем отражено тройчатое разделение вторичных ветвей у рода Trimerophyton.
Рис. 17-17. Жизненный цикл плауна. Это растение, как и псилот, равноспоровое, поэтому их жизненные циклы схожи. В результате мейоза образуются споры, дающие начало обоеполым гаметофитам. Последние у некоторых видов плауна подземные, и для их нормального роста, как показано на рисунке, требуется присутствие микоризного гриба. Чтобы произошло оплодотворение, необходима вода, в которой двужгутиковые спермии плывут к архегонию. При сингамии возникает зигота. Развитие зародыша идет внутри брюшка архегония. Молодой спорофит, иногда длительное время прикрепленный к гаметофиту, в конце концов становится независимым. Спорофиллы у многих видов плаунов собраны, как видно на рисунке, в стробилы
Рис. 17-18. Селагинелла. А. Поперечный срез стебля, на котором видны зрелые ткани. Протостела поддерживается в центре полого стебля удлиненными коровыми (эндодермальными) клетками, называемыми трабекулами. На рисунке видны только их фрагменты. Б. Протостела крупным планом
Рис. 17-19. Спорофит Isoetes muricata. Видны листья, похожие на иглы дикобраза, стебель и корни
Отдел Psilotophyta
Отдел включает два доживших до наших дней рода: псилот (Рsilotum) и тмезиптерис (Tmesipteris). Псилот распространен в тропиках и субтропиках; в США он встречается во Флориде, Луизиане, Аризоне, Техасе, на Гавайских островах и Пуэрто-Рико, а также как обычный оранжерейный сорняк. Ареал тмезиптериса ограничен Австралией, Новой Каледонией, Новой Зеландией и другими островами южной части Тихого океана. Оба рода — очень просто устроенные растения, напоминающие по своей основной структуре риниофиты.
Псилот уникален среди современных растений тем, что не имеет ни корней, ни листьев. Спорофит состоит из дихотомически ветвящейся надземной части с мелкими чешуевидными выростами и разветвленной подземной системы корневищ с многочисленными ризоидами (рис. 17-11). В наружных клетках коры корневищ присутствует эндомикоризный зигомицет. У псилота — протостела, следовательно, нет листовых прорывов (рис. 17-10).
Это растение равноспоровое; споры образуются в спорангиях, расположенных на концах коротких боковых ветвей. Прорастая, они дают начало обоеполым гаметофитам, которые похожи на фрагменты корневищ (рис. 17-11). Как и последние, подземный гаметофит содержит симбиотический гриб, а в некоторых случаях имеет проводящую ткань. Спермин псилота многожгутиковые и, чтобы достичь яйцеклетки, нуждаются в воде. Первоначально спорофит прикреплен к гаметофиту стопой — структурой, абсорбирующей питательные вещества из гаметофита. Впоследствии он от нее отделяется, а стопа остается погруженной в гаметофит.
Тмезиптерис растет как эпифит на древовидных папоротниках и других растениях (рис. 17-12). Листовидные придатки у него крупнее, чем чешуевидные выросты псилота, но в прочих отношениях эти два рода достаточно сходны.
Жизненный цикл псилота представлен на рис. 17-13.
Отдел Lycophyta
Четыре ныне живущих рода и примерно 1000 современных видов отдела Lycophyta (ликофиты, или плауновидные) представляют эволюционную линию, известную еще из девонского периода. Предками плауновидных почти с полной уверенностью можно считать зостерофиллофитов (см. рис. 17-8,5). Существует несколько порядков этого отдела; по крайней мере три из них, ныне вымершие, включали деревья, достигавшие иногда крупных размеров. Однако три современных порядка плауновидных состоят только из трав. И современные, и ископаемые ликофиты имеют микрофиллы; этот тип листьев — характернейшая черта отдела. Древовидные его представители были одними из доминирующих растений «углеобразующих» лесов каменноугольного периода (см. приложение к этой главе и рис. 18-1). Некоторые из них формировали структуры, сходные с семенами современных семенных растений. Большинство линий древесных плауновидных вымерло до конца палеозойской эры, т. е. не позже 248 млн. лет назад.
Lycopodium
Наверное, самыми известными из современных ликофитов являются плауны (род Lycopodium, см. рис. 10-7,5). Приблизительно 200 их видов встречаются от арктических областей до тропиков, хотя и редко играют заметную роль в растительных сообществах. Большинство тропических форм — эпифиты и поэтому редко бросаются в глаза, но несколько видов умеренных областей образуют ковры, иногда заметные в напочвенном покрове леса. Так как эти растения вечнозеленые, их лучше всего видно зимой.
Спорофит плауна состоит из ветвящегося корневища, от которого отходят надземные ветви и придаточные корни. И стебли, и корни протостелические (рис. 17-14). Микрофиллы обычно расположены спирально. Плаун — равноспоровое растение; спорангии расположены одиночно на верхней поверхности фертильных микрофиллов, называемых спорофиллами, — видоизменных листьев или листоподобных органов. У некоторых видов спорофиллы рассеяны среди стерильных микрофиллов и почти не отличаются от них (рис. 17-15). У других нефотосинтезирующие спорофиллы собраны в стробилы на верхушках надземных ветвей (см. рис. 10-7,5).
Споры плауна после прорастания дают начало обоеполым гаметофитам, которые в зависимости от вида представляют собой либо зеленые неправильно-лопастные образования, либо ветвящиеся подземные нефотосинтезирующие структуры. Как и у псилота и тмезиптериса, такие подземные гаметофиты вступают в симбиоз с грибом. Развитие и созревание архегониев и антеридиев у плаунов иногда продолжается от 6 до 15 лет. По мере роста гаметофита в последовательно созревающих архегониях может возникать целый ряд спорофитов.
Для оплодотворения необходима вода; двужгутиковый спермий подплывает в ней к архегонию и затем в ней же движется через его шейку. После оплодотворения зигота развивается в зародыш, растущий в брюшке архегония. Молодой спорофит может оставаться прикрепленным к гаметофиту в течение длительного времени, но в конце концов становится независимым.
Жизненный цикл плауна показан на рис. 17-17.
Selaginella
Среди современных родов плауновидных род селагинелла (Selaginella) насчитывает наибольшее число видов — около 700, распространенных главным образом в тропиках. Многие из них растут во влажных местообитаниях, но некоторые и в пустынных областях, впадая в покой вовремя наиболее сухой части года. Среди последних — так называемое «воскресающее» растение — Selaginella lepidophylla, ареал которого простирается от Мексики на север до Техаса и Нью- Мексико.
В основных чертах травянистые спорофиты селагинелл и плаунов схожи; здесь также наблюдаются микрофиллы, а спорофиллы образуют стробилы (рис. 17-16). В отличие от плаунов у селагинелл вблизи основания верхней поверхности каждого микрофилла и спорофилла развивается мелкий чешуевидный вырост, называемый язычком. Стебель и корень протостеличес
lifelib.info
Основы систематики - Flo natural flowers
Вид - основная таксономическая единица, объединяющая сходные между собой организмы, способные к скрещиванию друг с другом и прозводству плодовитого потомства. Виды, как таксоны, имеют двойные названия, в которых первое слово повторяет название рода, второе является видовым эпитетом.
