
Сельское хозяйство будущего: как выращивать овощи без Солнца. Что делают растения в темноте
Влияние различных сочетаний света и темноты на цветение растений
Современный южный ареал капусты абиссинской — Эфиопия, несмотря на это, она имеет четкую никтофобную реакцию, свидетельствующую о консервативности актиноритмической реакции растений и о северном происхождении данного вида. Следует отметить, что в условиях коротких дней Эфиопии она развивается медленно.
При непрерывном искусственном освещении период ее развития во много раз сокращается. Это подтверждают данные о выращивании капусты абиссинской при интенсивном (от 600 до 900 Вт/м2) ксеноновом освещении.
При непрерывном освещении ксеноновыми лампами бутонизация капусты абиссинской наблюдается на 17-й день после посева, а при длине дня 13 ч, близкой к продолжительности дня в Эфиопии, бутоны появляются на 85-й день.
Сильное тормозящее действие темноты наблюдалось начиная с 18-часового дня. При этом чем длинее был период темноты, тем дольше задерживалось цветение растений. Одновременно возрастало общее число часов света от начала опыта до появления первых бутонов: от 408 в условиях непрерывного освещения до 1105 при 13-часовом дне.
Как показывают результаты указанного опыта, капуста абиссинская способна зацветать, хотя и с большим опозданием, в условиях достаточно коротких дней. Даже 11 ч темноты не могут быть полным препятствием для цветения данной культуры. В этой связи становится понятным, что капуста абиссинская хоть и является никтофобным растением, но в конце концов она переходит к цветению и в условиях коротких дней, присущих Эфиопии.
Увеличение общего количества часов света, необходимое для формирования бутонов по мере возрастания темного суточного периода, свидетельствует о том, что темнота — это не нейтральный фактор, который просто отодвигает бутонообразбвание, напротив, она приводит к активному торможению процессов репродуктивного развития.
Мы изучали роль световых и темновых процессов в актиноритмической реакции капусты абиссинской. Для этого исследовали чередования света и темноты как суточные, так и несуточные, не препятствующие цветению и сильно тормозящие развитие растений. Основными источниками освещения в большинстве наших исследований были лампы накаливания, лучистый поток которых профильтровывался через воду для поглощения части длинноволновой инфракрасной радиации. Мощность на уровне вершин растений составляла 450 Вт/м2. В спектре ламп накаливания, как и ксеноновых, имеется большая доля красной и инфракрасной радиации. В этом отношении указанные спектры близки к солнечному. В этих условиях критическим для прохождения репродуктивного процесса капусты абиссинской является уже 17-часовой день. Более того, с уменьшением длины дня до 16 ч зацветают, как правило, не все растения и сроки бутонообразования затягиваются до 30 сут.
Результаты опытов, где в качестве источников света были использованы лампы накаливания, также свидетельствуют о тормозящем действии темноты на процессы репродуктивного развития капусты абиссинской.
Принимая во внимание отрицательную роль темноты в процессах репродуктивного развития никтофобных растений, следовало выявить, какие периоды ее, перемежающиеся с равновеликими периодами света, оказывают наименьшее тормозящее действие. Для этого была поставлена серия опытов, в которых чередование равновеликих периодов света и темноты проводилось по схеме.
В опытах использовали растения, предварительно выращенные при 12-часовом дне до 30-дневного возраста. Продолжительность опытов — 70 дней. При чередовании света и темноты через 30 мин бутоны образовывались на 13-й день опыта, т. е. всего на 2 дня позже, чем при непрерывном освещении. Таким образом, периоды света в 30 мин, чередующиеся с такими же периодами темноты, оказались значительно более благоприятными, чем периоды света и темноты длительностью 1 ч. Следовательно, за 30 мин и, конечно, за более короткое время отрицательное действие темноты не успевает проявиться.
С увеличением равновеликих интервалов света и темноты до 1, 2, 3, 4 и 6 ч бутоны у капусты абиссинской образовывались примерно через 30—32 дня от начала воздействия. При дальнейшем увеличении светлых и темных периодов до 18 ч процесс репродукции тормозился и растения до конца опыта оставались в вегетативном состоянии.
Бутонообразование наблюдалось у растений, находившихся в режимах: 19 ч света и 19 ч темноты, 24 ч света и 24 ч темноты, 32 ч света и 32 ч темноты, 48 ч света и 48 ч темноты, 72 ч света и 72 ч темноты. Чередования равновеликих периодов света и темноты длительностью в 32, 48 и 72 ч оказались более благоприятными, чем чередование 19 ч света и 19 ч темноты и 24 ч света с 24 ч темноты.
Сравнивая результаты, можно предположить, что при равновеликих интервалах света и темноты, начиная от 1 до 6 ч, темнота не устраняет бутонообразования, хотя и задерживает его на различные сроки. В этих случаях процессы онтогенеза, проходящие на свету, не снимаются последующими периодами темноты.
При чередовании периодов света и темноты с интервалами в 9, 10, 11, 12, 13, 15, 16 и 18 ч длительность темных периодов подавляет световые процессы и растения остаются вегетативными. За 24 ч световые процессы, связанные с репродуктивным развитием капусты абиссинской, продвигаются дальше, чем при чередовании 18 ч света и 18 ч темноты, и поэтому в последующие 24 ч темнота не снимает светового эффекта.
В сущности, аналогичная картина наблюдается и в вариантах, где свет и темнота чередуются через 32, 48 и 72 ч. Однако за 32 ч световые процессы продвигаются дальше, чем за 24 ч, а последующие 32 ч темноты менее тормозят световой эффект, чем в вариантах с 48 и 72 ч. При 72 ч света и 72 ч темноты световые процессы проходят также еще не до конца, а 72 ч темноты несколько задерживают появление бутонов.
Ранее было установлено, что после светлого периода в 5 сут, полностью обеспечивающего нормальное развитие бутонов, последующие 5 сут темноты нисколько не задерживали этого процесса по сравнению с ним при непрерывном освещении.
Следовательно, темнота, данная после завершения определенных световых процессов морфогенеза, не препятствует его выявлению, но в то же время тормозит его при незавершенности указанных процессов.
Проведенные эксперименты позволили выявить особенности актиноритмической реакции капусты абиссинской при чередовании равновеликих периодов света и темноты разной длительности. Прежде всего следует отметить, что краткие периоды темноты практически не влияют на скорость онтогенеза даже при чередовании с такими же интервалами света.
Сочетание различных отрезков света и темноты позволяет определить динамику световых процессов, связанных с бутонообразованием, в различных несуточных актиноритмических циклах. В частности, можно утверждать, что за 18 ч (режим — 18 ч света и 18 ч темноты) световые процессы, связанные с прохождением индукционного периода, не завершаются, и физиологическое состояние этих растений значительно отличается от состояния другой группы растений, получивших 19 ч света перед наступлением темноты (режим 19 ч света и 19 ч темноты).
При чередовании равновеликих отрезков света и темноты критическим периодом темноты является 9-часовой интервал. Именно поэтому капуста абиссинская не цветет за 3-месячный срок в актиноритмических условиях 15-часового и более короткого дня.