Высшие растения - растения, тело которых расчленено на стебель, корень, лист. Имеют закономерное чередование полового и бесполого поколений - гаметофита спорофита К высшим относят растения следующих отделов:
- Моховидные (Bryophyta)
- Плауновидные (Lycopodiophyta)
- Хвощевидные (Equsetophyta)
- Папоротниковидные (Polypodiophyta)
- Голосеменные (Gymnospermae или Pinophyta)
- Покрытосеменные или Цветковые (Angiospermae или Magnoliophyta)
Голосеменные (отдел Pinophyta или Gymnospermae) - семенные растения, семязачатки которых располагаются открыто на поверхности мегаспорофиллов.Спорофиллы собраны в однополые стробилы или собрания стробилов, часто именуемые мужскими и женскими шишками.
Отдел представлен исключительно древесными обычно вечнозелеными растениями.Двудольные (класс Magnoliopsida или Dicotyledones) - цветковые растения:
- с двумя зародышевыми семядолями
- с листьями простыми или сложными, обычно разделенными на черешок и пластинку
- с перистым или пальчатым жилкованием листовых пластинок
- обычно с 4-х или 5-ти членным планом строения цветка
Класс - таксономическая категория или таксон рангом ниже отдела. Латинские названия классов, как таксонов, имеют стандартное окончание - psida.Полиподиевые или многоножковые,настоящие папоротники (класс Polypodiopsida) - во флоре области "типичные" папоротники с листьями - вайями, имеющими в раннем возрасте характерную форму "улитки".Спорангии развиваются на обыкновенных зеленых листьях, либо на специальных частях листа или на специализированных отдельных спорофиллах, утративших способность к фотосинтезу. Спорангии обычно собраны в группы (сорусы) на нижней, лучше защищенной, стороне листа.Гаметофиты класса Polypodiopsida - мелкие напочвенные зеленые растеньица.
Однодольные (класс Liliopsida или Monocotyledones) - цветковые растения:
- с одной зародышевой семядолею;
- с листьями всегда простыми, обычно не разделенными на черешок и пластинку;
- с жилкованием параллельным или дуговидным;
- с цветками в своей основе обычно 3-членными, редко 4- или 2-членными.
Отдел - одна из наиболее крупных таксономических категорий, используемых в систематике растений. Латинские названия конкретных отделов имеют стандартные окончания - phyta.
Офиоглоссовые или ужовниковые (класс Ophioglossopsida) - наиболее древняя и примитивная группа современных папоротников.
Большинство видов образует ежегодно только по одному листу, развивающемуся медленно, часто до трех и более лет. При этом в пределах единственного листа развиваются отдельные вегетативная и спороносная зоны.
Гаметофиты ужовниковых ведут подземный образ жизни, связаны с почвенными грибами, долгоживущие (существуют до 20 лет).
Папоротниковидные или папоротники (отдел Polypodiophyta или Pterophyta) - споровые сосудистые растения с крупными листьями, эволюционно произошедшими в результате уплощения целых ветвей (у хвощей и плаунов листья мелкие, образованные как простые выросты стебля).
Следствием веточной природы листьев папоротников является их длительный верхушечный рост, большие размеры, сложная форма листовой пластинки.
Гаметофиты папоротников - маленькие зеленые растеньица, фотосинтезирующие и питающиеся самостоятельно. Реже лишены хлорофилла и живут под землей, питаясь микоризно.
Плауновидные (отдел Lycopodiophyta) - споровые сосудистые растения, представленные в современной флоре травянистыми обычно вечнозелеными растениями.
Имеют характерное вильчатое ветвление побегов; мелкие чешуевидные листья; длинные, часто ползучие стебли. Спорофиллы собраны в стробилы или образуют на стебле отдельные спороносные зоны. Имеются также водные представители отдела, обитатели дна олиготрофных водоемов.
Наиболее древний отдел современных высших растений. Его вымершие представители (лепидодендроны, сигиллярии) обладали гигантскими стволами до 40 метров высоты и вместе с рядом гигантских хвощевидных образовали на Земле основные запасы каменного угля.
Среди плауновидных имеются равноспоровые и разноспоровые представители, объдиняемые в два класса: Lycopodiopsida и Isoetopsida.
Плауновые (класс Lycopodiopsida) - равноспоровые представители отдела Lycopodiophyta.
Спорофиты плауновых являются вечнозелеными обычно ползучими травянистыми растениями, с мелкими чешуевидными листьями, с вильчатым ветвлением побегов, с расположением спорофиллов среди обычных вегетативных листьев или в отдельных стробилах.
Гаметофиты подземные или полуподземные, ведут самостоятельный образ жизни, созревают за 1-15 лет, связаны микоризно с почвенными грибами.
Покрытосеменные или цветковые (отдел Angiospermae или Magnoliophyta) - семенные растения, мегаспорофиллы которых, срастаясь краями, образуют полость, к внутренней поверхности которой прикреплены cемязачатки При созревании семян мегаспорофиллы образуют их вместилище, называемое плодом.
Для всех представителей характерен цветок, как специальный орган полового размножения.
Полушниковые, или шильниковые (класс Isoetopsida) - разноспоровые представители отдела Lycopodiophyta, с сильно редуцированными однополыми гаметофитами, обычно не покидающими оболочку споры.
Класс представлен семейством Selaginellaceae (селагинелловые), виды которого миниатюрны в размерах и внешне напоминают мхи, и семейством Isoetaceae (полушниковые), виды которого являются водными растениями с характерным пучком шиловидных листьев, встречающиеся только в особо чистых олиготрофных водоемах.
Из-за антропогенного загрязнения водоемов полушники в настоящее время довольно быстро вымирают.
Растения семенные - растения, имеющие в качестве единицы размножения и расселения семена, сложные многоклеточные образования, имеющие сформированный зародыш нового организма, определенный запас питательных веществ, защитные покровы.Семенными являются растения двух отделов сосудистых растений:
- Gymnospermae (Голосеменные)
- Angiospermae (Покрытосеменные)
Растения споровые - растения, имеющие в качестве единицы размножения и расселения споры, одноклеточные образования, способные при прорастании к образованию самостоятельного организма. Споровыми являются растения следующих отделов сосудистых растений:
- Lycopodiophyta (Плауновидные)
- Equisetophyta (Хвощевидные)
- Polypodiophyta (Папоротниковидные или Папоротники).
Род - таксономическая категория или таксон, объединяющая виды растений. В конкретный род, как таксон, входит один или несколько видов.
Семейство - таксономическая категория или таксон, объединяющая роды растений. Латинские названия семейств, как таксонов, имеют стандартные окончания - aceae.
Сосудистые растения - высшие растения, имеющие сосуды. К сосудистым относят растения следующих отделов:
- Плауновидные (Lycopodiophyta)
- Хвощевидные (Equsetophyta)
- Папоротниковидные (Polypodiophyta)
- Голосеменные (Gymnospermae или Pinophyta)
- Покрытосеменные или Цветковые (Angiospermae или Magnoliophyta)
Таксон - группа организмов, отнесенных в процессе классификации к определенной таксономической категории (иначе рангу таксона).
В классификации высших растений используются следующие ранги таксонов или таксономических категорий: отдел, класс, порядок, семейство, род и вид.
В названиях таксонов выше рода употребляются унифицированные окончания: для семейств - "aceae"; для порядков - "ales"; для классов - "psida"; для отделов - "phyta".
В основу названия таких таксонов кладется обычно название какого-нибудь рода, входящего в этот таксон. Однако допускается употребление давно установившихся и традиционных названий для некоторых таксонов. Например, семейство крестоцветных с равным правом может называться - Brassicaceae (от Brassica) и Cruciferae; бобовые - Fabaceae (от Faba) и Leguminosae и другие.
В названиях видов используются так называемый принцип бинарной номенклатуры или двойных названий, введенный в систематику К.Линнеем в 1753 году. Первое слово в названии обозначает род, к которому относится данный вид, второе - видовой эпитет.
Ученый, впервые описавший таксон, является его автором. Фамилия автора помещается после латинского названия таксона, обычно в сокращенной форме. Так буква L. указывает на авторство Линнея.
Хвойные (класс Pinopsida или Coniferae) - деревья и кустарники обычно с характерными игольчатыми листьями (хвоeй).
Хвощевидные (отдел Equsetophyta) - споровые сосудистые растения, с характерными членистыми побегами и мутовками бурых редуцированных листьев, утративших хлорофилл.
Имеют специфичные щитковидные спорофиллы, собранные в стробилы на верхушках стеблей. Стебли хвощей - зеленые фотосинтезирующие, часто со значительным содержанием кремния.
Гаметофиты современных видов - маленькие, величиной в несколько миллиметров, зеленые наземные растеньица.
Все современные хвощевидные - травянистые растения. Древние представители отдела, например каламиты, были древовидными и наряду с гигантскими плауновидными образовывали каменноугольные леса.
Хвощовые (класс Equsetopsida) - единственный класс современных хвощевидных, повторяющий общие черты отдела.
flo.com.ua
Сосудистые споровые растения - Википедия
Материал из Википедии — свободной энциклопедии
| Группа растений | |
| Сосудистые споровые растения | |
| Устаревшее таксономическое | |
| Pteridophyta | |
| подцарство Высшие споровые растения (Embryophytes) | |
Сосудистые споровые растения — собирательное название для группы высших растений, в которую входят папоротники и близкие к ним группы растений. В качестве научного названия для сосудистых споровых ранее использовалось название Pteridophyta[k 1]. В качестве русского названия для этой группы ранее использовался также термин «сосудистые тайнобрачные». Термин «папоротникообразные» обычно используется как синоним термина «сосудистые споровые растения», однако иногда под «папоротникообразными» понимают папоротниковидных — более узкую группу.
Группа Pteridophyta была общепринятой в системах классификации растений XIX века, позже она была признана искусственной[1].
Совокупность сосудистых споровых растений и мохообразных образуется группу высших споровых растений (ранее рассматривавшуюся как таксон, а в настоящее время — как неформальную группу).
Состав группы[ | ]
Обычно в состав сосудистых споровых растений включают следующие современные группы, рассматриваемые как самостоятельные ботанические отделы:
Кроме этих групп, к сосудистым споровым растениям относят три вымерших отдела:
В 35-м издании знаменитого немецкого учебника ботаники Э. Страсбургера для высшей школы Lehrbuch der Botanik für Hochschulen (нем.) (2002, русское издание — 2007) сосудистым споровым растениям соответствует подотдел Pteridophytina — один из четырёх подотделов в составе отдела Streptophyta: помимо Pteridophytina, в этот отдел в ранге других подотделов включены Зелёные водоросли (Streptophytina), Мхи (Bryophitina) и Семенные растения (Spermatophytina). В подотдел Pteridophytina в этом учебнике включено пять классов: †Псилофиты (Psilophytopsida), Плауны (Lycopodiophyta), Хвощевидные (Equisetopsida), Псилотовые (Psilotopsida) и Папоротники (Pteridopsida)[2].
Общая характеристика[ | ]
Спорофит (бесполое поколение) является для сосудистых споровых доминирующей формой жизненного цикла (в отличие от мохообразных, в жизненном цикле которых доминирует половое поколение). Для спорофита современных сосудистых споровых характерна дифференциация на корни, стебель и листья. Нередко имеются сосуды. Размножение происходит с помощью спорангия — специального органа, в котором развиваются споры. Из спор развивается половое поколение растений[1][2].
Для заростка — полового поколения сосудистых споровых (гаметофита) характерно отсутствие дифференциации тканей. На заростках развиваются половые органы размножения — женские (архегонии) и мужские (антеридии). Заростки имеют существенно меньшие размеры по сравнению со спорофитами того же вида (обычно не более нескольких сантиметров, заростки некоторых видов имеют микроскопические размеры). Время жизни заростка обычно очень небольшое (несколько недель): он отмирает вскоре после того, как на нём разовьются половые органы и произойдёт оплодотворение[2].
encyclopaedia.bid
Систематика высших споровых растений » СтудИзба
Лекция № 9
Систематика высших споровых растений
ЦАРСТВО РАСТЕНИЯ – РLАNТАЕ
В современной систематике царство Растения делят на три подцарства: Багрянки, или Красные водоросли; Настоящие водоросли и Высшие растения, или Листостебельные. Багрянки часто называют низшими растениями: вегетативное тело их не расчленено на органы и ткани и также называется талломом. Однако у багрянок по сравнению с настоящими водорослями есть некоторые отличия.
Растения — царство эукариотических организмов, способных к фотосинтезу, имеющих плотные, как правило, целлюлозные клеточные стенки. Следовательно, преобладающее большинство растений по способу питания — автотрофы. Растения-паразиты с гетеротрофным типом питания всегда вторичного происхождения. Древнейшие растения известны с протерозойской эры (примерно 1,5 млрд лет назад). Со второй половины силурийского периода палеозойской эры на суше стали расселяться высшие растения, которые впоследствии стали господствовать на планете.
Особая, космическая роль зеленых растений состоит в том, что без них невозможна жизнь всех других живых организмов, в том числе и человека. Только хлорофилл, содержащийся в зеленых растениях, способен аккумулировать энергию солнца и превращать ее в энергию химических связей, что ведет к образованию органического вещества из неорганических веществ.
ПОДЦАРСТВО ВЫСШИЕ РАСТЕНИЯ – ЕМВRУОРНУТА
Высшие растения — наиболее дифференцированные автотрофные многоклеточные организмы, приспособленные главным образом к наземной среде.
Тело подавляющего большинства высших растений расчленено на побеги (стебли и листья) и корни. Высшие растения имеют ткани. Формирование тканей — неизбежный результат переселения растений из водной среды на сушу. Питательные вещества всасываются не всей поверхностью растения, как у водных, а специализированными проводящими клетками.
В подцарстве не менее 300 тыс. ныне существующих видов и огромное число вымерших. Известные виды высших растений делят на 9 отделов:
1. Риниевые.
2. Зостерофилловые.
3. Моховидные.
4. Плауновидныеа.
5. Псилотовидные.
6. Хвощевидные.
7. Папоротниковидные.
8. Голосеменные.
9. Покрытосеменные, или Цветковые.
Риниевые и зостерофилловые полностью вымерли. В остальных отделах есть как вымершие, так и ныне существующие виды. Среди высших растений (за исключением моховидных) спорофит преобладает над гаметофитом. В органах спорофита есть сосуды и трахеиды, поэтому их еще называют сосудистыми растениями.