Что касается практического аспекта наших исследований, то пока можно сказать, что чередования света и темноты в пределах от 5 до 30 мин определяют наименьшие затраты электроэнергии. Если при непрерывном освещении, когда бутоны появляются на 10-й день после начала воздействия, электроэнергию, затраченную за этот период, принять за 100%, то при кратковременных чередованиях эта же величина сокращается более чем на 1/3 и составляет 59%. Данное явление интересно не только с практической точки зрения, поскольку указывает на энергетические изменения физиологических процессов, связанных с формированием бутонов.
Еще большее практическое значение имеют результаты опыта, в котором растения подвергались воздействию неравновеликих чередований света и темноты при длительности циклов от 8 до 16 мин. При чередовании 4 мин света с 8 мин темноты растения переходят к репродукции практически в нормальные сроки. В этом случае продолжительность светлого периода составляет только 1/2 периода темноты, что характерно для обычного суточного режима, в котором 8-часовой световой день чередуется с 16 ч темноты. В последнем случае капуста абиссинская не переходит к цветению. Таким образом, имеется реальная возможность выращивать растения, подобные капусте абиссинской, затрачивая на ее освещение электроэнергию всего в течение 8 ч в сутки.
Широкое распространение никтофобных видов в различных географических районах связано с их способностью развиваться в условиях, где ночь достигает 12—14 ч в сутки, хотя данный процесс и происходит значительно медленнее, чем при длинном дне. Таково поведение капусты абиссинской в Эфиопии. Если бы никтофобные виды не обладали этой способностью, они могли бы сохраниться только в умеренных широтах. Вместе с тем, несмотря на многолетнее пребывание в южных широтах, такие виды несомненно северного происхождения не теряют своей никтофобности.
Еще одно никтофобное растение — шпинат Виктория, цветет одновременно на 23—29-е сутки после появления всходов при длине дня от 24 до 15 ч. При 14-часовом дне цветение шпината уже задерживается на целый месяц, а в условиях более коротких дней, начиная с 13-часового, данная культура. в наших опытах не цвела. Очевидно, темнота не только не способствует цветению шпината, но, наоборот, при более 9 ч в сутки она делает его невозможным.
Тем интереснее сообщение Гентчева и Густафсона (Gentcheff, Gustafson, 1940) о том, что в их опыте шпинат (сорт Герта) цвел при непрерывной темноте на 21-е сутки.
Следовательно, не темнота сама по себе делает невозможным образование шпинатом органов плодоношения, а лишь определенные ее сочетания со светом.
Какими же должны быть суточные циклы свет — темнота, обеспечивающие актиноритмическую реакцию никтофильных по цветению растений? На этот вопрос отвечают результаты наших опытов.
Периллу масличную выращивали при 24-часовом цикле, начиная с непрерывного освещения и кончая 3-часовым днем. Оказалось, что данное растение начинает цвести с 15-часового дня и кончает при 4-часовом дне.
Иными словами, для цветения периллы масличной необходимо в сутки не менее 9 ч темноты и не менее 4 ч света. Если это так, то, очевидно, сочетание 4 ч света с 9 ч темноты (13-часовой цикл) должно обеспечить семенное размножение данной культуры. И первый же эксперимент, в котором периллу масличную выращивали при искусственных суточных циклах в 13 ч, из которых на долю света приходилось 4 ч, а на долю темноты — 9 ч, показал правильность нашего предположения. У растений периллы в этом опыте, как и во многих других, освещение было искусственным (250-ваттные дуговые ртутно-люминесцентные лампы ДРЛ), а включение и выключение света автоматическим.
Бутоны появлялись здесь несколько позднее (на 2 недели), чем при оптимальных актиноритмических условиях. Это характерно для всех случаев, когда света было значительно меньше, чем темноты. Аналогичные результаты получаются и в том случае, когда растения периллы масличной выращиваются при нормальных 24-часовых актиноритмических циклах с тем же отношением 1:2, что составляет 8 ч света и 16 ч темноты. В данном случае бутоны появлялись также на 32—34-й день.
Чтобы выявить наиболее оптимальные сочетания света и темноты, способствующие скорейшему переходу периллы масличной к цветению, мы провели большое число опытов с разными суточными циклами. Прежде всего было выяснено влияние на онтогенез периллы различных по продолжительности светлых периодов, чередовавшихся всегда с периодом темноты, равным 9 ч.
При продолжительности светлого периода от 4 до 7 ч бутонизация задерживалась, зато доведение светлого периода до 8 ч обеспечивало появление бутонов в кратчайший срок (20 дней).
Образование бутонов через 20 сут после начала опыта для молодых растений периллы, имеющих тот же возраст, — это нормальное явление для самых лучших световых условий. Дальнейшее наращивание светлого периода актиноритмических циклов, вплоть до 13—14 ч, не мешало нормальному онтогенезу периллы, и во всех указанных вариантах бутонизация наступала через 20— 22 сут после начала опыта. Но как только длина светлого периода доходила до 15 ч (в 24-часовом цикле), снова наблюдалась значительная задержка в появлении бутонов (на 16 дней против нормы). В двух следующих вариантах опыта эта задержка продолжала нарастать.
Таким образом, 9-часовой период темноты вызывает быстрое развитие периллы масличной только после светлого периода, длящегося от 8 до 14 ч.
Результаты нашего другого опыта с указанной культурой, где она получала 7-часовой период темноты в различных циклах, начиная с 12-часового и кончая 24-часовым. В этом случае все растения периллы остались до конца опыта, длившегося 60 сут, в вегетативном состоянии. Отсюда неизбежен вывод, что 7-часовой период темноты в цикле недостаточен для нормального онтогенеза типичного никтофильного растения — периллы масличной.
Если перилла масличная получает 8 ч темноты в нормальном 24-часовом цикле, то она тоже не приступает к бутонизации.
При любой продолжительности светлого периода — от 5 до 8 ч — бутонообразование периллы наступало, хотя и значительно позже, чем в оптимальных условиях. Как только период света становился длиннее на 1 ч 8-часового периода темноты, цветения периллы не происходило. Значит, сокращение ночного 9-часового периода на 1 ч приводит или к сильной задержке в репродуктивном развитии периллы, или вообще делает его невозможным. Вероятно, 8 ч являются критической продолжительностью темного периода цикла, обеспечивающего нормальный онтогенез периллы.
Любопытно, что при 8-часовой ночи появление бутонов наблюдается тем скорее, чем короче светлый период. Не служат ли эти факты прямым указанием на зависимость идущих в темноте процессов от количества лучистой энергии, связанной растениями за светлый период суток? Может быть, в процессе длительной эволюции у периллы, да и у других типичных никтофильных растений, возник суточный цикл метаболизма, в котором фото- и никтореакции находятся в оптимальных соотношениях, если периоды света и темноты равны друг другу.
Чтобы проверить возможное значение указанных равенств различных периодов света и темноты, проводились опыты с периллой масличной в условиях искусственного освещения с автоматическим включением и выключением ламп ДРЛ. Было испытано 15 различных циклов, начиная с 8-часового и кончая 52-часовым, с равными периодами света и темноты.
Актиноритмические циклы 4:4, 5:5, 6:6, 7:7, данные перилле в течение 80—90 календарных суток, не обеспечили репродуктивного развития растений, и все они оставались в вегетативном состоянии. В условиях 16-часового цикла с равными периодами света и темноты бутоны появились очень поздно, на 55-й календарный день после начала опыта. Зато в последующих циклах 9:9, 10:10, 11:11, 12:12 бутонообразование наблюдалось в нормальные сроки. В цикле 13: 13 бутоны появились на 5—6 сут позже, а при цикле 14:14 опоздание еще увеличивалось и дошло до 10 сут. Наконец, во всех следующих циклах, начиная с 32-часового и кончая 52-часовым, растения периллы за все время опыта оставались в вегетативном состоянии.
Далеко не все циклы, содержащие равное число часов света ц темноты, вызывают переход растений периллы масличной от вегетативного развития к репродуктивному. Обычно при выращивании периллы в условиях 24-часового цикла она дает наибольшие урожаи сухой массы и семян при равных (12-часовых) периодах света и темноты. Однако в условиях 20- и 18-часовых циклов общий урожай периллы оказывается более высоким. Причем в иных случаях прибавка сухой массы и массы семян доходила до 20— 30%. Это может иметь существенное практическое значение для культуры растений в условиях искусственного освещения, ибо в таких случаях несомненно повышается коэффициент использования растениями и лучистого потока, и электрической энергии. Весьма любопытным оказалось поведение растений периллы масличной при выращивании ее в двух вариантах 16-часового цикла. В одном случае, когда на долю света и темноты приходилось по 8 ч, бутоны образовались только на 55-й календарный день, т. е. более чем с месячным опозданием против нормы. В другом случае, когда на светлый период цикла приходилось 7 ч, а остальные 9 ч растения находились в темноте, бутоны появились почти в нормальный срок — на 25-й календарный день, или на 3—5 дней позднее, чем при оптимальных условиях. Из-за такого значительного различия наступления бутонизации в 50-дневном возрасте растения этих, казалось бы, близких по условиям освещения групп 16-часового цикла резко отличались друг от друга.
Растения, получавшие по 8 ч света и темноты, находились в вегетативном состоянии, а растения, получавшие 7 ч света, а затем 9 ч темноты, были покрыты почти созревшими семенами. Онтогенез этих растений периллы, выросших при 16-часовом актиноритмическом цикле, зависел от 1 ч, отнятого от светлого периода и добавленного к темному. Очевидно, 16-часовой цикл, как и многие другие циклы, вполне пригоден для быстрого репродуктивного развития периллы при условии, что светлый и темный периоды находятся в определенном соотношении друг с другом. Если в любом цикле период темноты меньше, чем 9 ч, то перилла, как правило, или совсем не цветет, или зацветает очень поздно.
Естественно, возникает предположение, что критическая граница между активными и пассивными для онтогенеза актиноритмами лежит в пределах значительно меньших, чем часовые интервалы времени. Поэтому с помощью специального автомата, включавшего и выключавшего свет в строго заданное время, были проведены эксперименты, в которых выяснялось значение 5-минутных интервалов в пределах часа, определяющего судьбу периллы масличной в 16-часовом цикле.
Данные составлены на основании описания всех опытных растений периллы масличной в возрасте 50 календарных суток. В это время все растения пяти первых групп, для которых была создана продолжительность ночи от 8 ч до 8 ч 20 мин, находились в вегетативном состоянии. Растения следующих трех групп (6, 7, 8-й), развивавшихся при длине темного периода от 8 ч 25 мин до 8 ч 35 мин, имели крупные бутоны. Растения же остальных пяти групп (9, 10, 11, 12, 13-й), росшие при длине темного периода от 8 ч 40 мин до 9 ч, были покрыты готовыми к уборке семенами. В пределах часа, лежащего на границе активного и неактивного 16-часовых актиноритмов, обнаружилось четкое значение 5-минутных интервалов, которые в зависимости от того, приходятся ли они на светлую или темную часть 16-часового цикла, определяют характер онтогенеза растений периллы масличной.
Теперь можно с большей точностью, чем прежде, сказать, что минимальным периодом темноты в актиноритмических циклах, вызывающих быстрое зацветание периллы, является не 9 ч, а 8 ч 40 мин. Если длительность темного периода уменьшается на 5 мин, то развитие задерживается, а если период темноты равен 8 ч 20 мин, то растения периллы или совсем не цветут, или дают бутоны лишь на 55-й день после начала жизни.
Думается, что в данном случае положительный результат зависит именно от длительности темного периода цикла, а не от строго определенных сочетаний в цикле света и темноты. Во всяком случае, только что изложенные результаты говорят об очень тонких и четких границах никтопериодов, определяющих никтопериодическую реакцию растений.
Результаты изложенных выше исследований, а также и хорошо согласующиеся с ними литературные данные приводят нас к неизбежному выводу о решающей роли темных периодов актиноритмических циклов в так называемой фотопериодической реакции растений. Несомненно, что все основные физиологические процессы, определяющие характер реагирования растений на суточные циклы смены света темнотой, проходят в темноте, а не на свету.
Среди никтофильных по цветению растений, как считают некоторые авторы, имеются такие, для образование цветочных зачатков которых нужен всего один темный период.
Так, японские исследователи Имамура и Такимото (Ifmamura, Takimoto, 1956) установили, что цветочный зачаток в виде чешуйки у ипомеи голубой (Pharbitis nil) появляется в результате воздействия на нее единственным периодом темноты длительностью 16 ч. На основании этого наблюдения, которое, по нашему мнению, не имеет никакого отношения к актиноритмизму, в литературе появились представления о короткодневных видах-одноночниках.
Чтобы показать несостоятельность таких высказываний об актиноритмической никтофильной реакции, мы провели специальные опыты с ипомеей голубой в условиях искусственного освещения лампами ДРЛ при интенсивности на уровне вершин растений около 40 В т/м2.
В первом опыте растения от момента появления всходов выращивали в условиях различных длин дня в течение 4 месяцев.
Максимальная скорость репродуктивного развития наблюдалась при 8- и 9-часовом дне. С удлинением светлого периода в суточном цикле переход растений к цветению замедлялся, однако даже при 16-часовом дне образовались единичные бутоны с задержкой почти на три месяца. Таким образом, ипомея проявляет четкую актиноритмическую реакцию короткого дня с критической длиной дня около 16 ч.
Сокращение длины дня до 4 ч задержало репродуктивное развитие растений. Световой день, равный 2 ч, привел к гибели растений. В том случае, когда 2-часовой день получали растения, предварительно до 2-месячного возраста выращенные при непрерывном освещении, цветочные зачатки образовались на 97-е сутки, но дальнейшего развития цветков не происходило.
На урожай надземной массы также в сильной степени влияла длина дня, при которой росли растения. При актиноритмических режимах, наиболее благоприятных для репродуктивного развития, рост побегов вскоре прекращался в связи с закладкой цветочных органов на верхушках растений. Наибольший урожай надземной массы был получен при 15-часовом дне, дальнейшее удлинение светового периода не стимулировало рост надземной массы, а при длине дня 20 ч и в условиях непрерывного освещения наблюдалось некоторое снижение массы растений.
В следующем опыте растения, выращенные до фазы полного раскрывания семядолей при непрерывном освещении, подвергались воздействию единственным темным периодом длительностью от 8 до 20 ч. Зачатки цветков образовались лишь при длительности темного периода более 15 ч, но в дальнейшем эти зачатки не развивались и опадали, верхушечные конусы нарастания вновь образовали вегетативные органы — листья.