Высшие растения делят на две очень неравные по значению и количеству видов группы — высшие споровые и семенные растения. У высших споровых гаметофиты и спорофиты — самостоятельные растения (за исключением моховидных, у которых спорофит развивается на гаметофите). Размножаются споровые растения спорами. К споровым относятся все отделы, кроме голосеменных и покрытосеменных растений.
Голосеменные и покрытосеменные — семенные растения, размножающиеся семенами. У семенных растений спорогенез и гаметогенез тесно связаны между собой. В процессе эволюции произошла сильная редукция женского и мужского гаметофита, поэтому редуцированный женский гаметофит (зародышевый мешок) развивается на спорофите, а мужской гаметофит (пылинка) переносится к яйцеклетке целиком. В результате оплодотворения яйцеклетки образуется диплоидная зигота, из которой развивается зародыш, окруженный специальными оболочками, или покровами. Зародыш с покровами образует семя. У голосеменных семена на семенных чешуях лежат открыто, а у покрытосеменных они находятся внутри завязи пестика, образованного одним или несколькими плодолистиками.
Считают, что высшие растения произошли от низших — обитателей водной среды, непосредственно из зеленых и бурых водорослей.
Считают, что высшие споровые растения появились около 415 — 430 млн лет назад. Существование растений на суше резко отличается от существования в водной среде. Вышедшие на сушу растения оказались в двух средах: воздушной и почвенной. В воздушной среде гораздо больше кислорода, растениям необходимо поддерживать стебель в вертикальном положении, поэтому сначала это были небольшие растения. Почвенная среда отличается иными условиями минерального питания и водоснабжения, и растениям необходимо было выработать специальные приспособления к жизни на суше, особенно для водоснабжения и транспорта питательных веществ, для защиты от высыхания и обеспечения полового процесса.
Первые примитивные высшие растения были дифференцированы на элементарные органы. Растения представляли собой дихотомически разветвленную ось, вверху заканчивающуюся органами спороношения, а снизу — корневищеподобными выростами — ризомоидами (прототипы корня) и ризоидами (прототипы корневых волосков). В результате длительного эволюционного процесса у спорофита высших растений появились ассимиляционные органы — листья, стебель, корень.
Листья, по-видимому, выполняли функцию фотосинтеза и спороносную функцию. В дальнейшем часть листьев несла ассимиляционную функцию, часть спороносную. Возможно, из спороносных листьев в процессе эволюции развились стробилы, или шишки, голосеменных и цветок покрытосеменных.
Споры у споровых растений образуются на диплоидном спорофите в многоклеточных спорангиях. Сначала это были одинаковые по размерам споры, а в дальнейшем разные: мелкие — микроспоры, крупные — мегаспоры.
Первые поселенцы суши — представители отдела риниофитов (псилофиты) вымерли в конце силурийского периода, более 415 млн лет назад. Виды риниофитов — влаголюбивые растения, произрастали на заболоченных почвах. Это были невысокие растения, до 50 см в высоту с диаметром стебля 5 мм. Надземные талломы заканчивались спорангиями, от корневищеобразных ризомоидов отходили ризоиды. Отдел включал один класс — риниевые и два порядка — риниевые и псилофитовые, которые отличались более высокой специализацией.
У представителей отдела зостерофилловых в отличие от риниевых спорангии имели боковое расположение. Это были небольшие дихотомически ветвящиеся растения, похожие на первичные плауновидные растения.
Общая характеристика. К отделу относят наиболее просто устроенные ныне существующие высшие растения. Тело представлено талломом или — у листостебельных форм — расчленено на стебель и листья. У листостебельных мхов ассимиляционная, механическая и проводящая ткани более или менее дифференцированы. Корни у мохообразных отсутствуют; к субстрату они прикрепляются с помощью ризоидов. Уникальность этой группы состоит в том, что у мохообразных в цикле развития гаметофит преобладает над спорофитом. Мохообразные — однодомные или двудомные растения. На разнополых гаметофитах образуются половые органы: антеридии и архегонии. В антеридиях созревают двужгути-ковые сперматозоиды, в архегониях — яйцеклетки. На женском гаметофите после оплодотворения формируется зигота, из которой развивается спорофит, представленный в виде коробочки со спорами. Оплодотворение возможно только во влажной среде. Развитие и питание спорофита — за счет женского гаметофита. Спорофит у мохообразных называется спорогоном и состоит из коробочки и ножки. Нижняя часть ножки расширена и называется гаусторией, ткань которой внедряется в гаметофит. Перед созреванием спор и открыванием коробочки внутри происходит редукционное деление. Высыпавшиеся гаплоидные споры дают начало новому гаплоидному растению.
Протонема — начальная стадия развития гаметофита — представлена в виде нити. Она делится на зеленую ассимиляционную часть - хлоронему и бесцветную подземную часть — ризонему. Эпидерма слоевищных и листостебельных мохообразных лишена кутикулы и типичных устьиц, в проводящей системе нет ситовидных трубок и трахеид.
Наука, изучающая мхи, носит название бриология. У мохообразных насчитывается примерно от 22 до 27 тыс. видов. В основном это низкорослые (от нескольких миллиметров до нескольких десятков сантиметров) многолетние растения влажных местообитаний.
Фотосинтез мохообразных менее интенсивен, чем у других высших споровых, у которых в цикле развития спорофит преобладает над гаметофитом; например, продуктивность фотосинтеза у мохообразных в 40 — 50 раз ниже, чем у цветковых растений (А. Г. Еленевский и др., 2000). Однако в связи с произрастанием мохообразных в условиях слабой освещенности под пологом других растений. У них выработалась способность фотосинтезировать при очень низкой освещенности, составляющей всего 4 % от полной. Фотосинтез у мохообразных длится (в условиях полярного дня) круглосуточно. Некоторые виды тундровых мхов способны фотосинтезировать даже под снегом на глубине до 20 см при температуре – 1400 С, т.е. круглогодично, что поддерживает обменные процессы и способствует небольшому зимнему приросту зеленой массы.
Мохообразные обладают очень древним примитивным типом водного режима, для которого характерна способность поглощать воду не столько физиологически, сколько физически: благодаря капиллярности, гигроскопичности, набуханию. Всасывается вода с помощью ризоидов, кроме того, мохообразные могут поглощать воду из атмосферы.
К примитивным признакам относят и способность мохообразных при неблагоприятных условиях переходить в состояние анабиоза, что подтверждает слабую активность обмена веществ в результате гаплоидности взрослого растения мохообразных. В условиях анабиоза обменные процессы снижаются и даже прекращаются и растения в таком состоянии могут находиться десятилетиями. К неблагоприятным условиям, а порой экстремальным, могут относиться не только недостаток влаги, но и высокая температура (до 70— 120°С), недостаток кислорода и бедные минеральными веществами почвы. Все многообразие частных специализаций мохообразных способствует их неизменному присутствию в разных растительных сообществах: влажных лугов, болот, лесов и прибрежно-водной растительности. Из природных зон мохообразные особенно богато представлены в тундре.