Опыты показали, что образование развитых цветочных органов и затем плодоношение растений ипомеи невозможны при воздействии одним периодом темноты, для этого необходимо чередование периодов света и темноты в течение определенного времени.
Чтобы выяснить минимальное число короткодневных циклов, необходимое для образования цветков и плодов ипомеи, были поставлены специальные опыты. В одном случае для опыта брали молодые растения в фазе развернутых семядольных листьев, в другом — взрослые, двухмесячные растения.
При индукции в фазе семядольных листьев растениям потребовалось для образования цветочных зачатков 3 коротких дня, а для формирования цветков и плодов — 19 коротких дней. В возрасте двух месяцев для закладки зачатков цветка было достаточно одного периода темноты, но для образования цветка потребовалось 11 коротких дней.
По-видимому, процесс физиологической перестройки листа у ипомеи завершается лишь после определенного числа коротких дней (не менее 11), только в этом случае лист становится устойчивым к действию непрерывного освещения и способным вызывать цветение и плодоношение растений.
В пользу такого предположения свидетельствуют результаты следующего опыта с ипомеей. Растения двухмесячного возраста, получившие 20 десятичасовых дней, были переведены в условия непрерывного освещения, где у них систематически по мере появления удаляли бутоны и вновь образующиеся листья.
Опытом установлено, что растения ипомеи сохраняют способность воспроизводить новые репродуктивные органы в течение 150 дней при непрерывном освещении, несмотря на старение и опадение значительной части индуцированных листьев.
Результаты проведенных исследований показывают, что цветение и плодоношение ипомеи возможны лишь после того, как она росла при определенном числе коротких дней. Длина индукционного периода сокращается с возрастом растений. Увеличение продолжительности короткодневного воздействия ускоряет образование репродуктивных органов. Скорость образования репродуктивных органов у ипомеи зависит от длины дня в суточном цикле значительно сильнее, чем у других короткодневных растений. При 16-часовом дне первый цветок образовался на 82 дня позднее, чем при 8-часовом дне.
Растения ипомеи, получившие достаточное число коротких дней, способны затем образовывать репродуктивные органы при непрерывном свете в течение, очень длительного времени — не менее пяти месяцев.
Таким образом, ипомея голубая переходит к цветению и плодоношению только после завершения 11-дневного индукционного периода в условиях актиноритмов с длиной дня от 6 до 10 ч, обнаруживая тем самым типичную никтофильную реакцию. Если бы ипомея была действительно одноночником, т. е. переходила бы к нормальному цветению после единственного периода темноты, то эта ее реакция на темноту была бы просто темновой, а не актиноритмической.
Помимо ипомей, на один период темноты, по данным Хамнера и Боннера (Hamner, Bonner, 1938), реагирует переходом к цветению и пенсильванский дурнишник, но и это положение оказалось недостоверным.
Прежде всего, в наших опытах выяснялось число индукционных циклов, обеспечивающих нормальное цветение пенсильванского дурнишника. Для этого ставили опыты с разновозрастными растениями. Им создавали от 1 до 15 индукционных циклов с длиною светлого периода 10 или 14 ч (темнота соответственно 14 и 10 ч).
Все опыты с дурнишником проводили в условиях искусственного освещения, источником которого в основном служили лампы ДРЛ по 250 и 500 Вт; в качестве арматуры использовали специальные глубокоизлучатели.
Как известно, дурнишник обладает раздельнополыми цветками. Это имеет большое значение для правильного истолкования результатов, полученных в актиноритмических исследованиях.
В наших опытах обнаружилось, что после одного или двух индукционных циклов появляются только мужские цветки, причем сколько бы эти растения ни держали после индукции в условиях непрерывного освещения, они так и не формировали женских цветков. Чтобы получить женские цветки, необходимо самое малое 5—6 индукционных циклов. Во всяком случае при четырех индукционных циклах женские цветки не появлялись. Следовательно, для формирования как женских цветков, так одновременно и мужских и женских, дурнишнику необходим не один, а по крайней мере пять индукционных циклов. Это сближает дурнишник по актиноритмической реакции цветения с другими никтофильными видами, в частности с периллой масличной. У дурнишника, как и у других видов, имеющих раздельнополые цветки, раньше и в более широком диапазоне внешних условий образуются мужские цветки. Непонятно, почему большинство физиологов растений, занимающихся актиноритмизмом дурнишника, обращали внимание на образование не женских, а мужских цветков. Это, несомненно, ошибка, приведшая к некоторым неясностям в оценке актиноритмической реакции пенсильванского дурнишника.
Несостоятельным оказалось и второе положение о том, что цветки у дурнишника могут образовываться только после восьмого листа. Напротив, нам без труда удалось получить и мужские, и женские цветки в пазухе первого листа через 10—11 дней после появления всходов, особенно если растения были декапитированы. Если указанное растение чем-то и отличается от таких видов, как перилла масличная, так это только большей актиноритмической чувствительностью уже первого малоразвитого листа. Несомненно, она у него выше, чем у периллы. Очень важен и тот факт, что пенсильванский дурнишник может образовывать нормальные цветки после появления листьев первого узла, что свидетельствует о их половой зрелости уже через 5—7 сут после всходов. Все сказанное и дает нам право отнести дурнишник к растениям, очень рано вступающим в период половой зрелости.
Переход к цветению у периллы и ипомеи регулируется никтофильными актиноритмическими реакциями, а у шпината и капусты абиссинской — никтофобными. Характерная особенность репродуктивного развития растений — это задержка его определенными периодами суточной темноты у растений, относимых к группе никтофобных, и, наоборот, необходимость суточной темноты для цветения другой — никтофильной группы. Однако есть и такие растения, у которых процессы репродукции идут с одинаковой активностью как при непрерывном освещении, так и при коротких днях, равных 4—5 ч. К таким растениям относятся исследованные в наших опытах гречиха и хлопчатник.
Опыты с хлопчатником (сорт Одесский 7) начались после того, как было выяснено, что при росте в условиях с электрическим освещением он оказывается значительно скороспелее и продуктивнее, чем в естественных условиях. Так, в обычной осветительной установке лаборатории светофизиологии, состоящей из ламп накаливания и водяного фильтра, хлопчатник дал три зрелые коробочки за 80—85 дней после появления всходов.
В первом опыте необходимо было установить влияние мощности лучистого потока при прочих равных условиях на синтетическую деятельность растений хлопчатника (для этого были использованы мощные лампы в режиме их перенакала). Спектральный состав лучистого потока и продолжительность ежесуточного освещения в опыте были одинаковыми, освещение непрерывное.
В опыте испытывали следующие мощности, Вт/м2: 100, 200, 400, 600, 800 и 1000, последняя равна максимальному количеству солнечной лучистой радиации. Под действием радиации указанной мощности растения хлопчатника выращивали в течение 15 дней, начиная с появления всходов. В конце данного периода все растения высушивали до абсолютно сухого состояния. Сухая масса растений характеризовала реакцию хлопчатника на соответствующие световые условия.