Значение мохообразных. Мохообразные способны аккумулировать многие, в том числе радиоактивные вещества, впитывать и удерживать большое количество воды, в связи с этим они играют большую роль в регулировании водного баланса местности. На сельскохозяйственных угодьях при неправильной агротехнике мхи могут интенсивно разрастаться и резко снижать продуктивность земель, способствуя их заболачиванию, ухудшать внешний вид газонов садов и парков. С другой стороны, разрастание мхов предохраняет почву от эрозии и обеспечивает равномерный перевод поверхностного стока вод в подземный.
Сфагновые мхи обладают антибиотическими, антисептическими свойствами и применяются в медицине. Так, во время Великой Отечественной войны дефицит ваты заменяли сфагновым мхом, который накладывали на раны. Кроме того, сфагновые мхи - образователи торфяников - источников топлива и органических удобрений.
Происхождение их относят на конец девона - начало карбона.
Согласно современной классификации, отдел Мохообразные подразделяется на три класса — Печеночники, или Печеночные мхи или Маршанциевые, Антоцеротовые и Листостебельные мхи. Листостебельные мхи включают три подкласса: сфагновые мхи, андреевые мхи и бриевые мхи. В основу классификации положены строение тела гаметофита, особенности строения ризоидов, строение и характер раскрывания коробочек, а также географическое распространение.
Класс Печеночники, или Печеночные мхи. Представитель класса — маршанция многообразная. Слоевище маршанции плоское, разветвленное в виде лопастей, размером до 10 см, плотно прилегающее к субстрату. Сверху слоевище покрыто однослойной эпидермой с устьицами. Фотосинтетическая ткань разделена на воздушные камеры перегородками. Снизу слоевище плотно прилегает к субстрату с помощью ризоидов. Слоевища раздельнополы, органы полового размножения расположены на особых подставках. На мужских гаметофитах антеридии находятся с верхней стороны подставки, а на женских гаметофитах архегонии расположены на нижней стороне подставки. После оплодотворения из образовавшейся зиготы развивается спорофит в виде коробочки на короткой ножке. Перед созреванием спор в коробочке происходит редукционное деление, споры в спорангиях разрыхляются гигроскопичными пружинистыми нитями — элатерами и выбрасываются наружу. Прорастающие споры дают начало гаплоидному гаметофиту в виде пластинчатой протонемы.
Печеночники способны и к вегетативному размножению частями гаметофита, чаще верхушками слоевища или краевыми выводковыми почками.
Маршанция живет во влажных местообитаниях: на сырой почве, на стенках канав, по берегам водоемов. Представитель свободноплавающих печеночников - риччия плавучая — хорошо известное и распространенное в аквариумах растение. Риччия служит убежищем для мальков рыб, нерестилищем для многих живородящих аквариумных рыбок. В естественных условиях риччия растет на Дальнем Востоке и в Предкавказье.
Класс Антоцеротовые включает около 300 видов, произрастающих в основном в тропиках.
Отличаются представители рода просто устроенным гаметофитом и сложно устроенным спорофитом. Растения произрастают на сырой почве. Антоцеросы однодомны, для предотвращения самоопыления антеридии созревают раньше архегониев. Антеридии и архегонии погружены в ткань слоевища. После оплодотворения вырастает щетинковидный спорогон до 10 см длиной. Спорогон растет своим основанием за счет вставочной меристемы. Основание спорогона бокальчатой формы. Спорофит имеет хлорофилл и ризоиды, поэтому женский гаметофит быстро разрушается и спорофит некоторое время существует самостоятельно. Вегетативное размножение происходит с помощью клубеньков, образующихся по краю или снизу слоевища. Как и лишайники, антоцеротовые мхи - пионеры влажных голых субстратов.
Класс Листостебельные мхи. Представители класса - наиболее широко распространенные по всему земному шару мхи. Класс включает более 14,5 тыс. видов из 700 родов, которые объединены в три подкласса.
Подкласс Сфагновые, Белые, или Торфяные мхи, представлен одним порядком - сфагновые мхи, одним семейством, одним родом, в котором более 300 видов. Сфагновые мхи наиболее широко представлены в умеренных и холодных областях Северного полушария. Наиболее типичный представитель торфяной мох, образующий сплошной покров на торфяных болотах и во влажных лесах. Его можно увидеть на зарастающих водоемах, в борах-беломошниках, он принимает участие в заболачивании лесов.
Растения имеют длинный (от 5 до 30 см) тонкий прямостоячий слабый стебель с пучковидно-олиствененными ветвями, на верхушке собранными в плотную головку.
Листья однослойные, состоят из 2 типов клеток: хлорофиллоносных и водоносных, бесцветных, выполняющих роль резервуаров без клеточного содержимого. Стенки водоносных клеток имеют кольчатые или спиральные утолщения, благодаря чему даже без воды эти клетки не спадаются. Торфяной мох способен притягивать и удерживать большое количество влаги. Сфагнум может всасывать в 37,5 раз больше воды, чем его сухая масса. Хлорофиллоносные клетки длинные и узкие, содержат хлоропласты. Эти живые клетки составляют основное “сетчатое” строение листа. Сфагновые мхи бывают как однодомными, так и двудомными, однако антеридии и архегонии образуются на разных побегах. После оплодотворения в присутствии воды из зиготы развивается бесполое поколение - спорофит в виде коробочки на ножке. После редукционного деления при созревании коробочки крышечка ее вскрывается, и споры выбрасываются наружу. Из спор прорастает гаметофит (протонема) - (половое поколение).
Торфяной мох сфагновых болот играет огромную роль в водном режиме ландшафтов. Сфагновые мхи преобладают в растительном покрове тундр и верховых болот умеренного и холодного поясов Северного полушария. В связи с избыточным увлажнением, недостатком кислорода, бактерицидной сильно кислой средой в глубине куртин сфагнового мха, его нижняя часть не разлагается. Происходит торфонакопление, образование залежей торфа, который широко используется в народном хозяйстве. Свойство наполнять водоносные клетки водой и надолго ее удерживать используют при выращивании комнатных растений влажных тропических лесов, особенно лиан, обертывая их мхом или направляя побеги к специально обернутым мхом стойкам или подвескам.
Подкласс Бриевые, или Зеленые мхи, наиболее распространенный подкласс всех листостебельных мхов. Зеленые мхи чаще всего многолетние растения от 1 мм до 60 см высотой зеленой буровато-красной и даже черной окраски (преобладают мхи с зеленой окраской). Зеленые мхи широко распространены на болотах, в хвойных лесах, на горах и в тундре (мниум точечный, фунария гигроскопическая, плевроциум шребера и др.). Обычно они растут на почве, на гнилой древесине, на коре, иногда даже на листьях деревьев. Среди них есть и водные обитатели - водный мох фонтиналис.
Представитель подкласса - широко распространенный кукушкин лен обыкновенный. Это многолетнее крупное растение до 50 см высоты. Произрастает группами на болотах, в заболачивающихся лесах, особенно хвойных, в долинах рек и т.д. Стебель растения прямостоячий, неразветвленный, густо покрыт многочисленными жесткими линейно-ланцетными листьями.
Мох кукушкин лен — растение двудомное. Раздельнополым гаметофитам для оплодотворения, как и другим мхам, необходима водная среда. После оплодотворения образуется зигота, из которой на женском гаметофите вырастает коробочка со спорами - спорофит. Спорофит сидит на ножке, питается первое время за счет гаметофита. Позднее коробочка с ножкой зеленеет, и спорогоний приобретает способность самостоятельно ассимилировать. Снаружи коробочка покрыта колпачком. После созревания коробочки со спорами внутри нее происходит редукционное деление, Крышечка открывается, и споры выбрасываются из коробочки на влажную почву. При благоприятных условиях спора прорастает и из нее развивается нитчатая протонема, которая дает начало новому гаметофиту, женскому или мужскому. Внешне молодые раздельнополые гаметофиты ничем не отличаются, но физиологически они разные. Гаметофит растет только во влажной среде. Спорофит более приспособлен к жизни в воздушной среде. Благодаря воздушной среде споры не прорастают внутри коробочки.
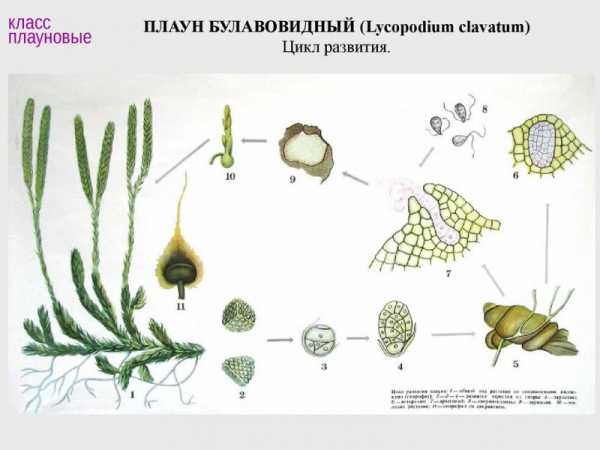
Плауновидные — самая древняя группа современных высших растений, ископаемые остатки растений рода плаун известны с конца силура — начала верхнего девона палеозойской эры (примерно 380 млн лет назад). В палеозое Плауновидные процветали, достигали огромных размеров (до 40 м), занимали господствующее положение в растительном покрове Земли. В настоящее время это угасающая группа растений. Все современные плауновидные — травянистые растения численностью примерно 1 тыс. видов. Это самые первые сосудистые растения: кроме хорошо развитых олиственных побегов у них есть настоящие корни. Для плауновидных характерна микрофилия, т.е. относительно мелкие размеры листьев. Листья плауновидных возникли как поверхностные выросты на осевых органах, что делает их уникальной группой. Стебель хорошо развит, листорасположение спиральное, супротивное и мутовчатое; ветвление дихотомическое. Молодые побеги нарастают за счет верхушечной меристемы, деятельность которой со временем угасает и поэтому Плауновидные ограничены в росте. Плауновидные не имеют главного корня. От подземного корневища отходят придаточные корни.
В жизненном цикле спорофит (взрослое растение) преобладает над гаметофитом. Спорангии у плауновидных формируются на специализированных спороносных побегах — стробилах в виде колосков. Гаметофит небольших размеров, называется заростком, который отмирает сразу же, как только из него образуется новый спорофит. Для оплодотворения яйцеклетки необходима водная среда.
Внутри отдела два класса: Плауновидные и Полушниковые.
Класс Плауновые объединяет равноспоровые растения. Современные плауновые представлены одним порядком, одним семейством Плауновые и Двумя родами: плаун и филлоглоссум.
У плауновых обоеполые подземные или полуподземные заростки (гаметофиты) созревают в течение 1 — 15 лет. У растений отсутствует камбий. Род плаун включает примерно 200 видов, как правило, это представители влажных тропических и субтропических лесов. В тропиках вертикальные стебли плауновых достигают 1,5 м высоты.
Типичный представитель зеленомошных хвойных лесов умеренного пояса Северного полушария — плаун булавовидный. Это вечнозеленое многолетнее травянистое растение с ползучим стеблем, достигающим 3 м длины. Стебель покрыт мелкими линейно-ланцетными листьями. Прикрепляется к земле боковыми корнями, внешне похожими на придаточные корни, живущими 2—5 лет. От ползучего стебля отходят невысокие вертикальные боковые побеги с дихотомическим ветвлением. На концах боковых побегов расположены спороносные колоски. Колосок состоит из споролистиков (спорофилл), в основании которых находится спорангий со спорами. Споры имеют трехгранную форму. Снаружи они покрыты сетчатым рисунком в результате утолщений стенок спор. При прорастании спор очень медленно (до 18 лет) развиваются обоеполые заростки. Заросток представляет собой беловатый клубенек диаметром до 2 см. На заростке формируются антеридии и архегоний. После оплодотворения из зиготы развивается молодой спорофит — бесполое поколение, на котором впоследствии образуются споры.
Споры плауна используют в медицине для обсыпки пилюль и в качестве детской присыпки. В большом количестве они представляют собой светло-желтый порошок, бархатистый, жирный на ощупь. Этот порошок предотвращает склеивание пилюль. Таким порошком раньше в металлургической промышленности обсыпали формы при выплавке чугуна.
Собирать колоски со спорами нужно очень осторожно, не повреждая самого растения, особенно корневую систему, так как растение восстанавливается очень медленно
Некоторые плауны ядовиты (баранец обыкновенный).
Класс Полушниковые включает разноспоровые травянистые растения. У некоторых видов в проводящей ткани есть сосуды. Из современно живущих полушниковых в классе два порядка: Селагинелловые и Полушниковые.
В порядке Селагинелловые одно семейство Селагинелловые с одним родом селагинелла. Большинство видов мелких размеров — 5—15 см, с нежными стелющимися побегами, произрастают под пологом влажных лесов, образуя зеленые подушки, похожие на моховые. Встречаются формы с лазающим и вьющимся побегами, достигающими в длину 20 м. Растения открытых сухих местообитаний имеют прямостоячий стебель от 20 см до 3 м в длину. У видов со стелющимся стеблем листья расположены в 4 ряда: более крупные листья в двух нижних рядах, а мелкие в двух верхних. Прикрепляются к почве с помощью тонких дихотомически разветвленнных корней, которые образуются на особых органах — ризофорах (корненосцах). У прямостоячих селагинелл листья мелкие (0,5 — 5 мм длины), спирально расположенные по стеблю. Мозаика листьев позволяет лучше улавливать свет. Некоторые селагинеллы — эпифитные растения.
Селагинеллы — разноспоровые травянистые растения. При бесполом размножении микро- и макроспоры возникают в микроспорангиях на микро- и мегаспорофиллах, в большинстве случаев расположенных на одном стробиле в форме колоска. Колосок может занимать различное положение. Прорастание микро- и макроспор начинается внутри спорангиев. При прорастании микроспоры образуется сильно редуцированный заросток. При первом делении появляются две неравноценные клетки: маленькая — проталлиальная (остаток вегетативного тела заростка), большая — антеридиальная. Антеридиальная клетка дает начало антеридию, где формируются многочисленные двужгутиковые сперматозоиды. К моменту созревания клетки стенки антеридия и проталлиальной клетки расплываются, и сперматозоиды плавают в общей массе цитоплазмы. Это и будет мужской гаметофит, похожий на пылинку семенных растений. Из мегаспор развиваются женские заростки. Ткань заростка многоклеточная, и после разрыва оболочки мегаспоры заросток выпячивается наружу, его клетки зеленеют и на нем образуются ризоиды. У некоторых видов этот заросток может выпасть наружу, попасть при благоприятных условиях на почву и прикрепиться к ней с помощью ризоидов. Ризоиды выполняют функцию поглощения воды, необходимой для оплодотворения. В данном случае женский заросток питается самостоятельно. У других видов женский гаметофит остается на мегаспоре, и тогда ризоиды выполняют функцию улавливания пылинок; в этом случае женский гаметофит живет за счет материнского растения. В обоих случаях после оплодотворения яйцеклетки развивается зародыш, который состоит из стебелька, листочков и ризофора, на котором впоследствии образуются корни. Сформировавшийся зародыш переходит к самостоятельной жизни.