Накопление сухой растительной массы хлопчатником следующим образом зависит от мощности лучистого потока.
Хлопчатник считается одной из самых светолюбивых культур. Однако, как видно из приведенных данных, накопление растительной массы в данном случае шло лучше всего при мощности лучистого потока 400 Вт/м2, т. е. примерно при половинной мощности прямого солнечного излучения. Во всяком случае совершенно ясно, что хлопчатник можно выращивать при мощности лучистого потока 400 Вт/м2 и что дальнейшее увеличение ее может быть не только бесполезным, но даже вредным. Правда, удвоение этой оптимальной мощности мало снижает продуктивность хлопчатника и тем самым указывает на его приспособленность переносить и высокие мощности солнечного излучения.
Нет ничего удивительного, что 0,1 прямого солнечного излучения не может обеспечить той же высокой продуктивности, какая получается при интенсивности освещения в 5 раз большей. Однако при мощности лучистого потока в 200 Вт/м2 растения оказываются более продуктивными, чем при освещении 1000 Вт/м2.
После указанных исследований были поставлены опыты, выявляющие реакцию хлопчатника на продолжительность периода ежесуточного освещения. Для этого использовали те же осветительные установки, что и в предыдущих опытах.
Мощность лучистого потока для всех вариантов актиноритмического опыта установлена в 500 Вт/м2. Продолжительность периода ежесуточного освещения равнялась 12, 15, 18 и 21 ч. Кроме того, был и пятый вариант, где растения освещались непрерывно в течение всего опыта, длившегося до появления бутонов у всех растений. После бутонизации отстающих по развитию растений все они одновременно, в возрасте 27 сут, были убраны для определения их массы в абсолютно сухом состоянии.
Прежде всего вслед за удлинением ежесуточного освещения при мощности лучистого потока 500 Вт/м2 увеличивается накопление сухой растительной массы, достигающей наибольшей величины при непрерывном освещении.
Образование значительной растительной массы растениями при непрерывном освещении — явление не редкое, но оно часто сопровождается задержкой в развитии. В описанном опыте с хлопчатником этого не произошло и раньше всего бутоны появились при непрерывном освещении. Растения, росшие при 12-часовом дне, образовали первые бутоны на 6 сут позже.
По результатам данного опыта нельзя считать исследуемый сорт хлопчатника (Одесский 7) растением короткого дня. Его актиноритмическая реакция по цветению, по общепринятой терминологии, оказалась нейтральной. Однако сказать о нем, что он вообще нейтрален к актиноритмам, тоже нельзя, ведь его растительная масса при непрерывном освещении была в 2 раза больше, чем при 12-часовом дне. Точно такое же явление наблюдается и у всех других так называемых нейтральных растений.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Подземная ферма в Лондоне – выращивание растений в темноте
В большинстве случаев письма приходят в течение одной минуты, но иногда для этого требуется до 10 минут. Возможно письмо еще не успело прийти. Проверьте пожалуйста внимательно папку Входящие (Inbox). В некоторых случаях письмо может попасть в папку Спам (Spam).
Логин или e-mail: Или войдите с помощью этих сервисов:www.ogorod.ru
GREENERGY » Реакция растений на длительную темноту
При выращивании растений исключительно в условиях светокультуры в фитотронах, камерах зимой в Заполярье и в других случаях иногда нарушается подача электроэнергии. Растения могут оказаться в темноте на длительное время, значительно превышающее фотопериодический цикл, выработанный в процессе эволюции.
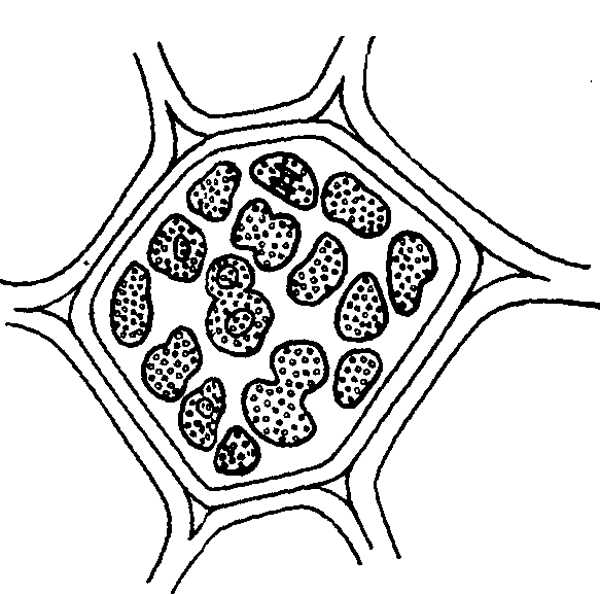
Длительное (порядка нескольких суток) отсутствие оптического излучения вызывает нарушение физиологических процессов, изменение структуры отдельных органелл, внешние повреждения органов и, наконец, гибель растений. По данным Т. Н. Шманаевой, у томатов уже на 2-е сутки уменьшается количество нуклеиновых кислот, размеры ядер, увеличивается плазмоядерное отношение, прекращаются митозы, задерживаются ростовые и формообразовательные процессы. Более длительная темнота вызывает фрагментацию ядер и полное их разрушение в клетках апикальной меристемы точек роста. На 7-е сутки растения погибают. При этом наблюдается большая потеря воды всем растением, некроз листьев, побурение и ослизнение корней. Нарушается обмен веществ, что проявляется в избыточном накоплении амидного и аммиачного азота. В листьях происходит одновременный и быстрый распад пигментов и частичный их ресинтез (хлорофилла – на 4-е сутки темноты, каротиноидов – на 6-е). Общее уменьшение количества пигментов понижает степень поглощения листом лучистой энергии за счет увеличения пропускания и отражения. Ход спектральной кривой при этом не смещается. Хлоропласты собираются к концам клетки или вокруг ядра. Задерживается их движение, изменяется форма – они становятся округлыми или угловатыми. Наблюдаются изменения и в структуре хлоропластов: у огурцов уже через 2 суток отмечена их деструкция. Уменьшается число гран, толщина мембран тилакоидов, увеличивается расстояние между ними. Постепенно разрушается строма хлоропластов. Через 5 суток образуются проламеллярные тела, затем разрушается оболочка, хлоропласт перестает существовать как автономное образование и прекращает функционировать. Разрушение структуры хлоропластов зависит от продолжительности темноты: сначала набухают тилакоиды гран и стромы, а затем появляются проламеллярные тела, которые со временем теряют решетчатую структуру; разрушается оболочка хлоропластов, остатки гран, и отдельные тилакоиды перемещаются непосредственно в цитоплазму.
Устойчивость растений к повреждающему действию длительной темноты определяется разными факторами: наследственными свойствами данной культуры, возрастом растений, этапом органогенеза, наличием запасных веществ, фотопериодической реакцией и т. п. Особое значение имеют параметры внешней среды: температура воздуха, условия минерального питания, режим предтемнового облучения и др. При температуре воздуха порядка 30° С огурцы погибают на 2-е, а томаты на 4-е сутки. Пониженная температура повышает устойчивость растений: томатов при 12° С – до 7 дней, пшеницы при 3° С -до 15 дней. Если после этого срока создать нормальные условия облучения, то растения возобновляют рост. При выращивании растений на питательной смеси исключение из нее азота повышает, а отсутствие фосфора понижает устойчивость растений. Интенсивность облучения в предтемновой период играет роль регуляторного механизма, определяющего направление и степень функциональных и структурных процессов, протекающих в растениях во время длительной темноты. В определенных условиях темнота играет важную защитную роль. Повреждающее действие на растения кислот, щелочей, гербицидов, рентгеновского излучения и низкой положительной температуры значительно слабее в темноте, чем на свету.
greenergy.org.ua
Сельское хозяйство будущего: как выращивать овощи без Солнца
В 21 веке пищевая индустрия претерпит существенный сдвиг: выращивание еды в немалой степени переместится в помещения. Идеальный урожай будет расти внутри здания без окон, с контролируемым светом, температурой, влажностью, качеством воздуха и питания. Такая ферма 21 века сможет находиться в подвале токийского небоскрёба, в старом складе в Иллинойсе, или даже в космосе. Взгляните на эту коллекцию замечательных внутренних ферм, где солнце никогда не светит, дождь никогда не идёт, а климат всегда идеален.
Базилик, руккола и микрозелень

Рабочий проверяет урожай на вертикальной ферме «FarmedHere» в Бедфорд Парк, Иллинойс, 20 февраля 2013 года. Эта ферма расположена внутри старого склада и продаёт свой урожай в зеленных лавках Чикаго и его пригородов.
Ваш салат растёт в полной темноте

Красный салат-эндивий, растущий на внутренней ферме «California Vegetable Specialties» в Рио Виста, Калифорния, апрель 2006. Процесс его роста долог и сложен, поскольку корни эндивия сначала растут наружу, а затем уходят внутрь, где в течение 11 месяцев созревают в полной темноте.
Под флуоресцентным светом

Тоширо Сакума проверяет состояние растений, растущих под флуоресцентными лампами в теплице, построенной в подвале токийского здания, 1 июля 2005 года.
Урожай без солнца

Растения фиттонии растут в специальной затемнённой комнате, подсвечиваемой синими и красными светодиодами в частной исследовательской лаборатории «PlantLab» в Ден Бош, центральная Голландия, 28 марта 2011.
Посадки медицинской марихуаны

Эта ферма находится в Окленде, Калифорния. Её счёт за электричество составляет больше 4 тысяч долларов в месяц.
Нелегальные посадки марихуаны

Эта внутренняя ферма конопли была арестована полицией Калифорнии.
Легальные посадки марихуаны

В Голландии выращивание конопли легально в течение уже долгого времени. Вот как выглядят посадки лучшей в мире марихуаны, сфотографированные несколько лет назад.
Японские внутренние теплицы

Цветы, которые выращивают под флуоресцентными лампами в теплице, названной «Pasona O2», в подвале офисного небоскрёба в Токио. В этой теплице применяются самые высокотехнологичные методы выращивания растений, включая гидропонику и светодиодную подсветку.
А вот так выглядят рисовые поля в «Pasona O2». Компания Pasona надеется, что её теплицы вдохновят новые поколения фермеров.

Гидропонная томатная ферма в «Pasona O2»

Персонал «Pasona O2» проверяет овощи, которые выращиваются при флуоресцентном освещении

Гидропонные сады для каждого

«Volksgarden приносит простые, чистые и удивительно эффективные гидропонные системы прямо в ваш дом», сообщает компания «Urban Led Growth». Такой юнит позволяет выращивать до 80 растений одновременно. В нём можно с лёгкостью выращивать травы, овощи, фрукты, и зерновые непрерывно, благодаря вращающейся цилиндрической основе.
Разместите AeroGarden у себя на кухне

Этот безгрунтовый внутренний садовый юнит использует аэропонику: овощи, салатная зелень, травы и цветы растут в этом горшке, будучи лишь немного выставленными на воздух, и немного погружёнными в питательный раствор. AeroGarden имеет встроенный источник света и систему «Smart Garden», которая подскажет вам, когда вашим растениям нужно больше питательных веществ или воды.
Автоматизированный, гидропонный, рециркуляционный вертикальный фермерский комплекс

Это один из четырёх подобных комплексов в «Green Farms Agronomics & Mycology». Эта компания расположена в Вальпараисо, Индиана, и была основана осенью 2010 года.
Чикагский городской сад

Это первый сад программы «Аэропонный сад в любой аэропорт мира». В 2011 году компании CDA и HMS Host Corporation объединились, чтобы построить этот сад в мезонине «O’Hare Rotunda Building». В этом саду корни растений помещены в 26 башен, содержащих более 1100 мест под посадку. Через башню с помощью насосов регулярно прокачивается питательный раствор, так что система не тратит и не испаряет лишнюю воду, что делает процесс само-поддерживающимся. Также в ней не используются удобрения или любые другие химикаты.
Астрокультура

Это внутренность блока для выращивания растений «Astroculture» в ходе миссии космического шаттла STS-73 в 1995 году. Астрокультурный комплекс совершил с тех пор восемь полётов на космическом шаттле, включая этот полёт 1995 года, когда в нём был выращен картофель, который вы можете видеть на фотографии.
Соевые бобы, выращенные на борту Международной космической станции

Член Пятой космической экспедиции, инженер Пегги Уитсон демонстрирует прогресс роста соевых бобов, выращенных в ходе эксперимента Advanced Astroculture (ADVASC) на борту МКС в 2002 году. Эти эксперименты призваны определить возможности роста растений в микрогравитационной среде.
ПОХОЖИЕ ЗАПИСИ
© Gearmix 2013 Права на опубликованный перевод принадлежат владельцам вебсайта gearmix.ru Все графические изображения, использованные при оформлении статьи принадлежат их владельцам. Знак охраны авторского права распространяется только на текст статьи. Использование материалов сайта без активной индексируемой ссылки на источник запрещено.
gearmix.ru
Комнатные растения повышают производительность у офисных работников