В порядок Полушниковые — входит одно семейство Полушниковые с единственным родом полушник, представленным примерно 70 видами. Это многолетние травянистые растения с утолщенным прямостоячим стеблем и розеткой линейных шиловидных цилиндрических листьев. У основания стебля находится ризофор, от которого отходят корни. На верхней поверхности в основании листьев расположены многочисленные микро- и мегаспорангии. Гаметофит у полушни-ковых еще более редуцирован, чем у селагинелловых.
Виды полушниковых встречаются в чистых олиготрофных озерах и по опресненным мелководьям морей, поэтому могут относиться к индикаторам чистой воды.
Псилотовидные представлены одним классом, порядком, семейством, в которое входят два рода: псилот с двумя видами и тмезиптерис с десятью видами. Псилотовидные - представители влажных тропических и некоторых субтропических областей земного шара.
Псилотовидные - травянистые многолетние растения, просто устроены и напоминают риниофиты. Псилот имеет дихотомически ветвящиеся побеги с плоским или трехгранным стеблем от 20 см до 1 м высотой, к почве прикрепляется дихотомически разветвленными ризоидами, что говорит об их древнем происхождении. Кроме того, растения не имеют типичных листьев. Псилофитовые ведут наземный или эпифитный образ жизни. Наземные растения имеют прямостоячий стебель, а у эпифитов стебель свисает вниз.
Псилот — равноспоровое растение. Споры формируются в спорангиях на концах боковых ветвей. Из споры вырастает подземный довольно крупный обоеполый зеленый гаметофит; половые органы рассеяны на его поверхности. Оплодотворение многожгутиковыми сперматозоидами в присутствии воды.
Тмезиптерис отличается от псилота более крупными листовыми придатками.
Отдел Хвощевидные.
Ныне живущие хвощевидные, или членистые, представлены многолетними корневищными травами с мутовками бурых редуцированных бесцветных листьев. В прошлом, будучи гигантских размеров, вместе с древовидными папоротниками и лепидодендронами они составляли каменноугольные леса. В конце палеозоя в связи с изменением климата древесные формы вымерли, и в настоящее время в этой группе растений остался только один класс Хвощевые, один порядок, одно семейство с одним родом хвощ, включающим 25 видов. Проводящая система представлена различными трахеидами и иногда сосудами. Во флоэме есть паренхима и ситовидные элементы. Листья чешуевидные, бурые или желтоватые. Функцию фотосинтеза выполняет зеленый стебель, на верхушке которого вырастает спороносный колосок. Стебли у хвощей умеренных широт не превышают 1 м, у представителей тропиков они достигают 10—12 м при толщине стебля 6 — 8 см.
В жизненном цикле спорофит преобладает над гаметофитом. Хвощ - равноспоровое растение. Для репродуктивных органов бесполого размножения характерны стробилы в виде колосков на верхушке стеблей. К оси стробил мутовками прикреплены спорангиофоры, которые представляют собой шестиугольные щитки. На внутренней стороне щитков находятся спорангии. При созревании спор щитки подсыхают, разрываются и споры разлетаются. Созревшая спора одета двумя оболочками, а снаружи спирально обернута своеобразной лентой — элатерой, способной отгибаться и совершать гигроскопические движения. При подсыхании элатеры раскручиваются и сцепляются друг с другом, образуя своеобразные клубочки спор. Это способствует распространению спор и прорастанию заростков недалеко друг от друга. Заростки обоеполые, представляют собой пластинку, прикрепленную к почве ризоидами. Наверху пластинки формируются антеридии и архегонии. Многие виды хвощей проявляют физиологическую разноспоровость, иногда в связи с условиями произрастания, иногда она генетически заложена. После оплодотворения яйцеклетки многожгутиковым сперматозоидом в условиях капельно-водной среды из зиготы формируется зародыш, который имеет все зачаточные вегетативные органы. Корешком зародыш внедряется в почву и начинает самостоятельное существование.
Хвощи — сильно разветвленные корневищные растения, в узлах которых иногда развиваются богатые крахмалом клубеньки. От узлов корневища отходят многочисленные придаточные корни.
Хвощи — представители разных растительных сообществ земного шара, но произрастающие в местах достаточного или избыточного увлажнения. Самыми распространенными видами для средней полосы европейской части России являются хвощ полевой, хвощ лесной, хвощ болотный и др. Хвощ полевой — особо злостный сорняк, однако молодые побеги его используют в медицине как мочегонное средство. Отмершие остатки древних каменноугольных гигантов хвощевидных дали начало залежам каменного угля.
Папоротникообразные — древняя группа высших споровых растений, геологический возраст которых такой же, как и у хвощевидных. Ископаемые формы известны с девона. Расцвет гигантских древовидных папоротников был в карбоне, остатки которых образовали залежи каменного угля. Предки папоротникообразных точно не установлены, предполагают, что это были риниофиты. В настоящее время известно более 10 тыс. видов.
Папоротникообразные в отличие от хвощевидных имеют крупные листья — вайи, в большинстве многократно рассеченные, перистые; произошли листья в результате уплощения крупных ветвей. Листья длительное время обладают верхушечным ростом, имеют черешок и пластинку. Пластинка листа прикреплена к оси, или рахису, который представляет продолжение черешка и соответствует главной жилке цельного листа. Размеры листьев от 1 — 2 мм до 10 м в длину. Стебель большинства папоротников короткий, горизонтально расположен в виде корневища, от нижней стороны его отходят придаточные корни. Камбий у папоротников отсутствует, у них нет вторичной древесины, прочность древовидных форм обусловлена склеренхимной обкладкой вокруг проводящих пучков стебля. Склеренхима присутствует и в корнях папоротников.
Папоротникообразные произрастают по всему земному шару, как правило, во влажных местообитаниях. В умеренном климатическом поясе это наземные многолетние (за исключением сальвинии плавающей) травянистые растения. Преобладающее большинство папоротникообразных встречается во влажных тропических областях, где встречаются папоротники разнообразной жизненной формы. В горных районах тропиков произрастают древовидные папоротники, в зарослях влажных тропических лесов — папоротники-лианы. В водоемах тропических областей живут плавающие многолетние папоротники. Некоторые древовидные виды папоротников достигают в высоту 20 —25 м при толщине ствола 50 см. У древовидных видов папоротников стебель представлен прямостоячим стволом, несущим на верхушке крону листьев. Во влажных тропических лесах широко распространены виды рода платицериум, ведущие эпифитный образ жизни.