Наличие зеленых растений на рабочем месте обогащает не только пространство, в котором мы работаем, но и улучшаем нашу производительность. Но почему?
Группа исследователей из школы психологии Кардиффского университета (Великобритания) провела натурное исследование, чтобы выяснить влияние на производительность людей в офисе наличие («зеленый офис») и отсутствие растений. После проверки восприятия персоналом качества воздуха, удовлетворенности работой, уровня концентрации и производительности в различных офисах ученые обнаружили, что офисы с зелеными растениями делали рабочих более счастливыми в своей рабочей среде. Они чувствовали себя более сосредоточенными и были на 15% эффективнее, чем в других офисах, где не было растений.
«Наше исследование показывает, что инвестиции в озеленение офисов с использованием зеленых растений будут окупаться за счет повышения качества жизни и производительности офисных работников», сказал руководитель исследования Марлон Нивенхюис, которое было опубликовано в «Журнале экологической психологии». (Journal of Environmental Psychology)
Причина этого огромного преимущества, согласно исследованию, заключается в том, что присутствие зеленых растений повышает трудовую активность сотрудников, делая их более физически, умственно и эмоционально вовлеченными в свою работу. Это происходит потому, что растения могут снизить нагрузку, повысить внимательность и улучшить общее самочувствие, что в конечном итоге приводит к повышению производительности.
Если вы захотели прислушаться к результатам этого исследования, а также преобразить растениями свой дом или офис, то качественные семена вам стоит поискать вот тут: https://leto.ua/. И тогда у вас и ваших работников все наладится.
Но растения кроме приносимой пользы способны и на другие интересные фокусы.
Растения сопротивляются лучше, чем животныеПять великих вымираний привели к глубоким изменениям в истории жизни на Земле. Однако новое исследование, проведенное группой ученых из Гётеборгского университета в Швеции, похоже, показывает, что растения лучше справляются с этими кризисами, чем животные.
Изучив более 20000 окаменевших растений, эти эксперты обнаружили, что вымирания затрагивали совершенно разные группы растений.
В частности, в исследовании, опубликованном в журнале «New Phytologist», они указывают, что после одного из этих событий с такой же скоростью исчезновения и появляются новые виды.
По сути, это указывает на то, что во время борьбы с последствиями вымирания, растения ведут себя более устойчиво. Например, падение астероида 66 миллионов лет назад ускорило исчезновение динозавров и многих других животных, но оказало ограниченное влияние на биоразнообразие растений.
«Среди растений массовое исчезновение действительно можно рассматривать как возможность для обновления», говорит Даниэле Сильвестро, эксперт в области филогенетики и экологии, который координировал исследование.
Но стоит также упомянуть, что сегодня есть стремления генетически модифицировать растения, чтобы сделать их светящимися в темноте и т.д., но подобные ГМО-растения плохо приживаются и не оправдывают тех надежд, о которых сообщают их создатели. Это касалось и светящихся в темноте растений.
gmoobzor.com
Комнатные растения растущие в темноте
данная презентация позволяет обучающимся познакомиться с комнатными растениями и правилами ухода за ними.
Просмотр содержимого документа «Комнатные растения растущие в темноте»

Тема: КОМНАТНЫЕ РАСТЕНИЯ
Автор
Курта Оксана Викторовна
учитель биологии
МБОУ СОШ № 189.

Комнатные растения, растущие в темноте.
Наверняка у всех есть темное место в квартире, которое так и хочется оживить, и сделать уютным, может это ванная, коридор, или гостиная, окна которой выходят на теневую сторону. Выход из ситуации есть, ведь не все растения любят солнечный свет, для некоторых их них он даже губителен. Есть золотое правило выбора тенелюбивых растений: чем темнее листья у растения — тем больше оно любит темный угол. Но нужно также понимать, что все растения нуждаются хотя бы в небольшом количестве рассеянного света, в условиях недостаточной освещенности растение не зацветет, а так как красота цветущих растений в цветках, мы сосредоточили свое внимание на красивых декоративных растениях, красота которых, в их листьях и окрасах.

Сансевиерия или «Тещин язык»
- В последнее время очень модная, украшает многие фешн рестораны и кафе, в которых зачастую нет окон, а используется искусственное освещение. Она может расти на солнце и в тени, она переносит сквозняки и перепады температур, редкие пересадки, растение очень неприхотливое, единственное чего боится сансеверия это переувлажнения и полива внутрь розетки, которая от этого может загнить.

Драцена Сандера
- Это растение также имеет называние «Счастливый бамбук» . Растение является одним из простых в выращивании, она имеет особенность расти в условиях низкой освещенности. Счастливый бамбук может расти в условиях многочасового затемнения, но небольшое количество света ему необходимо. Также популярность свою обрела из-за возможности формирования разных декоративных форм из стебля, чаще всего это спираль. Если планируете переставить драцену в более темное, или более светлое место, делайте это постепенно, понемногу сдвигая каждый день на несколько часов в новое место. В темном месте листья могут стать светло-зелеными.

Аспидистра
- Аспидистра проста на вид, но когда растение вырастает, то становится элегантным украшением интерьера. В последнее время мода на сочетание плетеной мебели и этого растения набирает оборотов. Растение неприхотливое, переносит небрежное обращение в прямом смысле слова, растение практически не нуждается в удобрении. Аспидистра не реагирует на избыток, или недостаток влаги, это растение для тех, кому некогда ими заниматься. Аспидистра незаменима при декорировании помещений с небольшой освещенностью или изменчивым температурным режимом .

Хлорофитум или «Растение паук»
- Неприхотливое растение, которое в добавок ко всему еще и цветет белыми цветами. Летом требует обильного полива, 2 раза в неделю, и опрыскивание. Зимой достаточно 1 раза. Растение отлично фильтрует воздух, он поглощает формальдегид и окись углерода и выделяет кислород. Растение лучше всего растет в полутени, но если его убрать в совсем темную комнату, например, офис, он утратит белые полоски на листьях.

Плющ или хедера
- Вечнозеленое растение, которое в природе выбирает себе место для роста в тенистых лесах. Благодаря большой массе листьев, хедера считается одним из самых эффективных очистителей воздуха в доме. Для того, чтобы ваш плющ превратился в шикарную зеленую стену, необходимо создать ему прохладу, плющ не выносит жары и солнца, он тенелюбив. Хедера влаголюбивое растение, корневая система у нее поверхностная, поэтому в весенне-летний период полив должен быть регулярным и достаточно обильным. Пересыхание земляного кома нежелательно, но и застой воды в горшке приведет к гибели растения. Зимой поливать часто при зимовке в теплом помещении, но если плющ зимует в прохладном месте, полив следует ограничить так, чтобы только не допускать пересыхания почвы.

Спатифиллюм
- Это экзотическое растение в народе называют «белый парус». Растение при благоприятных условиях цветет очень длительное время, цветы напоминают белый парус, некоторые виды имеют насыщенный аромат. Растение боится прямых солнечных лучей, может расти при рассеянном свете или в полутени. Полив в весенне-летний период и во время цветения обильный, между поливами верхний слой должен подсохнуть. В осенне-зимнее время полив умеренный. При поливе субстрат не должен пересыхать, но и не должен быть чрезмерно переувлажнен. Для спатифиллума важно правильно выбрать место – он не любит холода. Поставьте его подальше от сквозняков и открывающихся зимой для проветривания окон, и никаких проблем с растением не будет.

Папоротник
- Папоротник любит тень и влажный воздух. Самые простые в содержании и распостраненные папоротники: цитромиум, даваллия, птерис критский, нефролепис, костенец гнездовой, пеллея круглолистная. Но даже они нуждаются в частом опрыскивании листьев, особенно во время отопительного сезона. Следите, чтобы земля в горшке с папоротником всегда была влажной.

Сциндапсус (Потос)
- Растение приспособлено к условиям комнатной культуры, влаголюбивое и теневыносливое, но при недостаточном освещении пестрота листвы может появляться или исчезать. В комнатной культуре растения практически не цветут. Применяют как ампельное растение и как вьющееся на опоре, обвернутой сфагновым мхом. Используется в разнообразных аранжировках.

Гардения
- Менее капризная из этих прекрасных цветков является гардения жасминовая. При выборе места для гардении выбирайте южные или юго-западные окна. Растение любит частые опрыскивания. Может расти в притененных местах, с пониженным освещением в течении всего дня, но выбор места определите заранее и не изменяйте его, даже самое небольшое перемещение приводит к опаданию цветов и даже листьев. Ни в коем случае нельзя допускать пересыхания земляного кома. Грунт всегда должен быть слегка влажный. Но при этом он должен хорошо пропускать воздух к корням. Поэтому и рекомендуют для выращивания гардении жасминовидной использовать лёгкую почву, как для Азалий.

Аглаонема
- Эти яркие растения с зелеными листьями прекрасно растут в местах, с недостаточным освещением. Прямые солнечные лучи могут привести к ожогам листьев. Растение боится сквозняков. Несмотря на способность аглаонемы очищать воздух от бензола и убивать стрептококковую инфекцию, существует мнение, что из-за содержания в ней щавелевой кислоты, токсичных энзимов и протеинов, при неправильном обращении с растением может вызвать сильные раздражения, и даже ожоги кожных покровов. Выделение сока растения может вызвать отеки слизистых оболочек, затруднение дыхания. В некоторых случаях аглаонема может вызывать аллергические реакции, именно поэтому аглаонема и закрывает список растений, любящих тень. Поэтому при проведении каких-либо работ с растением нужно пользоваться защитными перчатками, а по окончании нужно хорошо промыть руки с мылом. Лучше убрать растение от животных и детей.

Спасибо за внимание!
Литература
http://yandex.ru/clck/jsredir?from=yandex.ru%3Bimages%2Fsearch%3Bimages%3B%3B&text=&etext=1327.5k6JOm0Z82T3auDA_DN5ny8MsrR0ciJ0Lsmycxpnyy5ZorSN8Z4oxlEHptOAJ3M3xaNPujZG0l6xYZbCkMQoLA.1c7d3c56eebade5dc98864f5f5e27762e9e96be2&uuid=&state=tid_Wvm4RM28ca_MiO4Ne9osTPtpHS9wicjEF5X7fRziVPIHCd9FyQ,,&data=UlNrNmk5WktYejVVaS1zZThueDVKOXVnTXpDa1ZockZfZVR6aHdLTUVSREpwN19UVlljRkdsSm8xbnZSUG1ZMURZT1lmTUxyNUZ3eFdod21QcGtFN3ZvWlhRbVdVNEU0WmJGVjFmRWhBN210dm9JeTlLNjFPN3ZnMlFDN19JSDlYQ2xBOU1nMFQ0OCw,&sign=9b5676cccf2d049456374daca0a138f5&keyno=0&b64e=2&l10n=ru
1.
https://kashevarnya.com/recept/komnatnye-rasteniya-rastushchie-v-temnotetop-10-neprihotlivyh-rasteniy
http://yandex.ru/clck/jsredir?from=yandex.ru%3Bimages%2Fsearch%3Bimages%3B%3B&text=&etext=1327.t7crNRUu6dcvJ1p7dmRetmDtd4atQqEisGzIzTDdrwmAPerxibDXtnRknXMJZcDF4fHb7leHbcLOBxkB0vZ6a3qYBNNLdrIOLc0pZhkJdANsJsUgWpJlCAAl_1AuB7-LF-JMG_KX1xVMWd6AZ5Y7rA.3f9747cd0ae1ea916d6666341573776ba3a437b2&uuid=&state=tid_Wvm4RM28ca_MiO4Ne9osTPtpHS9wicjEF5X7fRziVPIHCd9FyQ,,&data=UlNrNmk5WktYejdJbjBTS0RYZ2lucHZNMDdheldQUEpnenhEb0Y0Y2x3cmRrUUtsZ3Q4WFpfOUZrVUd1clNQTTg0bXVKemN6WEs4YmNwSlVyTWd0SllmR3BXZ21UUS1LaENyT2FwLW8tbWVRTDJWMjJfOWtLcEpIYWRIQXllaTliLVBSYkhzMXFhQ252RmRkOUtNZE1MY1o3SXUySC1tOVBhOTBGQlIyZWY4WkFrSThSMDhoRUljSkcyZV8waTJWam54RWlPX3MtcjJua2VzRTB4dnBZZyws&sign=8d5ffef4cc2d947b3f7f8d0a7f0b6456&keyno=0&b64e=2&l10n=ru
http://yandex.ru/clck/jsredir?from=yandex.ru%3Bimages%2Fsearch%3Bimages%3B%3B&text=&etext=1327.t7crNRUu6dcvJ1p7dmRetmDtd4atQqEisGzIzTDdrwmAPerxibDXtnRknXMJZcDF4fHb7leHbcLOBxkB0vZ6a3qYBNNLdrIOLc0pZhkJdANsJsUgWpJlCAAl_1AuB7-LF-JMG_KX1xVMWd6AZ5Y7rA.3f9747cd0ae1ea916d6666341573776ba3a437b2&uuid=&state=tid_Wvm4RM28ca_MiO4Ne9osTPtpHS9wicjEF5X7fRziVPIHCd9FyQ,,&data=UlNrNmk5WktYejY1MXJJdGM1QmRPZHh3dzg4azBGQktCWV9jcm5yMzlVUHBfS3V0aHhiTVg1QnBTOTJ2T1U3WlQxWVBrNlh4UzF1VldmbzViNXRZWEg3dVQ4cWJhWVJvckNtMW13YnowRUt6TmMtSW5tUnI1WnQ2bDQ0UFFQU1VHT0hBcF9kbXIxVG55ckduZHBraTdYblpUYkRoSGNvNA,,&sign=e821de35005f5dcee9a6b25dc89ff8f4&keyno=0&b64e=2&l10n=ru
kopilkaurokov.ru