В жизненном цикле преобладает спорофит, представляющий собой взрослое многолетнее растение папоротникообразных. Рассмотрим жизненный цикл на примере мужского папоротника. Спорангии развиваются на нижней стороне зеленых листьев на специальных спороносцах — сорусах или на специализированных листьях. Например, у страусника листья дифференцированы на фотосинтезирующие и фертильные, несущие спорангии. У представителей порядка Ужовниковые часть листа выполняет вегетативную функцию, а другая часть спороносную, или фертильную. Сорусы могут располагаться одиночно или группами. Место прикрепления спорангия к листу называется плацентой. У многих папоротников сорусы состоят из выпуклого ложа — рецептакула, к которому с помощью ножек прикрепляются спорангии. Снаружи спорангии защищены специальными покрывальцами, или индузиями, сформированными в результате местного разрастания плаценты или разрастания поверхностных тканей листа. При подсыхании спорангия он разрывается в местах тонкостенных клеток. Споры высыпаются, и из них развивается гаметофит в виде заростка. Большинство папоротников — равноспоровые растения. Гаметофиты их обоеполые, зеленые, размером с пятирублевую монету, сердцевидной формы, обитают на поверхности почвы. У некоторых папоротников гаметофиты лишены хлорофилла и живут под землей. Прикрепляются к субстрату с помощью ризоидов. На нижней, брюшной стороне гаметофита развиваются архегонии и антеридии. Антеридии находятся у основания пластинки заростка и созревают раньше. Чуть позднее на вершине пластинки развиваются архегонии. Такая неравномерность развития способствует перекрестному оплодотворению. Из оплодотворенной яйцеклетки образуется зигота, которая дает начало дигогоидному зародышу, из которого формируется диплоидный спорофит.
У разноспоровых папоротников гаметофит, особенно мужской, редуцирован до микроскопических размеров.
Папоротники размножаются также вегетативно: с помощью выводковых почек, образующихся на листьях, стеблях и корнях.
Отдел Папоротникообразные делят на 7 классов, из которых до наших дней дожили только представители трех классов: Ужовниковые, Мараттиевые и Полиподиевые.
Класс Полиподиевые включает три подкласса: Полиподииды, Марсилеиды, Сальвинииды.
Из подкласса Полиподииды в лесах умеренных широт Европы, Азии и Америки широко распространен папоротник щитовник мужской. Он относится к порядку Полиподиевые, или Настоящие папоротники, семейству Асплениевые, роду щитовник. Растение представляет собой розетку дваждыперисторассеченных листьев, отходящих от толстого ползшего корневища. Молодые листья на концах свернуты в “улитку”, растут верхушкой, как и стебель. От корневища отходят придаточные корни.
Стебель снаружи одет эпидермисом, дальше расположена кора. Вдоль стебля тянутся крупные проводящие пучки, более мелкие проводящие пучки проходят в листья. Вся система пучков, или стела, похожа на сетку с крупными ячеями, из которых каждая представляет собой прорыв пучка в местах отхождения от него крупного листового следа. Такая проводящая система называется сетчатой, или диктиостелой (диктион — сетка). Основания корневищ молодых папоротников имеют проводящие пучки простого строения, или протостелу. Это одно из доказательств того, что диктиостела произошла от протостелы, свойственной более примитивным папоротникообразным и псилофитовым.
Спорангии, имеющие вид двояковыпуклой линзы, сидят на ножках на нижней стороне листа. Спорангии собраны в сорусы, одетые индузием. При созревании спор и подсыхании тонких стенок индузия стенка спорангия прорывается и споры разбрасываются в разные стороны на расстояние до 1 м.
После прорастания споры образуется заросток сердцевидной формы диаметром около 1 см. Заросток прикрепляется к почве с помощью ризоидов, на нем развиваются половые органы: антеридии и архегонии. Оплодотворение происходит в водной среде. Из оплодотворенной яйцеклетки развивается зародыш, который через некоторое время (образование стебелька с листочком и корешка) переходит к самостоятельной жизни спорофита. В хвойных (особенно сосновых) и лиственных лесах, на горах, вырубках и других сухих и бедных почвах широко распространен орляк с крупными длинночерешковыми листьями. Пластинка листа широкотройчаторассеченная, напоминающая со стороны треугольник. В умеренных областях Северного полушария произрастает “кружевной” на вид с сильно рассеченной нежной листовой пластинкой женский кочедыжник, редкий и очень изящный папоротник линнея и величественный, похожий на страусово перо страусник и др.
Подкласс Марсилиевые включает всего около 70 видов разноспоровых водных или прибрежно-водных папоротников, встречающихся в основном в тропических областях земного шара. Несколько видов можно встретить в низовьях р. Волги. В умеренных широтах некоторые виды культивируют в водоемах ботанических садов или любители-аквариумисты.
Подкласс Сальвиниевые состоит из семейства Азолловые и семейства Сальвиниевые. Род сальвиния включает 8 видов пресноводных, большей частью тропических растений. В Сибири, на Дальнем Востоке, на Кавказе и в Средней Азии встречаются 2 вида сальвинии. Сальвиния плавающая — однолетний водный папоротник небольших размеров (15 — 20 см) со стелющимися по воде побегами. В каждой мутовке побега из трех листьев два листа плавающие, один подводный; корней нет. Плавающие листья цельные, покрыты восковым налетом; нижний лист сильно рассеченный, с множеством нитевидных долей, покрытых жесткими волосками, внешне похож на корни. Шаровидные сорусы формируются на коротких черешках подводных листьев. Сальвиния — разноспоровый папоротник. Вегетативное размножение осуществляется за счет отделения боковых побегов, образующихся из почек между листьями. В благоприятных условиях произрастания сальвиния сильно разрастается и мешает хозяйственной деятельности. В умеренных широтах сальвиния часто используется в аквариумоводстве.
Семейство Азолловые с единственным родом азолла включает 6 тропических видов. Особенность азоллы — ее симбиоз с цианобактерией анабеной азоллы, благодаря чему папоротник способен усваивать атмосферный азот, а поэтому его используют в качестве зеленого удобрения, особенно в странах Юго-Восточной Азии.
Семенные растения
Семенные растения существенно отличаются от споровых растений. Общее направление эволюции шло по линии дальнейшего развития спорофита и редукции гаметофита. Семенные растения размножаются семенами, а не спорами. Половой процесс не связан с капельно-жидкой средой, гаметофиты развиваются и проходят полный цикл своего развития на спорофите. В борьбе за существование на суше появление семени имело большое эволюционное значение, так как зародыш находится под покровом оболочек семени и семя в отличие от споры содержит питательные вещества для развития зародыша. Все семенные растения разноспоровые. Микроспоры дают начало мужскому гаметофиту, мегаспоры — женскому. Мегаспоры развиваются в семяпочках, или семязачатках, которые представляют собой мегаспорангии. Семяпочка состоит из нуцеллуса и интегумента, или покрова (одного или двух). Мегаспора постоянно заключена в мегаспорангии (нуцеллус гомологичен мегаспорангию). В мегаспоре развивается женский гаметофит, происходит процесс оплодотворения и формируется зародыш. Со временем семязачаток превращается в семя, внутри которого развивается зародыш. Зародыш семени имеет зародышевый корешок, зародышевые листочки (семядоли) и почечку. Семенные растения представлены двумя отделами: Голосеменные и Покрытосеменные.
studizba.com